Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
SACHARIDOVÝ METABOLISMUS KOSTERNÍCH SVALŮ BĚHEM TĚLESNÉ PRÁCE
Fyziologie a fyziologie zátěže Fakulta tělesné kultury UP

2
Dostupnost a využitelnost sacharidů během prolongované (dlouhotrvající) zátěže.
Svalová únava - někdy spojená s nedostatkem sacharidů - deplece (snížení) glykogenu (GG) nebo hypoglykémie. Nedostatek sacharidů - redukce hladiny pyruvátu. Lipidy nelze využívat jako energetický substrát při intenzitě zatížení (IZ) > 60% VO2 max. Laktát - preferovaný substrát pro glukoneogenezi v játrech a pro oxidaci ve svalech a v myokardu.
 glykogenu (GG) nebo hypoglykémie. Nedostatek sacharidů - redukce hladiny pyruvátu. Lipidy nelze využívat jako energetický substrát při intenzitě zatížení (IZ) > 60% VO2 max. Laktát - preferovaný substrát pro glukoneogenezi v játrech a pro oxidaci ve svalech a v myokardu.")
3
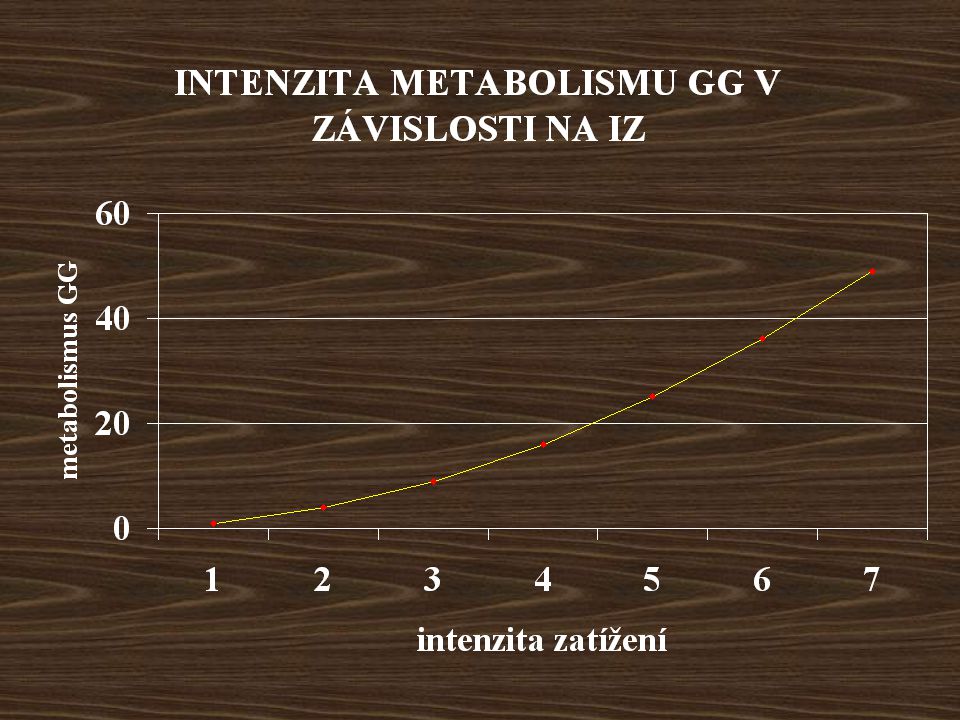
Metabolismus svalového GG během tělesné práce.
Nejintenzivnější na začátku zatížení. Intenzita metabolismu GG roste exponenciálně s IZ.

5
lipolýza aerobní glykogenolýza anaerobní glykogenolýza Převažující metabolismus

6
Zapojování jednotlivých energetických systémů
v průběhu maximální práce. Závažný omyl staré koncepce energetického metabolismu: Energetické systémy odpovídají požadavkům intenzity zatížení téměř ve všech sekvencích zátěže Alaktátová zóna Laktátová zóna Aerobní sacharidová zóna Aerobní lipidová zóna t

7
4. Zapojování jednotlivých energetických systémů v průběhu maximální práce.
Jaký je aerobní a anaerobní podíl v jednotlivých fázích maximální práce? Je stejný? (???) Mění se? (???) Gastin (2001) pět příkladů odhadů podílů energetických systémů 1. 90sekundová maximální zátěž na kole 2. běh na 800 m 3. běh na 1500 m 4. vyčerpávající jízda na bicyklu při intenzitě zatížení 110 % VO2 max 5. jednostranné extenze kolenního kloubu při zatížení 65 W
 Mění se ( ) Gastin (2001) pět příkladů odhadů podílů energetických systémů sekundová maximální zátěž na kole. 2. běh na 800 m. 3. běh na 1500 m. 4. vyčerpávající jízda na bicyklu při intenzitě zatížení 110 % VO2 max. 5. jednostranné extenze kolenního kloubu při zatížení 65 W.")
8
Odhad podílu aerobního energetického systému (%) v po sobě následujících fázích během různých druhů pohybové aktivity a při různých intenzitách zatížení Po sobě následující fáze výkonu (s) Maximální 90 s cyklistický výkon 800 m běh 1500 m běh 110 % VO2 max bicykl Jednostranná extenze kolena při 65 W 0-30 30 41 57 35 20 30-60 73 84 66 44 60-90 91 76 87 90-120 88 81 62 89 85 70 20 – 57 % Tak např. v prvních 30 sekundách maximální práce trvající 90 sekund bylo převážné krytí energetického požadavku v oblasti anaerobní glykolysy.
 Maximální 90 s. cyklistický výkon. 800 m běh m běh. 110 % VO2 max. bicykl. Jednostranná extenze. kolena při 65 W – 57 % Tak např. v prvních 30 sekundách maximální práce trvající 90 sekund bylo převážné krytí energetického požadavku v oblasti anaerobní glykolysy.")
9
Odhad podílu aerobního energetického systému (%) v po sobě následujících fázích během různých druhů pohybové aktivity a při různých intenzitách zatížení Po sobě následující fáze výkonu (s) Maximální 90 s cyklistický výkon 800 m běh 1500 m běh 110 % VO2 max bicykl Jednostranná extenze kolena při 65 W 0-30 30 41 57 35 20 30-60 73 84 66 44 60-90 91 76 87 90-120 88 81 62 89 85 70 70 – 91 % (v prvních 30 s 20 – 57 %) Po 60 s maximálního výkonu výrazně dominuje aerobní fosforylace, která v poslední půlminutě zabezpečila přes 90 % energie
 Maximální 90 s. cyklistický výkon. 800 m běh m běh. 110 % VO2 max. bicykl. Jednostranná extenze. kolena při 65 W – 91 % (v prvních 30 s 20 – 57 %) Po 60 s maximálního výkonu výrazně dominuje aerobní fosforylace, která v poslední půlminutě zabezpečila přes 90 % energie.")
10
35-75 s 0-30 30-60 60-90 90-120

11
Při tělesné práci vysoké intenzity trvající déle než 15 sekund
4. Zapojování jednotlivých energetických systémů v průběhu maximální práce. Při tělesné práci vysoké intenzity trvající déle než 15 sekund nemůžeme hovořit o homogenních energetických zónách V jakém pásmu se tedy pohybovalo energetické krytí při maximálním 90 s cyklistickém výkonu? V anaerobním laktátovém pásmu? Ano, ale jen 30 sekund. Ve zbývajícím čase už o anaerobní zóně mluvit absolutně nemůžeme.

12
Při tělesné práci vysoké intenzity trvající déle než 15 sekund
4. Zapojování jednotlivých energetických systémů v průběhu maximální práce. Při tělesné práci vysoké intenzity trvající déle než 15 sekund nemůžeme hovořit o homogenních energetických zónách

13
Podíl jednotlivých energetických systémů
Gastin PB. Energy system interaction and relative contribution during maximal exercise. Sports Med 2001; 31 (10): Podíl jednotlivých energetických systémů na hrazení celkového energetického výdeje při různém trvání maximální zátěže. 100 aerobní fosforylace 80 ATP-CP Podíly anaerobní glykolýzy a systému ATP-CP na resyntéze ATP po 4 minutách maximální práce činí jen asi 20 % 60 glykolysa 40 Příspěvek k celkovému energetickému výdeji (%) 20 50 100 150 200 250 300 Trvání práce (s)
: Podíl jednotlivých energetických systémů. na hrazení celkového energetického výdeje při různém trvání maximální zátěže aerobní fosforylace. 80. ATP-CP. Podíly anaerobní glykolýzy a systému ATP-CP na resyntéze ATP po 4 minutách maximální práce činí jen asi 20 % 60. glykolysa. 40. Příspěvek k celkovému energetickému výdeji (%) Trvání práce (s)")
14
Vliv tréninku na zapojení
jednotlivých energetických systémů při maximální práci. Gastin a Lawson (1996) rychlostně a vytrvalostně trénovaní sportovci podíl jednotlivých energetických systémů na celkovém energetickém výdeji při 90sekundové práci maximální intenzity na bicyklovém ergometru
 rychlostně a vytrvalostně trénovaní sportovci. podíl jednotlivých energetických systémů na celkovém energetickém výdeji. při 90sekundové práci maximální intenzity na bicyklovém ergometru.")
15
Relativní podíl tří energetických systémů na hrazení celkového
180 Relativní podíl tří energetických systémů na hrazení celkového energetického výdeje během 90sekundové vyčerpávající zátěže 160 140 Energetický výdej Vytrvalostně trénovaní (triatlonisti VO2 max 65 ml/kg/min) 120 Rychlostně trénovaní (cyklisti VO2 max 58 ml/kg/min) 100 Energetický přínos (ml O2 ekviv./kg/min) 80 Glykolysa 60 40 Aerobní fosforylace 20 ATP-CP čas (s) 10 20 30 40 50 60 70 80 90
 120. Rychlostně trénovaní (cyklisti VO2 max 58 ml/kg/min) 100. Energetický přínos (ml O2 ekviv./kg/min) 80. Glykolysa Aerobní fosforylace. 20. ATP-CP. čas (s)")
16
5. Vliv tréninku na zapojení
jednotlivých energetických systémů při maximální práci. Rychlostně trénovaní sportovci - větší silový výkon podporovaný vyšší maximální rychlostí resyntézy ATP ze systému ATP-PC a anaerobní glykolysy I když hodnoty celkové VO2 během testu byly ve skupině sprintérů nižší než u vytrvalců, kinetika VO2 byla u sprintérů překvapivě rychlejší

17
Při prolongovaném zatížení využití GG klesá
pokles dostupnosti (pokles aktivity GP-lázy) vzestup dostupnosti alternativních zdrojů (GL a FFA). Při IZ 60% - 75% VO2 max probíhá rozklad GG zejména ve svalových vláknech I. typu, méně ve IIa. Při zvyšování IZ až ke 100% VO2 max se postupně do glykogenolýzy zapojují všechna vlákna, dominuje zapojení vláken II. typu. Zvýšený rozklad GG i v nepracujících svalech, uvolněný laktát zdrojem glukoneogeneze nebo oxidace.
 vzestup dostupnosti alternativních zdrojů (GL a FFA). Při IZ 60% - 75% VO2 max probíhá rozklad GG zejména ve svalových vláknech I. typu, méně ve IIa. Při zvyšování IZ až ke 100% VO2 max se postupně do glykogenolýzy zapojují všechna vlákna, dominuje zapojení vláken II. typu. Zvýšený rozklad GG i v nepracujících svalech, uvolněný laktát zdrojem glukoneogeneze nebo oxidace.")
18
Regulace glykogenolýzy v kosterních svalech.
PFK - senzitivní na metabolické meziprodukty (ATP, ADP, AMP, F-6-P a F-1,6-PP, NH2, citrát, H+). Při práci o vysoké IZ klesá množství ATP (v důsledku deplece GG), tím klesá poměr ATP:ADP, tím se zvyšuje aktivita PFK a udržuje se intenzita GG-lýzy. Např. při maximálním zatížení redukce F-6-P (důsledek zhoršené dostupnosti svalového GG - deplece - vyčerpání) je kompenzována zvýšenou aktivitou PFK - udržení stejné rychlosti GG-lýzy. Totéž při dlouhodobé práci o střední IZ (75% VO2 max).
. Při práci o vysoké IZ klesá množství ATP (v důsledku deplece GG), tím klesá poměr ATP:ADP, tím se zvyšuje aktivita PFK a udržuje se intenzita GG-lýzy. Např. při maximálním zatížení redukce F-6-P (důsledek zhoršené dostupnosti svalového GG - deplece - vyčerpání) je kompenzována zvýšenou aktivitou PFK - udržení stejné rychlosti GG-lýzy. Totéž při dlouhodobé práci o střední IZ (75% VO2 max).")
19
Regulace glykogenolýzy v kosterních svalech.
GG-fosforyláza - senzitivní na katecholaminy. Aktivace neaktivní b-formy c-AMP po stimulaci adrenalinem, po svalové kontrakci, v důsledku uvolnění Ca++, v důsledku poklesu dostupnosti substrátů, atd. Inhibice aktivní a-formy GG-fosforylázy, aktivace GG-syntetázy a defosforylace G-6-P působením ATP, G-6-P a inzulínu.

20
Regulace glykogenolýzy v kosterních svalech.
GG vazbou na GG-fosforylázu zvyšuje její aktivitu (mění inaktivní b-formu na a-formu) - proto při větším obsahu GG ve svalech je zvýšená pracovní kapacita svalu = zvýšená vytrvalostní výkonnost. Vzestup glykémie snižuje rozklad GG. Podání sacharidů během práce nízké a střední intenzity (pod anaerobním prahem) zvyšuje glykémii, zpomaluje svalovou glykogenolýzu, šetří glykogen a zvyšuje pracovní kapacitu svalu. Při zatížení nad anaerobním prahem nemá podání sacharidů během vytrvalostní práce žádný efekt (GL z krve jako zdroj nestačí).
 - proto při větším obsahu GG ve svalech je zvýšená pracovní kapacita svalu = zvýšená vytrvalostní výkonnost. Vzestup glykémie snižuje rozklad GG. Podání sacharidů během práce nízké a střední intenzity (pod anaerobním prahem) zvyšuje glykémii, zpomaluje svalovou glykogenolýzu, šetří glykogen a zvyšuje pracovní kapacitu svalu. Při zatížení nad anaerobním prahem nemá podání sacharidů během vytrvalostní práce žádný efekt (GL z krve jako zdroj nestačí).")
21
Odhad anaerobního prahu
. . % VO2/kg max = 60 + (VO2/kg max : 3,5) Např. při VO2/kg max = 35 ml bude anaerobní práh zhruba na 70% VO2/kg max % VO2/kg max = (60 + VO2/kg max : 3,5) = 60 + (35 : 3,5) = = 70. . . .
 Např. při VO2/kg max = 35 ml bude anaerobní práh zhruba na 70% VO2/kg max. % VO2/kg max = (60 + VO2/kg max : 3,5) = 60 + (35 : 3,5) = =")
22
Lokální faktory Např. kalcium - aktivuje GG-fosforylázu (kalcium může být lokálně i celkově působící prostředek). Např. ADP, AMP, Pi a intermediáty - aktivizují GG-fosforylázu v místě svého vzniku (zejména ve svalových vláknech II. typu).
.")
23
lokálně i celkově působící Ca++

24
Hormonální regulace GG-fosforylázy
Adrenalin zvyšuje GG-lýzu Inzulín snižuje GG-lýzu ve svalech a snižuje lipolýzu v tukové tkáni inzulín adrenalin

25
Hormonální regulace GG-fosforylázy
Opačný metabolický účinek inzulín adrenalin

26
Spotřeba GL ve svalové tkáni během práce
V klidu - svaly využívají asi 15% - 20% z celkového využití GL. Při 60% VO2 max svaly využívají asi 80% - 85% (zvýšená potřeba a zvýšené prokrvení). Utilizace je ovlivněná trváním (s prolongací práce stoupá využití GL) IZ (se zvyšující se IZ stoupá využití GL, ale POZOR - při vysoké IZ se redukuje využití GL - vzestup G-6-P inhibuje hexokinázu)
. Utilizace je ovlivněná. trváním (s prolongací práce stoupá využití GL) IZ (se zvyšující se IZ stoupá využití GL, ale POZOR - při vysoké IZ se redukuje využití GL - vzestup G-6-P inhibuje hexokinázu)")
27
Regulace spotřeby GL ve svalové tkáni
Lokální faktory - GLUT - transportní jednotka glukózy (nejčastější typ GLUT1 a GLUT4)
")
30
Regulace spotřeby GL ve svalové tkáni
GLUT je aktivována inzulínem uvolněním sarkoplazmatického vápníku hypoxií

31
Regulace spotřeby GL ve svalové tkáni
GLUT je inhibována adrenalinem, který aktivuje GG-fosforylázu (zvyšuje glykogenolýzu)
")
32
Regulace spotřeby GL ve svalové tkáni
Přesto, že při práci hladina inzulínu klesá, jeho účinnost stoupá, neboť se zvyšuje senzitivita svalových buněk na inzulín a zvyšuje se prokrvení svalů a tím se zvyšuje množství inzulínu ve svalových vláknech Vliv inzulínu a zátěže na GL ve svalech je synergní, avšak během zátěže není přítomnost inzulínu nezbytná pro zvýšení transportu GL z krve do svalů (uplatňují se další mechanismy).
.")
33
utilizace glukózy stoupá inzulín při práci klesá!
fyzická práce inzulín při práci klesá! inzulín není nezbytný

34
POKLES INZULINÉMIE ZVYŠUJE SENZITIVITU BUNĚK NA INZULÍN
(negativní zpětná vazba)
")
35
ZVÝŠENÍM SENZITIVITY NA INZULÍN SNIŽUJE POTŘEBU JEHO ZVÝŠENÉ
TĚLESNÁ PRÁCE ZVÝŠENÍM SENZITIVITY NA INZULÍN SNIŽUJE POTŘEBU JEHO ZVÝŠENÉ HLADINY U DIABETES MELLITUS Citlivost na inzulín Citlivost na inzulín

36
Dostupnost substrátu Spotřeba GL je závislá na množství svalového GG.
Na začátku práce při dostatku GG je využití GL svalem minimální. Glykémie má při zátěži souhlasnou dynamiku s využitím svalové GL. Podání GL během práce zvyšuje její utilizaci pracujícím svalem.

37
glykémie utilizace GL

38
glykémie utilizace GL

39
glykémie utilizace GL

40
a tím ke snížení využití GL.
Dostupnost substrátu Zvýšená hladina FFA (např. působením lipoproteinové lipázy v krvi nebo hormon-senzitivní lipázy v adipocytech) vede ke zvýšení jejich utilizace k inhibici PFK ke zpomalení glykolýzy k vzestupu GL-6-P k inhibici hexokinázy a tím ke snížení využití GL.
 vede. ke zvýšení jejich utilizace. k inhibici PFK. ke zpomalení glykolýzy. k vzestupu GL-6-P. k inhibici hexokinázy. a tím ke snížení využití GL.")
41
Stoupají FFA v krvi

42
Stoupají FFA v krvi

44
HSL FA + glycerol triglyceridy FFA FFA FFA FFA FFA FFA

46
Klesá utilizace krevní GL

47
a tím ke snížení využití GL. a klesá utilizace krevní GL.
Dostupnost substrátu Zvýšená hladina FFA (např. působením lipoproteinové lipázy v krvi nebo hormon-senzitivní lipázy v adipocytech) vede ke zvýšení jejich utilizace k inhibici PFK ke zpomalení glykolýzy k vzestupu GL-6-P k inhibici hexokinázy a tím ke snížení využití GL. a klesá utilizace krevní GL.
 vede. ke zvýšení jejich utilizace. k inhibici PFK. ke zpomalení glykolýzy. k vzestupu GL-6-P. k inhibici hexokinázy. a tím ke snížení využití GL. a klesá utilizace krevní GL.")
48
Ještě jednou k metabolismu kyseliny mléčné:
kyseliny mléčná není jen slepý produkt anaerobní glykogenolýzy glukoneogenetický prekrusor GG v játrech oxidativní substrát v pracujících svalech oxidativní substrát v myokardu !

49
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Intenzita zatížení (čím vyšší IZ, tím vyšší podíl anaerobní glykogenolýzy). Trvání práce (čím delší práce, tím větší podíl aerobní glykogenolýzy).
. Trvání práce (čím delší práce, tím větší podíl aerobní glykogenolýzy).")
50
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Typ (způsob) zátěže Při přibližně stejné IZ je utilizace GG při běhu nižší než při jízdě na kole při běhu je zapojeno více svalů (stejné zatížení rozděleno na větší množství svalové tkáně) zapojení jiných svalových vláken (díky relativně nižší IZ připadající na jednotku svalové tkáně je při běhu zapojeno více vláken typu I - červená vlákna s větší aktivitou mitochondriálních enzymů) větší metabolický stres m. vastus lateralis při cyklistice
 zátěže. Při přibližně stejné IZ je utilizace GG při běhu nižší než při jízdě na kole. při běhu je zapojeno více svalů (stejné zatížení rozděleno na větší množství svalové tkáně) zapojení jiných svalových vláken (díky relativně nižší IZ připadající na jednotku svalové tkáně je při běhu zapojeno více vláken typu I - červená vlákna s větší aktivitou mitochondriálních enzymů) větší metabolický stres m. vastus lateralis při cyklistice.")
51
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Trénovanost Vytrvalostní trénink zvyšuje anaerobní práh a a tím zvyšuje preference aerobní glykolýzy před anaerobní glykolýzou zvyšuje využití FFA na úkor GG a GL Příklad Převažující energetický substrát (T = tuky, C = cukry) 55 % VO2 max 65% VO2 max 75% VO2 max 85% VO2max A (netrén.) T C C C B (trén.) T T C C
 55 % VO2 max 65% VO2 max 75% VO2 max 85% VO2max. A (netrén.) T C C C. B (trén.) T T C C.")
52
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Dieta vysoký obsah sacharidů zvýšená oxidace sacharidů během práce zvýšená glykogenolýza ve svalech zvýšená spotřeba glukózy nízký obsah sacharidů nízká oxidace sacharidů během práce snížená glykogenolýza ve svalech snížená spotřeba glukózy PŘÍČINA?

53
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Dieta 1. Změněná produkce inzulínu, glukagonu a katecholaminů 2. Změna v dostupnosti substrátů

54
příjem sacharidů - glykémie - inzulín
Inzulín reguluje GLUT4 (transportní protein) Inzulín se váže na inzulínový receptor a fosfátová skupina z ATP se váže na proteinovou molekulu Fosforylace proteinů změní jejich vlastnosti = translokace GLUT4 z intracelulárních zásobních míst k buněčné membráně Inzulín tím zvýší maximální rychlost transportu, ale pouze v těch buňkách, které mají geny GLUT4 (sval a tuk). Ale: Při práci inzulín klesá, avšak pracující sval má zvýšenou schopnost získávat GL z krve, nezávisle na vlivu inzulínu!!! Tento vliv svalových kontrakcí přetrvává i v časné fázi zotavení = doplnění zásob energetických zdrojů
 Inzulín se váže na inzulínový receptor a fosfátová skupina z ATP se váže na proteinovou molekulu. Fosforylace proteinů změní jejich vlastnosti = translokace GLUT4 z intracelulárních zásobních míst k buněčné membráně. Inzulín tím zvýší maximální rychlost transportu, ale pouze v těch buňkách, které mají geny GLUT4 (sval a tuk). Ale: Při práci inzulín klesá, avšak pracující sval má zvýšenou schopnost získávat GL z krve, nezávisle na vlivu inzulínu!!! Tento vliv svalových kontrakcí přetrvává i v časné fázi zotavení = doplnění zásob energetických zdrojů.")
55
příjem sacharidů - glykémie - inzulín
Nízký inzulín sníží rychlost transportu v těch buňkách, které mají geny GLUT4 (sval a tuk). Opačný vliv vysokosacharidové a nízkosacharidové diety
. Opačný vliv vysokosacharidové. a nízkosacharidové diety.")
56
Adrenalin - hormon dřeně nadledvin
Při tělesné práci stoupá v závislosti na intenzitě zatížení (čím vyšší IZ, tím vyšší produkce adrenalinu) Zvyšuje glykogenolýzu v játrech i ve svalech Zvyšuje uvolňování glukózy a FFA do krve Má opačnou dynamiku i účinek jako inzulín = zvyšuje glykémii
 Zvyšuje glykogenolýzu v játrech i ve svalech. Zvyšuje uvolňování glukózy a FFA do krve. Má opačnou dynamiku i účinek. jako inzulín = zvyšuje glykémii.")
57
Glukagon - hormon produkovaný alfa-buňkami pankreatu
Při tělesné práci stoupá Zvyšuje glykogenolýzu v játrech i ve svalech Zvyšuje uvolňování glukózy a FFA do krve Jeho produkce je snížená při zvýšené dodávce sacharidů a tuků potravou Jeho produkce je stimulovaná adrenalinem Má opačnou dynamiku i účinek jako inzulín = zvyšuje glykémii

58
Při tělesné práci adrenalin stoupá - glykémie
glukagon stoupá - glykémie inzulín klesá - glykémie Přes zvýšenou spotřebu GL a GG, která by sama o sobě vedla k poklesu svalového a jaterního GG a glykémie, se výsledná glykémie většinou nemění, pouze při dlouhodobé intenzivní práci může dojít k hypoglykémii (vyčerpání glykogenu v játrech i ve svalech)
")
59
Při dietě chudé na sacharidy
adrenalin stoupá - glykémii glukagon stoupá - glykémii inzulín klesá - glykémii Podobná dynamika uvedených hormonů jako při tělesné práci - společná snaha zabránit hypoglykémii, i přes nedostatečný příjem sacharidů potravou

60
Hladovění mobilizuje tuky a snižuje utilizaci svalového GG během práce. Snížená produkce inzulínu a zvýšená produkce adrenalinu zvyšují aktivitu hormon-senzitivní lipázy - tím zvyšují metabolizaci tuků v adipoctytech - a tím zvyšují uvolňování MK do oběhu (akcentování tuku jako energetického zdroje).
.")
61
Redukční dieta + pravidelné cvičení
mobilizuje tuky a zvyšuje jejich využití jako energetického substrátu. Výsledkem je redukce tukové tkáně. Samostatná redukční dieta vede k redukci aktivní tělesné hmoty (zejména svalů) a ke ztrátám vody! Ztráty tuků jsou relativně menší. Samostatná redukční dieta je 1. méně účinná než v kombinaci se cvičením, 2. nezdravá (ztráta svalů a vody).
 a ke ztrátám vody! Ztráty tuků jsou relativně menší. Samostatná redukční dieta je. 1. méně účinná než v kombinaci se cvičením, 2. nezdravá (ztráta svalů a vody).")
62
Při dietě bohaté na sacharidy
adrenalin klesá - glykémii glukagon klesá - glykémii inzulín stoupá - glykémii Společná snaha zabránit hyperglykémii, ke které by mohlo dojít při vysokém příjmu sacharidů potravou

63
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Dieta 1. Změněná produkce inzulínu, glukagonu a katecholaminů 2. Změna v dostupnosti substrátů

64
Změna v dostupnosti substrátů
Při dostatku sacharidů v potravě se stávají sacharidy hlavním energetickým zdrojem při pohybové aktivitě - výhodné při intenzivní zátěži. Při nedostatku sacharidů v potravě se stávají hlavním energetickým zdrojem tuky (při dlouhodobém hladovění i bílkoviny - kachektizace) - tuky nejsou výhodným energetickým zdrojem při intenzivní zátěži.
 - tuky nejsou výhodným energetickým zdrojem při intenzivní zátěži.")
65
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Prostředí Tepelný stres zvyšuje glykogenolýzu ve svalech (zvýšený adrenalin - stresový hormon) tvorbu laktátu (redukce krevního průtoku a tím i kyslíku) glykémii (zvýšený adrenalin) Po aklimatizaci na zvýšenou teplotu okolí se glykogenolýza a produkce laktátu redukují. Stejný efekt na utilizaci substrátů při práci má trénink (snižuje adrenalin a zvyšuje prokrvení svalů).
 tvorbu laktátu (redukce krevního průtoku a tím i kyslíku) glykémii (zvýšený adrenalin) Po aklimatizaci na zvýšenou teplotu okolí se. glykogenolýza a produkce laktátu redukují. Stejný efekt na utilizaci substrátů při práci má. trénink (snižuje adrenalin a zvyšuje prokrvení svalů).")
66
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Prostředí Hypoxie (nedostatek kyslíku) zvyšuje glykogenolýzu (zvýšený adrenalin - stresový hormon) zvyšuje tvorbu laktátu (nedostatek kyslíku) Po aklimatizaci na hypoxii se glykogenolýza a produkce laktátu redukují (zvýšené využití FFA a snížená produkce adrenalinu). Stejný efekt na utilizaci substrátů při práci má trénink (snižuje adrenalin a zvyšuje využití tuků).
 zvyšuje glykogenolýzu (zvýšený adrenalin - stresový hormon) zvyšuje tvorbu laktátu (nedostatek kyslíku) Po aklimatizaci na hypoxii se glykogenolýza. a produkce laktátu redukují (zvýšené využití FFA. a snížená produkce adrenalinu). Stejný efekt na utilizaci substrátů při práci má. trénink (snižuje adrenalin a zvyšuje využití tuků).")
67
Faktory ovlivňující metabolismus sacharidů během tělesné práce
Pohlaví Ženy mají relativně větší předpoklady pro vytrvalostní práci relativně lepší využití tuků relativně horší využití sacharidů

68
SHRNUTÍ Svalový GG a krevní GL jsou důležité substráty pro resyntézu ATP v kontrahujících se svalech během tělesné práce. Kyselina mléčná je důležitý glukoneogenetický a oxidativní substrát. Intenzita svalové glykogenolýzy a spotřeby GL během práce je primárně determinovaná IZ a trváním práce.

69
SHRNUTÍ Další vlivy Druh zátěže Trénovanost Dieta Prostředí Pohlaví

70
SHRNUTÍ Regulace metabolismu sacharidů zahrnuje interakci mezi
lokálně vzniklými změnami svalového vápníku, anorganického fosforu a dalších metabolitů hormony dostupností substrátů.

71
SHRNUTÍ KONEC KAPITOLY
Zvýšená svalová glykogenolýza a spotřeba GL tak reagují na metabolické požadavky vzniklé během práce. KONEC KAPITOLY

Podobné prezentace












.>")









