Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Maticové modely Kategorie definované jako věkové třídy (častější u živočichů) Velikostní kategorie (častější u rostlin) Všechny parametry jsou konstantní (není nutné, ale pak se to špatně analyzuje)
 Velikostní kategorie (častější u rostlin) Všechny parametry jsou konstantní (není nutné, ale pak se to špatně analyzuje)")
2
Věkové třídy; předpokládám, že šírka kategorie a délka kroku jsou stejné (obvykle jedna sezóna). Potom Už bez nulových položek Obecně: P - pravděpodobnosti přežití, F - Fertility, počet živých potomků na jedno individuum za časový interval.

3
pro zjednodušení nahradíme zápisem n(t+1)=An(t) Počítáme:
=An(t) Počítáme:")
4
n(t+1)=An(t) logicky taky platí n(t+2)=An(t+1) atd. tedy n(t+k)=A k n(t) nebo se dá psát n(t)=A t n(0)
=A k n(t) nebo se dá psát n(t)=A t n(0).")
5
když vektor mnohokrát (před)násobím touže maticí (nebo násobím A k, kde k je dost vysoké), dostanu dominantní charakteristický vektor dané matice (jsou výjimky pro „divný“ matice). Z maticové algebry Když n je charakteristickým vektorem, můzeme matici v rovnici n(t+1)=An(t) nahradit jejím charakteristickým číslem λ a rovnici přepsat n(t+1)= λ n(t) pak bude pro součet všech individuí N=Σn i také platit N t+1 = λ N t a dostáváme klasický exponenciální růst
=An(t) nahradit jejím charakteristickým číslem λ a rovnici přepsat n(t+1)= λ n(t) pak bude pro součet všech individuí N=Σn i také platit N t+1 = λ N t a dostáváme klasický exponenciální růst.")
6
Celková velikost populace

7
Relativní zastoupení jedné kategorie se ustaluje

8
Smysl jednotlivých parametrů závisí na tom, jaká je to populace, a kdy dělám census!!! - Protože krok je jeden rok, může mě zajímat jen jeden okamžik v roce. Zajíci: rodí v dubnu, census dělám těsně než rodí (prebreeding c.), v úvahu beru pouze samice, potom v rovnici: N i - počet samic věku 1 až 4. V dubnu jsou jednoleté nejmladší. P i,j - pravděpodobnost přežití z věkové kategorie j do i F i - fertilita, vyjádřená jako počet samičích mláďat, které přežijí do jara dalšího roku, na jednu samici dané věkové kategorie
, v úvahu beru pouze samice, potom v rovnici: N i - počet samic věku 1 až 4. V dubnu jsou jednoleté nejmladší. P i,j - pravděpodobnost přežití z věkové kategorie j do i F i - fertilita, vyjádřená jako počet samičích mláďat, které přežijí do jara dalšího roku, na jednu samici dané věkové kategorie.")
9
F 3 bych tedy odhadl jako (přepokládám sex ratio 1:1) průměrný počet mláďat na jednu tříletou samici/2 * pravděpodobnost, že mládě přežije první zimu Tak, jak je model formulován, předpokládá, že plodné jsou jen samice tří- a čtyřleté, čtyřleté ještě porodí, ale další zimu už nepřežijí. Mláďata, která první zimu nepřežijí se v modelu vůbec nevyskytnou (v prvním dubnu ještě, v druhém dubnu už nejsou)
.")
10
Census dělám v květnu, malí zajíčči už jsou narození (postbreeding c.)(a zase mě zajímají pouze samice). Pokud bude situace stejná, potom model musí vypadat pak N 1 je počet čerstvých mláďat, N 2 počet jednoletých atd., N 5 je počet čtyřletých. Hodnoty P i,j jsou opět příslušné pravděpodobnosti přežívání, hodnoty F jsou nyní průměrné počty vyvedených (samičích) mláďat na jednu samici (živě narozená mláďata přežívající do doby censu) násobená pravděpodobností, že samice přežije. Ale pozor! Samice rodí až rok po censu. Proto F 3 je průměrný počet mláďat na samici tříletou, tj. samici čtvrté věkové kategorie, násobený pravděpodobností, že dvouletá samice přežije rok, tj. P 4,3.
 mláďat na jednu samici (živě narozená mláďata přežívající do doby censu) násobená pravděpodobností, že samice přežije. Ale pozor. Samice rodí až rok po censu. Proto F 3 je průměrný počet mláďat na samici tříletou, tj. samici čtvrté věkové kategorie, násobený pravděpodobností, že dvouletá samice přežije rok, tj. P 4,3..")
11
Pokud používám Leslieho matici pro jakýkoliv reálný systém, musím jednoznačně vědět, kterou roční dobu modeluji. Pokud populace rodí kontinuálně: za časový interval dochází ke třem procesům - umírají samice, rodí se mláďata (ale jen těm samicím, co do té doby přežily, a mláďata umírají), ale můžeme i tak parametry F odhadnout, z počtu individuí na začátku intervalu, a počtu jejich živých potomků na konci intervalu, nebo to můžeme relativně složitě vypočítávat.
, ale můžeme i tak parametry F odhadnout, z počtu individuí na začátku intervalu, a počtu jejich živých potomků na konci intervalu, nebo to můžeme relativně složitě vypočítávat..")
12
když vektor mnohokrát (před)násobím touže maticí (nebo násobím A k, kde k je dost vysoké), dostanu dominantní charakteristický vektor dané matice (jsou výjimky pro „divný“ matice). Z maticové algebry Když n je charakteristickým vektorem, můzeme matici v rovnici n(t+1)=An(t) nahradit jejím charakteristickým číslem λ a rovnici přepsat n(t+1)= λ n(t) pak bude pro součet všech individuí N=Σn i také platit N t+1 = λ N t a dostáváme klasický exponenciální růst
=An(t) nahradit jejím charakteristickým číslem λ a rovnici přepsat n(t+1)= λ n(t) pak bude pro součet všech individuí N=Σn i také platit N t+1 = λ N t a dostáváme klasický exponenciální růst.")
13
Charakteristický vektor představuje stabilní věkovou strukturu populace tj. struktura, kdy se poměrné zastoupený věkových kategorií nemění λ je růstová rychlost. Jak budeme v praxi parametry odhadovat?

14
Tabulky přežívání Mám kohortu. Tu sleduji po mnoho let. l(x) - věkově specifické přezívání; P(x) - pravděpodobnost přežití intervalu; 1-P(x) - věkově specifická mortalita; m(x) - věkově specifická plodnost (předpokládám sex ratio 1:1, proto je m počet mláďat dělený dvěma)
 - věkově specifické přezívání; P(x) - pravděpodobnost přežití intervalu; 1-P(x) - věkově specifická mortalita; m(x) - věkově specifická plodnost (předpokládám sex ratio 1:1, proto je m počet mláďat dělený dvěma).")
15
Prebreeding census, kategorie 1 jsou jednoletý Postbreeding census, kategorie 1 jsou čerstvě narozený

16
Čistá reprodukční rychlost R 0 =Σl(x).m(x) průměrný počet mláďat (samic) na samici za celý život Generační doba: vážený průměr věku porodu G= {Σl(x).m(x).x}/R 0 R0R0
.m(x) průměrný počet mláďat (samic) na samici za celý život Generační doba: vážený průměr věku porodu G= {Σl(x).m(x).x}/R 0 R0R0")
17
Nemám kohortu po mnoho let, ale mám různě stará individua ve dvou po sobě následujících letech. Platí l(x) =ΠP(x) /např. pravděpodobnost, že přežije do druhého roku, je pravděpodobnost že přežije první rok * pravděpodobnost, že přežije druhý rok. Řada metod odhadování pro spojitá data (např. Kaplan - Meier) Pravděpodobnost pro každou věkovou kategorii je procento přeživších z těch, co tam byli na začátku intervalu. Když mám kohortu (časově velmi náročné, “zuby” na křivce dané různými sezónami (např. 2002 se jich moc utopilo). Když mám jednu sezónu, tak jsou věkové kategorie ve srovnatelných podmínkách, ale vše je ovlivněné danou sezónou. Nejlépe - kombinovat.
 =ΠP(x) /např. pravděpodobnost, že přežije do druhého roku, je pravděpodobnost že přežije první rok * pravděpodobnost, že přežije druhý rok. Řada metod odhadování pro spojitá data (např. Kaplan - Meier) Pravděpodobnost pro každou věkovou kategorii je procento přeživších z těch, co tam byli na začátku intervalu. Když mám kohortu (časově velmi náročné, zuby na křivce dané různými sezónami (např se jich moc utopilo). Když mám jednu sezónu, tak jsou věkové kategorie ve srovnatelných podmínkách, ale vše je ovlivněné danou sezónou. Nejlépe - kombinovat..")
18
Statická life table Mám v jednom okamžiku věkové složení populace. Pouze pokud je λ=1 a populace má stabilní věkovou strukturu, může jako odhad pravděpodobnosti přežívání sloužit n x+1 /n x. POKUD NE, JE TO VELMI PROBLEMATICKÉ. (klasický příklad ovce podle mrtvol). narození smrt
. narození smrt.")
19
Funkce přežívání Log na svilsé ose! Konstantní mortalita je typ II je potom přímka. Typ I člověk na Západě, Typ III ryby. Typ I velmi vzácně. Když už, tak spíš mortalita mláďat bývá vysoká

20
Stage-classified (populace rozdělená do velikostních tříd) - často u rostlin Např. pro strom: Semenáč, malý sapling, velký sapling, malý dospělý velký dospělý Důsledky: může v kategorii zůstat, ale může i “přeskočit kategorii, případně jít do kategorie nižší => matice je “plnější”, nenulové hodnoty nejen pod diagonálou Pro odhad: bezpodmínečně potřebuji značená individua (u věku je možné vystačit si s počty)
.")
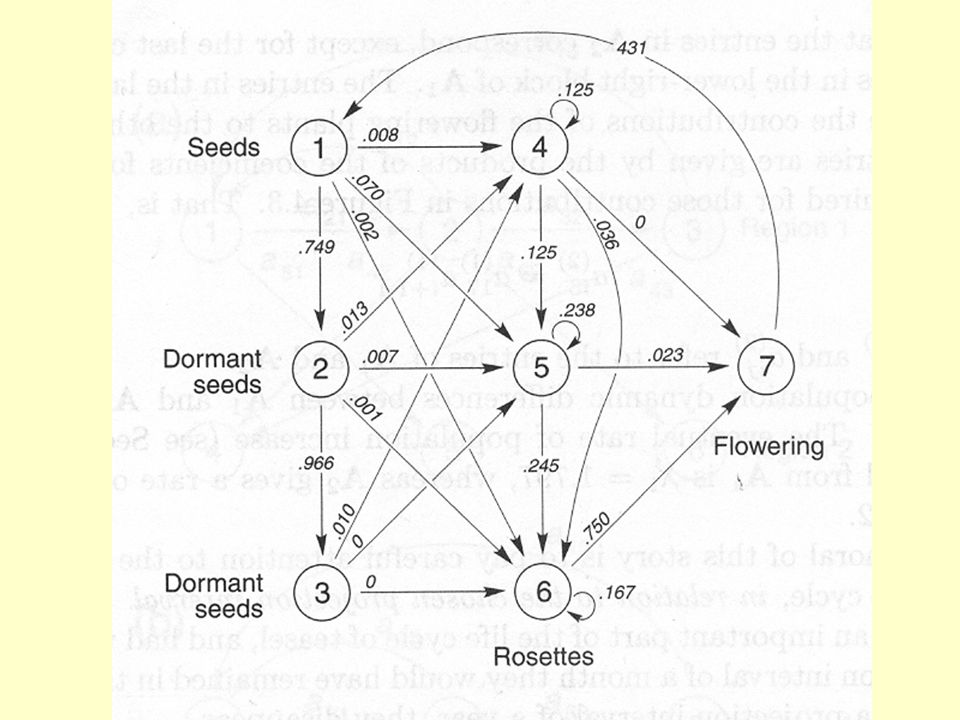
21
1-MR2-VR3-KV P 1,1 P 2,1 P 3,2 P 2,2 P 1,3 - Průměrný počet nových růžic vyprodukovaný jednou kvetoucí Monocarpic perennial census v době květu Předopokládám: každá kvetoucí chcípne, do květu může příští rok jít jen velká růžice, není zásoba semen. MR- malá růžice VR- velká růžice KV- kvetoucí

22
Nejsou zde semena (protože není semenná banka, tak v době před květem jsou z nich buď malé růžice, pokud nevyklíčily, jsou odsouzeny k zániku). Realistické může být i následující: tj. některá semena vyklíčí a hned 1-MR2-VR3-KV P 2,1 P 3,2 P 1,3 P 2,3 vytvoří velkou růžici

23
Problém odhadování: jak široké mají být kategorie (vždy arbitrární). Měly by být vnitřně homogenní a rozdílné mezi sebou. Široké: jsou heterogenní, a pak dávají nerealistický obraz. Úzké: pak je jich mnoho, a to vede k nižšímu počtu individuí pro odhad. POZOR, některé pravděpodobnosti mohou být blízké nule nebo jedničce, pak potřebujeme hodně individuí. Všeobecně: pravděpodobnosti přežívání pomocí značených individuí. “Recruitment”, tj. objevování se nových je obtížné, na základě nově se objevivších jedinců.

24
Špatná matice klasika λ=1,797

26
Takhle to má bejt správně (vymyslel Caswell sám) λ=2.322
 λ=2.322")
27
Platí Pokud jsou parametry konstantní, tak jak věková, tak matice z velikostních tříd vede na exponenciální model (neomezený růst nebo vymírání), z obojího odhadneme λ. V modelu totiž nikde není density dependence Paramtery modelu mohou odrážet i působení kompetice, ale protože se nemění v průběhu času, předpokládáme, že se kompetice též nemění (i kdyby se N měnilo)
.")
28
Projekce - co by se stalo, kdyby současné podmínky trvaly Predikce - co se stane. Proto jméno PROJEKČNÍ MATICE

29
Užití výhoda - jedná se o modely, jejich parametry lze rozumně odhadnout Odhaduji λ - odhad, zda populace roste nebo vymírá. Mohu kombinovat matice (pro suchou a vlhkou sezonu) Z dvouletého sledování mohu odhadnout matici, a z ní potom stabilní věkovou (velikostní) strukturu. Tu porovnám s realitou. Pokud není, hledám příčiny Porovnávám několik po sobě jdoucích přechodů, odhaduji variabilitu. Population viability analysis.
 Z dvouletého sledování mohu odhadnout matici, a z ní potom stabilní věkovou (velikostní) strukturu. Tu porovnám s realitou. Pokud není, hledám příčiny Porovnávám několik po sobě jdoucích přechodů, odhaduji variabilitu. Population viability analysis..")
30
Analýza senzitivity (citlivost na absolutní změnu velikosti parametru) a elasticity (citlivost na relativní změnu velikosti parametru) Vochysia ferruginea Matice senzitivita elasticita
 a elasticity (citlivost na relativní změnu velikosti parametru) Vochysia ferruginea Matice senzitivita elasticita")
31
Vochysia ferruginea

32
Analýza senzitivity (citlivost na absolutní změnu velikosti parametru) a elasticity (citlivost na relativní změnu velikosti parametru) Vochysia ferruginea Matice senzitivita elasticita Elasticitu nejsem schopen počítat pro nuly – spíš bych měl psát n.a. – Součet všech elasticit je 1

33
U každé sensitivity/elasticity Musím říci Čeho (nemusí to být vždy λ, ale třeba změna stabilní věkové struktury, dumping ratio u oscilujících), a vzhledem k čemu (můžu měnit jednotlivé členy matice, ale také víc členů najednou, třeba všechny plodnosti)
, a vzhledem k čemu (můžu měnit jednotlivé členy matice, ale také víc členů najednou, třeba všechny plodnosti)")
34
Na co slouží analýza senz./elast. Každý parametr je odhadnut s nějakou chybou – parametry, s velkou senzitivitou- elasticitou (tj. na jejichž změnu je cílová funkce citlivá) by měly být odhadnuty velmi přesně Managementové úvahy – o změnu kterého parametru se mám managementem snažit Evoluční úvahy
 by měly být odhadnuty velmi přesně Managementové úvahy – o změnu kterého parametru se mám managementem snažit Evoluční úvahy.")
35
* Můžu mít několik matic za sebou pro různé části sezóny (pozor záleží na pořadí) - zjišťuji, co je důležité v které části roku Rozšíření modelu * Můžu mít parametry stochastické, nebo závislé na vnějších podmínkách * Můžu mít parametry závislé na hustotě Pak budu většinou potřebovat simulační přístup
 - zjišťuji, co je důležité v které části roku Rozšíření modelu * Můžu mít parametry stochastické, nebo závislé na vnějších podmínkách * Můžu mít parametry závislé na hustotě Pak budu většinou potřebovat simulační přístup")
36
Individual based models Modely založené na chování individuí

37
Individual based models Mají (spatially explicit) nebo nemají popsáno umístění individuí v prostoru – ploše Neprostorové, tzv. JABOWA type models Nejsou nutně jednopopulační – vlastností individua je pak i druhová identita Každé individuum je popsáno stavovou proměnnou (nebo více proměnnými) V každém kroku, růst individua závisí na jeho velikosti, a na kompetici Podobně, pravděpodobnost prěžití je závislá na velikosti individua a kompetičním tlaku Deterministické (pravidlo rozhoduje, kdo přežije), nebo stochastické (spočítám pravděpodobnost přežití, a pak Monte Carlo simulace rozhodne, zda přežije)
 V každém kroku, růst individua závisí na jeho velikosti, a na kompetici Podobně, pravděpodobnost prěžití je závislá na velikosti individua a kompetičním tlaku Deterministické (pravidlo rozhoduje, kdo přežije), nebo stochastické (spočítám pravděpodobnost přežití, a pak Monte Carlo simulace rozhodne, zda přežije).")
38
„Fibichův“ model

39
Většinou simulační modely Velikost individua i (t+1) = Velikost individua i (t) + přírůst přírůst = fce 1 (Velikost individua i (t), kompetice) Bez kompetice Pravděpodobnost přežití = fce 2 (Velikost individua i (t), kompetice) velikost ind. přír. max. vel. indiv.

40
Individuum je popsáno jednou proměnnou - ostatní jsou alometrickými charakteristikami realističtější Více proměnnými, např. Výška a DBH kompetice přírůstvýškový kompetice přírůst DBH Stromy v mírné kompetici rostou do výšky, ale netloustnou

41
Nebo mechanističtěji Odhadnu biomasu, kterou za jednotku daný jedinec vytvoří Podle kompetice určím, jakou část bude investovat do výškového růstu, a jakou část do horizontálního růstu (charakterizovaného třeba šířkou koruny)
")
42
Jak počítám kompetici 1, Na základě density (plus velikostní struktury, včetně biomasy) individuí v celé simulované ploše 2. Na základě těch, co jsou blízko. Potom potřebuju pro každé individuum jeho umístění v ploše, tzv. spatially explicit models (jde jen u rostlin, resp. u sedentárních organismů)
.")
43
Spatially explicit models - the competition is based on the neighbours only

46
Problém – co s rozmnožováním Jednodušší u těch, co nejsou spatially explicit Problém (u stromů) – teoreticky jde nasimulovat každý rok semenáče jako nová individua (buď s lokalizací, nebo bez ní) – ale chci-li slušný porost, potřebuji 50m x 50m, abych měl dost velkých stromů – a na m 2 mohou být i desítky semenáčů.
 – teoreticky jde nasimulovat každý rok semenáče jako nová individua (buď s lokalizací, nebo bez ní) – ale chci-li slušný porost, potřebuji 50m x 50m, abych měl dost velkých stromů – a na m 2 mohou být i desítky semenáčů.")
47
Často řesení pomocí kombinace – seedling recruitment a mladá stádia simulovaná pomocí maticového modelu s density dependencí, starší stádia pak jako IBM

Podobné prezentace