Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Rozkladači a detritovoři

2
Rozklad MRTVÉ organické hmoty
celá řada predátorů žere mrtvé tkáně, protože kořist nejprve zabije. Ale rozkladači a detritovoři (decomposers & detritovores), na rozdíl od predátorů (včetně parazitů a herbivorů), neovlivňují PŘÍMO rychlost tvorbu svých potravních zdrojů. (Výjimka – někteří nekrotrofní paraziti: žerou nejdřív živé, a pak i mrtvé tkáně uhynulých hostitelů) Vztah mezi rozkladači-detritovory a jejich zdroji se dá popsat jako řízený dárcem (donor-controlled). Dynamika vztahu se v mnoha bodech liší od klasické L&V dynamiky dravec-kořist, nejdůležitějším rozdílem je STABILITA těchto vztahů, která je navíc nezávislá (nebo dokonce pozitivně úměrná) na zvyšující se druhové bohatosti a komplexitě trofických vazeb. Existuje ovšem zjevná NEPŘÍMÁ zpětná vazba mezi rozkladači/detritovory a jejich zdroji: uvolněné živiny jsou opět k dispozici primárním producentům. Ve svém důsledku je tato skutečnost nejdůležitější rolí detritovorů / rozkladačů v ekosystémech. Zdrojem pro rozkladače a detritovory nemusí být nutně pouze mrtvá těla jiných organismů. Mnoho unitárních organismů se během svého životního cyklu zbavuje „opotřebovaných“ částí – exoskelety u členovců, pokožka, peří, srst a paroží u obratlovců, a tyto zdroje mají své specialisty. Nepřetržité „odhazování“ mrtvých částí je ještě charakterističtější pro organismy modulární – koráli, sezónní opad listů, buňky z růstových vrcholů kořenů ( rhizosféra). A v neposlední řadě nelze zapomínat na organický odpad, který vypadává z opačného konce trávicí trubice konzumentů.
, na rozdíl od predátorů (včetně parazitů a herbivorů), neovlivňují PŘÍMO rychlost tvorbu svých potravních zdrojů. (Výjimka – někteří nekrotrofní paraziti: žerou nejdřív živé, a pak i mrtvé tkáně uhynulých hostitelů) Vztah mezi rozkladači-detritovory a jejich zdroji se dá popsat jako řízený dárcem (donor-controlled). Dynamika vztahu se v mnoha bodech liší od klasické L&V dynamiky dravec-kořist, nejdůležitějším rozdílem je STABILITA těchto vztahů, která je navíc nezávislá (nebo dokonce pozitivně úměrná) na zvyšující se druhové bohatosti a komplexitě trofických vazeb. Existuje ovšem zjevná NEPŘÍMÁ zpětná vazba mezi rozkladači/detritovory a jejich zdroji: uvolněné živiny jsou opět k dispozici primárním producentům. Ve svém důsledku je tato skutečnost nejdůležitější rolí detritovorů / rozkladačů v ekosystémech. Zdrojem pro rozkladače a detritovory nemusí být nutně pouze mrtvá těla jiných organismů. Mnoho unitárních organismů se během svého životního cyklu zbavuje „opotřebovaných částí – exoskelety u členovců, pokožka, peří, srst a paroží u obratlovců, a tyto zdroje mají své specialisty. Nepřetržité „odhazování mrtvých částí je ještě charakterističtější pro organismy modulární – koráli, sezónní opad listů, buňky z růstových vrcholů kořenů ( rhizosféra). A v neposlední řadě nelze zapomínat na organický odpad, který vypadává z opačného konce trávicí trubice konzumentů.")
3
Kdo jsou rozkladači a detritovoři?
Rozkladači (decomposers) ranní kolonisté - bakterie a houby: jsou všudypřítomné > jsou obvykle prvními (spolu s nekrotrofními parazity) na „zdroji“. Využívají především rozpustné látky obsahující aminokyseliny a cukry, nemají ezymy na štípání chitinu, celulozy, ligninu či keratinu. např. Penicillium, Mucor, Rhizopus, kvasinky, bakterie (Lactobacillus) V areobních podmínkách je výsledným produktem CO2, v anaerobních alkoholy a organické kyseliny. V anaerobních sedimentech často bakterie s jinými typy metabolismu, kdy je místo kyslíku je donorem elektronů nějaká anorganická látka. pozdní kolonisté – specializované bakterie a houby rezistence organických látek vůči dekompozici roste zhruba v tomto pořadí: cukry < škrob < hemicelulózy, pektiny a proteiny < celulóza < lignin < suberin (korek)< kutin (kutikula). Specializované bakterie a houby už mají příslušné ezymy na štípání chitinu, celulozy, ligninu či keratinu, spíše ze skupin Actinomycetes, Basidiomycetes a Ascomycetes. Často pozorovatelná posloupnost = sukcese hub a/nebo bakterií.
 ranní kolonisté - bakterie a houby: jsou všudypřítomné > jsou obvykle prvními (spolu s nekrotrofními parazity) na „zdroji . Využívají především rozpustné látky obsahující aminokyseliny a cukry, nemají ezymy na štípání chitinu, celulozy, ligninu či keratinu. např. Penicillium, Mucor, Rhizopus, kvasinky, bakterie (Lactobacillus) V areobních podmínkách je výsledným produktem CO2, v anaerobních alkoholy a organické kyseliny. V anaerobních sedimentech často bakterie s jinými typy metabolismu, kdy je místo kyslíku je donorem elektronů nějaká anorganická látka. pozdní kolonisté – specializované bakterie a houby. rezistence organických látek vůči dekompozici roste zhruba v tomto pořadí: cukry < škrob < hemicelulózy, pektiny a proteiny < celulóza < lignin < suberin (korek)< kutin (kutikula). Specializované bakterie a houby už mají příslušné ezymy na štípání chitinu, celulozy, ligninu či keratinu, spíše ze skupin Actinomycetes, Basidiomycetes a Ascomycetes. Často pozorovatelná posloupnost = sukcese hub a/nebo bakterií.")
4
Kdo jsou rozkladači a detritovoři?
Detritovoři (a mikrobovoři) Mikrobovoři (ti, co žerou bakterie a jiné mikroorganismy) jdou obyčejně s rozkladači ruku v ruce a občas se od nich špatně odlišují. Žerou ale živé buňky. Jsou to (narozdíl od rozkladačů) obvykle volně žijící pohyblivé organismy (améby, nematodi – Pelodera, Ditylenchus) Detritovoři – čím jsou větší, tím hůře rozlišují mezi detritem a mikroorganimsy jej rozkládající. V terestrických ekosystémech škatulkování obvykle podle velikosti: mikrofauna – protozoa, nematodi a vířníci mesofauna – (mezi 100 um a 2 mm) roztoči, chvostoskoci, Enchytreidae makrofauna (2 – 20 mm) a megafauna (> 20 mm) – Isopoda, Diplopoda, Annelida, Mollusca, larvy dvoukřídlých a brouků Darwin (1888): okolo jeho domu žížaly během 30 let vyprodukovaly vrstvu 18 cm nové půdy a na povrch vynesly každý rok tj. 5.1 kg.m-2. V Nigerii až 17.3 kg.m-2 během období dešťů (tj. 2-6 měsíců) – a to jen na povrch, skutečné číslo musí být mnohem vyšší.
 Mikrobovoři (ti, co žerou bakterie a jiné mikroorganismy) jdou obyčejně s rozkladači ruku v ruce a občas se od nich špatně odlišují. Žerou ale živé buňky. Jsou to (narozdíl od rozkladačů) obvykle volně žijící pohyblivé organismy (améby, nematodi – Pelodera, Ditylenchus) Detritovoři – čím jsou větší, tím hůře rozlišují mezi detritem a mikroorganimsy jej rozkládající. V terestrických ekosystémech škatulkování obvykle podle velikosti: mikrofauna – protozoa, nematodi a vířníci. mesofauna – (mezi 100 um a 2 mm) roztoči, chvostoskoci, Enchytreidae. makrofauna (2 – 20 mm) a megafauna (> 20 mm) – Isopoda, Diplopoda, Annelida, Mollusca, larvy dvoukřídlých a brouků. Darwin (1888): okolo jeho domu žížaly během 30 let vyprodukovaly vrstvu 18 cm nové půdy a na povrch vynesly každý rok tj. 5.1 kg.m-2. V Nigerii až 17.3 kg.m-2 během období dešťů (tj. 2-6 měsíců) – a to jen na povrch, skutečné číslo musí být mnohem vyšší.")
5
Relativní význam detritovorů v terestrických ekosystémech
Záleží hodně na teplotě a množství vody (její nedostatek limituje pohyblivé mikroorganismy)
")
6
Detritovoři ve vodních ekosystémech
Nejde o velikost, ale spíš o způsob získávání potravy Např. v tocích: drtiči (shredders) – živí se velkými částicemi, které drtí na menší. Larvy chrostíků (např. Stenophylax), blešivci a Isopoda – v tocích často na opadaných listech. Pak sběrači (collectors) – a to buď smetači (gatherers; larvy jepic Ephemera, larvy pakomárů, nítěnky) nebo filtrátoři (larvy muchniček, larvu chrostíků Hydropsyche). Dále škrabači (grazers-scrapers) oškrabávají vrstvy nárostů (např. larvy jepic Heptagenia, někteří chrostíci se schránkami - Glossosoma) Ve vodě obyčejně generalisté - rozdíl od suchozemských detritovorů.
 – živí se velkými částicemi, které drtí na menší. Larvy chrostíků (např. Stenophylax), blešivci a Isopoda – v tocích často na opadaných listech. Pak sběrači (collectors) – a to buď smetači (gatherers; larvy jepic Ephemera, larvy pakomárů, nítěnky) nebo filtrátoři (larvy muchniček, larvu chrostíků Hydropsyche). Dále škrabači (grazers-scrapers) oškrabávají vrstvy nárostů (např. larvy jepic Heptagenia, někteří chrostíci se schránkami - Glossosoma) Ve vodě obyčejně generalisté - rozdíl od suchozemských detritovorů.")
7
Relativní význam detritovorů a dekompozitorů
Porovnáváme-li počty, vedou bakterie. Porovnáváme-li biomasu, tak mikroflora taky převažuje. Ale biomasa a počty špatným měřítkem, protože různé generační doby a metabolické aktivity. Proces rozkladu organické hmoty není pouhým součtem činností rozkladačů a detritovorů, ale častěji výsledek jejich interakce. Fragmentace napomáhá dekompozici a vice versa.

8
Relativní význam detritovorů a dekompozitorů
Poměr látek ve zdroji vs. rychlost jeho rozkladu dekompozice závisí na poměru C : N. Pokud v rostlinném materiálu C : N = 40:1, tak rozklad pomalejší, protože dekompozitoři (mikroflora) potřebují asi 10:1. V případě zbytků živočichů tyto problémy nejsou – poměr C:N v jejich tělech zhruba stejný jako v rozkladačích. To má některé významné následky – např. po sezónním odumření rostlinné hmoty dochází k deficitu dusíku v půdě, protože se přesouvá do mikroorganismů. Zaorávání strnišť – deficit dusíku příští rok.
 potřebují asi 10:1. V případě zbytků živočichů tyto problémy nejsou – poměr C:N v jejich tělech zhruba stejný jako v rozkladačích. To má některé významné následky – např. po sezónním odumření rostlinné hmoty dochází k deficitu dusíku v půdě, protože se přesouvá do mikroorganismů. Zaorávání strnišť – deficit dusíku příští rok.")
9
Interakce mezi detritovory a zdroji
Konzumace rostlinných zbytků celulóza a lignin. Katabolismus celulózy vyžaduje celulázu. Ovšem tu umí vyrábět jen málo detritovorů. U živočichů jsou důkazy o přímé tvorbě jen u termitů podčeledi Nasutitermitinae a u jednoho švába. Ostatní detritovoři (a herbivoři) závisí na produkci celuláz jinými organismy, a to jak mikroflórou, tak někdy i prvoky. Vztah mezi detritovorem a producenty celuláz sahá od obligátního mutualismu (specializovaná stálá mikroflora a mikrofauna v žaludku) přes fakultativní mutualismus (živočichové využívají celulázu produkovanou mikroorganismy, které sežerou s detritem) k formě „vnějšího bachoru“, kdy živočichové asimilují již rozložené látky, metabolické produkty mikroorganismů s celulázou. Příklady: obligátní symbiosa Termiti (Eutermes) – symbiotická protozoa až 60% hmotnosti termita. Nedovedou lignin, což je u termitů obecné, i když Reticulitermes naopak umí. částečná symbiosa terestrická Isopoda – pokusy s antibiotiky a následným krmením. asimilace rozložených metabolitů - chvostoskoci, nejdokonalejší příklady termiti a mravenci, co si aktivně zakládají a pěstují houbové kolonie.
 závisí na produkci celuláz jinými organismy, a to jak mikroflórou, tak někdy i prvoky. Vztah mezi detritovorem a producenty celuláz sahá od obligátního mutualismu (specializovaná stálá mikroflora a mikrofauna v žaludku) přes fakultativní mutualismus (živočichové využívají celulázu produkovanou mikroorganismy, které sežerou s detritem) k formě „vnějšího bachoru , kdy živočichové asimilují již rozložené látky, metabolické produkty mikroorganismů s celulázou. Příklady: obligátní symbiosa Termiti (Eutermes) – symbiotická protozoa až 60% hmotnosti termita. Nedovedou lignin, což je u termitů obecné, i když Reticulitermes naopak umí. částečná symbiosa terestrická Isopoda – pokusy s antibiotiky a následným krmením. asimilace rozložených metabolitů - chvostoskoci, nejdokonalejší příklady termiti a mravenci, co si aktivně zakládají a pěstují houbové kolonie.")
10
Interakce mezi detritovory a zdroji
Konzumace exkrementů Od masožravců – špatná kvalita, protože potrava je využita s vysokou účinností (až 80%). Navíc je toho relativně málo, stejně jako jeho původců. Od býložravců – větší pochoutka, protože hodně nestrávených zbytků, na kterých se leckdo uživí. Příklad sloního trusu: v období dešťů je trus ihned (během 24 hodin) rozkouskován a „pohřben“ chrobáky. V období sucha je jen velmi málo broučích imág, proto mikrobi mají relativně hodně času, ale jde jim to pomalu – až dva roky (protože brouci nežerou starý trus, ani když se namočí). V Austrálii problém s kravincema, protože na kontinentu chybí na kravince specializovaní detritofágové. Ti místní „umějí“ jen suchý trus klokanů. Proto v 1963 introdukováni i afričtí chrobáci. Tento problém je navíc násoben existencí domácích i introdukovaných much, které kladou do exponovaných kravinců a způsobovaly svým přemnožením kalamity. Po introdukci brouků trochu lepší.
. Navíc je toho relativně málo, stejně jako jeho původců. Od býložravců – větší pochoutka, protože hodně nestrávených zbytků, na kterých se leckdo uživí. Příklad sloního trusu: v období dešťů je trus ihned (během 24 hodin) rozkouskován a „pohřben chrobáky. V období sucha je jen velmi málo broučích imág, proto mikrobi mají relativně hodně času, ale jde jim to pomalu – až dva roky (protože brouci nežerou starý trus, ani když se namočí). V Austrálii problém s kravincema, protože na kontinentu chybí na kravince specializovaní detritofágové. Ti místní „umějí jen suchý trus klokanů. Proto v 1963 introdukováni i afričtí chrobáci. Tento problém je navíc násoben existencí domácích i introdukovaných much, které kladou do exponovaných kravinců a způsobovaly svým přemnožením kalamity. Po introdukci brouků trochu lepší.")
11
Interakce mezi detritovory a zdroji
Konzumace mršin Od předchozích se hodně liší, a je to vidět už na skladbě enzymů. Karbohydrázy jsou řídké a málo aktivní, naopak jsou četné lipázy a proteázy. V zásadě totéž, co mají predátoři-masožravci. Nakonec, řada z nich se živí i mršinami (polární lišky, chaluhy, vrány, jezevci, luňáci, hyeny apod.). To, co nechají velcí mrchožrouti, je k dispozici mikroflóře a dalším, jako například larvám masařek. Ale záleží na počasí, pokud moc sucho, tak mumifikace. (Ta může být i v zimě). I mezi mrchožrouty najdeme specialisty na špatně rozložitelné tkáně. Např. masařky Lucilia mají enzym kolagenázu. Můry a kožojedi se zase potýkají s keratinem v peří a srsti pomocí silně redukujících látek vylučovaných do žaludku. Tak se rozruší vazby mezi peptidovými řetězci, a nastoupí hydrolytické enzymy. Houby čeledi Onygenaceae jsou specialisty na rohy a peří. Obecně – mrtvoly velkých zvířat poskytují zdroje mnoha různým mrchožroutům. Naopak, mrtvolky malých zvířátek (dejme tomu slimáka) slouží jen pár hmyzům. Zajímavá je skupina „hrobníků“, tedy těch živočichů, kteří mršiny pohřbívají. Třeba dospělý hrobařík Necrophorus najde po čichu mrtvolu během 1-2 hodin. Začne žrát tkáně, a pokud je mrtvolka starší, též larvy masařek. Přitom začne mrtvolu pohřbívat (někdy si ji odtáhne až pár metrů jinam), některé druhy až do hloubky 20 cm. Během zahrabávání aktivně vyhání další hrobaříky (dokonce i souboje na život a na smrt). Přiletí-li samička, začne pomáhat.
. To, co nechají velcí mrchožrouti, je k dispozici mikroflóře a dalším, jako například larvám masařek. Ale záleží na počasí, pokud moc sucho, tak mumifikace. (Ta může být i v zimě). I mezi mrchožrouty najdeme specialisty na špatně rozložitelné tkáně. Např. masařky Lucilia mají enzym kolagenázu. Můry a kožojedi se zase potýkají s keratinem v peří a srsti pomocí silně redukujících látek vylučovaných do žaludku. Tak se rozruší vazby mezi peptidovými řetězci, a nastoupí hydrolytické enzymy. Houby čeledi Onygenaceae jsou specialisty na rohy a peří. Obecně – mrtvoly velkých zvířat poskytují zdroje mnoha různým mrchožroutům. Naopak, mrtvolky malých zvířátek (dejme tomu slimáka) slouží jen pár hmyzům. Zajímavá je skupina „hrobníků , tedy těch živočichů, kteří mršiny pohřbívají. Třeba dospělý hrobařík Necrophorus najde po čichu mrtvolu během 1-2 hodin. Začne žrát tkáně, a pokud je mrtvolka starší, též larvy masařek. Přitom začne mrtvolu pohřbívat (někdy si ji odtáhne až pár metrů jinam), některé druhy až do hloubky 20 cm. Během zahrabávání aktivně vyhání další hrobaříky (dokonce i souboje na život a na smrt). Přiletí-li samička, začne pomáhat.")
12
Interakce mezi detritovory a zdroji
pokračování: Pohřbením je sice mršina méně vystavena dalším zájemcům, ale přesto má další ochranu díky mutualistickým roztočům Poecilochirus necrophori, kteří se nechají přenášet brouky od mršiny k mršině. Při zahrabávání brouci sice odstraní veškerou srst s vajíčky masařek, ale pokud je mršina pohřbená jen mělce, tak se tam masařky mohou znovu dostat. Pokud se tak stane, roztoči vajíčka masařek napíchnou a vysají. Pak brouci vymodelují z odchlupené/odpeřené mršiny kouli, a po kopulaci samička snese do chodbičky vedle mršiny vajíčka. Obvykle přinejmenším jeden dospělec zůstává, aby se o mršinu „staral“ – má se za to, že látky ze slin a exkrementů hrobaříka potlačují růst plísní. Po vylíhnutí larviček vyhloubí brouk na vrchu mršiny důlek, do kterého vyvrhává kapky natrávené potravy. Na nich se larvičky živí, dokud nejsou samy schopny žrát tkáně. Dospělec opouští mršinu, když se larvy zakuklí.

13
SOUHRN Nízká aktivita dekompozitorů a detritovorů je při nízké teplotě, špatné aeraci, nízkému obsahu vody v půdě a nízkém pH. Velký význam má porozita a struktura prostředí – většina detritovorů a rozkladačů se musí aktivně „prodírat“ prostředím, aby našla to, co hledá. Aktivita mikroflóry (rozkladačů) a detritovorů je tak vzájemně provázaná a synergická, že nemá skoro cenu se pídit, kdo je relativně významnější. Řada z nich jsou specialisté, a proto je rozklad organické hmoty dílem kombinovaných aktivit velmi různých organismů. Organická hmota může opakovaně „cyklovat“ skrz řadu mikrohabitatů uvnitř a vně žaludků a exkrementů různých organismů – nakonec skončí jako CO2 a minerální živiny. Aktivita dekompozitorů uvolňuje dusík a fosfor vázaný v mrtvých tkáních. Rychlost této dekompozice určuje dostupnost nutrientů pro rostliny. Mrtvé organismy jsou obvykle v prostředí rozmístěny ostrůvkovitě. Kolonizace takových zdrojů je částečně dílem náhody (kdo dřív přijde, ten …), a úspěšné druhy se mohou lišit případ od případu Dynamika kompetice mezi těmito druhy má své vlastní modely, často založené na „ostrovní“ koncepci. Nakonec, změňme optiku pohledu z úspěšnosti dekompozice na její selhávání – lesy existují jen díky neschopnosti mikroorganismů rozkládat rychle dřevo. Díky selhání mikroorganismů tu máme a užíváme uhlí, ropu a rašelinu!
 a detritovorů je tak vzájemně provázaná a synergická, že nemá skoro cenu se pídit, kdo je relativně významnější. Řada z nich jsou specialisté, a proto je rozklad organické hmoty dílem kombinovaných aktivit velmi různých organismů. Organická hmota může opakovaně „cyklovat skrz řadu mikrohabitatů uvnitř a vně žaludků a exkrementů různých organismů – nakonec skončí jako CO2 a minerální živiny. Aktivita dekompozitorů uvolňuje dusík a fosfor vázaný v mrtvých tkáních. Rychlost této dekompozice určuje dostupnost nutrientů pro rostliny. Mrtvé organismy jsou obvykle v prostředí rozmístěny ostrůvkovitě. Kolonizace takových zdrojů je částečně dílem náhody (kdo dřív přijde, ten …), a úspěšné druhy se mohou lišit případ od případu Dynamika kompetice mezi těmito druhy má své vlastní modely, často založené na „ostrovní koncepci. Nakonec, změňme optiku pohledu z úspěšnosti dekompozice na její selhávání – lesy existují jen díky neschopnosti mikroorganismů rozkládat rychle dřevo. Díky selhání mikroorganismů tu máme a užíváme uhlí, ropu a rašelinu!")
14
Symbiosa a mutualismus

15
Mutualismus vs. symbiosa
symbiosa = organismy „nerozlučně“ pohromadě, v blízkém fyzickém vztahu. mutualismus = širší pojem, popisující oboustranně výhodnou interakci mezi organismy. Mutualisté nemusí být nutně symbionty, i když řada z nich je. A naopak, ne všichni symbionti jsou mutualisty. Mutualismus – důležitější, než by se zdálo Mutualisté tvoří většinu biomasy na Zemi. Téměř všechny stepní a lesní rostliny jsou (svými kořeny) v úzkém symbiotickém vztahu s houbami. Většina korálů obsahuje jednobuněčné řasy, celá řada kvetoucích rostlin je závislá na opylovačích a mnoho živočichů má v zažívacím traktu mikroorganismy.
 v úzkém symbiotickém vztahu s houbami. Většina korálů obsahuje jednobuněčné řasy, celá řada kvetoucích rostlin je závislá na opylovačích a mnoho živočichů má v zažívacím traktu mikroorganismy.")
16
Mutualismus – typy vztahů
Vzájemné vazby v chování Mravenec – akácie (ochrana pro rostlinu, potrava a bydlo pro mravence) Ryby-čističi a jejich „zákazníci“ (čističi se živí ektoparazity, a hostitelé se jich zbavují) Medojed a medojedka (medojedka umí najít hnízdo včel, medojed ho zas „otevřít“) Slepí garnáti (Alphaeus) a ryby z čeledi Gobiidae (chodby pro rybu, doprovod pro garnáta) Ryby-klauni (Amphiprion) a sasanky (ochrana ryby a ochrana sasanky) Mutualismus – cílené pěstování rostlin nebo zvířat Člověk a kulturní plodiny/domácí zvířata Mravenci a medovici produkující hmyz Pěstování hub brouky a mravenci (brouci skupiny Scolytidae tunelují dřevo, aby tam rostly houby, mravenci Atta a Acromyrmex nosí listí na kompost, kde pěstují houby) Mutualismus při rozšiřování semen nebo opylovaní bobule a jiné dužnaté plody pojídané ptáky – semena se šíří (obecně málo specialistů, ale jsou) Rostliny využívající opylovače jim na oplátku nabízejí nektar (ten jinak nemá pro rostlinu žádnou cenu). Mezi opylovači jak generalisté, tak specialisté. Specializace nejvyšší u orchidejí, naopak příkladem generalisty třeba ostružiník Rubus fruticosus. Opylovači mohou rozšiřovat „pohlavní“ nemoci rostlin.
 Ryby-čističi a jejich „zákazníci (čističi se živí ektoparazity, a hostitelé se jich zbavují) Medojed a medojedka (medojedka umí najít hnízdo včel, medojed ho zas „otevřít ) Slepí garnáti (Alphaeus) a ryby z čeledi Gobiidae (chodby pro rybu, doprovod pro garnáta) Ryby-klauni (Amphiprion) a sasanky (ochrana ryby a ochrana sasanky) Mutualismus – cílené pěstování rostlin nebo zvířat. Člověk a kulturní plodiny/domácí zvířata Mravenci a medovici produkující hmyz Pěstování hub brouky a mravenci (brouci skupiny Scolytidae tunelují dřevo, aby tam rostly houby, mravenci Atta a Acromyrmex nosí listí na kompost, kde pěstují houby) Mutualismus při rozšiřování semen nebo opylovaní. bobule a jiné dužnaté plody pojídané ptáky – semena se šíří (obecně málo specialistů, ale jsou) Rostliny využívající opylovače jim na oplátku nabízejí nektar (ten jinak nemá pro rostlinu žádnou cenu). Mezi opylovači jak generalisté, tak specialisté. Specializace nejvyšší u orchidejí, naopak příkladem generalisty třeba ostružiník Rubus fruticosus. Opylovači mohou rozšiřovat „pohlavní nemoci rostlin.")
17
Mutualismus – typy vztahů
Mutualismus organismů v trávicích traktech Většina předchozích vztahů byla víceméně volnější, zahrnovala (v nějaké podobě) nějaké vyhledávání a fungovala většinou na principu odměny ve formě potravy. Jednalo se o vyvinuté, komplexní organismy, které tráví přinejmenším část života zcela samostatně. V řadě jiných případu mutualismu se jedná naopak o víceméně pevnou vazbu mezi jednobuněčným eukaryotem/prokaryotem a tělem partnera – extracelulární i intracelurání. Pro mnoho živočichů je přítomnost mikroflóry v trávicím traktu zásadní podmínkou umožňující trávit celulózu, a možná též pro syntézy vitaminů. Trávení pomocí mikroflóry je vypracovaný a řízený proces – a velmi se podobá kontinuální fermentaci, jakou známe např. z pivovaru. Klasika – bachor přežvýkavců: první rozmělnění redukuje velikost potravy na částice o objemu 1 – 1000 l, ovšem jen částice pod 5 l jdou do dalších částí žaludku, větší jsou vyvrženy do huby a znovu přežvýkány. Mikroorganismy tvoří až 20% obejmu bachoru, jsou specializované na určité látky, mezi ty hlavní patří Bacteriodes succinogenes (celulosa), Ruminococcus (celulosa, celobiosa, xylosa), z dalších Clostridium locheadii (celulosa, i škrob). Z prvoků Holotricha a Entodinomorpha, z nich jen málo umí celulosu, ale žerou třeba bakterie, nebo jiné prvoky = je to komplexní ekosystém. Produktem bachoru jsou nenasycené mastné kyseliny (ty jsou to hlavní pro přežvýkavce), amoniak, C02 a metan. V dalších částech žaludku už přežvýkavec tráví pomocí svých vlastních enzymů. Vzájemné výhody: mikroorganismy mají zajištěn neustálý přísun potravy a stabilní podmínky, přežvýkavec zase stravitelnou potravu. Termiti – tři skupiny: 1. mikroorganismy v trávicí trubici nebo 2. vlastní celulása nebo 3. pěstování hub. Termiti tzv. refekují, tj. znovu požírají výkaly (poprvé se mikrobi namnoží, podruhé jsou sežraní).
 nějaké vyhledávání a fungovala většinou na principu odměny ve formě potravy. Jednalo se o vyvinuté, komplexní organismy, které tráví přinejmenším část života zcela samostatně. V řadě jiných případu mutualismu se jedná naopak o víceméně pevnou vazbu mezi jednobuněčným eukaryotem/prokaryotem a tělem partnera – extracelulární i intracelurání. Pro mnoho živočichů je přítomnost mikroflóry v trávicím traktu zásadní podmínkou umožňující trávit celulózu, a možná též pro syntézy vitaminů. Trávení pomocí mikroflóry je vypracovaný a řízený proces – a velmi se podobá kontinuální fermentaci, jakou známe např. z pivovaru. Klasika – bachor přežvýkavců: první rozmělnění redukuje velikost potravy na částice o objemu 1 – 1000 l, ovšem jen částice pod 5 l jdou do dalších částí žaludku, větší jsou vyvrženy do huby a znovu přežvýkány. Mikroorganismy tvoří až 20% obejmu bachoru, jsou specializované na určité látky, mezi ty hlavní patří Bacteriodes succinogenes (celulosa), Ruminococcus (celulosa, celobiosa, xylosa), z dalších Clostridium locheadii (celulosa, i škrob). Z prvoků Holotricha a Entodinomorpha, z nich jen málo umí celulosu, ale žerou třeba bakterie, nebo jiné prvoky = je to komplexní ekosystém. Produktem bachoru jsou nenasycené mastné kyseliny (ty jsou to hlavní pro přežvýkavce), amoniak, C02 a metan. V dalších částech žaludku už přežvýkavec tráví pomocí svých vlastních enzymů. Vzájemné výhody: mikroorganismy mají zajištěn neustálý přísun potravy a stabilní podmínky, přežvýkavec zase stravitelnou potravu. Termiti – tři skupiny: 1. mikroorganismy v trávicí trubici nebo 2. vlastní celulása nebo 3. pěstování hub. Termiti tzv. refekují, tj. znovu požírají výkaly (poprvé se mikrobi namnoží, podruhé jsou sežraní).")
18
Mutualismus – typy vztahů
Mutualismus organismů v trávicích traktech Termiti – tři skupiny: 1. mikroorganismy v trávicí trubici nebo 2. vlastní celulása nebo 3. pěstování hub. Termiti tzv. refekují, tj. znovu požírají výkaly (poprvé se mikrobi namnoží, podruhé jsou sežraní). Z eukaryot je tam primitivní bičíkovec Trichomonas termopsidis, z bakterií dva typy mutualistů: 1. spirochety, hlavně na povrchu bičíkovců (pohyb bičíkovců?) 2. některé bakterie jsou schopny fixovat vzdušný dusík – Citrobacter freundii a Enterobacter (! jediný známý případ u hmyzu !), Mutualismus – organismy uvnitř živočišných tkání nebo buněk bakterie ve hmyzích mycetocytech (švábi, rovnokřídlí, mšice, brouci..) – snad pomáhají syntetizovat esenciální aminokyseliny. Mutualismus – vyšší rostliny a houby existuje mnoho vztahů rostlina – houba, ale ne všechny nutně mutualistické. Ale na druhou stranu – většina vyšších rostlin nemá kořeny, ale mykorhizu – těsné spojení rostlinných tkání a pletiv hub (jen málo čeledí rostlin ne, např. Cruciferae). Formy mykorhizy: ektomykorhiza (hlavně stromy, kořeny v vrchním humózním horizontu, obaluje povrch kořenů, hyfy do pletiv kořenů MEZI buňky, potřebuje karbohydráty, dodává minerální živiny, hodně malých spor). Vesikulárně arbuskularní (VA) (hyfy dovnitř buněk kořenů, málo velkých spor, zisk pro rostlinu = více fosforu z půdy, nebo obrana před patogeny, pro houbu = karbohydráty od rostliny) Rod Endogone, Glomus. Při nízkém P spíš parazit? Další typy: třeba u některých orchidejí, které nemají chlorofyl, zprostředkovává houba transport asimilátů z jiných fotosyntetizujících rostlin. V mnoha takových případech je však těžké rozhodnout, zda vlastně nejde o parasitismus.
. Z eukaryot je tam primitivní bičíkovec Trichomonas termopsidis, z bakterií dva typy mutualistů: 1. spirochety, hlavně na povrchu bičíkovců (pohyb bičíkovců ) 2. některé bakterie jsou schopny fixovat vzdušný dusík – Citrobacter freundii a Enterobacter (! jediný známý případ u hmyzu !), Mutualismus – organismy uvnitř živočišných tkání nebo buněk. bakterie ve hmyzích mycetocytech (švábi, rovnokřídlí, mšice, brouci..) – snad pomáhají syntetizovat esenciální aminokyseliny. Mutualismus – vyšší rostliny a houby. existuje mnoho vztahů rostlina – houba, ale ne všechny nutně mutualistické. Ale na druhou stranu – většina vyšších rostlin nemá kořeny, ale mykorhizu – těsné spojení rostlinných tkání a pletiv hub (jen málo čeledí rostlin ne, např. Cruciferae). Formy mykorhizy: ektomykorhiza (hlavně stromy, kořeny v vrchním humózním horizontu, obaluje povrch kořenů, hyfy do pletiv kořenů MEZI buňky, potřebuje karbohydráty, dodává minerální živiny, hodně malých spor). Vesikulárně arbuskularní (VA) (hyfy dovnitř buněk kořenů, málo velkých spor, zisk pro rostlinu = více fosforu z půdy, nebo obrana před patogeny, pro houbu = karbohydráty od rostliny) Rod Endogone, Glomus. Při nízkém P spíš parazit Další typy: třeba u některých orchidejí, které nemají chlorofyl, zprostředkovává houba transport asimilátů z jiných fotosyntetizujících rostlin. V mnoha takových případech je však těžké rozhodnout, zda vlastně nejde o parasitismus.")
19
Mutualismus – typy vztahů - mykorhiza
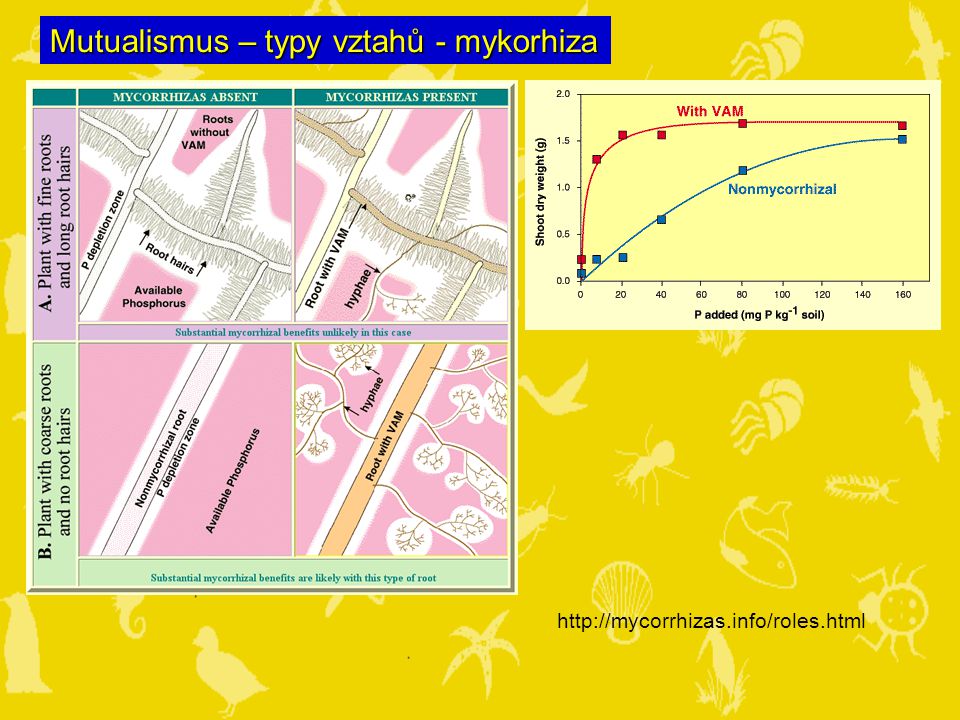
20
Mutualismus – typy vztahů

21
Mutualismus – typy vztahů - mykorhiza
A. Benefits to plants Increased plant nutrient supply by extending the volume of soil accessible to plants as explained below. Increased plant nutrient supply by acquiring nutrient forms that would not normally be available to plants (Tarafdar & Marschner 1994, Schweiger et al. 1995, Kahiluoto & Vestberg 1998). Some ECM and ericoid fungi have the capacity to breakdown phenolic compounds in soils which can interfere with nutrient uptake (Bending & Read 1997). Root colonisation by ECM and VAM fungi can provide protection from parasitic fungi and nematodes (Duchesne et al. 1989, Grandmaison et al. 1993, Newsham et al. 1995, Little & Maun 1996, Cordier et al. 1998, Morin et al. 1999). Non-nutritional benefits to plants due to changes in water relations, phytohormone levels, carbon assimilation, etc. have been reported, but are difficult to interpret (Brundrett 1991, Smith & Read 1997). Mycorrhizal benefits can include greater yield, nutrient accumulation, and/or reproductive success (Lewis & Koide 1990, Stanley et al. 1993). Mycorrhizas can cause growth form changes to root architecture, vascular tissue, etc. (Daniels Hetrick et al. 1988, Miller et al. 1997). Suppression of competing non-host plants, by mycorrhizal fungi has been observed (Allen et al. 1989). Significant amounts of carbon transfer through fungus mycelia connecting different plant species has been measured (Simard et al. 1997). This could reduce competition between plants and contribute to the stability and diversity of ecosystems. Networks of hyphae supported by dominant trees may help seedlings become established or contribute to the growth of shaded understorey plants (Hogberg et al. 1999, Horton et al. 1999). Nutrient transfer from dead to living plants may occur (Eason et al. 1991).
. Some ECM and ericoid fungi have the capacity to breakdown phenolic compounds in soils which can interfere with nutrient uptake (Bending & Read 1997). Root colonisation by ECM and VAM fungi can provide protection from parasitic fungi and nematodes (Duchesne et al. 1989, Grandmaison et al. 1993, Newsham et al. 1995, Little & Maun 1996, Cordier et al. 1998, Morin et al. 1999). Non-nutritional benefits to plants due to changes in water relations, phytohormone levels, carbon assimilation, etc. have been reported, but are difficult to interpret (Brundrett 1991, Smith & Read 1997). Mycorrhizal benefits can include greater yield, nutrient accumulation, and/or reproductive success (Lewis & Koide 1990, Stanley et al. 1993). Mycorrhizas can cause growth form changes to root architecture, vascular tissue, etc. (Daniels Hetrick et al. 1988, Miller et al. 1997). Suppression of competing non-host plants, by mycorrhizal fungi has been observed (Allen et al. 1989). Significant amounts of carbon transfer through fungus mycelia connecting different plant species has been measured (Simard et al. 1997). This could reduce competition between plants and contribute to the stability and diversity of ecosystems. Networks of hyphae supported by dominant trees may help seedlings become established or contribute to the growth of shaded understorey plants (Hogberg et al. 1999, Horton et al. 1999). Nutrient transfer from dead to living plants may occur (Eason et al. 1991).")
22
Mutualismus – typy vztahů - mykorhiza
B. Other roles in ecosystems Soil hyphae are likely to have an important role in nutrient cycling by helping to prevent losses from the system, especially at times when roots are inactive (Lussenhop & Fogel 1999). Hyphae are conduits that may transport carbon from plant roots to other soil organisms involved in nutrient cycling processes. Thus, cooperating with other members of the decomposition soil food-web. Soil hyphae may have an important role in nutrient cycling by acquiring nutrients from saprophytic fungi (Lindahl et al. 1999). Epigeous and hypogeous sporocarps of ECM and VAM fungi are important food sources for placental and marsupial mammals (McGee & Baczocha 1994, Janos et al. 1995, Reddell et al. 1997, Mcilwee & Johnson 1998, Claridge 2002). Mycorrhizal roots and fungus fruit bodies are important as food sources and habitats for invertebrates (Fogel & Peck 1975, Rabatin & Stinner 1989, Lawrence & Milner 1996). Mycorrhizal fungus hyphae are an important food source for soil invertibrates (Setala 1995, Ingham & Massicotte 1994). Mycorrhizas influence soil microbial populations and exudates in the mycorrhizosphere and hyphosphere (Ames et al. 1984, Bansal & Mukerji 1994, Olsson et al. 1996, Andrade et al. 1998). Hyphae of VAM fungi are considered to contribute to soil structure. Their role in mechanical aggregation has been questioned (Degens et al. 1994), but secretions such as glomalin may be more important (Wright & Upadhyaya 1998). Hyphal mats produced by ECM fungi considerably alter soil structure (Griffiths et al. 1994). Mycorrhizal fungi contribute to carbon storage in soil by altering the quality and quantity of soil organic matter (Ryglewicz & Andersen 1994).
. Hyphae are conduits that may transport carbon from plant roots to other soil organisms involved in nutrient cycling processes. Thus, cooperating with other members of the decomposition soil food-web. Soil hyphae may have an important role in nutrient cycling by acquiring nutrients from saprophytic fungi (Lindahl et al. 1999). Epigeous and hypogeous sporocarps of ECM and VAM fungi are important food sources for placental and marsupial mammals (McGee & Baczocha 1994, Janos et al. 1995, Reddell et al. 1997, Mcilwee & Johnson 1998, Claridge 2002). Mycorrhizal roots and fungus fruit bodies are important as food sources and habitats for invertebrates (Fogel & Peck 1975, Rabatin & Stinner 1989, Lawrence & Milner 1996). Mycorrhizal fungus hyphae are an important food source for soil invertibrates (Setala 1995, Ingham & Massicotte 1994). Mycorrhizas influence soil microbial populations and exudates in the mycorrhizosphere and hyphosphere (Ames et al. 1984, Bansal & Mukerji 1994, Olsson et al. 1996, Andrade et al. 1998). Hyphae of VAM fungi are considered to contribute to soil structure. Their role in mechanical aggregation has been questioned (Degens et al. 1994), but secretions such as glomalin may be more important (Wright & Upadhyaya 1998). Hyphal mats produced by ECM fungi considerably alter soil structure (Griffiths et al. 1994). Mycorrhizal fungi contribute to carbon storage in soil by altering the quality and quantity of soil organic matter (Ryglewicz & Andersen 1994).")
23
Mutualismus – typy vztahů
Fotosyntetizující a chemosyntetizující symbionti bezobratlých Řasy nacházíme v tkáních různých organismů, nejvíce u láčkovců (nezmar Hydra a řasa Chlorella – až buněk na jedince, v trávicích buňkách nezmara). Nezmar má karbohydráty a kyslík, řasy stabilní a bezpečné prostředí. V mořích nejčastěji dinoflagelát Symbiodinium v sasankách, korálech, medúzách apod. Kromě „primární produkce“ vysrážení CaCO3 = podstatná komponenta těl korálů. Též v mořském prvokovi Mesodinum rubrum symbiotické řasy, v jeho případě může dojít až ke „kvetení vody“, při něm je primární produkce až 2 g m-3 h-1, tj. jedna z největších mezi mikroorganismy. Chloroplasty ze strávených řas (Siphonales) fungují v plžích až několik měsíců – zdroj kyslíku! Lišejníky houba + řasa. Řasy tvoří jen 3-10% hmotnosti lišejníku. Z počtu asi známých druhů hub asi 25% lichenizuje.Houba má asimiláty, u řas to není tak jasné. Možná jen komfortnější podmínky k růstu. Při lichenizaci se dramaticky mění morfologie a růst houby – pokud roste volně, vypadá jak houba, pokud s řasou, tak tvoří úplně jinak vypadající, ale zase typický tvar – to vede až k taxonomické klasifikaci lišejníků. 90% lišejníků má zelené řasy, proto nemohou vázat vzdušný dusík. Zbylých 10% má sinice, takže mohou. Ale kupodivu nerostou na dusík chudých místech !?
. Nezmar má karbohydráty a kyslík, řasy stabilní a bezpečné prostředí. V mořích nejčastěji dinoflagelát Symbiodinium v sasankách, korálech, medúzách apod. Kromě „primární produkce vysrážení CaCO3 = podstatná komponenta těl korálů. Též v mořském prvokovi Mesodinum rubrum symbiotické řasy, v jeho případě může dojít až ke „kvetení vody , při něm je primární produkce až 2 g m-3 h-1, tj. jedna z největších mezi mikroorganismy. Chloroplasty ze strávených řas (Siphonales) fungují v plžích až několik měsíců – zdroj kyslíku! Lišejníky. houba + řasa. Řasy tvoří jen 3-10% hmotnosti lišejníku. Z počtu asi známých druhů hub asi 25% lichenizuje.Houba má asimiláty, u řas to není tak jasné. Možná jen komfortnější podmínky k růstu. Při lichenizaci se dramaticky mění morfologie a růst houby – pokud roste volně, vypadá jak houba, pokud s řasou, tak tvoří úplně jinak vypadající, ale zase typický tvar – to vede až k taxonomické klasifikaci lišejníků. 90% lišejníků má zelené řasy, proto nemohou vázat vzdušný dusík. Zbylých 10% má sinice, takže mohou. Ale kupodivu nerostou na dusík chudých místech !")
24
Život při vysokých teplotách
Pozoruhodný typ „horkého“ stanoviště byl objeven nedávno (1979) - jedná se o vývěry vroucí vody z hlubokomořských tektonických švů, které vytváří z vysrážených minerálů „komíny“. Jde o hloubky m, kde se díky vysokému tlaku vaří voda až při 370 – 400 °C. Při vývěru z „komínu“ se voda prudce ochlazuje, protože okolí má 2 °C – vzniká tak obrovský gradient přechodných teplot. Bakterie odtud odebrané rostou při 110 °C, avšak jsou nepřímé důkazy, že přímo v komínech žijí ještě daleko odolnější bakterie. V okolí komínů žije v teplíčku mnoho dalších organismů (55 taxonů), jejichž společenstvo je založeno na přísunu energie od lithotrofních H2S využívajících bakterií, přičemž některé jsou i endosymbionti (např. v žábrách zdejších červů, gastropodů a škeblí).
 - jedná se o vývěry vroucí vody z hlubokomořských tektonických švů, které vytváří z vysrážených minerálů „komíny . Jde o hloubky m, kde se díky vysokému tlaku vaří voda až při 370 – 400 °C. Při vývěru z „komínu se voda prudce ochlazuje, protože okolí má 2 °C – vzniká tak obrovský gradient přechodných teplot. Bakterie odtud odebrané rostou při 110 °C, avšak jsou nepřímé důkazy, že přímo v komínech žijí ještě daleko odolnější bakterie. V okolí komínů žije v teplíčku mnoho dalších organismů (55 taxonů), jejichž společenstvo je založeno na přísunu energie od lithotrofních H2S využívajících bakterií, přičemž některé jsou i endosymbionti (např. v žábrách zdejších červů, gastropodů a škeblí).")
25
Calyptogena Riftia pachyptila (Vestimentifera - Pogonophora)
")
26
Mutualismus – typy vztahů
Fixace vzdušného dusíku Neschopnost vázat vzdušný dusík většinou organismů je další evoluční hádankou. Umí to jen některá prokaryota, a z nich je řada pevně vázána v mutualistických vztazích s různými eukaryoty. Ti, co známe ze symbiosy, patří k Azotobacteriaceae (Azotobacter, Azotococcus), vážou v aerobním prostředí, jsou běžně na povrchu listů a kořenů Rhizobia (Rhizobium, Bradirhizobium, Azorhizobium), v nodech kořenů Fabaceae (a jilmu). Umějí vázat v anaerobiose. Bacillaceae (Clostridium, Desulfotoaculum), v trávicím traktu savců (přežvýkavců) Enterobacteriaceae (Enterobacter, Citrobacter), v termitech, ale i na kořenech Spirillaceae (Spirillium), obligátní aerob na kořenech trav Actinomycetes – rod Frankia, na kořenech nebobovitých rostlin (třeba olše) Sinice čeledi Nostocaceae, s mnoha různými rostlinami. Nejlíp studováno u Fabaceae Olše a Frankia
, vážou v aerobním prostředí, jsou běžně na povrchu listů a kořenů. Rhizobia (Rhizobium, Bradirhizobium, Azorhizobium), v nodech kořenů Fabaceae (a jilmu). Umějí vázat v anaerobiose. Bacillaceae (Clostridium, Desulfotoaculum), v trávicím traktu savců (přežvýkavců) Enterobacteriaceae (Enterobacter, Citrobacter), v termitech, ale i na kořenech. Spirillaceae (Spirillium), obligátní aerob na kořenech trav. Actinomycetes – rod Frankia, na kořenech nebobovitých rostlin (třeba olše) Sinice čeledi Nostocaceae, s mnoha různými rostlinami. Nejlíp studováno u Fabaceae. Olše a Frankia.")
27
Fixace vzdušného dusíku
Nejlíp studováno u Fabaceae Bakterie volně v půdě, exudáty z kořenů je stimulují k růstu, a také „zapnou“ geny, jež jsou odpovědné za indukci nodulace-tvorby rhizobií na kořenech hostitele. Po vytvoření nodu (hlízky) jsou cévním systémem přiváděny asimiláty, a odváděny dusíkaté látky, zejména asparagin. Nitrogenáza tvoří až 40% všech bílkovin v hlízkách a její aktivita je limitována nízkou tenzí kyslíku, proto je jeho přísun bráněn tlustou stěnou hlízky. V hlízkách se navíc tvoří hemoglobin (leghemoglobin), hlízky mají díky němu růžovou barvu. Váže volný kyslík a jeho pomocí jsou zajištěny anaerobní podmínky pro nitrogenázy. Hodnocení výhod a ztrát mykorhizy je složité, je nutno počítat s alternativními zdroji dusíku. Tvorba a udržování hlízek stojí rostlinu až 12% fotosyntetické aktivity. Ale má se za to, že řada zelených rostlin patří mezi „patologické nadvýrobce karbohydrátů“, proto jsou pro ně náklady ve formě asimilátů vlastně nepodstatné, velmi levné. Rostliny fixující dusík jsou svým způsobem „lokální sebevrazi“. Fixací dusíku zlepšují podmínky pro jiné rostliny, se kterými potom prohrávají v kompetici.
 jsou cévním systémem přiváděny asimiláty, a odváděny dusíkaté látky, zejména asparagin. Nitrogenáza tvoří až 40% všech bílkovin v hlízkách a její aktivita je limitována nízkou tenzí kyslíku, proto je jeho přísun bráněn tlustou stěnou hlízky. V hlízkách se navíc tvoří hemoglobin (leghemoglobin), hlízky mají díky němu růžovou barvu. Váže volný kyslík a jeho pomocí jsou zajištěny anaerobní podmínky pro nitrogenázy. Hodnocení výhod a ztrát mykorhizy je složité, je nutno počítat s alternativními zdroji dusíku. Tvorba a udržování hlízek stojí rostlinu až 12% fotosyntetické aktivity. Ale má se za to, že řada zelených rostlin patří mezi „patologické nadvýrobce karbohydrátů , proto jsou pro ně náklady ve formě asimilátů vlastně nepodstatné, velmi levné. Rostliny fixující dusík jsou svým způsobem „lokální sebevrazi . Fixací dusíku zlepšují podmínky pro jiné rostliny, se kterými potom prohrávají v kompetici.")
28
Modelovní mutualistických vztahů
Na první pohled, proč ne jako Lotka-Volterra pro dravce, ale s opačným znaménkem u působení druhého druhu. Ovšem takový model vede k explozivnímu růstu obou druhů do nekonečné velikosti. Musí se zahrnout nějaké limitující faktory-nosné kapacity (ovšem odjinud). Pro určitý typ mutualismu modely založené na rozložení aktivity mezi hledání a zpracování (viz Hollingova rovnice), ale univerzální model se stále hledá.
. Pro určitý typ mutualismu modely založené na rozložení aktivity mezi hledání a zpracování (viz Hollingova rovnice), ale univerzální model se stále hledá.")
29
Mutualismus – obecné shrnutí
Symbiotičtí mutualisté mají obvykle velmi jednoduchý životní cyklus (narozdíl od parazitů) Sexuální rozmnožování je u endosymbiontů obvykle potlačeno (pokud srovnáváme s parazity či volně žijícími příbuznými) Endosymbionti nemají zřetelnou dispersi (snad jen nějaké houby) Dá se čekat, že díky koevoluci se budou mutualisté šířit pospolu. To platí jen někde (mravenčí královna si odnáší inokulum houby do nového mraveniště, lišejníky), jinde ne – např. mykorhitické bakterie. Populace mutualistů jsou stabilní, neznáme epidemie (jako u parazitů). Počet endosymbiontů na hostitele je velmi stálý, musí existovat nějaké hustotně závislé regulace. Ekologická nika druhů v mutualistickém vztahu je vždy širší než když žijí tyto druhy samostatně (opět opak od parazitů)
 Sexuální rozmnožování je u endosymbiontů obvykle potlačeno (pokud srovnáváme s parazity či volně žijícími příbuznými) Endosymbionti nemají zřetelnou dispersi (snad jen nějaké houby) Dá se čekat, že díky koevoluci se budou mutualisté šířit pospolu. To platí jen někde (mravenčí královna si odnáší inokulum houby do nového mraveniště, lišejníky), jinde ne – např. mykorhitické bakterie. Populace mutualistů jsou stabilní, neznáme epidemie (jako u parazitů). Počet endosymbiontů na hostitele je velmi stálý, musí existovat nějaké hustotně závislé regulace. Ekologická nika druhů v mutualistickém vztahu je vždy širší než když žijí tyto druhy samostatně (opět opak od parazitů)")
Podobné prezentace



















