Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Biosyntéza aminokyselin
1. Fixace dusíku 2. Tvorba aminokyselin z meziproduktů citrátového cyklu a dalších hlavních metabolických drah 3. Regulace biosyntézy aminokyselin zpětnou vazbou 4. Aminokyseliny jako prekurzory dalších biomolekul

2
Fixace dusíku (redukce N2 na NH3)
Schopnost fixovat dusík mají bakterie, vyšší organismy ne !! Anorganický průmysl, výroba NH3 – Fritz Haber (1910). Vodík, katalyzátory (Fe), 500oC a tlak 300 atm. Trojná vazba dusíku je velmi rezistentní vůči chemickým reakcím (225 kcal/mol)
. Vodík, katalyzátory (Fe), 500oC a tlak 300 atm. Trojná vazba dusíku je velmi rezistentní vůči chemickým reakcím (225 kcal/mol)")
3
Nitrogenasový komplex: reduktasa a nitrogenasa. Účast ATP
Nitrogenasový komplex: reduktasa a nitrogenasa. Účast ATP. Komplex je extrémně citlivý na kyslík. Motýlokvěté rostliny chrání nitrogenasu v kořenových hlízách leghemoglobinem, který váže kyslík.

4
Redukce N2 na NH3 je šestielektronový proces.
Biologický proces obvykle vede ke tvorbě 1 molu H2 při tvorbě 2 molů NH3 z každého molu N2. V tom případě je nutný vstup osmi elektronů. N2 + 8 e- + 8 H+ = 2 NH3 + H2 Zdrojem elektronů je redukovaný ferredoxin (vzniká při fotosyntéze nebo při oxidacích). K transportu každého elektronu se hydrolyzují 2 ATP, tedy celkem 16 ATP na jednu molekulu N2.
. K transportu každého elektronu se hydrolyzují. 2 ATP, tedy celkem 16 ATP na jednu molekulu N2.")
5
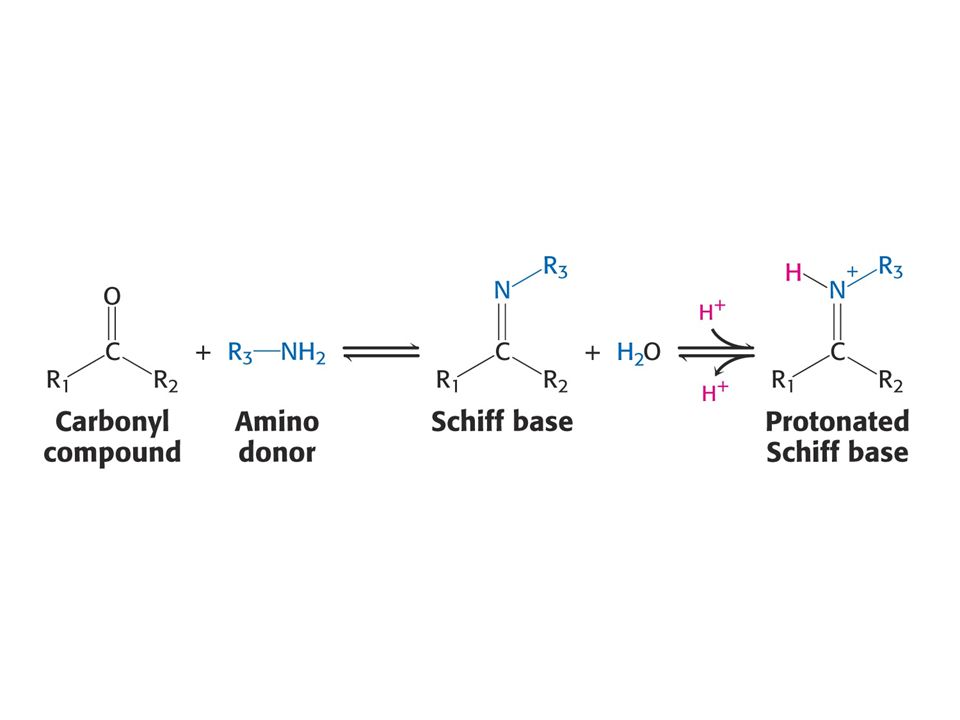
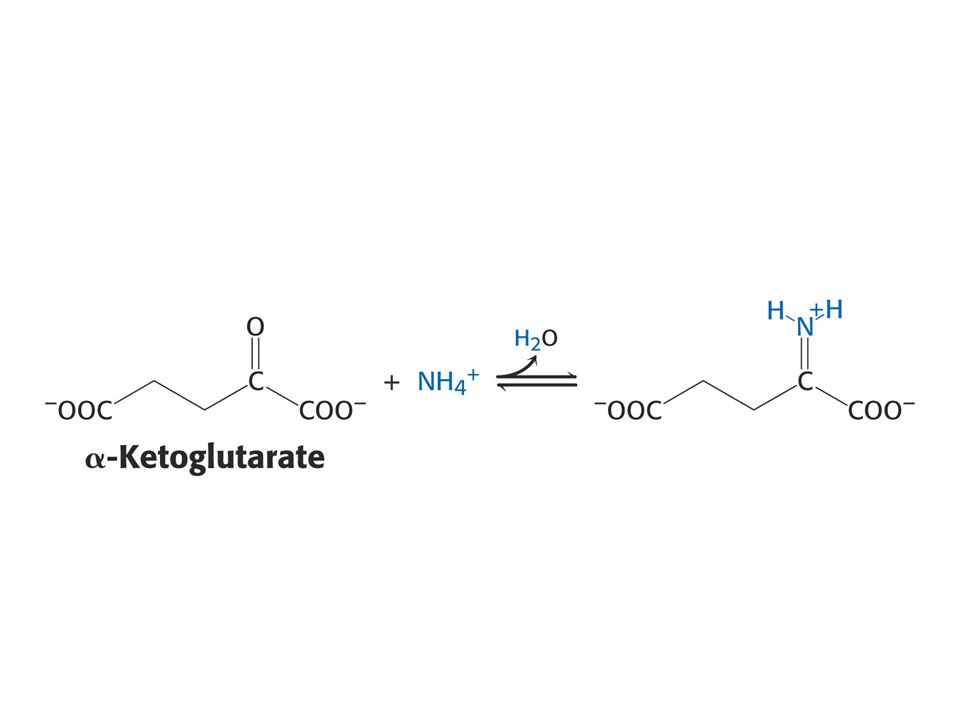
NH4+ je asimilován do aminokyselin přes Glu a Gln
Enzym: glutamátdehydrogenasa (nerozlišuje mezi NADH a NADPH). NADPH je redukční činidlo. NH4+ + a-oxoglutarát + NADPH + H+ = Glutamát + NADP+ + H2O Reakce probíhá ve dvou stupních: tvorba Schiffovy báze a potom po protonaci, redukce přenosem hydridového iontu z NADPH.
. NADPH je redukční činidlo. NH4+ + a-oxoglutarát + NADPH + H+ = Glutamát + NADP+ + H2O. Reakce probíhá ve dvou stupních: tvorba Schiffovy báze a potom po protonaci, redukce přenosem hydridového iontu z NADPH.")
9
Reakce glutamátdehydrogenasy je stereospecifická. Vzniká jen L-glutamát.

10
Glutaminsynthetasa. Vstup druhého NH4+ do Glu.

11
Glutamátdehydrogenasa a glutaminsynthetasa jsou přítomny ve všech organismech.
Většina prokaryot obsahuje enyzm glutamátsynthasu, která katalyzuje reduktivní aminaci a-oxoglutarátu za účasti Gln jako donoru dusíku: a-Oxoglutarát + Gln + NADPH + H+ = 2 Glu + NADP+ Za situace, kdy je nedostatek NH4+ probíhá následná reakce glutaminsynthetasy a glutamátsynthasy – sumárně: NH4+ + a-oxoglutarát + NADPH + ATP = Glu + NADP+ + ADP + Pi.

12
Stechiometrie této reakce se liší od glutamátdehydrogenasové
Stechiometrie této reakce se liší od glutamátdehydrogenasové. Při této je hydrolyzováno ATP. Proč prokaryonti někdy využívají tuto dražší alternativu ? KM glutamátdehydrogenasy pro NH4+ je vysoké (asi 1 mM) a enzym není saturován za situace nedostatku NH4+. Na rozdíl, glutaminsynthetasa má vysokou afinitu k NH4+. Proto je nutná hydrolýza ATP k zachycení nedostatkového NH4+.
 a enzym není saturován za situace nedostatku NH4+. Na rozdíl, glutaminsynthetasa má vysokou afinitu k NH4+. Proto je nutná hydrolýza ATP k zachycení nedostatkového NH4+.")
13
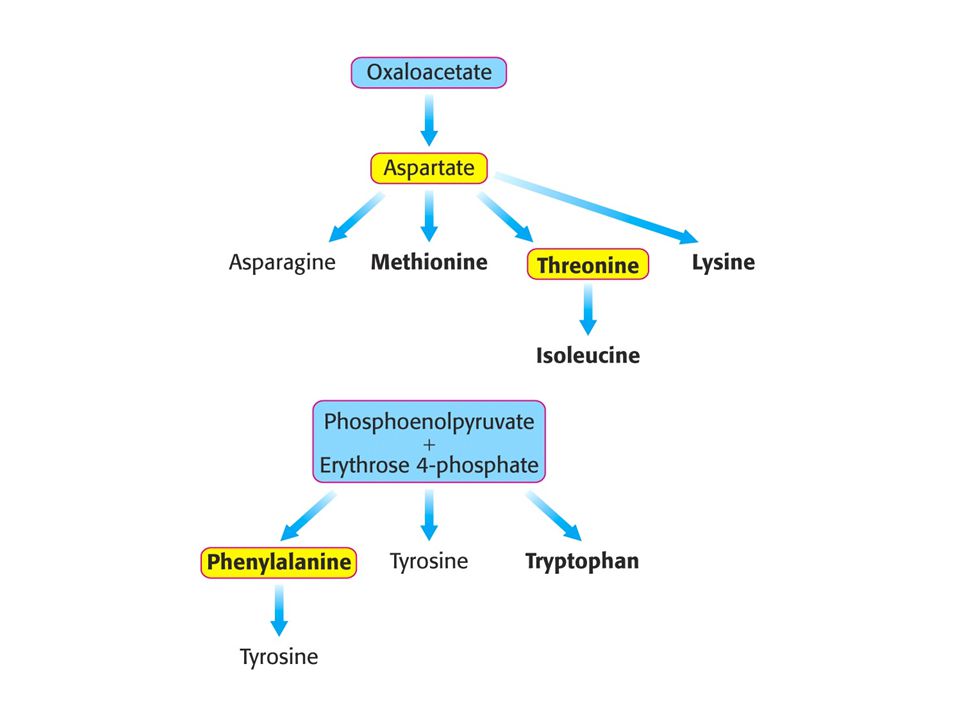
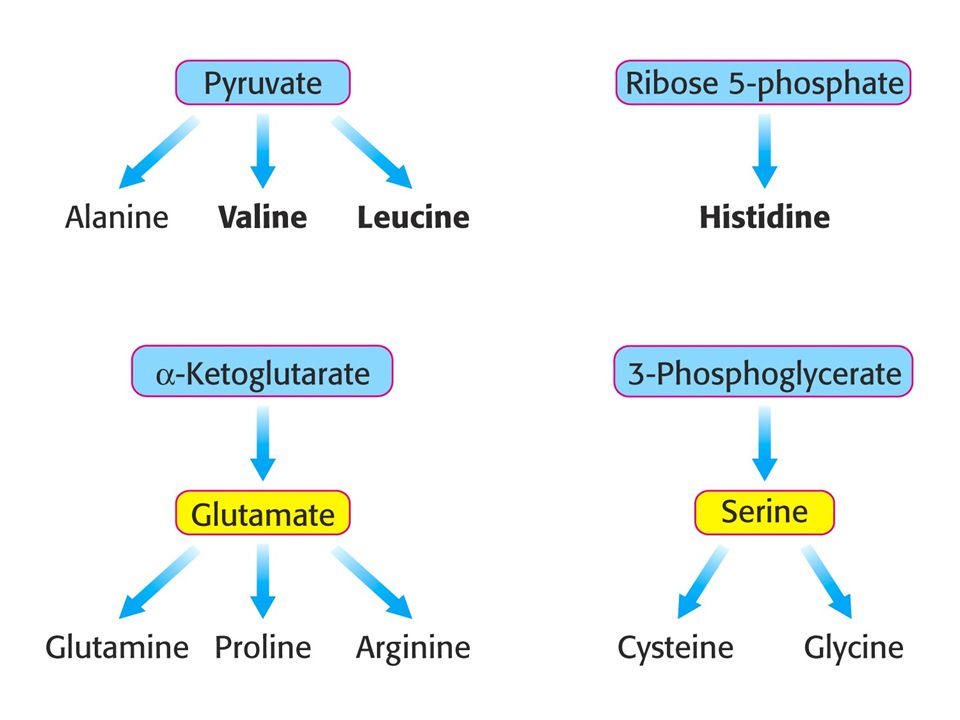
Tvorba aminokyselin z meziproduktů citrátového cyklu a dalších hlavních metabolických drah (glykolýzy a pentosafosfátové dráhy). Hlavní prekursory. Esenciální aminokyseliny tištěny tučně.

17
Společný krok určuje chiralitu všech aminokyselin.
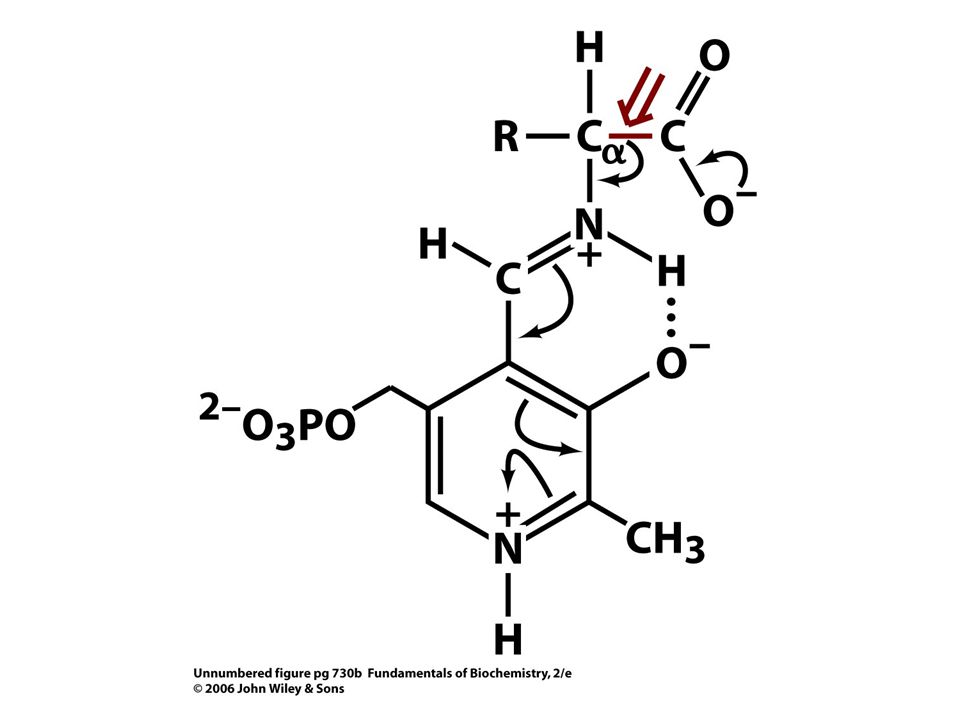
Tři a-ketokyseliny mohou být převedeny v jednom kroku na aminokyseliny (pyruvát, a-oxoglutarát a oxaloacetát. Katalyzováno pyridoxalfosfát dependentními transaminasami. Njdůležitějším krokem transaminace je protonizace chinoidního meziproduktu za vzniku externího aldiminu. Chiralita vznikající aminokyseliny je určována směrem ze kterého se proton připojuje na chinoidní formu.

18
Adenylátový meziprodukt je nutný pro tvorbu Asn z Asp.

19
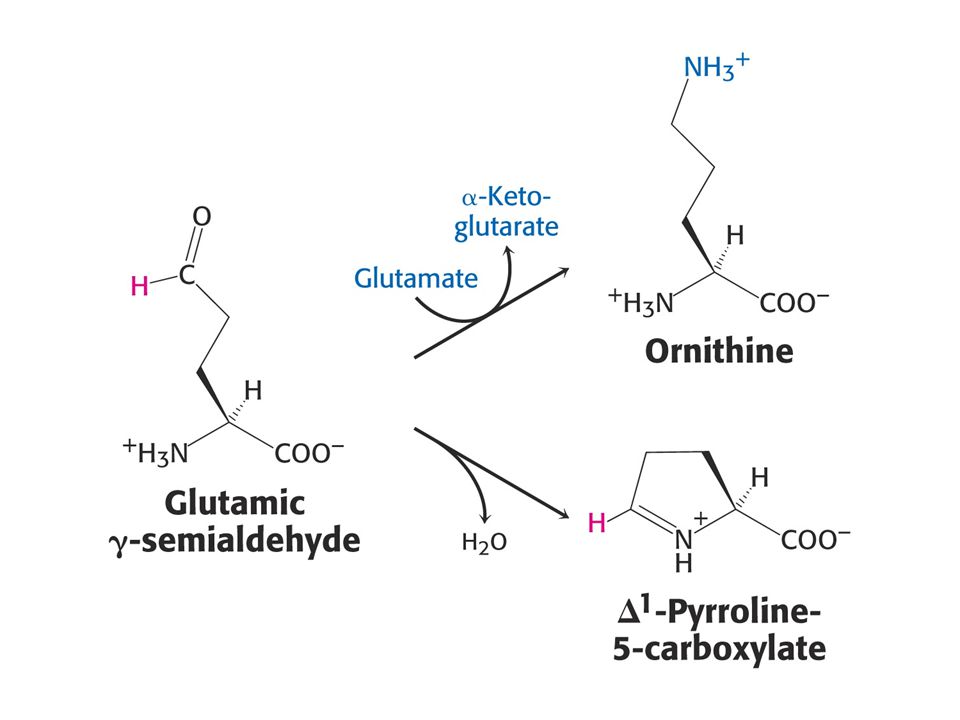
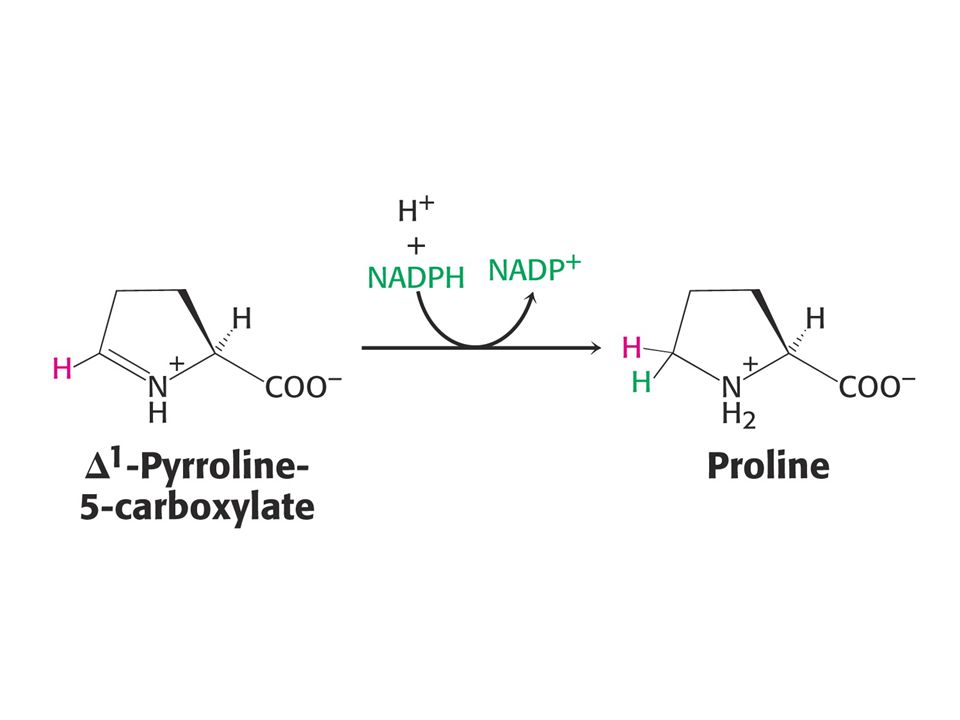
Glu je prekursorem Gln, Pro a Arg.

22
Ser, Cys a Gly se tvoří z 3-fosfoglycerátu.

23
TETRAHYDROFOLÁT –TF4 (také tetrahydropteroylglutamát) – je universální přenašeč jednouhlíkatých jednotek. Savci umí syntetizovat pteridinový skelet, ale nedokáží ho spojit se dvěma dalšími součástmi a proto ho získávají z potravy nebo z mikroorganismů střevní mikroflóry.

24
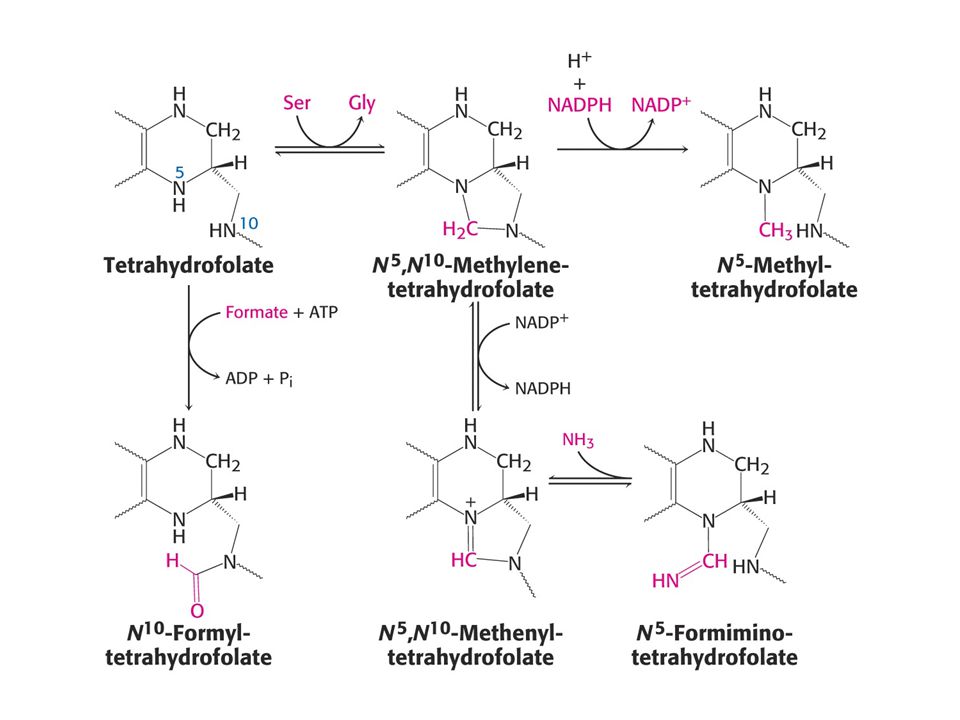
Jednouhlíkatá jednotka přenášená TF4 je vázána do poloh N-5 a N-10 (označeno N5 a N10 nebo do obou.
Jednotka může existovat ve třech oxidačních stavech: Redukovaná – CH3 Částečně oxidovaná - -CH2- (methylen) Více oxidovaná: formyl, formimino a methenyl. Plně oxidovaná je CO2 – přenáší biotin. Jednotlivé formy jsou vzájemně převoditelné. N10 –formyltetrahydrofolát může být také syntetizován z tetrahydrofolátu, mravenčanu a ATP.
 Více oxidovaná: formyl, formimino a methenyl. Plně oxidovaná je CO2 – přenáší biotin. Jednotlivé formy jsou vzájemně převoditelné. N10 –formyltetrahydrofolát může být také syntetizován z tetrahydrofolátu, mravenčanu a ATP.")
27
Využití derivátů tetrahydrofolátu:
Regenerace Met z homocysteinu přenosem methylu N5-methyltetrahydrofolátu. Biosyntéza purinových nukleotidů. Methylskupina Thyminu je z N5, N10-methylentetrahydrofolátu. Alternativní syntéza Gly (glycinsynthasa): CO2 + NH4+ + N5, N10-methylentetrahydrofolát + NADH = Gly + tetrahydrofolát + NAD+. TF4 je akceptorem jednouhlíkatých zbytků při degradacích. Hlavním zdrojem jednouhlíkatých skupin je snadná konverze Ser na Gly poskytující N5, N10-methylentetrahydrofolát.
: CO2 + NH4+ + N5, N10-methylentetrahydrofolát + NADH = Gly + tetrahydrofolát + NAD+. TF4 je akceptorem jednouhlíkatých zbytků při degradacích. Hlavním zdrojem jednouhlíkatých skupin je snadná konverze Ser na Gly poskytující N5, N10-methylentetrahydrofolát.")
30
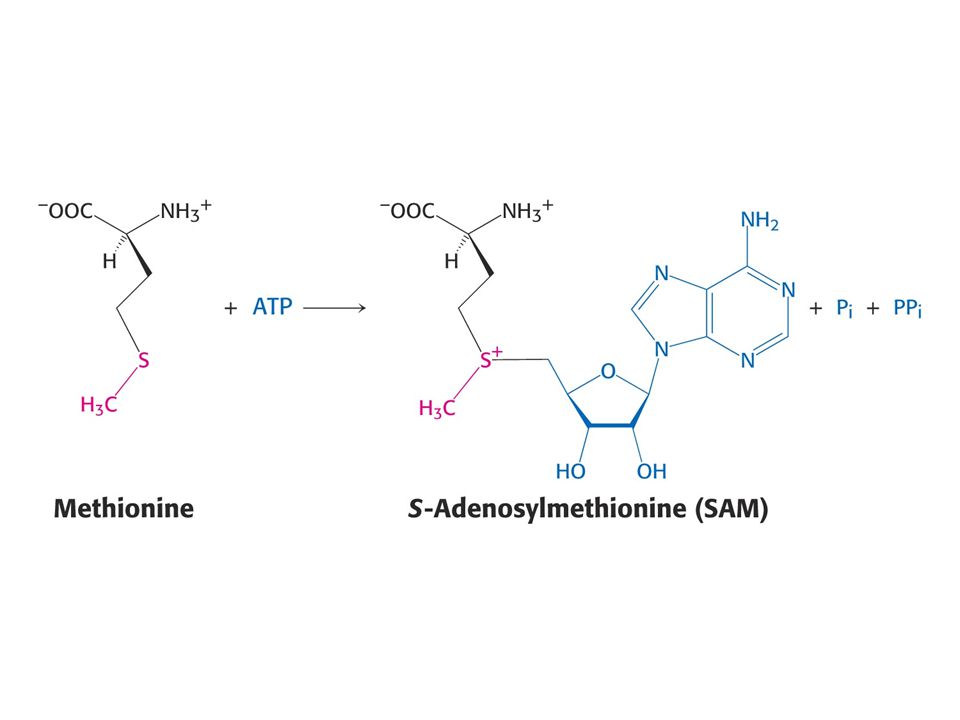
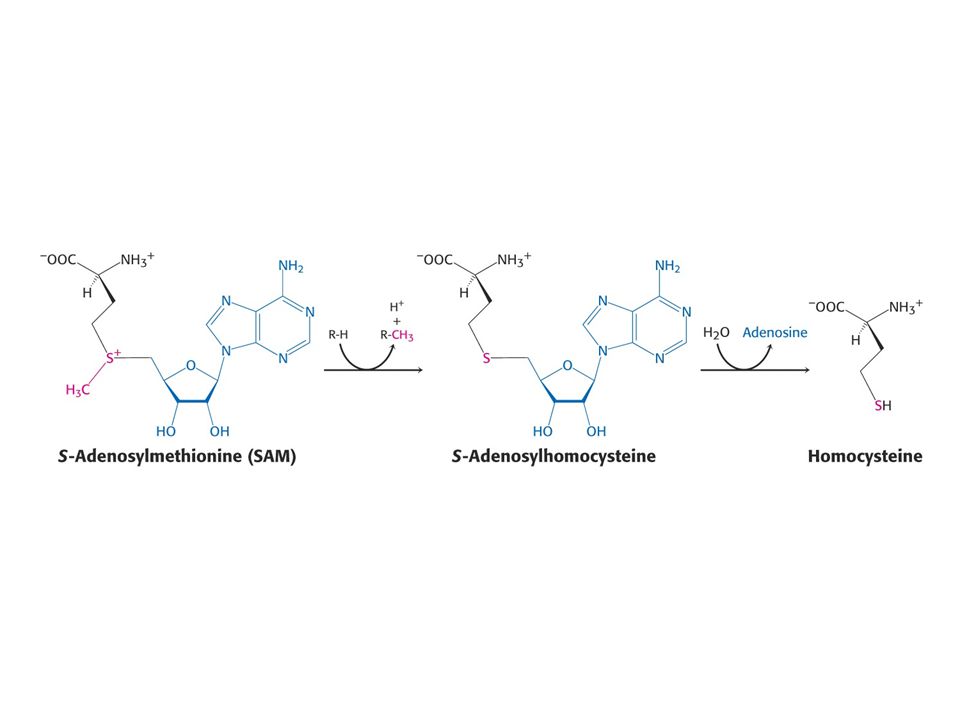
Methioninsynthasa, také homocysteinmethyltransferasa
Methioninsynthasa, také homocysteinmethyltransferasa. Koenzymem je methylkobalamin, prekurzorem vitamin B12

31
Aktivovaný cyklus methylu.

32
Methylace DNA.Methylace DNA chrání bakteriální DNA před štěpením restrikčními enzymy. Struktura DNAmethylasy

33
SAM jako prekursor ethylenu. Zrání plodů.

34
Cys je syntetizován ze Ser a homocysteinu
Cys je syntetizován ze Ser a homocysteinu. Enzym: cystathionin-b-synthasa. Atom síry Cys je z homocysteinu a uhlíkatý skelet ze Ser

35
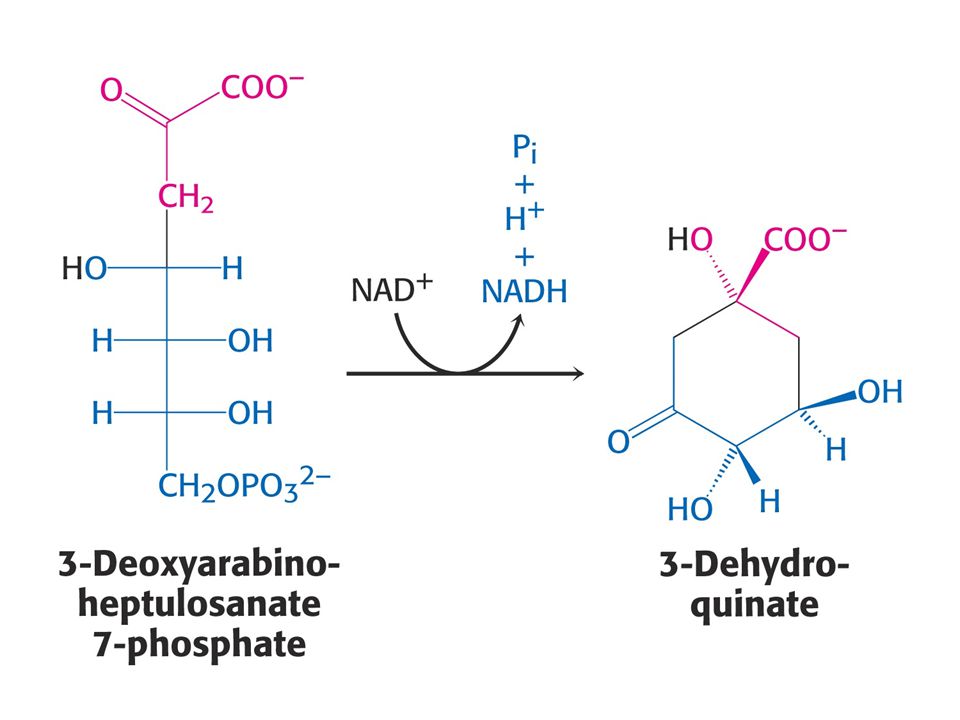
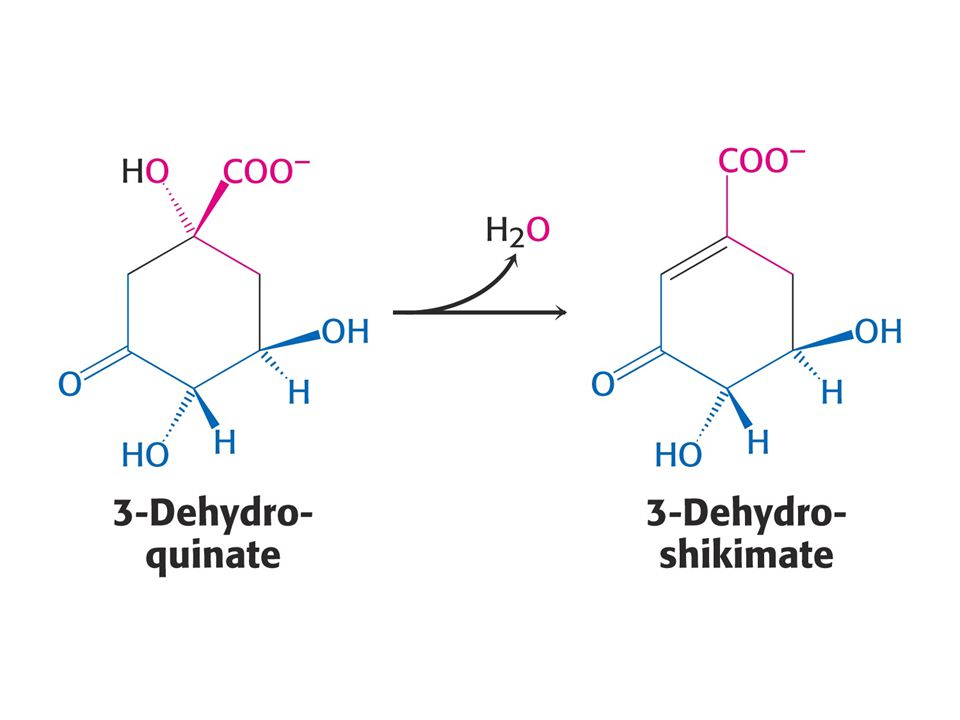
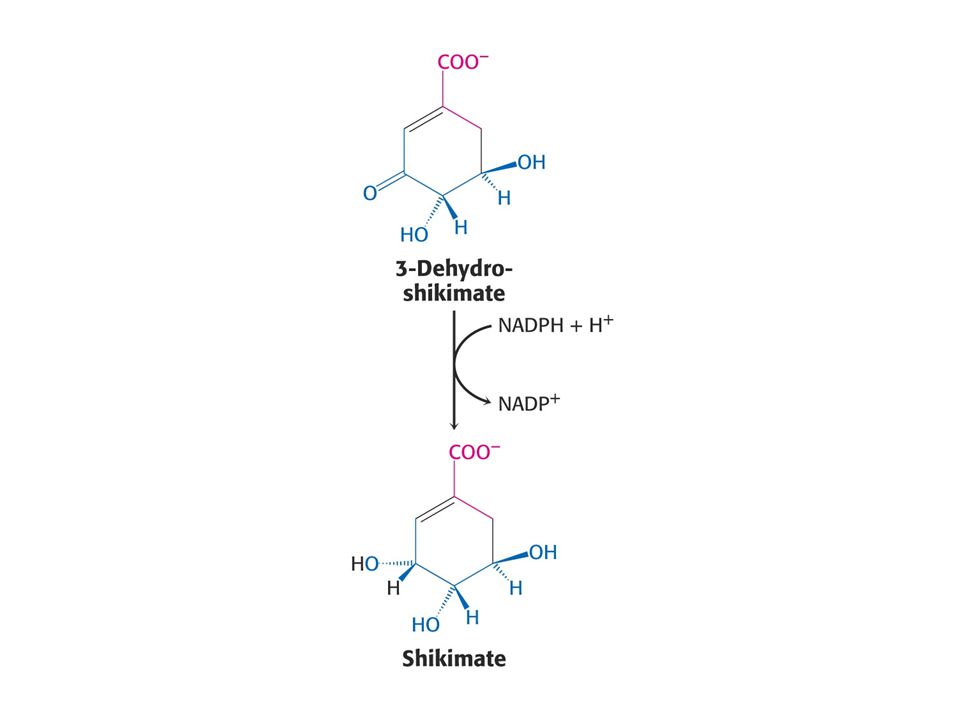
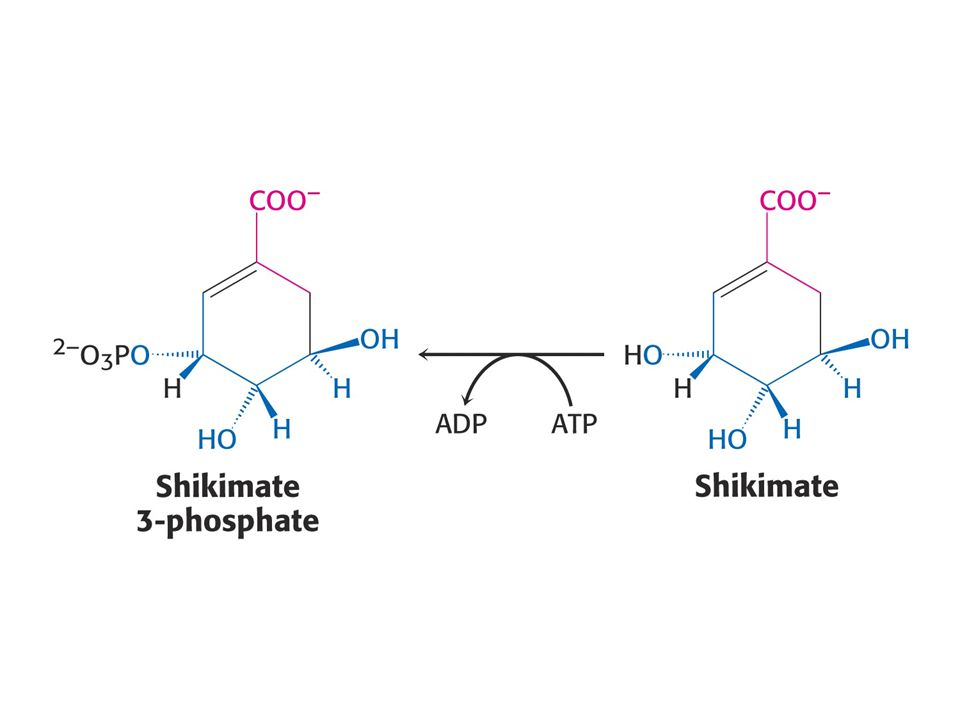
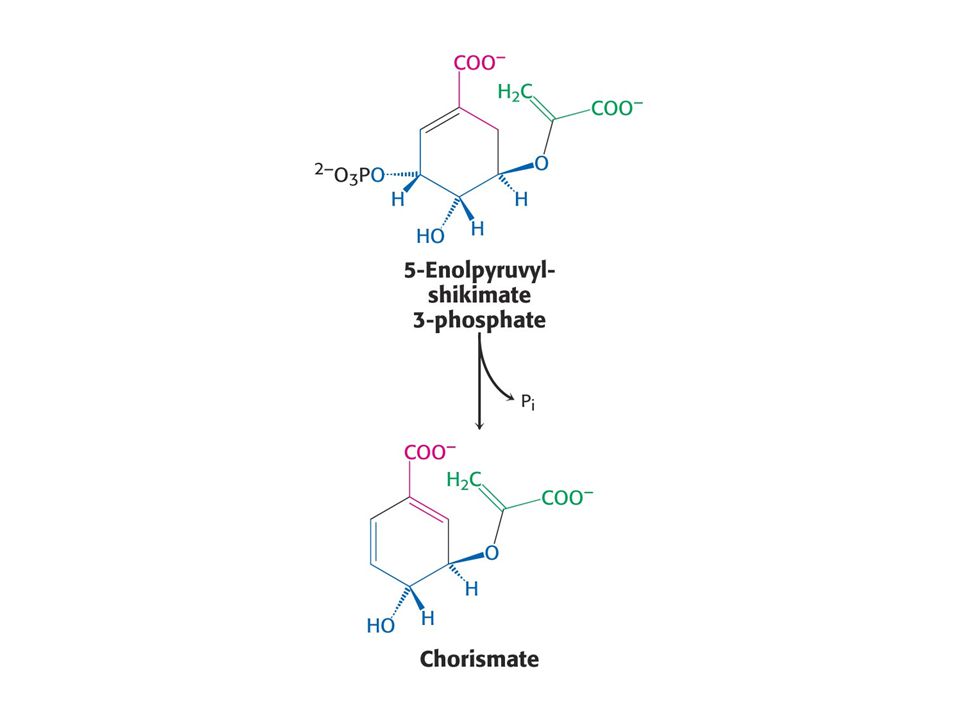
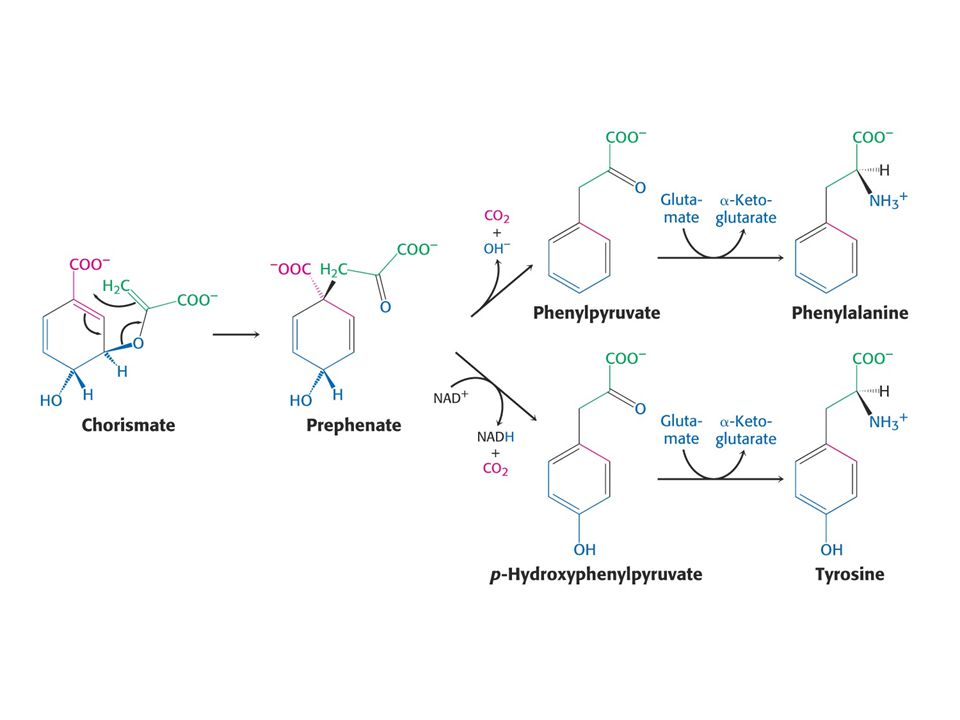
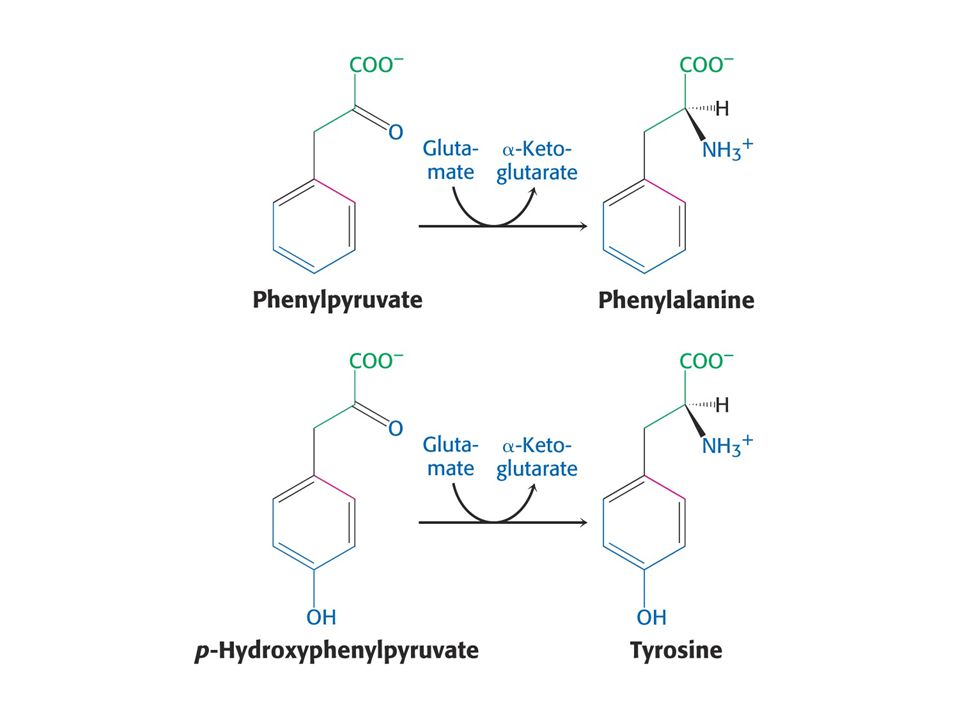
Biosyntéza aromatických aminokyselin – šikimátová dráha.
Prekurzory: 2 x fosfoenolpyruvát a 1 x erythrosa-4-fosfát Významné meziprodukty: Šikimát a chorismát.

41
Glyfosát (Raundup) inhibuje enzym tvorby 5-enoylpyruvylšikimát-3-fosfátu. Zvířata tento enzym nemají a proto je Raundup pro ně netoxický.

49
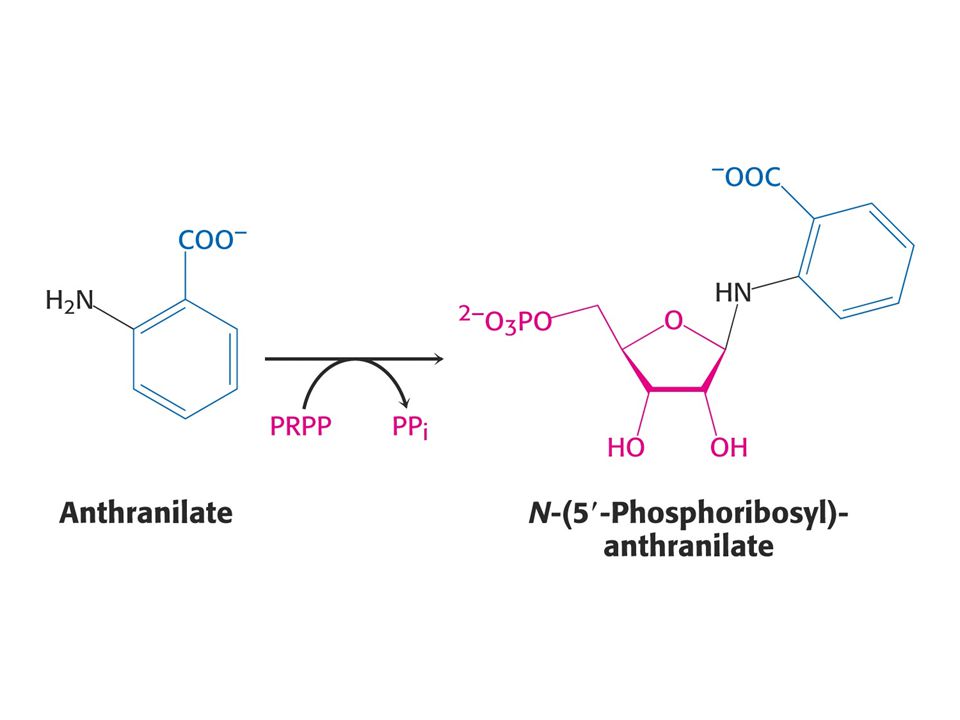
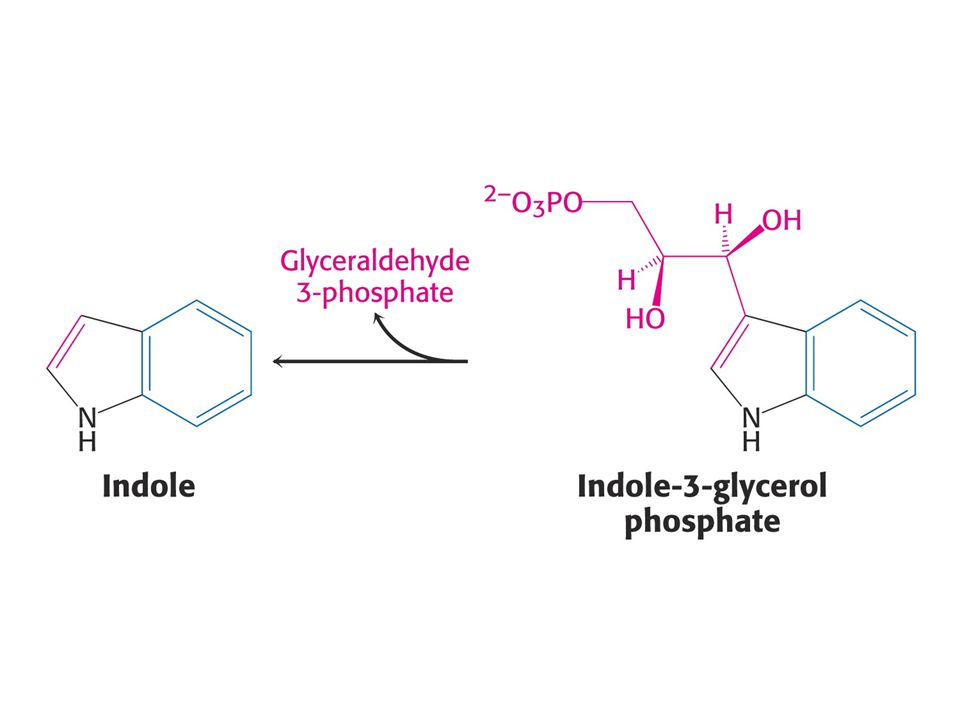
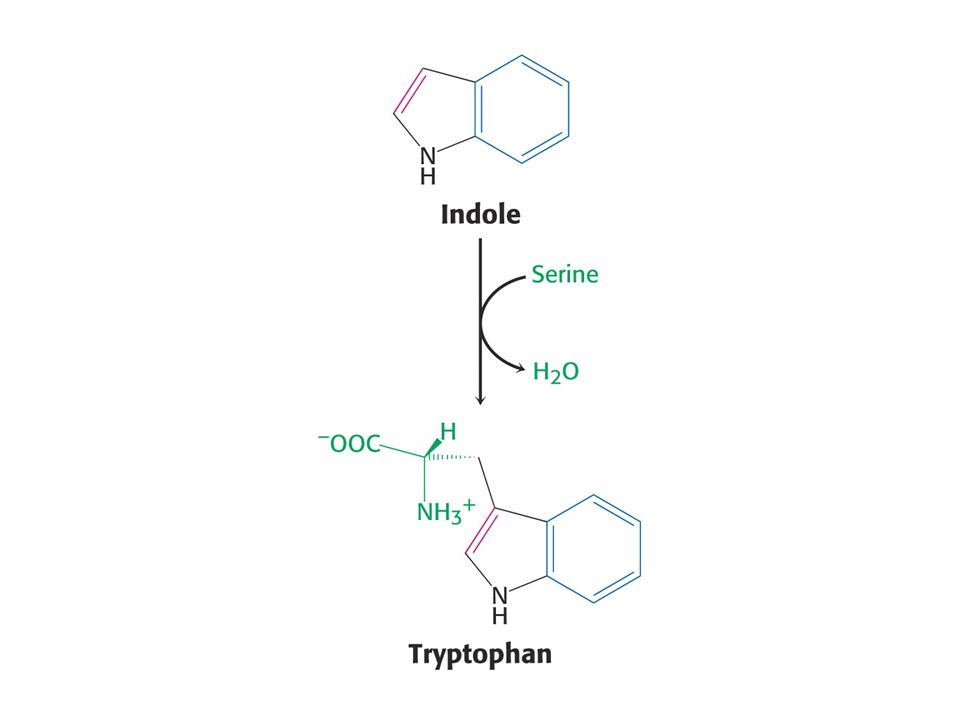
Anthranilát – odbočka na Trp

56
5-Fosforibosyl-1-pyrofosfát (PRPP).
Aktivovaná forma ribosafosfátu. Meziprodukt při syntéze His, purinových a pyrimidinových nukleotidů.

57
Biosyntéza aminokyselin je regulována zpětnovazebnou inhibicí.
Při biosyntetické dráze je obvykle první ireversibilní reakce nazvaná rozhodující stupeň („committed step“) důležitým regulačním místem. Konečný produkt je často inhibitorem enzymu této reakce. Např. při biosyntéze Ser je tímto stupněm oxidace 3-fosfoglycerátu za katalýzy 3-fosfoglycerátdehydrogenasy. U E. coli je enzym homotetramer mající aktivní místo a vazebné místo pro Ser. Po vazbě Ser do vazebných míst se snižuje hodnota KM pro substrát. Dostatek Ser vede k zablokování jeho syntézy a 3-fosfoglycerát může být využit pro jiné procesy. Nehromadí se další meziprodukty.
 důležitým regulačním místem. Konečný produkt je často inhibitorem enzymu této reakce. Např. při biosyntéze Ser je tímto stupněm oxidace. 3-fosfoglycerátu za katalýzy 3-fosfoglycerátdehydrogenasy. U E. coli je enzym homotetramer mající aktivní místo. a vazebné místo pro Ser. Po vazbě Ser do vazebných míst. se snižuje hodnota KM pro substrát. Dostatek Ser vede k zablokování jeho syntézy. a 3-fosfoglycerát může být využit pro jiné procesy. Nehromadí se další meziprodukty.")
58
Aminokyseliny jako prekursory mnoha biomolekul
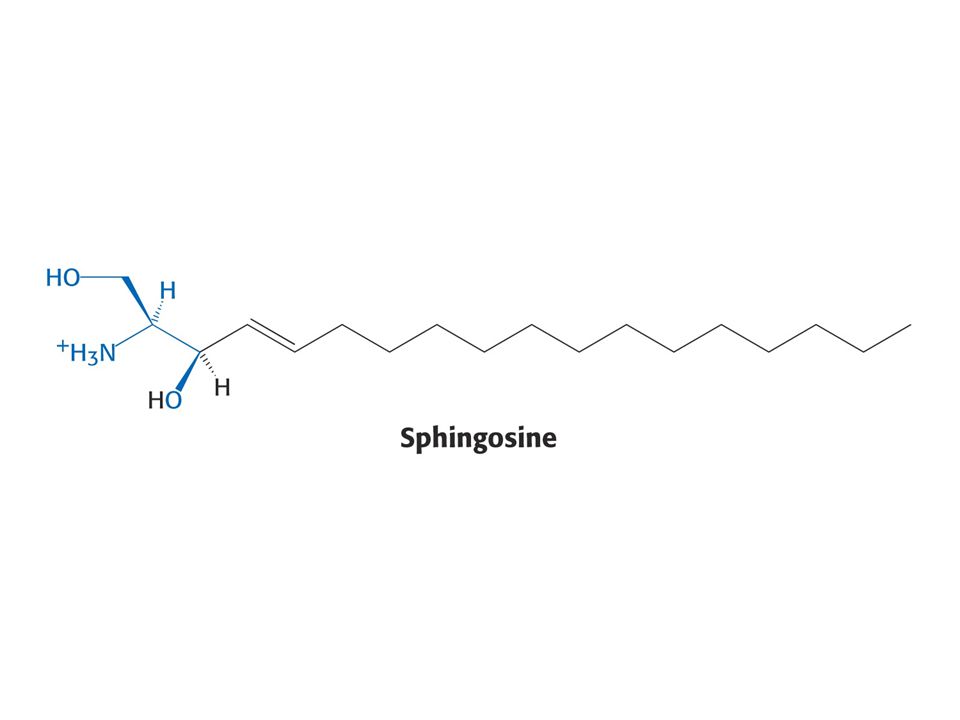
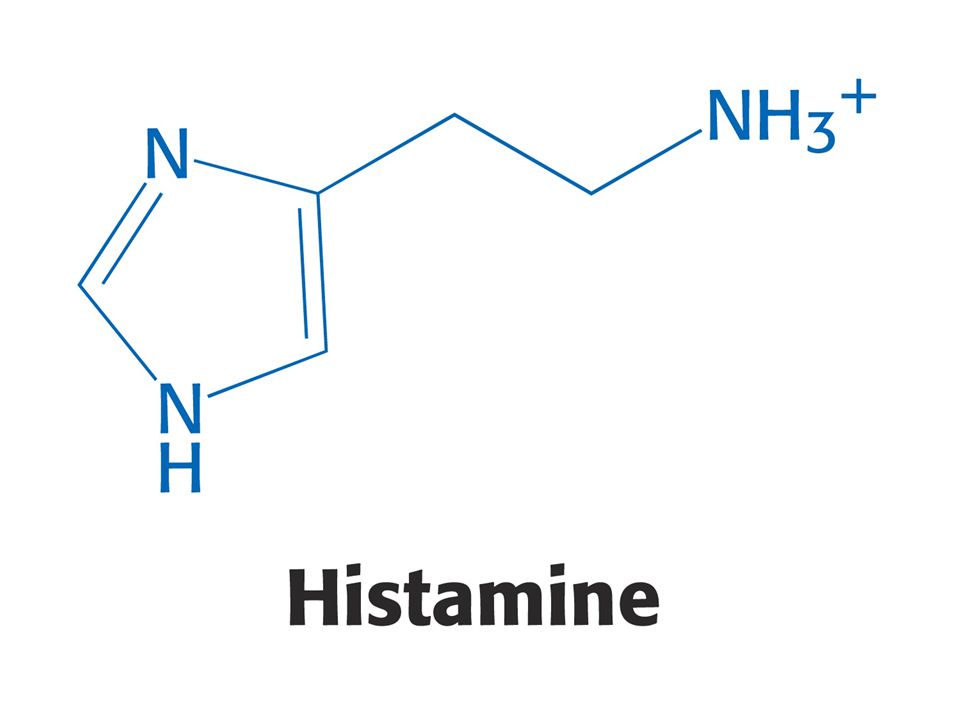
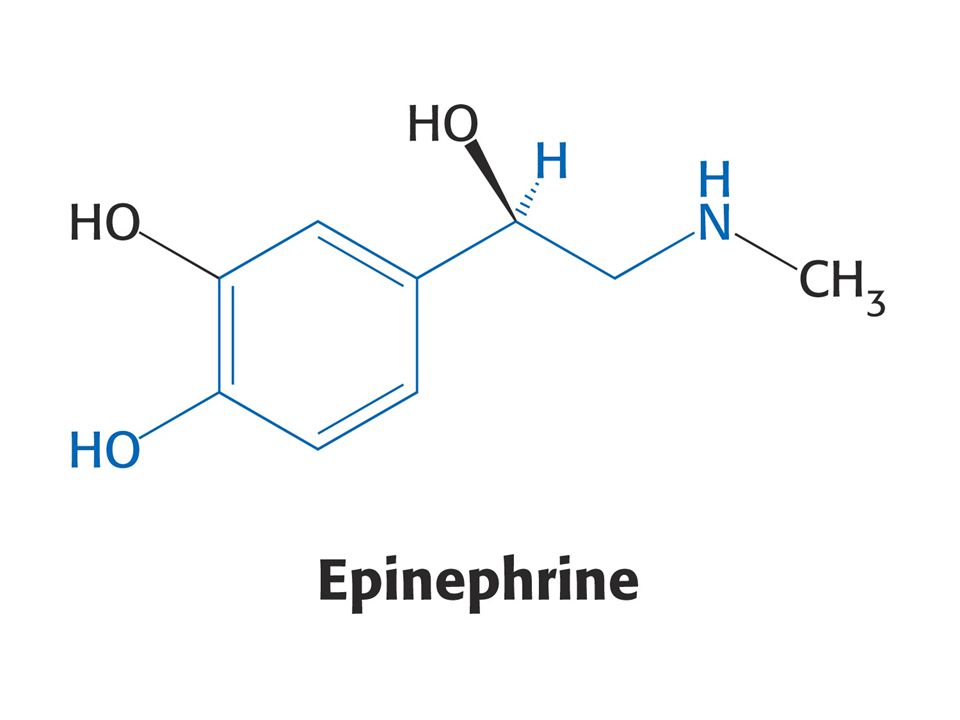
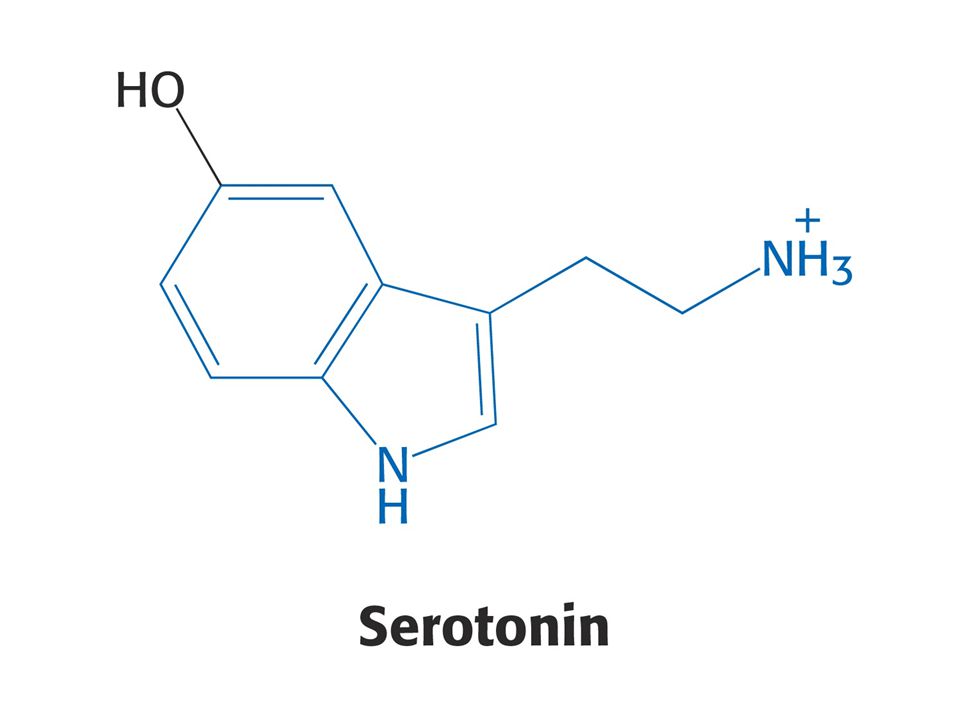
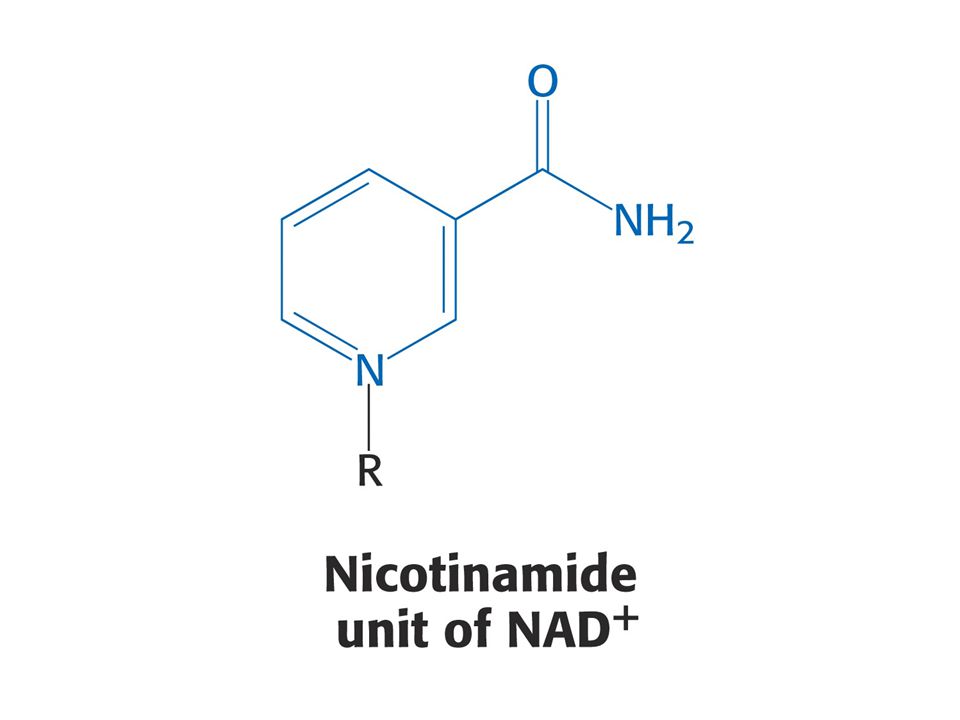
Puriny a pyrimidiny - z různých aminokyselin. Reaktivní konec sfingosinu, meziprodukt biosyntézy sfingolipidů, pochází ze Ser. Histamin, vasodilatans, z His. Prekurzorem hormonů thyroxinu, adrenalinu a komplexu polymerního pigmentu melaninu je Tyr. Neurotransmiter serotonin a nikotiamidový kruh NAD+ jsou syntetizovány z Trp.

68
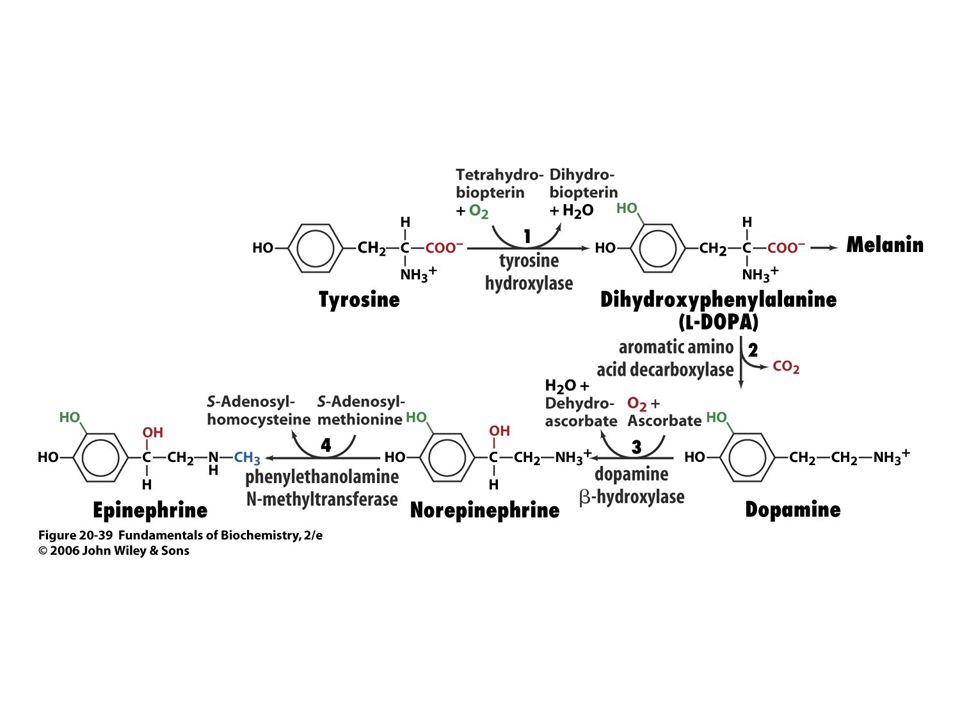
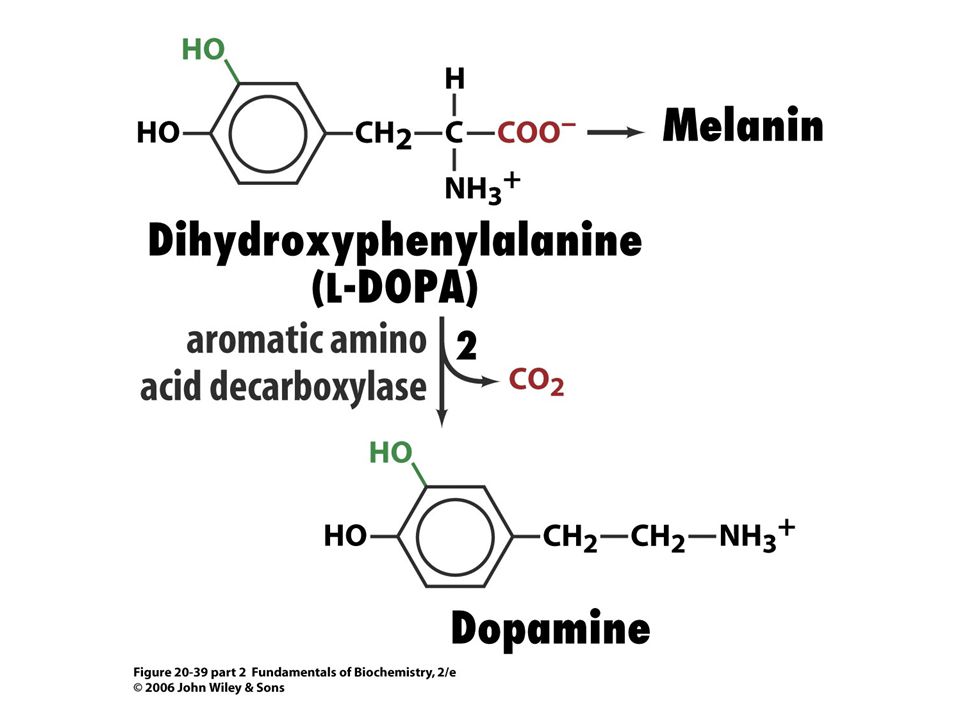
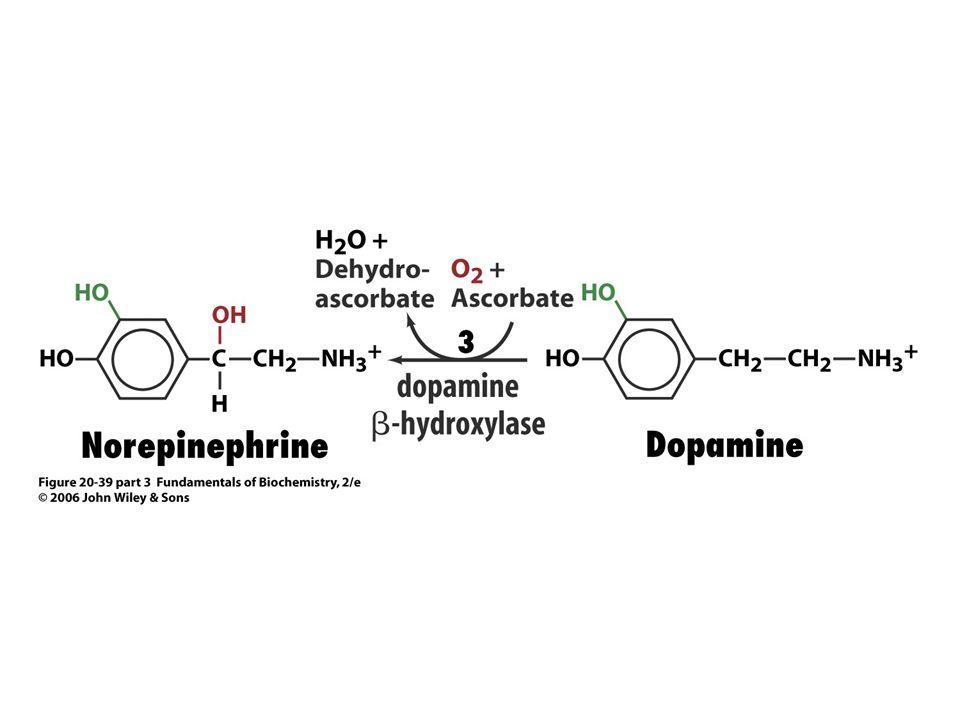
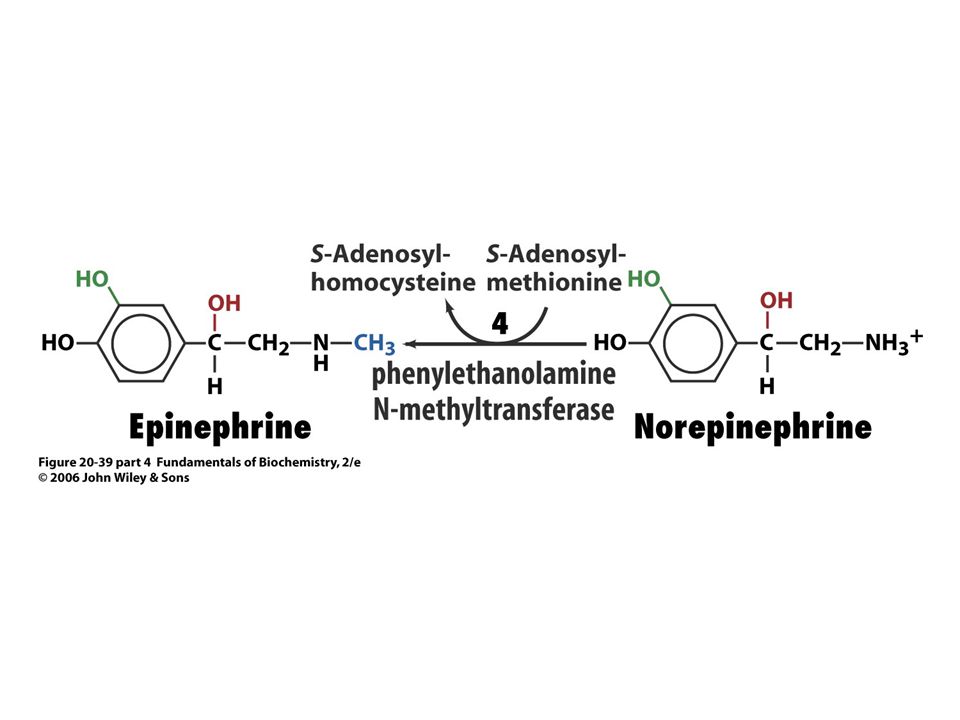
Biosyntéza katecholaminů.

76
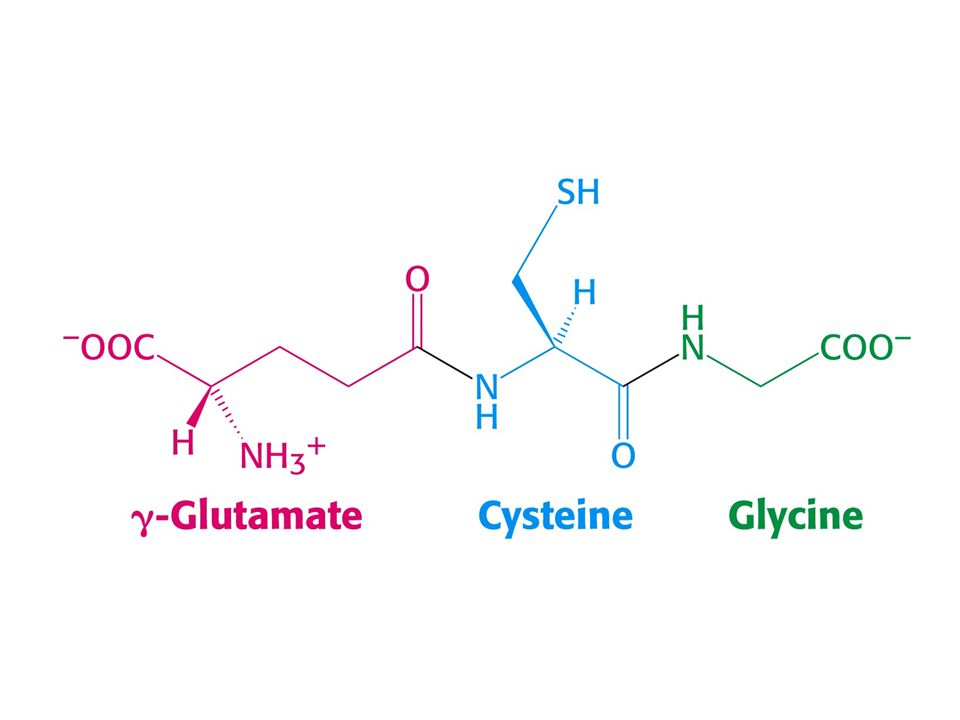
Glutathion je zásobou SH (sulfhydrylu) a antioxidant.
Koncentrace glutathionu v živočišných buňkách je 5 mM. Chrání červené krvinky před oxidačním poškozením. Glutathion cykluje mezi oxidovanou GSSG a redukovanou GSH formou. Je redukován za katalýzy glutathionreduktasy (flavoprotein využívající jako zdroj elektronů NADPH). Poměr GSH/GSSG je ve většině buněk větší než 500. Glutathion má významnou roli při detoxifikaci. Reguje s peroxidem vodíku a organickými peroxidy. 2 GSH + RO-OH = GSSG + H2O + ROH. Katalyzuje glutathionperoxidasa
. Poměr GSH/GSSG je ve většině buněk větší než 500. Glutathion má významnou roli při detoxifikaci. Reguje s peroxidem vodíku a organickými peroxidy. 2 GSH + RO-OH = GSSG + H2O + ROH. Katalyzuje glutathionperoxidasa.")
77
Struktura glutathionperoxidasy.

78
Glutathionperoxidasa obsahuje aminokyselinu SELENOCYSTEIN
Glutathionperoxidasa obsahuje aminokyselinu SELENOCYSTEIN v aktivním místě. Selenolát E-Se- redukuje peroxid a oxiduje se na kyselinu. Poté vstupuje do reakce GSH a vytváří selenosulfidový adukt (E-Se-S-G). Druhá molekula GSH poté regeneruje enzym reakcí se selenosulfidem za tvorby oxidovaného glutathionu GSSG.
. Druhá molekula GSH poté regeneruje enzym reakcí se selenosulfidem za tvorby oxidovaného glutathionu GSSG.")
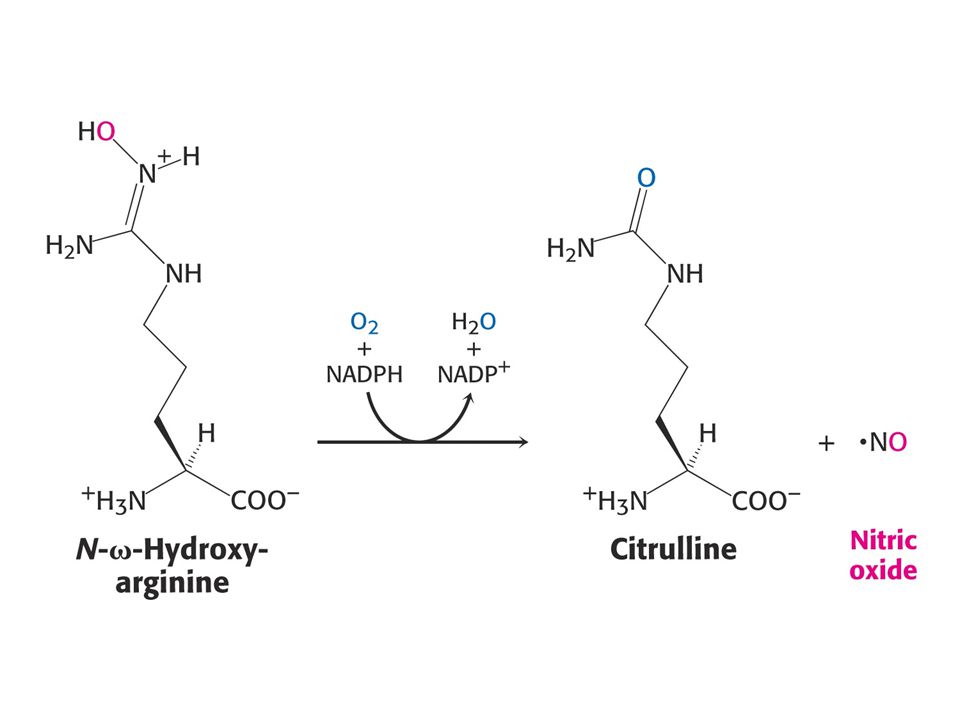
81
Oxid dusnatý (NO), signální molekula s krátkou dobou života se tvoří z Arg. Enzym je NO synthasa (NOS) využívající kyslík a NADPH. Reakce probíhá přes na enzymu vázaný hydroxyArg. NOS je homodimer mezi 120 až 160 kd/podjednotka. Každá podjednotka obsahuje jeden FMN, jeden FAD, jeden tetrahydrobiopterin a jeden Fe3+ hem. Pětielektronová redukce Arg.
 využívající kyslík a NADPH. Reakce probíhá přes na enzymu vázaný hydroxyArg. NOS je homodimer mezi 120 až 160 kd/podjednotka. Každá podjednotka obsahuje jeden FMN, jeden FAD, jeden tetrahydrobiopterin a jeden Fe3+ hem. Pětielektronová redukce Arg..")
84
NO je plyn rychle difundující přes buněčné membrány, vysoce reaktivní (poločas 5 s) působí do vzdálenosti 1 mm od místa tvorby. Tvoří se v buňkách endothelu. Neurony také syntetizují NO (neurony jsou z 55% homologní s endothelem). NO syntetizovaný neurony rozšiřuje mozkové a jiné artérie. Nejvíce NOS obsahuje mozek. NO je důležitý pro funkci CNS. NO se také syntetizuje v leukocytech – zde je NO součástí cytotoxické funkce. NO spolu se superoxidem vytváří reaktivní hydroxylový radikál, který zneškodňuje vstupující bakterie. NO se váže a tak aktivuje guanylátcyklasu – enzym uplatňující se při přenosu signálu. Enzym je homologem adenylátcyklasy. Obsahuje hem s doménou vážící NO.

85
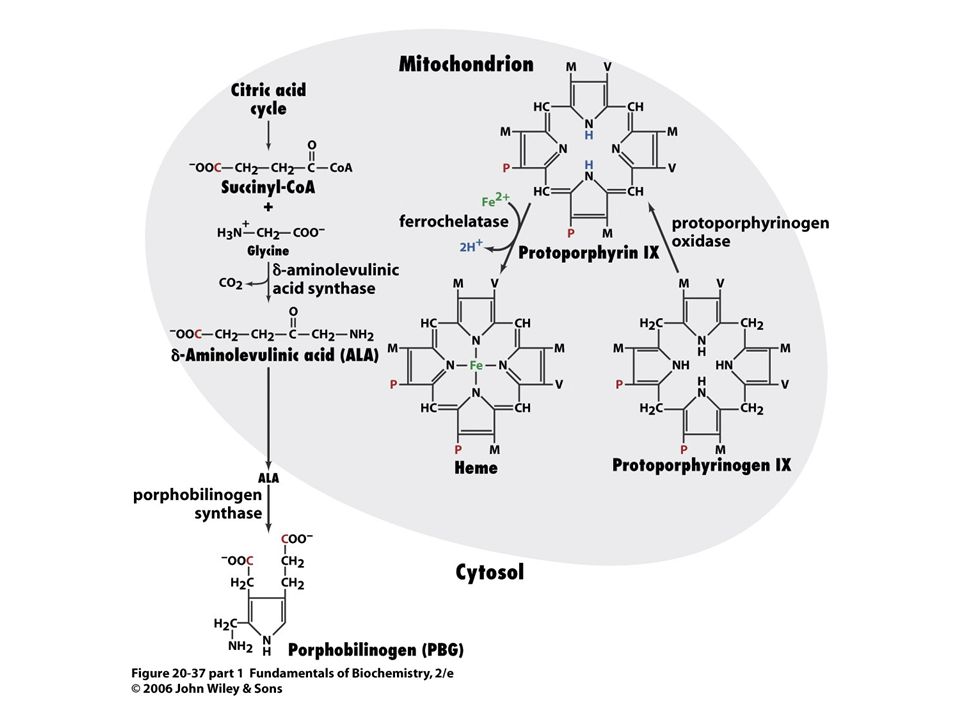
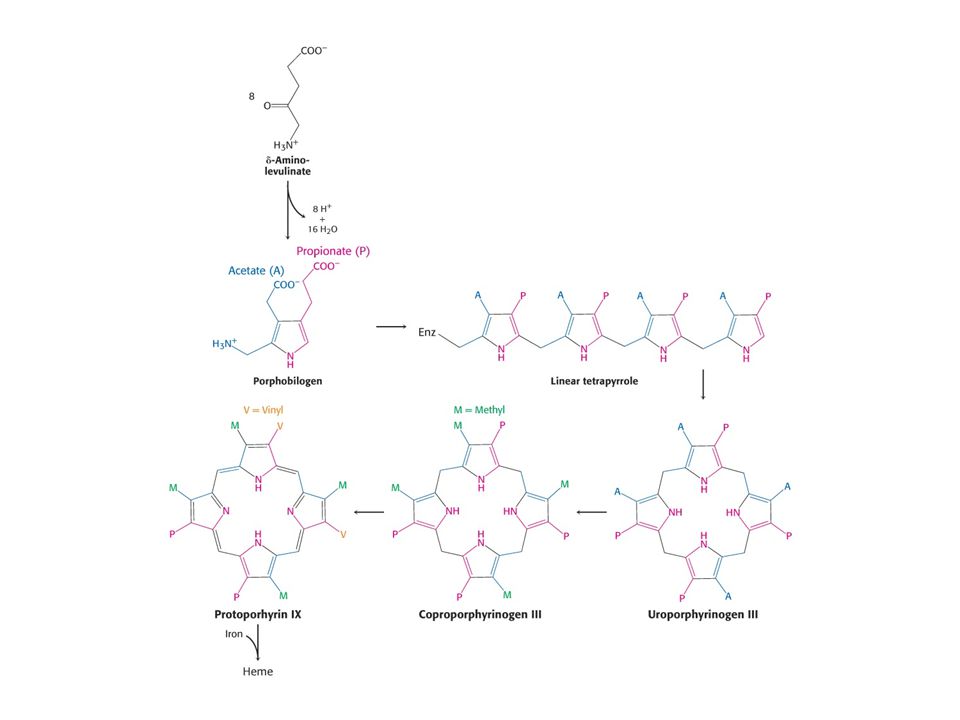
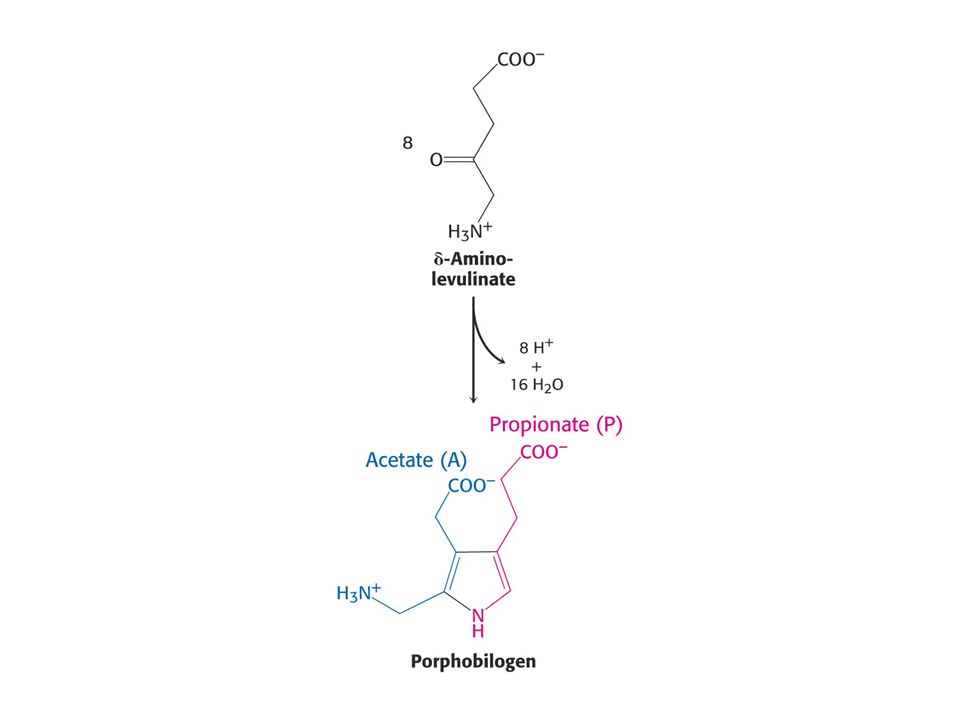
Savčí porfyriny se syntetizují z Gly a sukcinylCoA.
Reakce je katalyzována d-aminolevulinátsynthasou (PLP, v mitochondrii).
.")
90
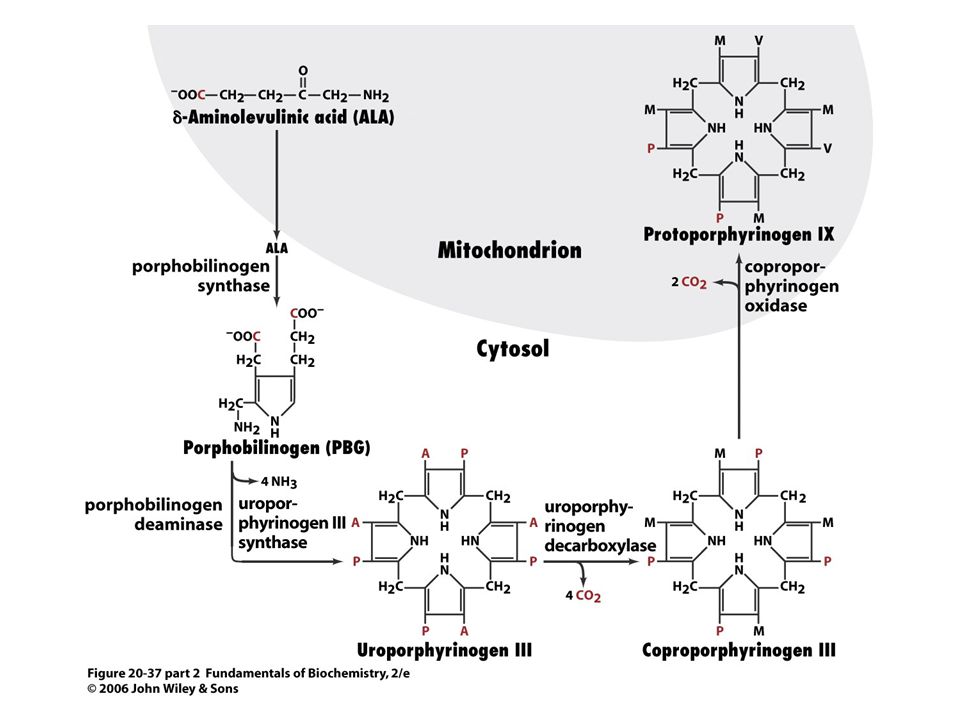
Porfobilinogendeaminasa, kondenzce hlava – ocas.

91
Je nutný enzym KOSYNTHASA-vzniká uroporfyrinogen III, který má asymetrické vedlejší řetězce. Meziprodukt při syntéze vitaminu B12 a chlorofylu u baktérií a rostlin.

92
Koproporfyriogen III se tvoří dekarboxylací acetátových vedlejších řetězců

93
Desaturace porfyrinového kruhu a převedení dvou propionátových vedlejších řetězců na vinyly.

95
Regulace biosyntézy hemu.
Největší podíl biosyntézy hemu v erythroidních buňkách (kostní dřeň) 85%-zbytek v játrech. Biosyntéza v játrech závisí a metabolickém stavu. Nutnost detoxikace vede k syntéze hemu pro cytochrom P450. V erythroidních buňkách dochází k syntéze tak, aby se proteinová část hemoglobinu (globin) a hem mohly obměnit za 120 dnů, což je životnost erythrocytu. V játrech je kontrolním bodem synthasa d-aminlevulinátu (ALA). Aktivitu kontroluje hladina hemu a jeho oxidačního produktu heminu. Inhibují transport ALA z cytosolu do matrix a jeho biosyntézu. V erytroidních buňkách indukuje hem biosyntézu globinu v retikulocytech (nezralé erythrocyty). Hlavními rychlost limitujícími enzymy jsou ferrochelatasa a porfobilinogendeaminasa a ne ALA synthasová reakce.
 85%-zbytek v játrech. Biosyntéza v játrech závisí a metabolickém stavu. Nutnost detoxikace vede k syntéze hemu pro cytochrom P450. V erythroidních buňkách dochází k syntéze tak, aby se proteinová část hemoglobinu (globin) a hem mohly obměnit za 120 dnů, což je životnost erythrocytu. V játrech je kontrolním bodem synthasa d-aminlevulinátu (ALA). Aktivitu kontroluje hladina hemu a jeho oxidačního produktu heminu. Inhibují transport ALA z cytosolu do matrix a jeho biosyntézu. V erytroidních buňkách indukuje hem biosyntézu globinu v retikulocytech (nezralé erythrocyty). Hlavními rychlost limitujícími enzymy jsou ferrochelatasa a porfobilinogendeaminasa a ne ALA synthasová reakce.")
96
Hem je prosthetická skupina myoglobinu, hemoglobinu, katalasy, peroxidasy a cytochromu cytochromu P450. Vstup Fe++ je katalyzován ferrochelatasou. Železo je transportováno plasmou transferinem, proteinem vážícím dva atomy Fe3+ . Skladován je ve tkáních uvnitř ferritinu, který ve své kavitě může pojmout až Fe3+.

97
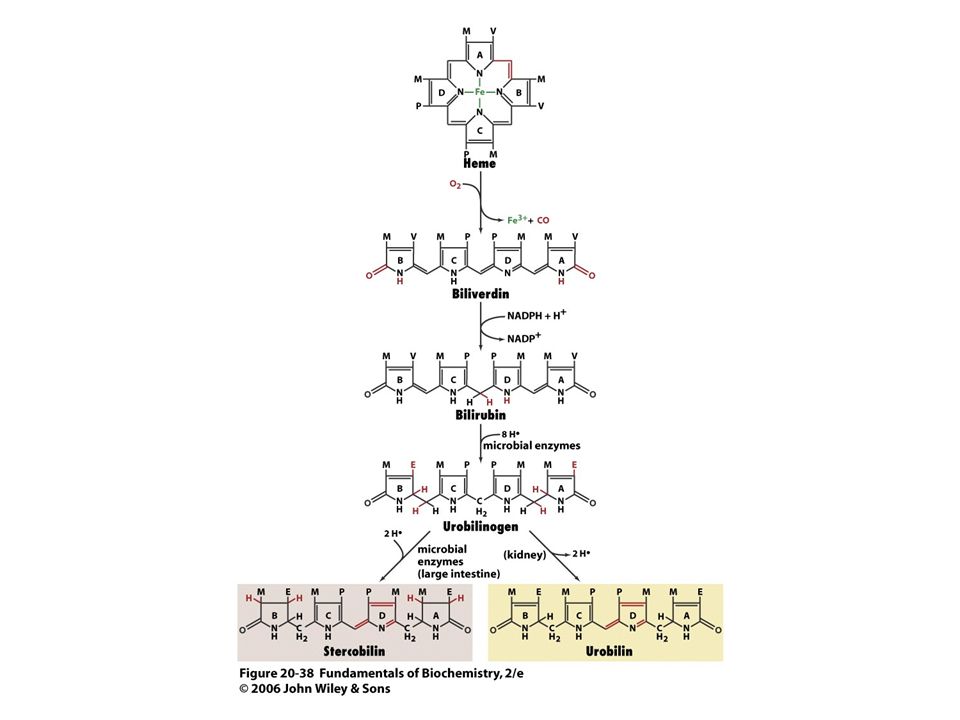
Degradace hemu. Doba života erythrocytu je asi 120 dnů.
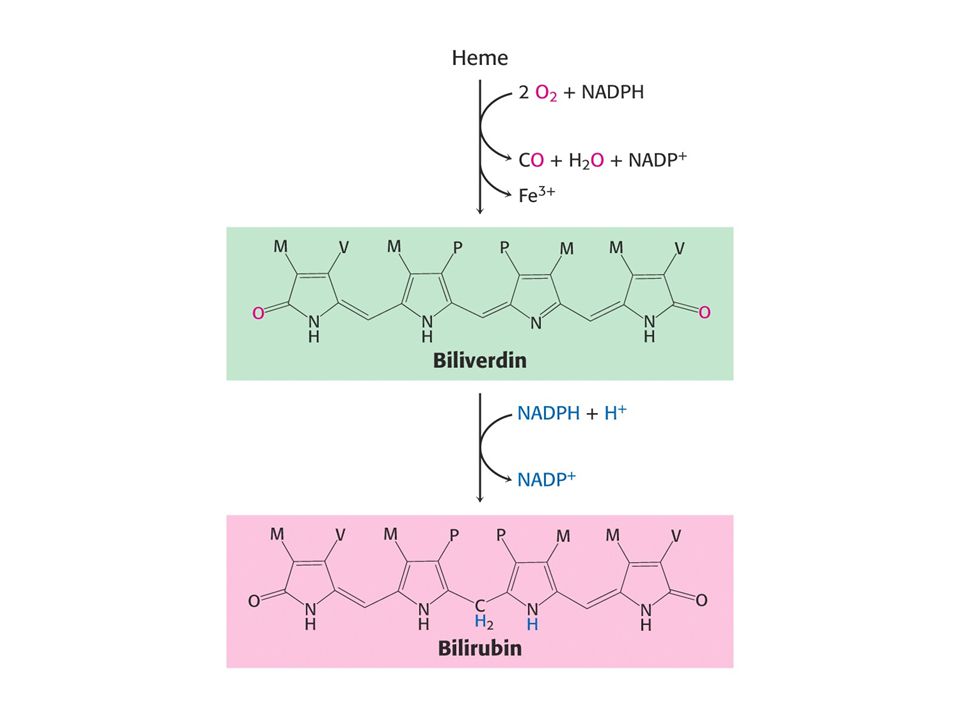
Prvním stupněm při degradaci hemu je štěpení a-methenového můstku za tvorby zeleného biliverdinu (lineární tetrapyrrol). Následuje redukce centrálního methenového můstku enzymem biliverdinreduktasa za tvorby bilirubinu (červené barvivo).
. Následuje redukce centrálního methenového můstku enzymem biliverdinreduktasa za tvorby bilirubinu (červené barvivo).")
100
Dědičné poruchy metabolismu porfyrinů.
Porfyrie – deficience některých enzymů syntézy hemu. Spojené se změnou barvy např. moči. Congenital erythropoietic porphyria (vrozená erythrocytární porfyrie) – rozpad ethrocytů. Nedostatek aktivity kosynthasy. Přebytek symetrického uroporfyrinogenu I. Moč pacientů je červená, zuby červeně fluoreskují při ozáření UV světlem. Pokožka je velmi citlivá a světlo, protože fotoexcitované porfyriny jsou velmi reaktivní. Akutní intermittent (občasná, přerušovaná) porfyrie – nadprodukce porfobilinogenu a d-aminolevulinátu. Poškozuje játra. Bolesti břicha a neurologické potíže. The „madness“ of George III King of Egland během Americké revoluce byla důsledkem této nemoci.
 – rozpad ethrocytů. Nedostatek aktivity kosynthasy. Přebytek symetrického uroporfyrinogenu I. Moč pacientů je červená, zuby červeně fluoreskují při ozáření UV světlem. Pokožka je velmi citlivá a světlo, protože fotoexcitované porfyriny jsou velmi reaktivní. Akutní intermittent (občasná, přerušovaná) porfyrie – nadprodukce porfobilinogenu a d-aminolevulinátu. Poškozuje játra. Bolesti břicha a neurologické potíže. The „madness of George III King of Egland během Americké revoluce byla důsledkem této nemoci.")
101
Kontrola pochopení přednášky.
Napište rovnovážnou rovnici syntézy alaninu z glukosy. Které meziprodukty jsou na cestě od dusíku (N2) k hemu ? Které deriváty folátu jsou reaktanty při přeměně: a) Gly na Ser ? b) Homocysteinu na Met ? Většina cytosolárních proteinů postrádá disulfidové vazby, zatímco extracelulární proteiny je obvykle obsahují. Proč ? Syntéza d-aminolevulinátu probíhá v matrix mitochondrie, zatímco tvorba profobilinogenu v cytoplasmě. Jaký je důvod proto, aby první krok biosyntézy probíhal v matrix ? ě
 k hemu Které deriváty folátu jsou reaktanty při přeměně: a) Gly na Ser b) Homocysteinu na Met Většina cytosolárních proteinů postrádá disulfidové vazby, zatímco extracelulární proteiny je obvykle obsahují. Proč Syntéza d-aminolevulinátu probíhá v matrix mitochondrie, zatímco tvorba profobilinogenu v cytoplasmě. Jaký je důvod proto, aby první krok biosyntézy probíhal v matrix ě.")
102
Pro následující příklad větvené dráhy navrhněte zpětnovazebné inhibiční schéma, které povede k produkci stejného množství Y a Z.

103
Jakým může zvýšená syntéza Asp a Glu ovlivnit produkci energie v buňce ?
Jak může buňka odpovědět na takový efekt ?

Podobné prezentace