Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Miroslav Strnad / Dušan Lazár
oddělení biofyziky CRH a Katedra biofyziky, Př.f., UP Fotosyntéza - světelné reakce [kapitola 7] Olomouc

2
Úvod Fotosyntéza je jeden z nejdůležitějších procesů probíhajících na Zemi, protože vede k vývoji kyslíku a produkci biomasy Snaha o umělou fotosyntézu: - vývoj kyslíku - abychom žili nový zdroj energie: tvorba vodíku u některých řas a sinic (přímá fotolýza (viz obr. dále) nebo nepřímá fotolýza (anaerobní fermentace)) a bakterií (fotofermantace) schopnost fotosyntetického aparátu přeměňovat dopadající světelnou energie v elektrický proud
 nebo nepřímá fotolýza (anaerobní fermentace)) a bakterií (fotofermantace) schopnost fotosyntetického aparátu přeměňovat dopadající světelnou energie v elektrický proud.")
3
použití fotosystému jako generátoru elektrického napětí napříč biologickou membránu - fotosystém I, po zabudování do elektricky nefunkčních (neexcitovatelných) membrán živočišných buněk, je schopen obnovit excitovatelnost (ve smyslu vzniku akčního potenciálu) těchto membrán. Čili, elektrické napětí generované skrz membránu fotosystémem I je schopné otevřít napěťově řízené kanály v membráně, čehož lze využít při aktivování nefunkčních nervových nebo svalových buněk. - použití mechanismu nábojové separace v reakčním centru ke tvorbě procesorů do PC - ve fotosyntéze se nábojová separace odehrává v čase okolo 1 ps, kdežto taktovací frekvence procesorů ve standardním PC jsou řádově GHz (stovky pikosekund) => fotosyntéza je řádově 100 krát rychlejší.
 => fotosyntéza je řádově 100 krát rychlejší.")
4
použití fotosystému jako fotoaparátu - fotosystém 1 sinice připojen pomocí molekulárního drátu (vitamín K1) na vstup tranzistoru řízeného elektrickým polem => čím více excitací dopadá na fotosystém 1, tím více elektronů se z něho uvolňuje a tím větší je elektrický proud generovaný v tranzistoru použití sinic jako zdroj látek - jednobuněčné sinice (prokaryotní autotrofy) samy o sobě, ale také jako hostující organismus, mohou být využity pro syntézu důležitých přírodních organických látek => je snaha detailně poznat metabolismus sinic a to jak pomocí experimentů, tak i pomocí matematického modelování; nejdále je výzkum se sinicí Synechocystis sp. PCC 6803.
 samy o sobě, ale také jako hostující organismus, mohou být využity pro syntézu důležitých přírodních organických látek => je snaha detailně poznat metabolismus sinic a to jak pomocí experimentů, tak i pomocí matematického modelování; nejdále je výzkum se sinicí Synechocystis sp. PCC")
5
Fotosyntézu lze rozdělit na 2 fáze:
- světlená fáze - pro průběh těchto reakcí je nutná přítomnost excitační energie ve formě absorbovaného světla - temnostní fáze - pro průběh těchto reakcí není potřeba světla, ale produktů světelné fáze fotosyntézy

6
list a buňka chloroplast a thylakoidní membrána

7
PS II, cyt b6f, PS I, FNR, ATP-synt.
Světelná fáze fotosyntézy – thylakoidní membrána PS II, cyt b6f, PS I, FNR, ATP-synt. hn + H2O + NADP+ + ADP + Pi O2 + NADPH + ATP

8
Lipidová dvojvsrsrva - thylakoidní membrána
- thylakoidní membrána je lipidová dvojvrstva - lipid, který převážně tvoří thylakoidní membránu je digalaktosyldiacylglycerid (DGDG) - má polární hydrofilní (= snáší se s vodou) hlavičku a hydrofobní (nesnáší se s vodou) ocasy - pro lipidovou dvouvrstvu obecně existuje model “tekuté mozaiky” - integrální proteiny a proteiny na vnitřní a vnější straně membrány volně “plují” v membráně
 - má polární hydrofilní (= snáší se s vodou) hlavičku a hydrofobní (nesnáší se s vodou) ocasy. - pro lipidovou dvouvrstvu obecně existuje model tekuté mozaiky - integrální proteiny a proteiny na vnitřní a vnější straně membrány volně plují v membráně.")
9
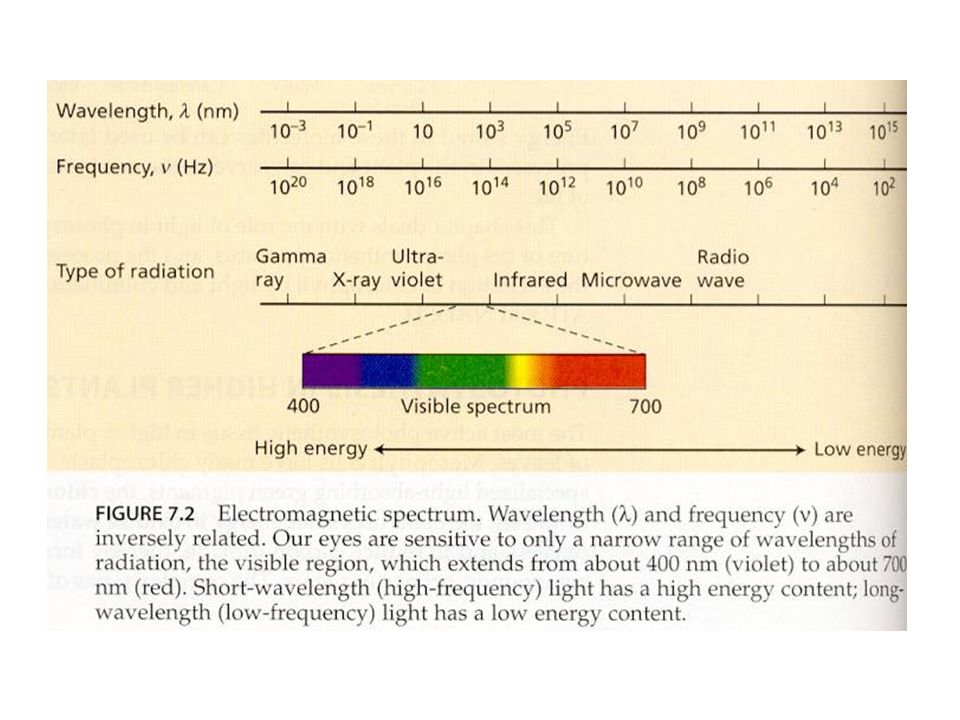
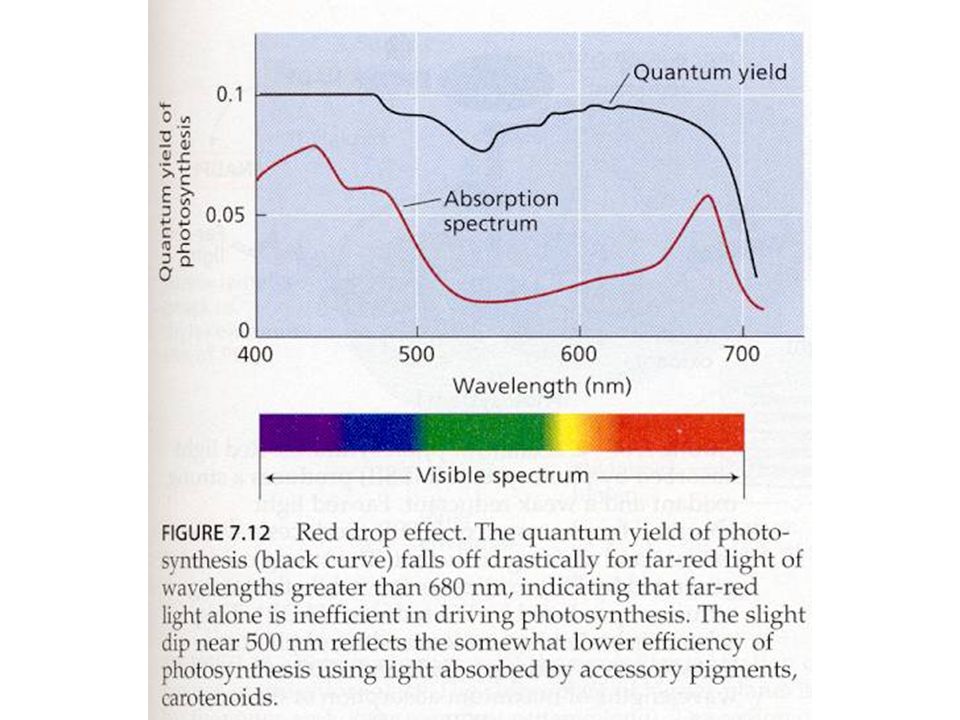
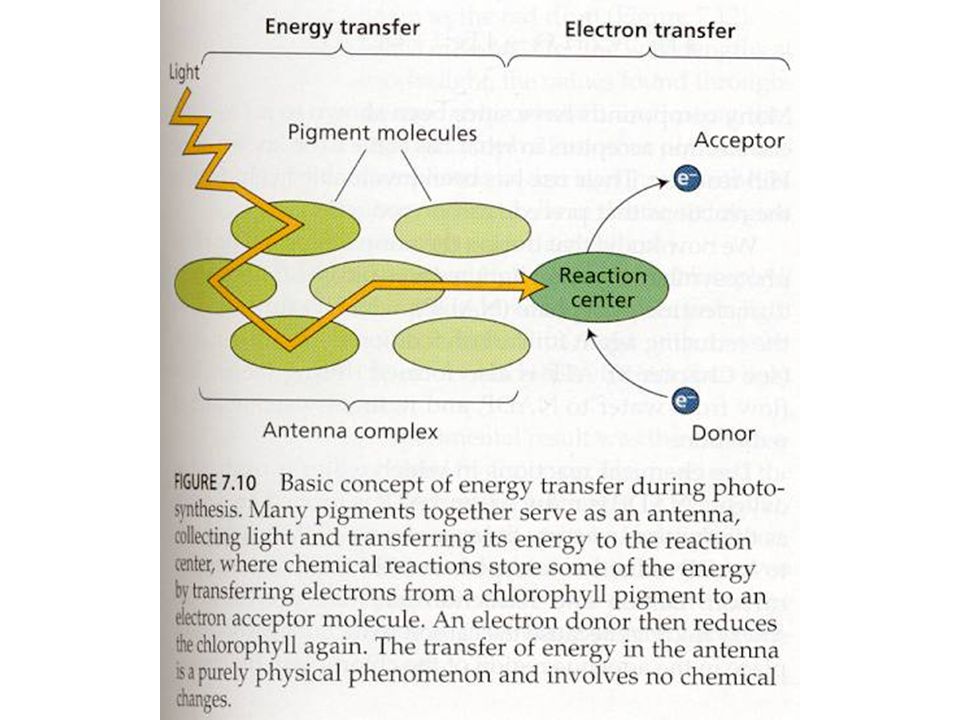
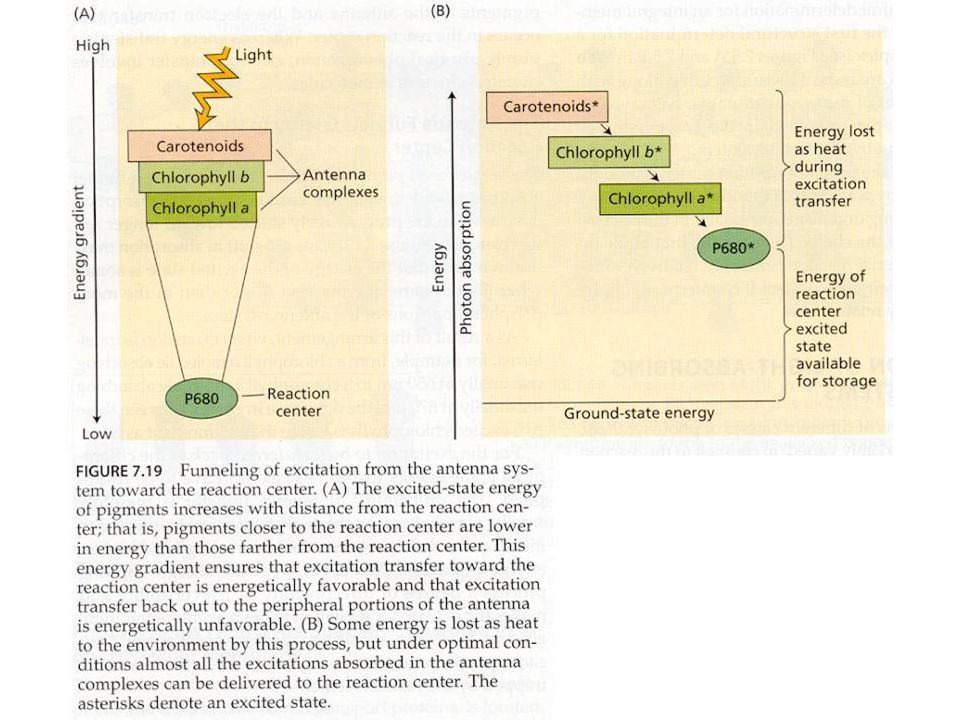
Světlo

14
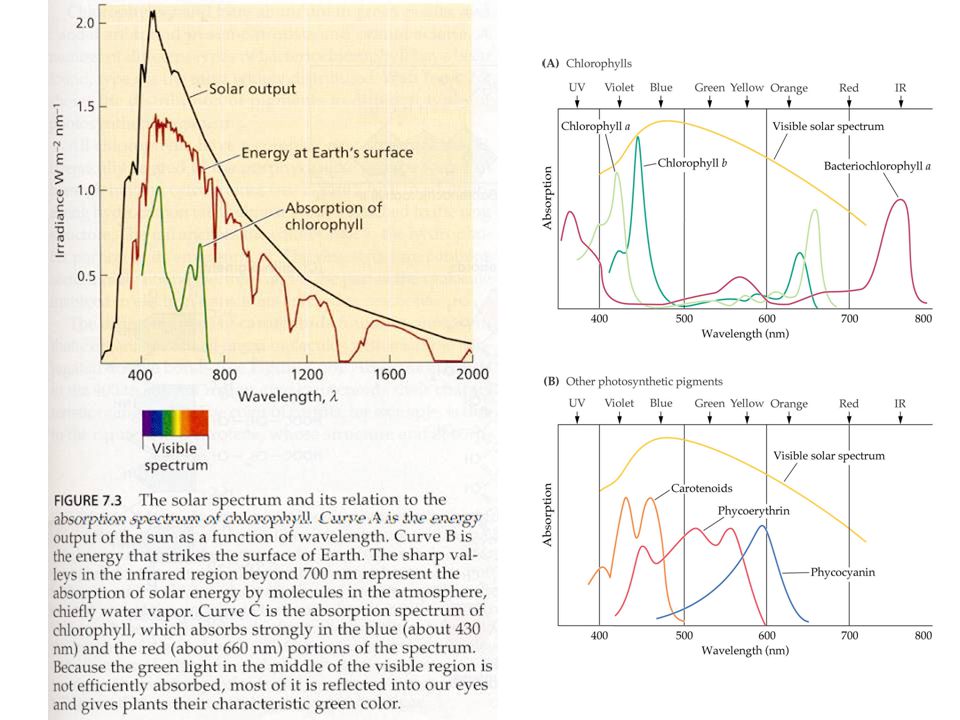
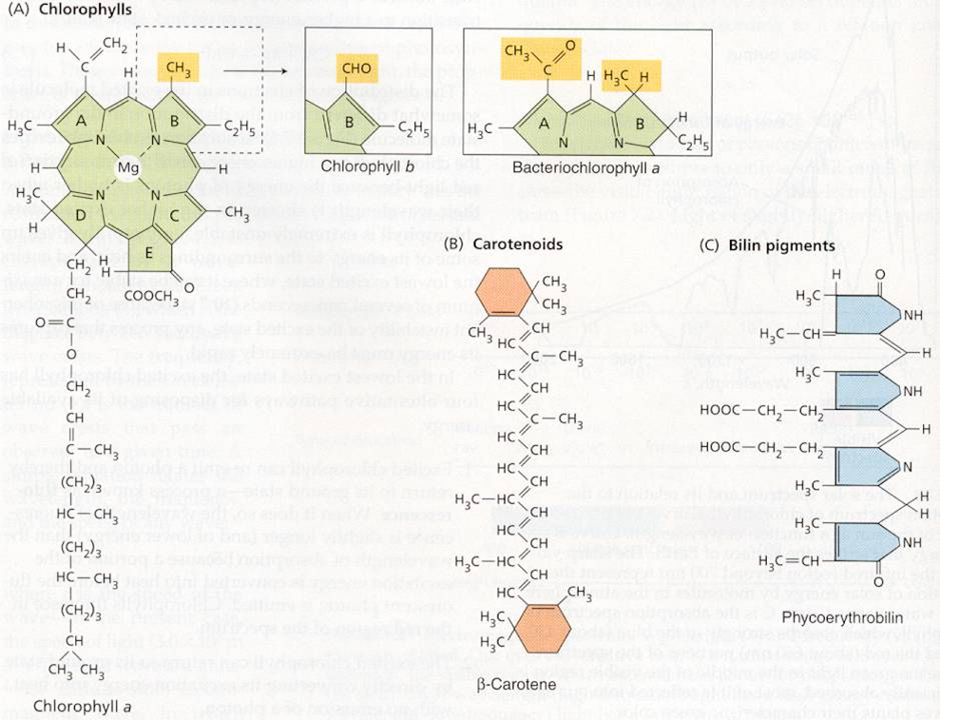
Světlosběrné komplexy
světlosběrné komplexy (LHC) vyšších rostlin a řas (chlorofyly a karotenoidy) fykobilisomy sinic a některých řas (fykocyanin, fykoerytrin)
 vyšších rostlin a řas (chlorofyly a karotenoidy) fykobilisomy sinic a některých řas. (fykocyanin, fykoerytrin)")
20
Přenos elektronů přenos elektronů z H2O na NADP+ se děje díky vhodným redoxním potenciálům jednotlivých přenašečů – to zobrazuje tzv. „Z-schéma“

21
Redoxní potenciál: - je to relativní schopnost (vyjádřena ve voltech) dané látky přijmout elektron (= být redukována) vzhledem k atomu vodíky, jehož redoxní potenciál je roven 0 V (dohoda) - čím je redoxní potenciál dané látky více kladný, tím větší má táto látka tendenci přijímat elektrony a být redukována - standardní redoxní potenciál (E0) je měřen za standardních podmínek (25 C, koncentrace 1 M všech látek, tlak 1 atm (101,325 kPa)) čím je větší rozdíl mezi redoxními potenciály 2 látek (DE), tím větší je „síla“, s kterou elektrony budou samovolně putovat z látky s méně pozitivním na látku s více pozitivním redoxním potenciálem redoxní potenciály přenašečů elektronů v Z-schématu představují jakési efektivní potenciály (přenašeče jsou umístěny v proteinech), které se navíc mění v čase přenos elektronů v proteinech a jeho rychlost se striktně neřídí pouze DE, protože v proteinech dochází k tunelovému jevu (Marcusova teorie), který může být i proti spádu energie
 dané látky přijmout elektron (= být redukována) vzhledem k atomu vodíky, jehož redoxní potenciál je roven 0 V (dohoda) - čím je redoxní potenciál dané látky více kladný, tím větší má táto látka tendenci přijímat elektrony a být redukována. - standardní redoxní potenciál (E0) je měřen za standardních podmínek (25 C, koncentrace 1 M všech látek, tlak 1 atm (101,325 kPa)) čím je větší rozdíl mezi redoxními potenciály 2 látek (DE), tím větší je „síla , s kterou elektrony budou samovolně putovat z látky s méně pozitivním na látku s více pozitivním redoxním potenciálem. redoxní potenciály přenašečů elektronů v Z-schématu představují jakési efektivní potenciály (přenašeče jsou umístěny v proteinech), které se navíc mění v čase. přenos elektronů v proteinech a jeho rychlost se striktně neřídí pouze DE, protože v proteinech dochází k tunelovému jevu (Marcusova teorie), který může být i proti spádu energie.")
22
Fotosystém II (PSII) tzv. reakční centrum (RC) PSII tvoří 2 proteiny (D1 a D2) a cyt b559 - PS II má také svoje vnitřní světlosběrné komplexy CP43 a CP47 PSII je pravděpodobně jako dimér - přenos elektronů skrz PheoD2 (dolní obr.) je neaktivní - tzv. “neaktivní B-větev” - cytochrom b559 je důležitý pro tvorbu RC PS II a zprostředkovává cyklický transport elektronů okolo PSII - TyrD velmi pomalu donuje elektrony QB - sekundární chinonový akceptor elektronů QA - primární chinonový akceptor elektronů PheoD1 (feotytin) - primární akceptor elektronů ChlD1 (chlorofyl) - primární donor elektronů PD1 a PD2 (chlorofylový dimer, P680) - sekundární donor elektronů TyrZ (= YZ, tyrosin) - terciální donor elektronů OEC - komplex vyvíjející kyslík
 je neaktivní - tzv. neaktivní B-větev - cytochrom b559 je důležitý pro tvorbu RC PS II a zprostředkovává cyklický transport elektronů okolo PSII. - TyrD velmi pomalu donuje elektrony. QB - sekundární chinonový akceptor elektronů. QA - primární chinonový akceptor elektronů. PheoD1 (feotytin) - primární akceptor elektronů. ChlD1 (chlorofyl) - primární donor elektronů. PD1 a PD2 (chlorofylový dimer, P680) - sekundární donor elektronů. TyrZ (= YZ, tyrosin) - terciální donor elektronů. OEC - komplex vyvíjející kyslík.")
23
Komplex vyvíjející kyslík (OEC, WOC)
Redukovaný TyrZ je oxidován elektrony z OEC, který tímto postupně přecházi v cyklu přes své tzv. S-stavy. OEC je tvořen manganovým komplexem (4 atomy Mn), ze kterého se postupně v jednotivých S-přechodech uvolňují elektrony a protony. Před uzavřením S-přechodů do cyklu dojde ke štěpení H20 na O2 a uvolnění elektronů (a protonů) z vody, kterými je manganový komplex zase redukován na “počáteční” stav. Pro stabilizaci manganového komplexu jsou důležité i Ca2+ a Cl- ionty.
, ze kterého se postupně v jednotivých S-přechodech uvolňují elektrony a protony. Před uzavřením S-přechodů do cyklu dojde ke štěpení H20 na O2 a uvolnění elektronů (a protonů) z vody, kterými je manganový komplex zase redukován na počáteční stav. Pro stabilizaci manganového komplexu jsou důležité i Ca2+ a Cl- ionty.")
24
Cytochrom (cyt) b6/f Cyt b6f zprostředkovává přenos elektronů od PS II na PS I, tzv. lineární přenos elektronů, ale i cyklický přenos elektronů okolo PS I (na obr. neukázáno). Obsahuje nízko- (bL) a vysoko- (bH) poteniálové formy hemu b, Rieskeho železnatosirný komplex, hem f a hem c (v okolí bH, není na obrázku). Při přenosů elektronů skrz cyt b6f, tento prochází tzv. Q-cyklem (existuje i semi Q-cyklus), který kromě oxidace redukovaného plastochinonu (PQH2) umožňuje redukci oxidovaného PQ. Při lineárním přenosu elektronů pouze jeden elektron z PQH2 jde na cyt f a druhý elektron jde na Q cyklus.
. Obsahuje nízko- (bL) a vysoko- (bH) poteniálové formy hemu b, Rieskeho železnatosirný komplex, hem f a hem c (v okolí bH, není na obrázku). Při přenosů elektronů skrz cyt b6f, tento prochází tzv. Q-cyklem (existuje i semi Q-cyklus), který kromě oxidace redukovaného plastochinonu (PQH2) umožňuje redukci oxidovaného PQ. Při lineárním přenosu elektronů pouze jeden elektron z PQH2 jde na cyt f a druhý elektron jde na Q cyklus.")
25
Fotosystém I (PSI) Fdx - feredoxin
Fx, FA, FB - železnatosirné akceptory elektronů A1 (fylochinon) - sekundarní akceptor elektronů A0 (chlorofyl) - primární akceptor elektronů ChlB(A) (chlorofyl) - primární donor elektronů P700 (chlorofylový dimer) - sekundární donor elektronů PC (plastocyanin) - terciální donor elektronů PSI je pravděpodobně jako monomér - jako u PS II, i zde existuje tzv. “neaktivní B-větev” (na obr. nahoře nezobrazena)
 - sekundarní akceptor elektronů. A0 (chlorofyl) - primární akceptor elektronů. ChlB(A) (chlorofyl) - primární donor elektronů. P700 (chlorofylový dimer) - sekundární donor elektronů. PC (plastocyanin) - terciální donor elektronů. PSI je pravděpodobně jako monomér. - jako u PS II, i zde existuje tzv. neaktivní B-větev (na obr. nahoře nezobrazena)")
26
Porovnání redoxních potenciálů přenašečů elektronů v PSII a PSI a rychlostních konstant přenosů:

27
Plastochinon, plastocyanin, feredoxin

28
ATP-syntáza Tvorba ATP je určována (chemiosmóza) elektrochemickým potenciálem protonů, respektive tzv. protonmotivní silou (pmf), která nezávisí pouze na DpH, ale i na elektrickém napětí DY napříč thylakoidní membránou (pmf = DY + (2.3 R T DpH)/F) a ve fotosyntéze podíl DY na celkové pmf může být až 50%.
 elektrochemickým potenciálem protonů, respektive tzv. protonmotivní silou (pmf), která nezávisí pouze na DpH, ale i na elektrickém napětí DY napříč thylakoidní membránou (pmf = DY + (2.3 R T DpH)/F) a ve fotosyntéze podíl DY na celkové pmf může být až 50%.")
29
Rotor je g podjednotka uvnitř statoru tvořeného a a b podjednotkami.
Točivý momemt je 40 pN nm (= 40 x 10-3 N m; motor „standardního“ auta má řádově 100 N m).
.")
30
Lineární x cyklický přenos elektronů
Za normálních podmínek (ustálené světelné podmínky) se děje převážně lineární přenos elektronů (obrázek vlevo). Při stresu fotosyntetického aparátu, za účelem „udržení“ tvorby příslušného množství NADPH, ATP a gradientu protonů napříč thylakoidní membránou, je aktivován cyklický přenos elektronů okolo PS I (obrázek vpravo). Existuje několik cest toku elektronů při tomto cyklickém transportu, i cesty které nejdou přes feredoxin (nejsou ukázány na obrázku vpravo).
 se děje převážně lineární přenos elektronů (obrázek vlevo). Při stresu fotosyntetického aparátu, za účelem „udržení tvorby příslušného množství NADPH, ATP a gradientu protonů napříč thylakoidní membránou, je aktivován cyklický přenos elektronů okolo PS I (obrázek vpravo). Existuje několik cest toku elektronů při tomto cyklickém transportu, i cesty které nejdou přes feredoxin (nejsou ukázány na obrázku vpravo).")
31
Heterogenita fotosystémů
Neexistuje pouze jeden typ PSII a jeden typ PSI, ale různé typy => fotosystémy jsou obecně heterogenní. Pro PSII i PSI existují různé typy heterogenit vzhledem k tomu, která funkční vlastnost se konkrétně sleduje. Tyto funkční heterogenity jsou spojené i s heterogenitou rozmístění jednotlivých typů PSII a PSI v thylakoidní membráně.

32
Stres, degradace a oprava
Působením stresů dochází k tvorbě reaktivních forem kyslíku (ROS), které pak způsobují degradaci fotosyntetických proteinů. Existuje mnoho mechanismů, jak předejít působení těchto ROS (zhášení excitovaných stavů a tzv. voda-voda cyklus). Pokud k degradaci dojde, existují pak opravné mechanismy, které umožňují syntézu nových proteinů.
, které pak způsobují degradaci fotosyntetických proteinů. Existuje mnoho mechanismů, jak předejít působení těchto ROS (zhášení excitovaných stavů a tzv. voda-voda cyklus). Pokud k degradaci dojde, existují pak opravné mechanismy, které umožňují syntézu nových proteinů.")
33
Zhášení excitovaných stavů – nefotochemické zhášení (qN, NPQ)
Existuje několik typů zhášení excitovaných stavů: Energeticky závislé (qE) zhášení – „nejčastější“ závisí na akumulaci protonů v lumenu (u řas a vyšších rostlin), vedoucí k různým změnám na úrovni PSII, viz obrázek níže, které způsobují zhášení excitovaných stavů. Zhášení způsobené stavovými přechody (qT) – v delší časové škále, než se uskutečňuje qE, pokud je pořád přebytek excitaci na PSII, dojde k odpojení části LHCII a k jejímu připojeni k PSI, tzv. stavový přechod (stav 1 - stav 2 přechod). Fotoinhibiční zhášení (qI) – pokud je delší dobu fotosyntetický aparát vystavený hodně silné intenzitě světla.
 zhášení – „nejčastější závisí na akumulaci protonů v lumenu (u řas a vyšších rostlin), vedoucí k různým změnám na úrovni PSII, viz obrázek níže, které způsobují zhášení excitovaných stavů. Zhášení způsobené stavovými přechody (qT) – v delší časové škále, než se uskutečňuje qE, pokud je pořád přebytek excitaci na PSII, dojde k odpojení části LHCII a k jejímu připojeni k PSI, tzv. stavový přechod (stav 1 - stav 2 přechod). Fotoinhibiční zhášení (qI) – pokud je delší dobu fotosyntetický aparát vystavený hodně silné intenzitě světla.")
34
Zhášení excitovaných stavů – nefotochemické zhášení (qN, NPQ)
U sinic qE funguje úplně jinak – světlem dojde k aktivaci „orange carotenoid protein“ (OCP) a k jeho navázání k fykobylisomům, čímž dojde ke zhášení excitovaných stavů. Pro dosažení původního stavu je třeba navázání „fluorescence recovery protein“ (FRP), který způsobí oddělení OCP. Zhášení excitovaných stavů u sinic, na rozdíl od zhašení excitovaných stavů u řas a vyšších rostlin, nezávisí na akumulaci protonů v lumenu.
 a k jeho navázání k fykobylisomům, čímž dojde ke zhášení excitovaných stavů. Pro dosažení původního stavu je třeba navázání „fluorescence recovery protein (FRP), který způsobí oddělení OCP. Zhášení excitovaných stavů u sinic, na rozdíl od zhašení excitovaných stavů u řas a vyšších rostlin, nezávisí na akumulaci protonů v lumenu.")
35
Voda-voda cyklus Je to sled chemických enzymatických i neenzymatických reakcí (viz obrázek), které mají za účel odstranění ROS. Název voda-voda proto, že elektrony v celém sledu reakcí tečou původně z vody v PSII a končí opět na vodě (viz obrázek).
, které mají za účel odstranění ROS. Název voda-voda proto, že elektrony v celém sledu reakcí tečou původně z vody v PSII a končí opět na vodě (viz obrázek).")
Podobné prezentace


>")

















