Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Obrat proteinů a katabolismus aminokyselin
Proteiny se odbourávají na aminokyseliny Obrat proteinů je přísně regulován Prvním stupněm odbourávání aminokyselin je odstranění dusíku Amoniak je u většiny suchozemských obratlovců odbouráván na močovinu. Uhlíkatý skelet aminokyselin patří k hlavním metabolickým meziproduktům. Vrozené vady metabolismu aminokyselin.

2
Trávení a absorpce proteinů
Žaludek – kys. prostředí – denaturace proteinů. Poté pepsin – nespecifická proteinasa. Lumen tenkého střeva – proteinasy pankreatu trypsin, chymotrypsin (sekrece ve formě zymogenů). Vznikají volné aminokyseliny, zčásti di- a tripeptidy. Aminopeptidasy N – plasmová memrána tenkého střeva. Od N konce. Směs aminokyselin a krátkých peptidů vstupuje z lumen do buněk tenkého střeva a poté do krve.
. Vznikají volné aminokyseliny, zčásti di- a tripeptidy. Aminopeptidasy N – plasmová memrána tenkého střeva. Od N konce. Směs aminokyselin a krátkých peptidů vstupuje. z lumen do buněk tenkého střeva a poté do krve.")
3
Trávení a absorpce proteinů.

4
Obrat proteinů – degradace a syntéza.
Poločas života proteinů se pohybuje od minut do celoživotní doby. Např. ornithindekarboxylasa má t1/2 = 11 min. (podílí se na syntéze polyaminů a tím na růstu a diferenciaci). Hemoglobin má t1/2 shodný s erythrocytem. Protein oční čočky, krystalin, se nemění po celou dobu života.
. Hemoglobin má t1/2 shodný s erythrocytem. Protein oční čočky, krystalin, se nemění po celou dobu života.")
6
Technion – Israel Institute of Technology Haifa, Israel , born 1947
Laureáti Nobelovy ceny za chemii 2004 „for the discovery of ubiquitin-mediated protein degradation" Aaron Ciechanover Technion – Israel Institute of Technology Haifa, Israel , born 1947 Avram Hershko Technion – Israel Institute of Technology Haifa, Israel, born 1937, Karcag, Maďarsko Irwin Rose University of California Irvine, CA, USA , born 1926

7
Aaron Ciechanover Israeli citizen. Born 1947 (57 years) in Haifa, Israel. Doctor's degree in medicine in 1981 at the Technion (Israel Institute of Technology), Haifa. Distinguished Professor at the Unit of Biochemistry, the Rappaport Family Institute for Research in Medical Sciences at the Technion, Haifa, Israel.
 in Haifa, Israel. Doctor s degree in medicine in 1981 at the Technion (Israel Institute of Technology), Haifa. Distinguished Professor at the Unit of Biochemistry, the Rappaport Family Institute for Research in Medical Sciences at the Technion, Haifa, Israel.")
8
Avram Hershko Israeli citizen. Born 1937 (67 years) in Karcag, Hungary. Doctor's degree in medicine in 1969 at the Hadassah and the Hebrew University Medical School, Jerusalem. Distinguished Professor at the Rappaport Family Institute for Research in Medical Sciences at the Technion, Haifa, Israel.
 in Karcag, Hungary. Doctor s degree in medicine in 1969 at the Hadassah and the Hebrew University Medical School, Jerusalem. Distinguished Professor at the Rappaport Family Institute for Research in Medical Sciences at the Technion, Haifa, Israel.")
9
Irwin Rose American citizen. Born 1926 (78 years) in New York, USA. Doctor's degree in in 1952 at the University of Chicago, USA. Specialist at the Department of Physiology and Biophysics, College of Medicine, University of California, Irvine, USA.
 in New York, USA. Doctor s degree in in 1952 at the University of Chicago, USA. Specialist at the Department of Physiology and Biophysics, College of Medicine, University of California, Irvine, USA.")
10
Jak se rozpoznávají proteiny, které se budou odbourávat ?
UBIQUITIN !!! Černá tečka !! Polibek smrti !! Zkratka Ub. Malý protein – 8, 5 kd. C konec pro vazbu na jiné proteiny. Lys 48 vazba dalších molekul ubiquitinu. Přítomen ve všech eukaryotních buňkách.

11
Karboxyl terminálního Gly Ub se kovalentně váže na e-aminoskupinu různých lysinových zbytků proteinů určených k degradaci (isopeptodvá vazba).
.")
12
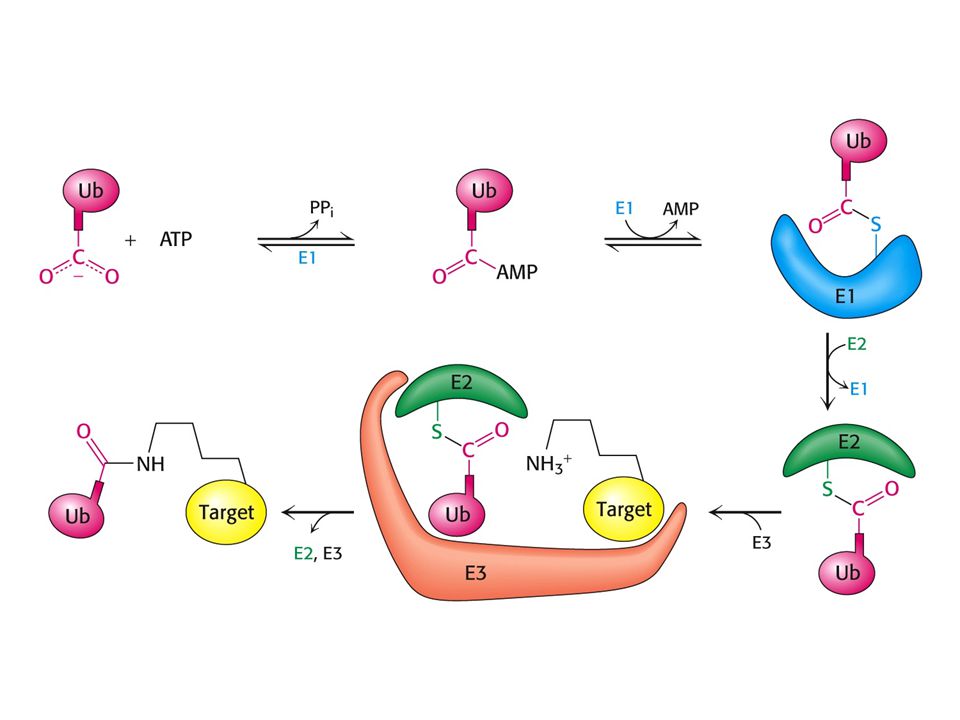
Konjugace ubiquitinu. E1 - ubiquitin aktivující enzym
E2 – ubiquitin konjugační enzym E3 – ubiquitinproteinligasa ATP aktivována – podoba s aktivací mastných kyselin. Efektivní jsou až čtyři navázané molekuly Ub

14
Tetraubiquitin

15
Co je určující pro to, který protein bude označen Ub ?
Délka, poločas života cytosolárních proteinů, je dán charakterem aminokyselin na N konci. Označeno jako N-terminální pravidlo. Např. protein s Met na konci má poločas života více než 20 hod. Stejný protein s Arg nebo Leu na N konci, jen 2 minuty. Enzymy E3 čtou N-konce. Druhým markerem jsou tzv. cyklin destrukční partie (části) řetězce proteinu. Jsou to sekvence aminokyselin, které označují destrukční proteiny buněčného cyklu a proteiny bohaté na Pro, Glu, Ser a Thr (PEST sekvence).
 řetězce proteinu. Jsou to sekvence aminokyselin, které označují destrukční proteiny buněčného cyklu a proteiny bohaté na Pro, Glu, Ser a Thr (PEST sekvence).")
16
Vykonavatel destrukce proteinů.
PROTEASOM – proteinasový komplex nazývaný také 26S proteasom. Poháněno ATP. Komplex: 20S proteasom s katalytickou aktivitou 19S regulační podjednotka. Ub je recyklován

18
Odstranění dusíku – první stupeň odbourávání aminokyselin.
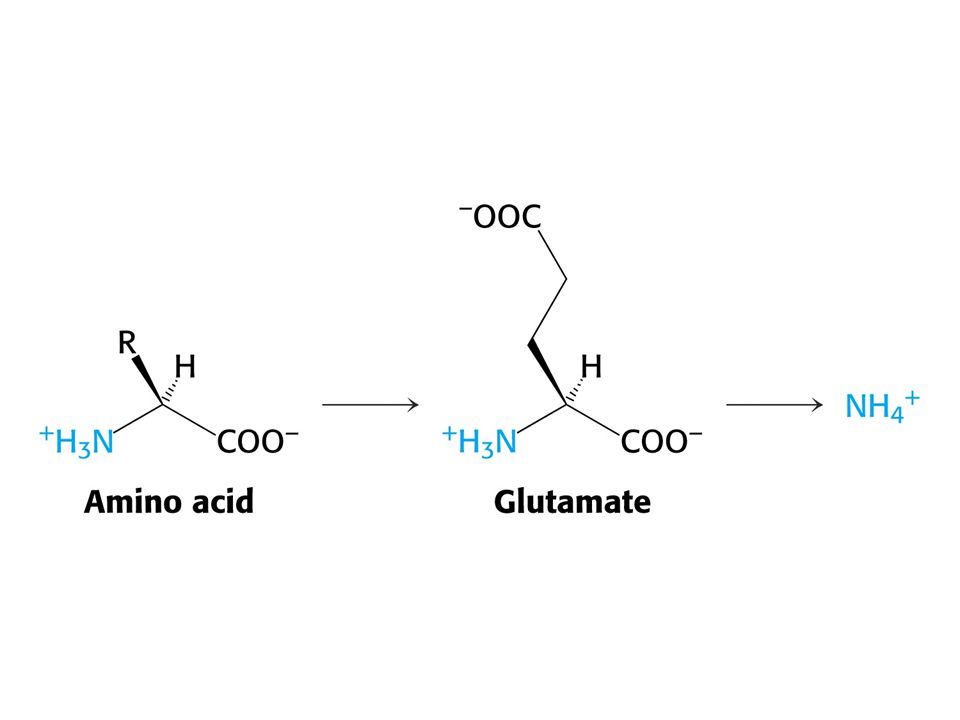
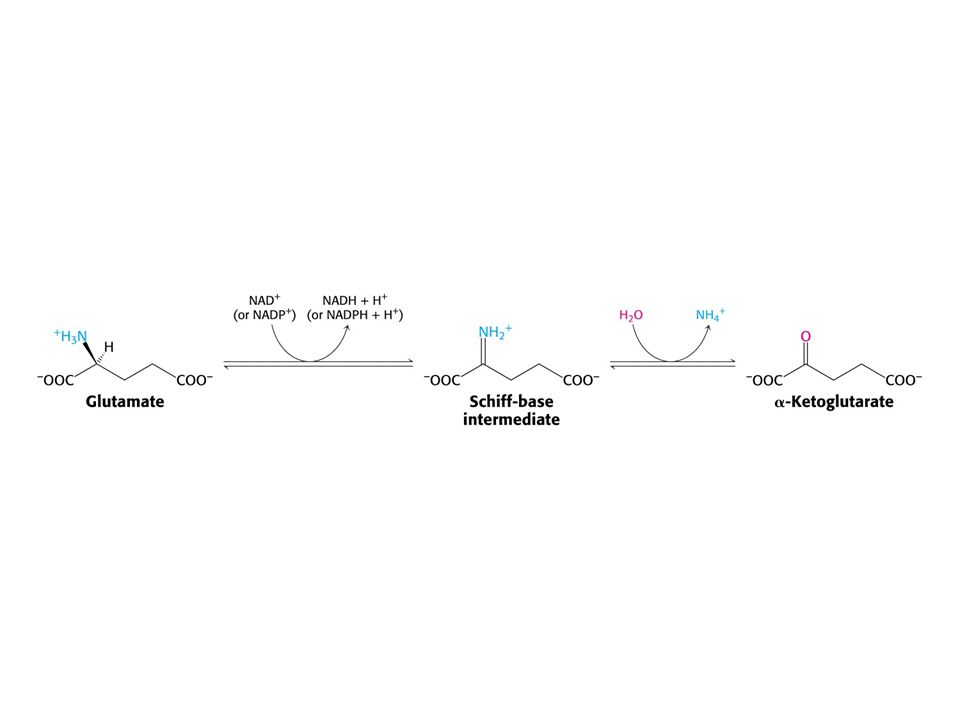
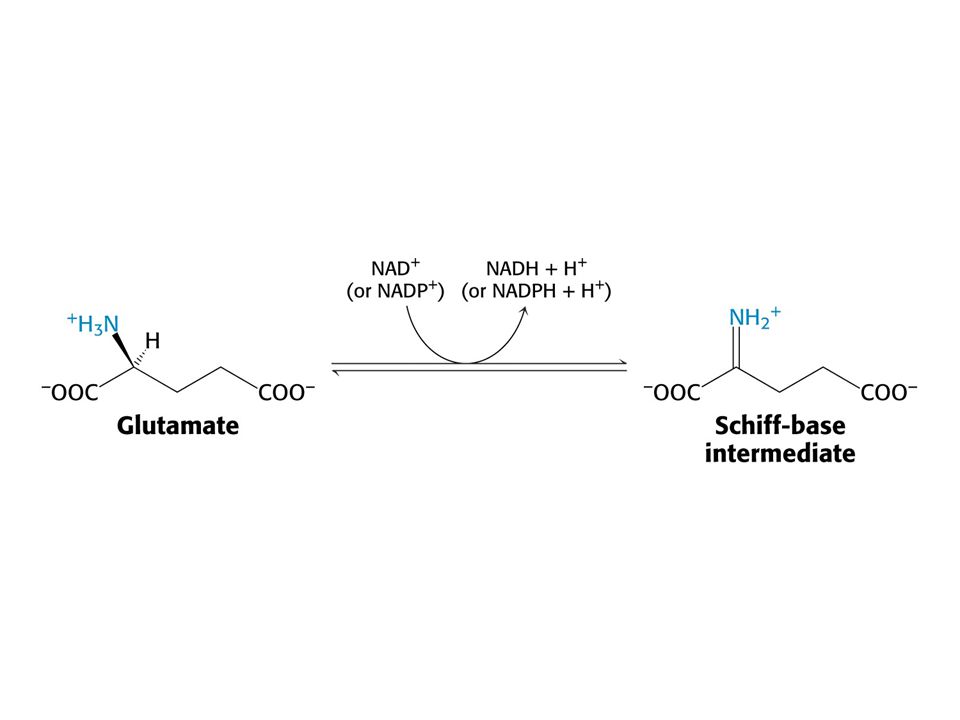
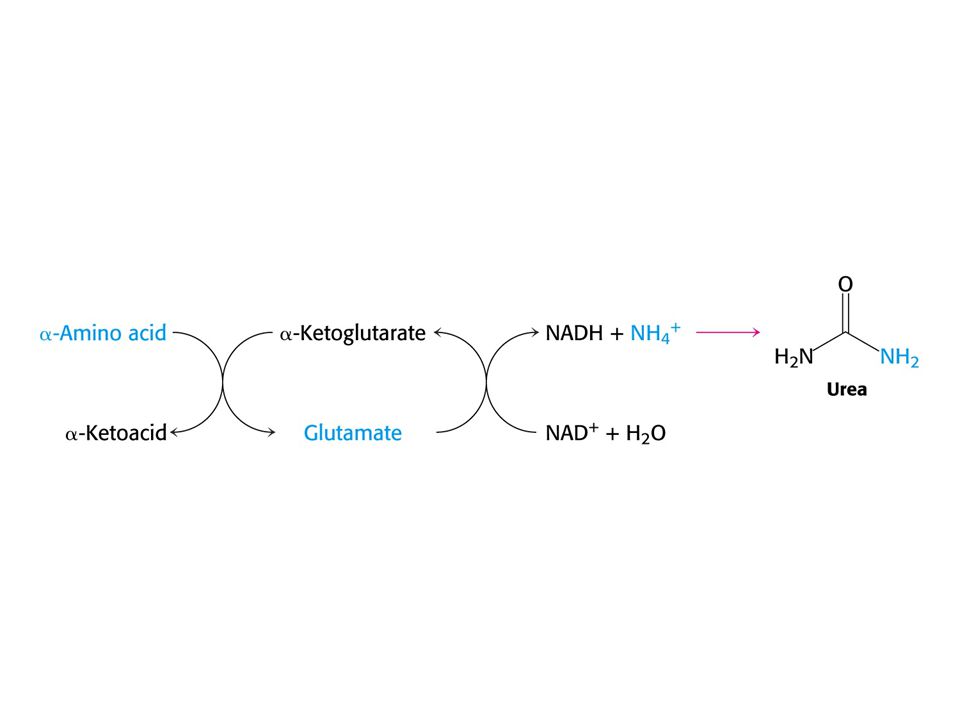
Hlavním místem odbourávání aminokyselin jsou u savců játra. Dusík musí být odstraněn, protože zde nejsou žádné dusíkaté sloučeniny vhodné k zisku energie. Uhlíkatá kostra aminokyselin vstupuje do citrátového cyklu nebo je jeho meziprodukty. Oxidativní deaminace Glu Aminotransferasy: Aspartátaminotransfersa Alaninaminotransferasa

29
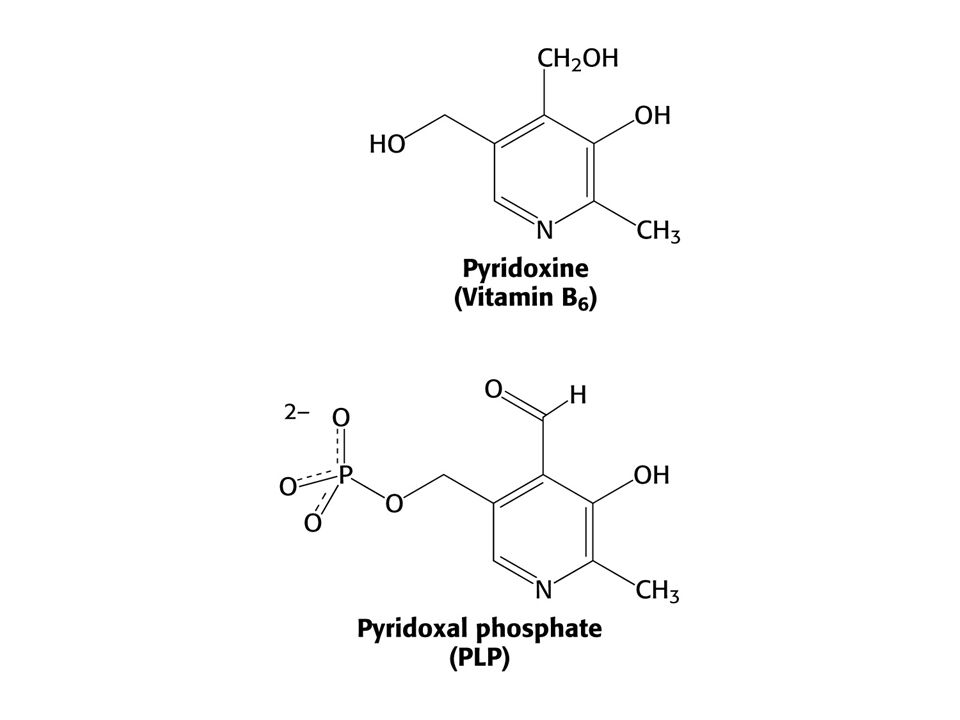
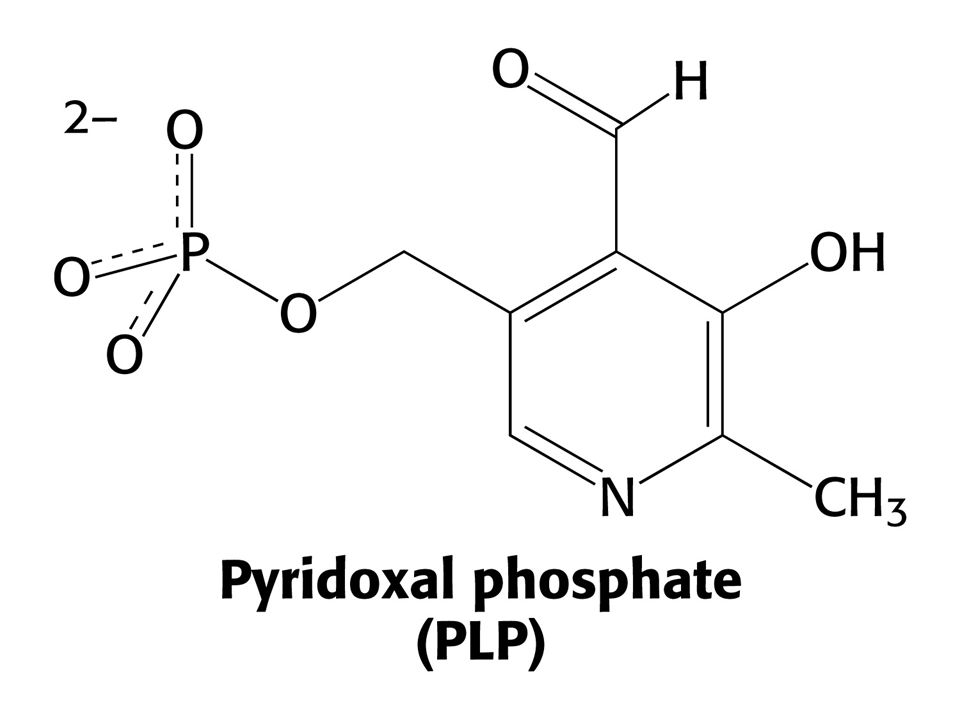
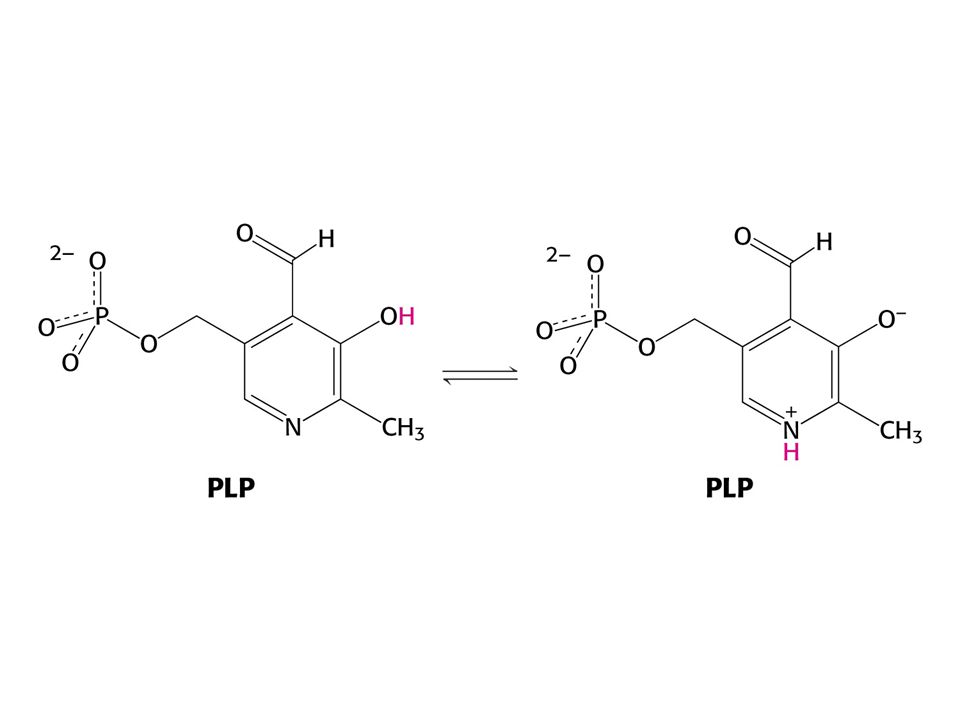
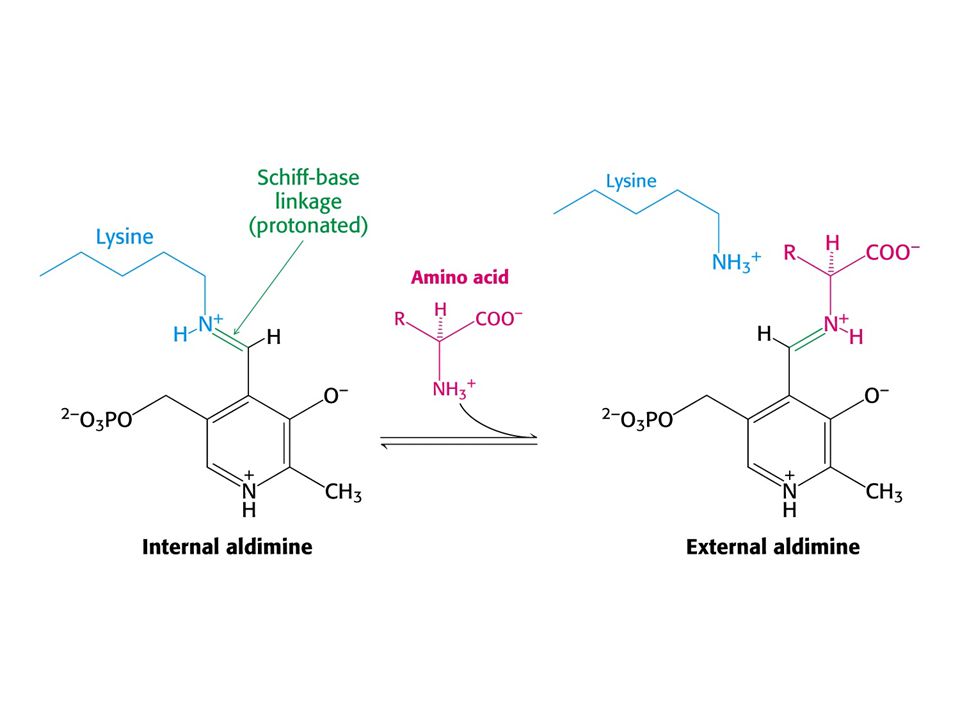
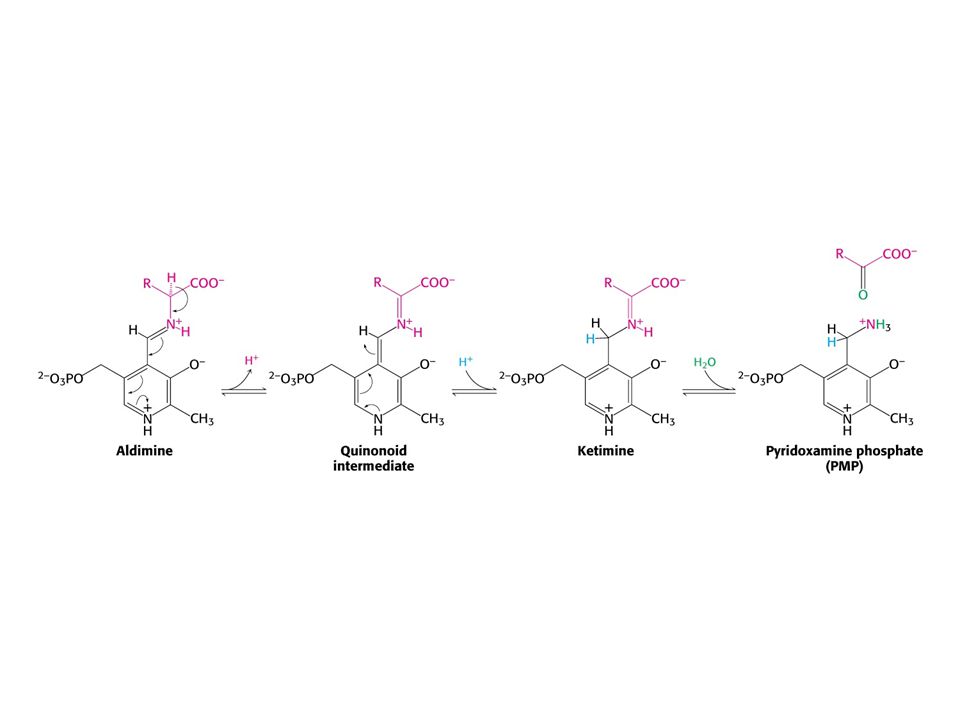
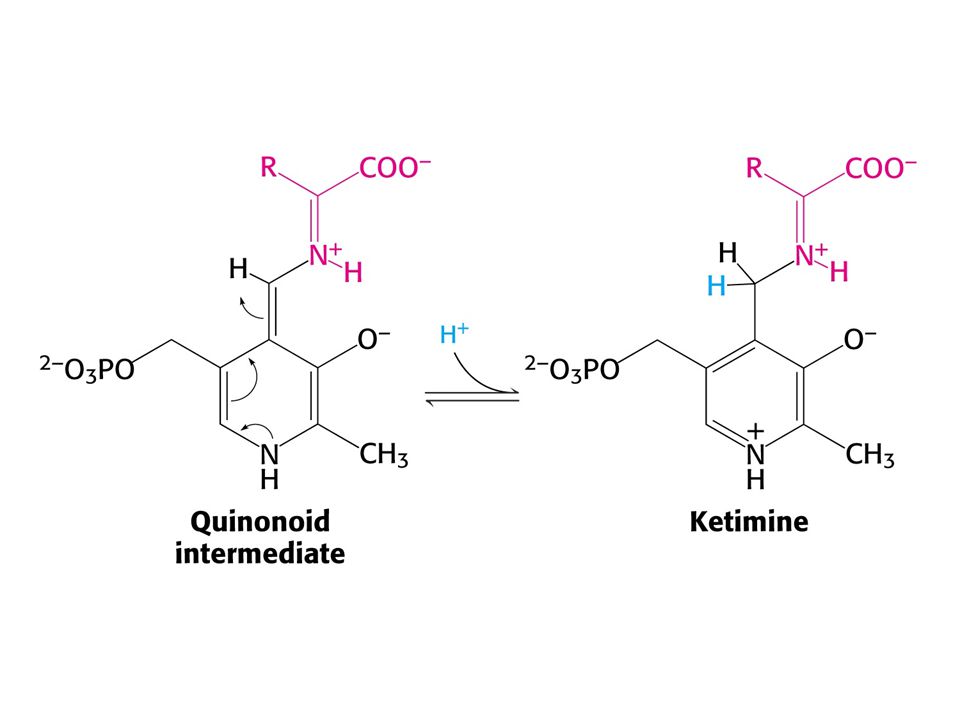
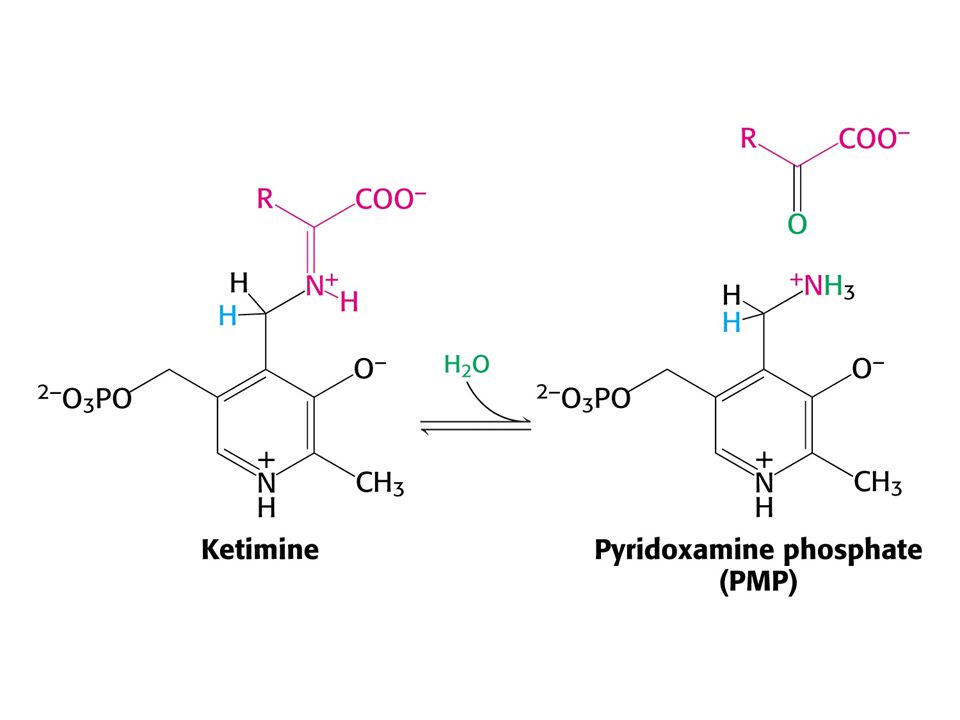
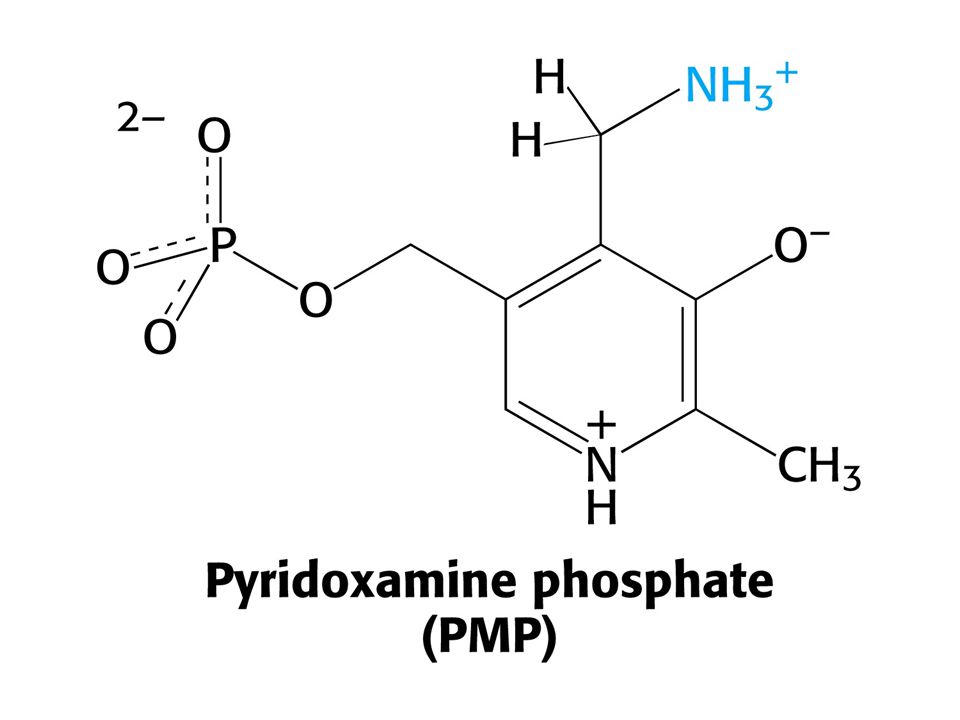
AMINOTRANSFERASOVÁ REAKCE (účast PLP jako prosthetické skupiny)
")
36
Aspartátaminotranferasa

37
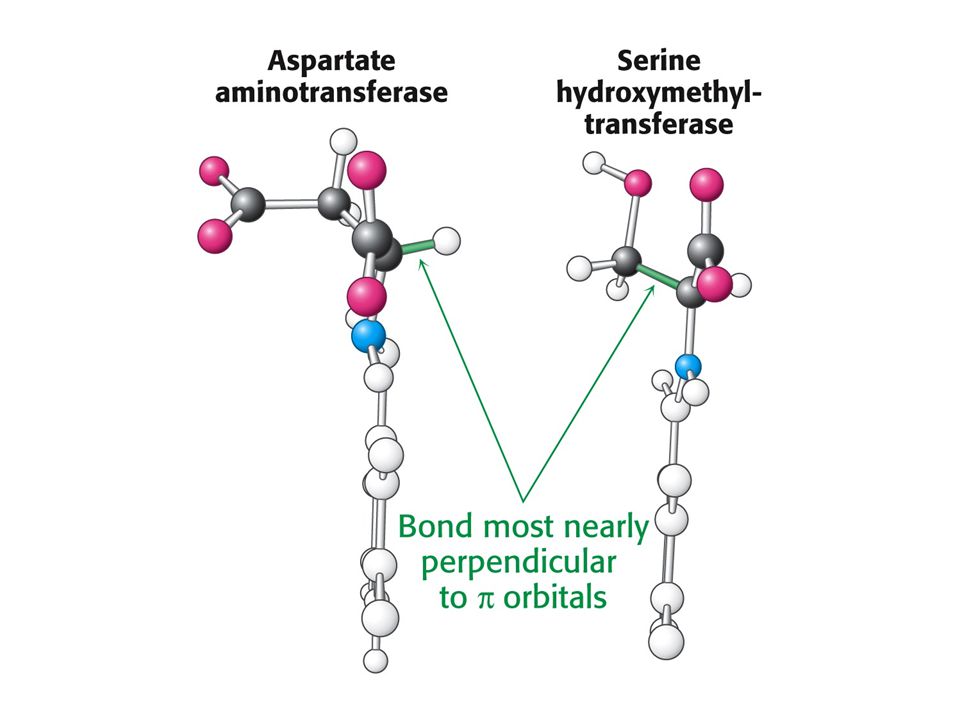
PLP enzymy labilizují jednu ze tří vazeb
PLP enzymy labilizují jednu ze tří vazeb. Vazba a je labilizována aminotransferasami, vazba b dekarboxylasami a vazba c aldolasami.

38
Stereochemický efekt. Vazba rovnoběžná s p orbitalem PLP se nejsnáze štěpí.

40
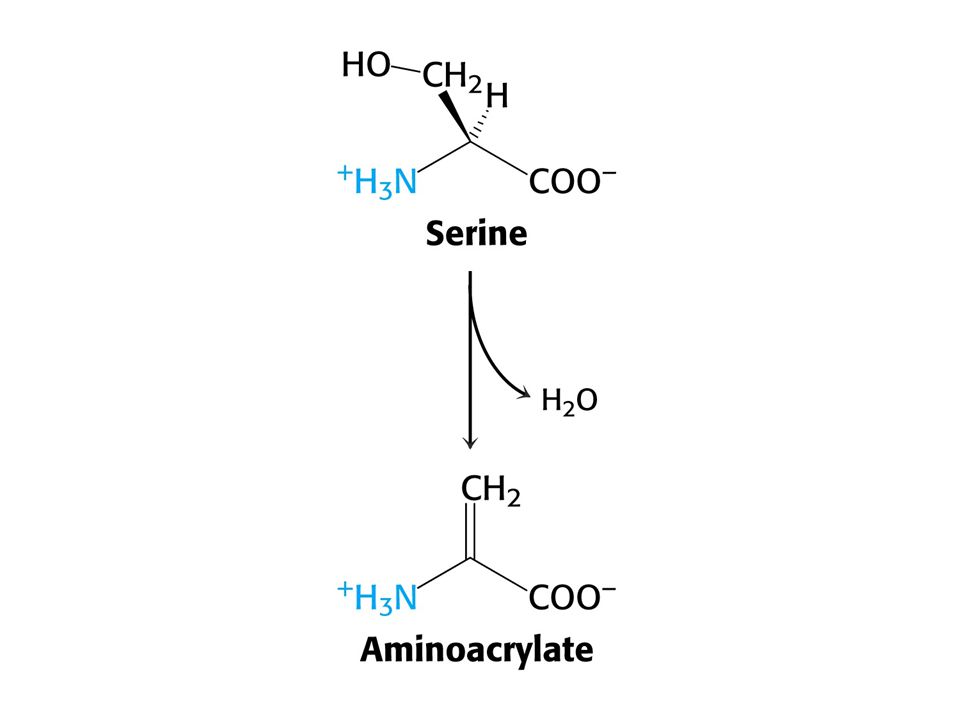
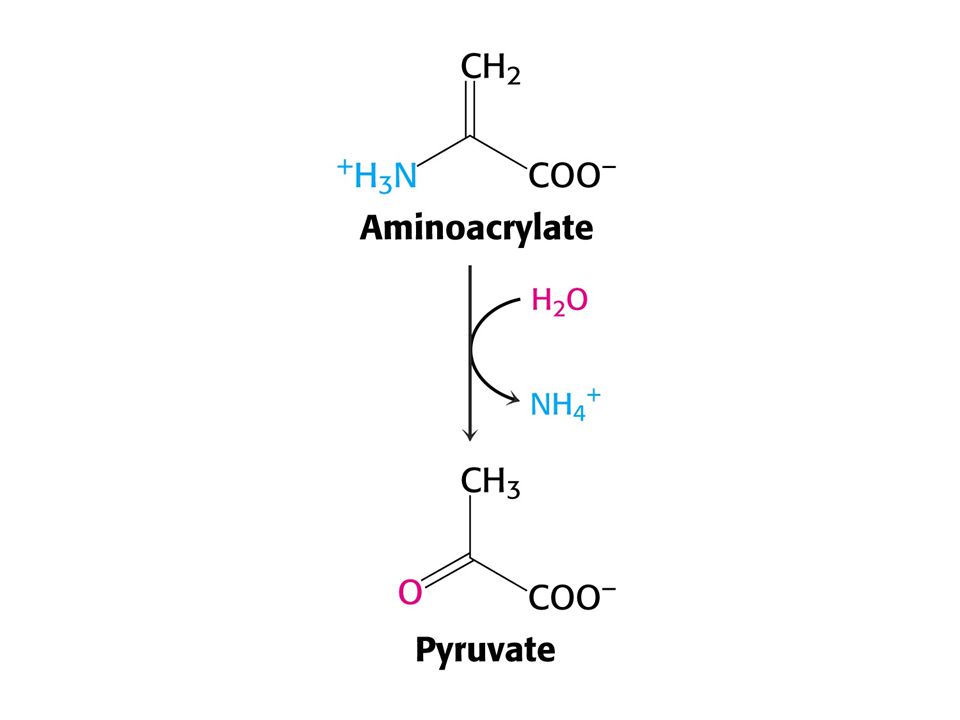
Serin = pyruvát + NH4+ (serindehydratasa-PLP)
Aminokyseliny Ser a Thr mohou být přímo deaminovány PLP jako prosthetická skupina u obou enzymů. Serin = pyruvát + NH4+ (serindehydratasa-PLP) Threonin = a-oxobutyrát + NH4+ (threonindehydratasa - PLP)
 Threonin = a-oxobutyrát + NH4+ (threonindehydratasa - PLP)")
44
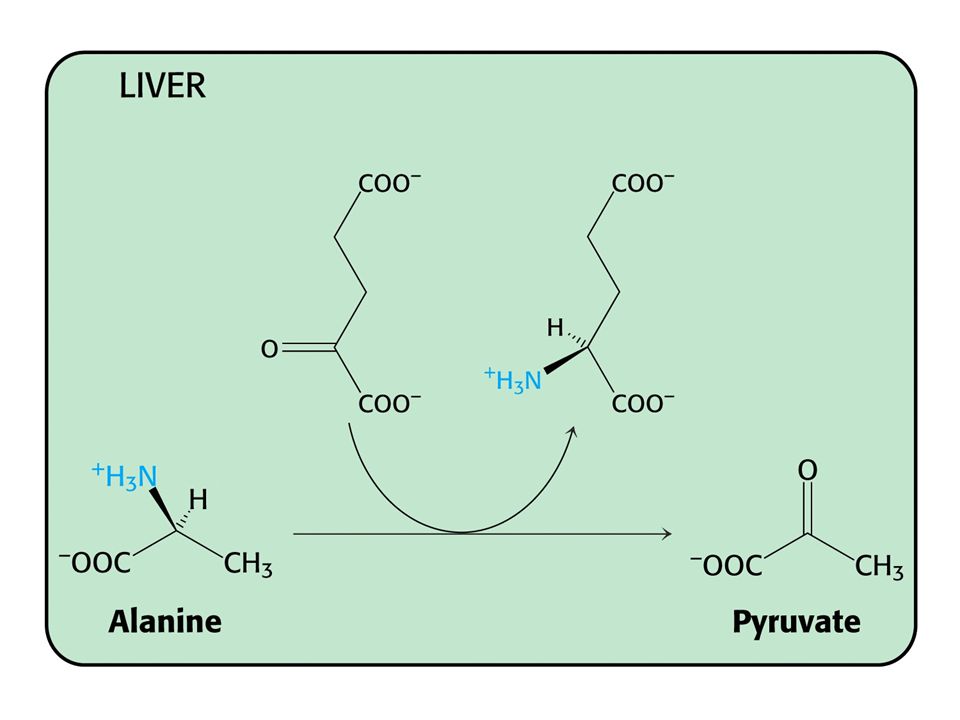
Alaninový cyklus. Transport dusíku ze svalů do jater
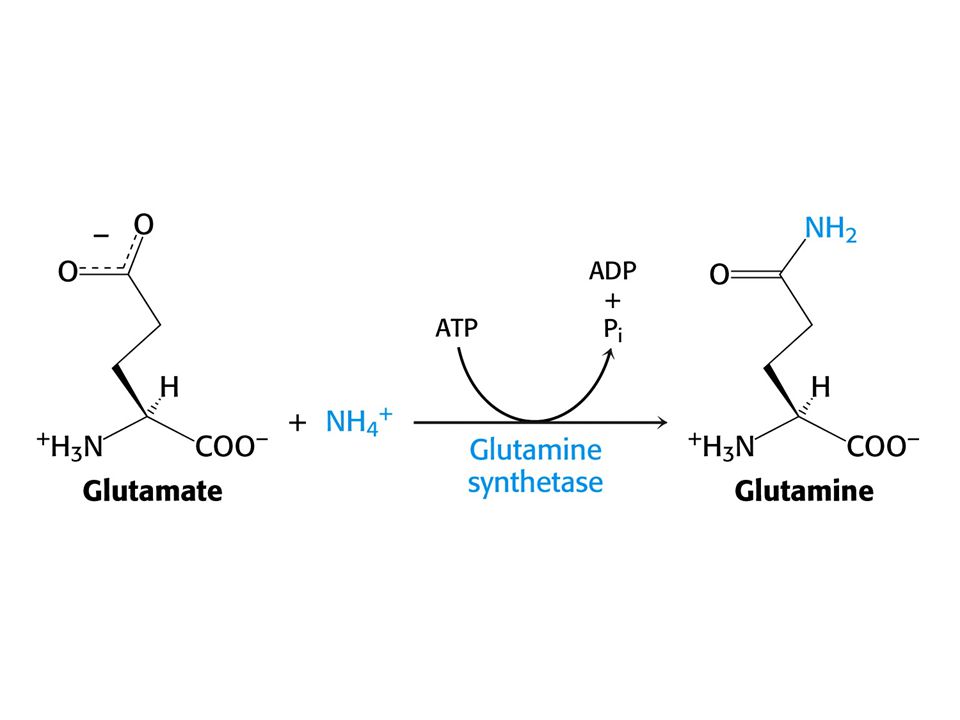
Alaninový cyklus. Transport dusíku ze svalů do jater. Dusík může být také transportován jako Gln. NH4+ + Glu + ATP glutaminsynthetasa Gln + ADP + Pi

47
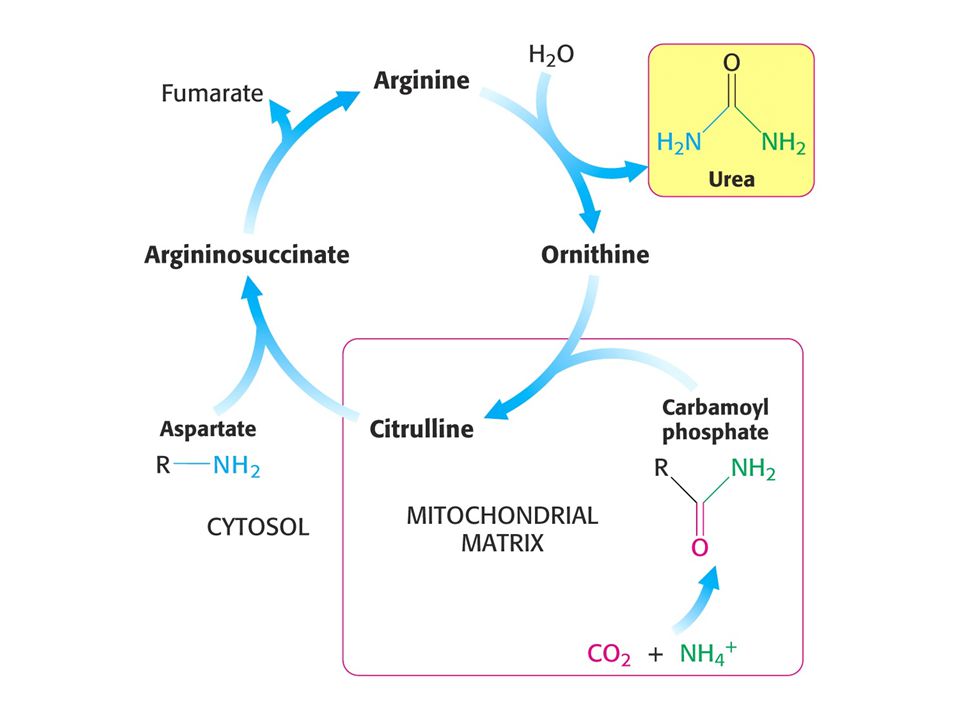
Amoniak je převáděn na močovinu u většiny suchozemských obratlovců
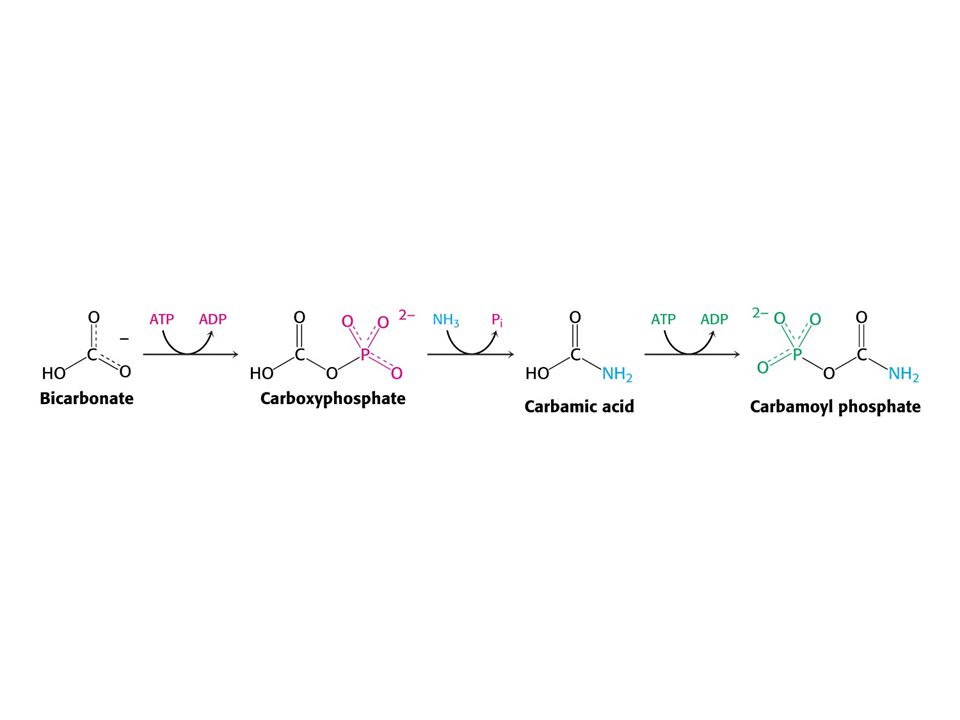
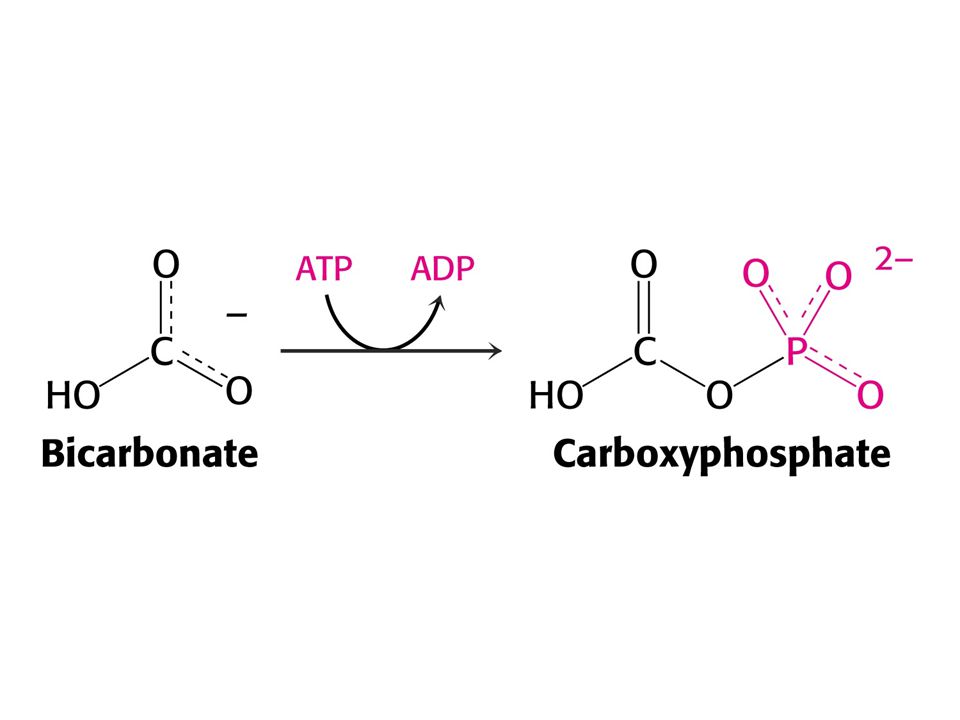
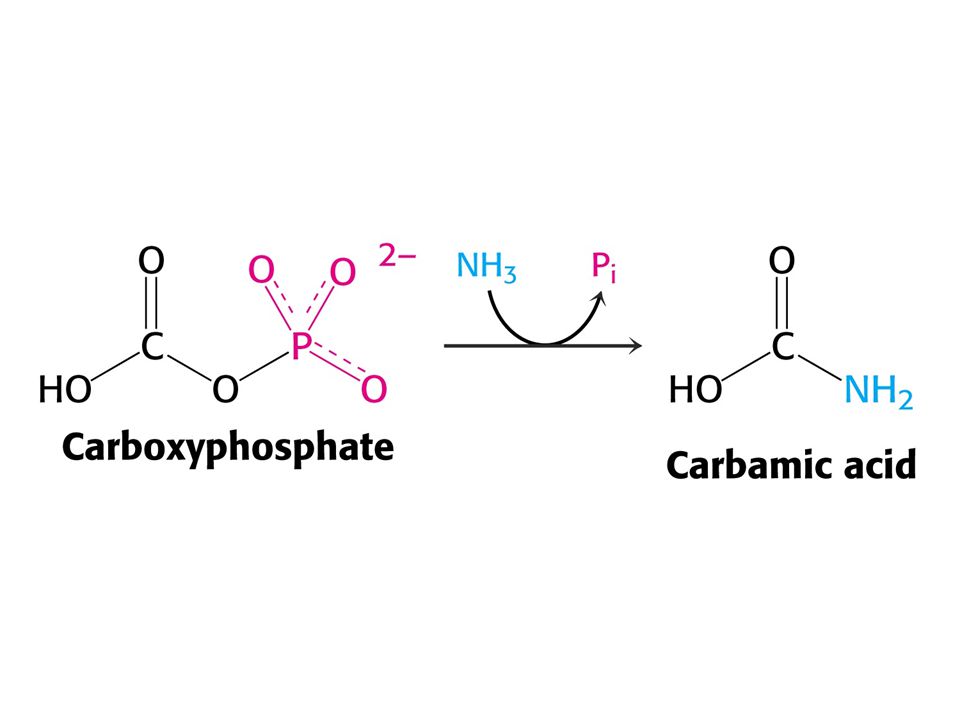
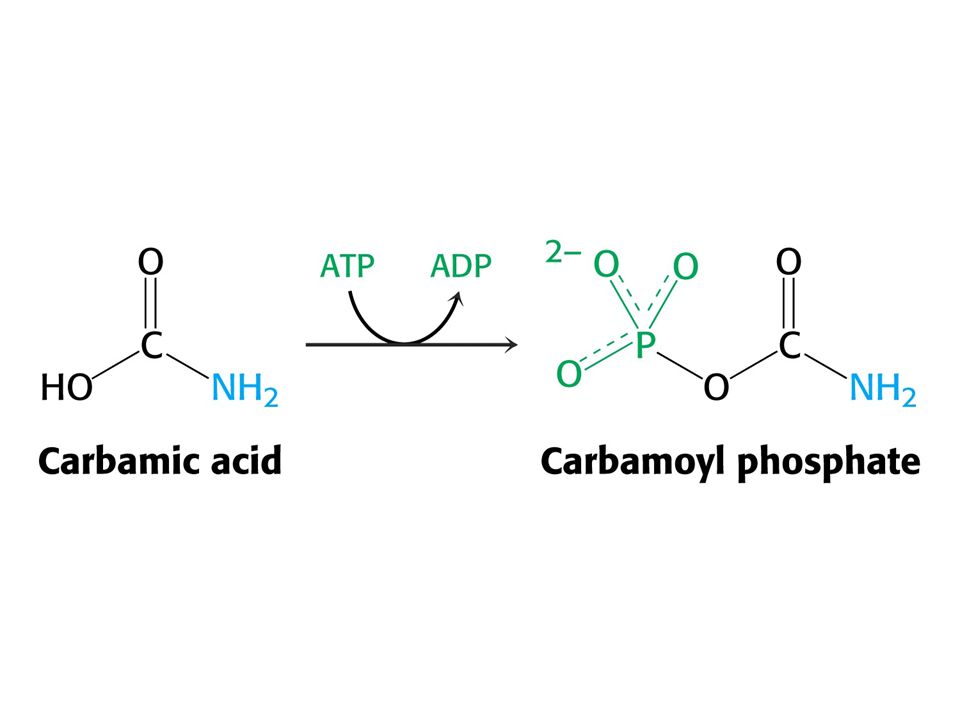
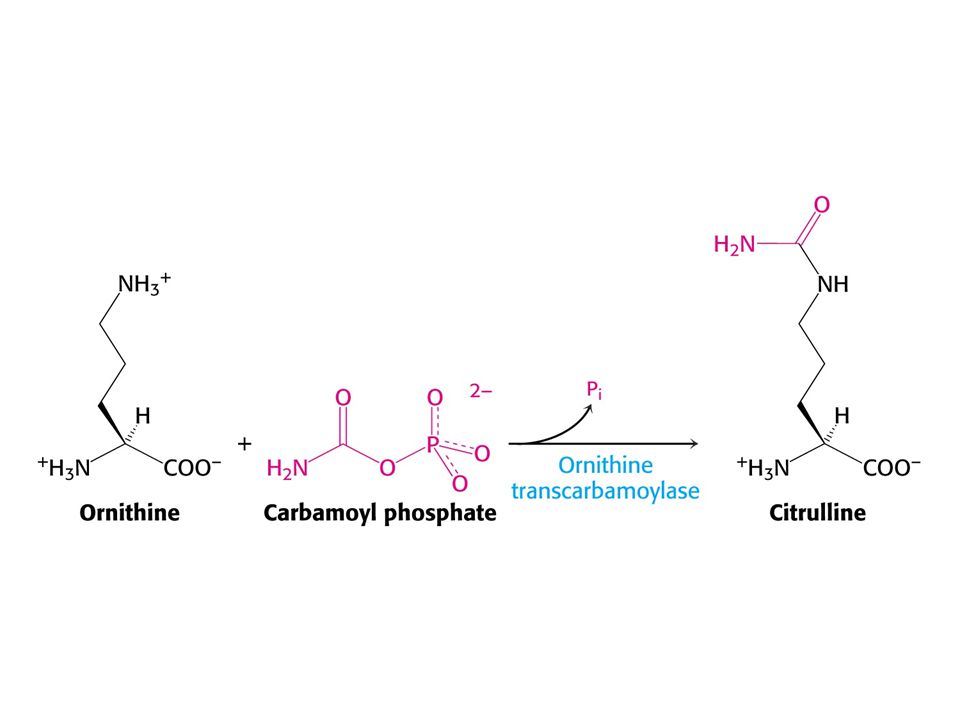
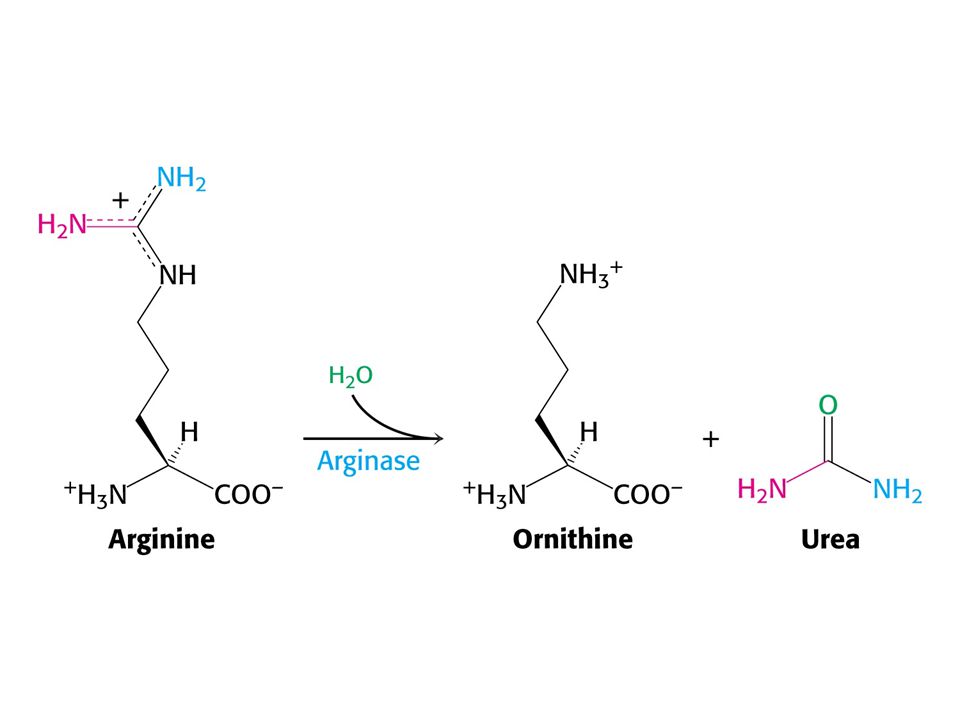
Amoniak je převáděn na močovinu u většiny suchozemských obratlovců. Močovinový (ornithinový) cyklus. Hans Krebs a Kurt Henseleit, 1932 – první metabolický cyklus. První reakcí je tvorba karbamoyl fosfátu z NH4+ , HCO3- a ATP. Enzym: Karbamoylfosfátsynthetasa. Spotřeba dvou ATP činí reakci ireversibilní! Savčí enzym potřebuje k plné aktivitě N-actelyglutamát.
 cyklus. Hans Krebs a Kurt Henseleit, 1932 – první metabolický cyklus. První reakcí je tvorba karbamoyl fosfátu z NH4+ , HCO3- a ATP. Enzym: Karbamoylfosfátsynthetasa. Spotřeba dvou ATP činí reakci ireversibilní! Savčí enzym potřebuje k plné aktivitě. N-actelyglutamát.")
57
Metabolická integrace dusíkového metabolismu.

58
Stechiometrie močovinového cyklu.
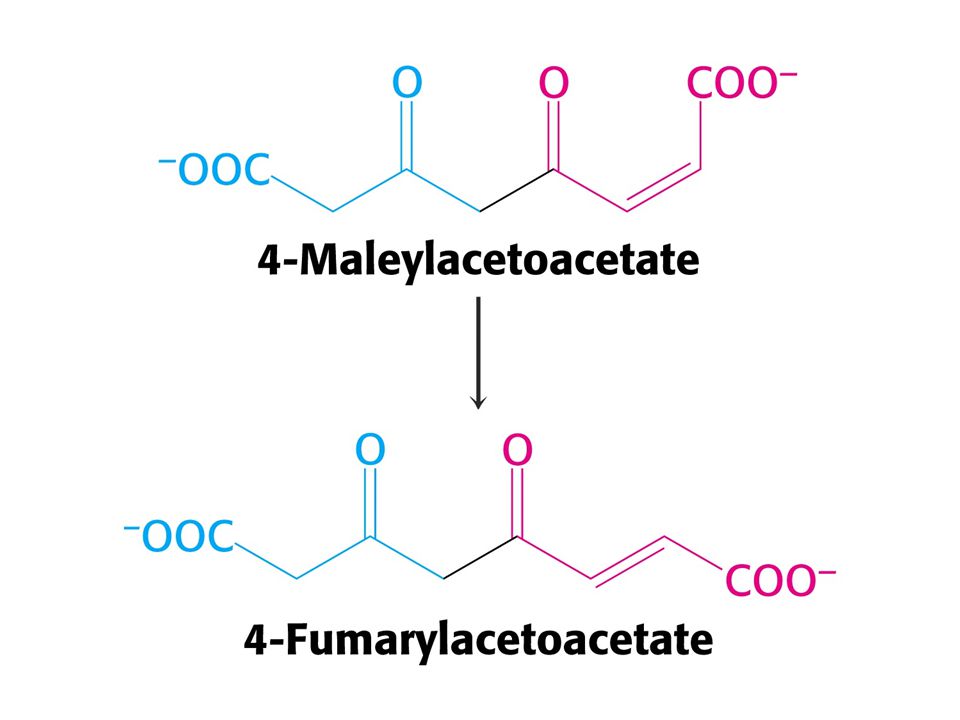
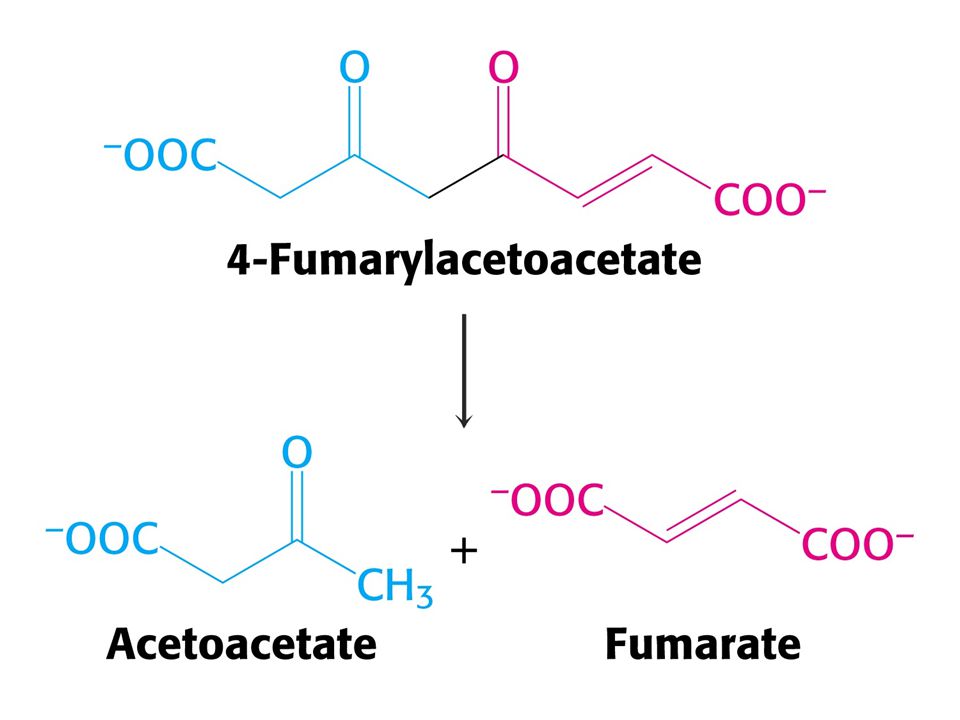
CO2 + NH4+ + 3ATP + Asp + 2 H2O = močovina + 2 ADP + 2 Pi + AMP + PPi + fumarát Spotřebuje se ekvivalent 4 ATP !!! Syntéza fumarátu v močovinovém cyklu je důležitá neboť spojuje močovinový cyklus s cyklem citrátovým. Enzym karbamoylfosfátsynthetasa katalyzuje také syntézu karbamoylfosfátu jako výchozí látku pro syntézu pyrimidinových nukleotidů. U savců existují dva isoenzymy. Při biosyntéze pyrimidinů je zdrojem dusíku Gln a enzym je součástí velkého polypeptidu zvaného CAD (OBSAHUJE TŘI ENZYMY: karbamoylfosfátsynthetasu, aspartáttranskarbamoylasu a dihydroorotasu. Čtyři z pěti enzymů močovinového cyklu byly adaptovány z enzymů syntézy pyrimidinových nukleotidů.

59
Vrozené defekty močovinového cyklu.
Všechny poruchy močovinového cyklu vedou ke zvýšené hladině NH4+ v krvi (hyperamonemie). Proč jsou vysoké hladiny NH4+ toxické ? Zvýšené hladiny glutaminu a glutamátu tvořené při vysoké koncentraci NH4+ vytváří osmotický efekt, který vede k otoku mozku. Vyčerpává se a-oxoglutarát a je tak omezen citrátový cyklus.
. Proč jsou vysoké hladiny NH4+ toxické Zvýšené hladiny glutaminu a glutamátu tvořené při vysoké koncentraci NH4+ vytváří osmotický efekt, který vede k otoku mozku. Vyčerpává se a-oxoglutarát a je tak omezen citrátový cyklus.")
61
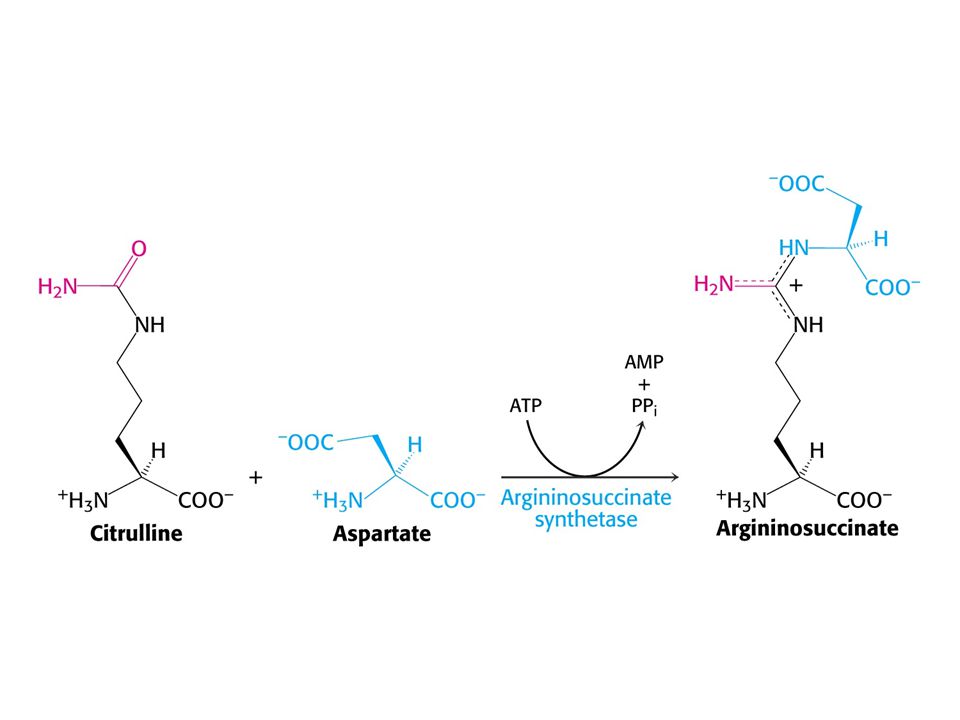
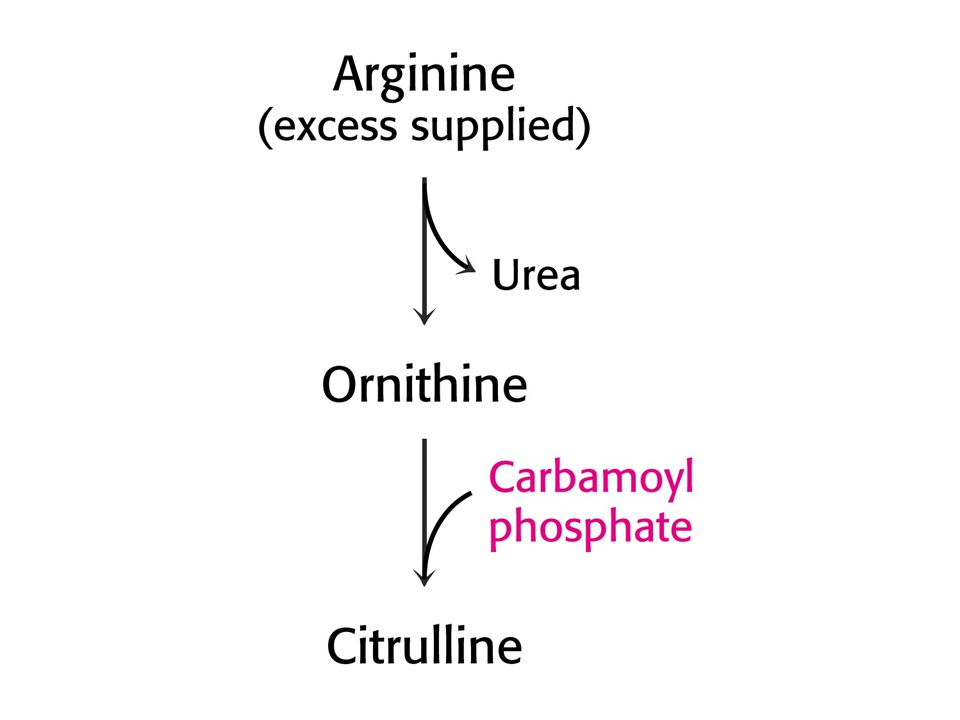
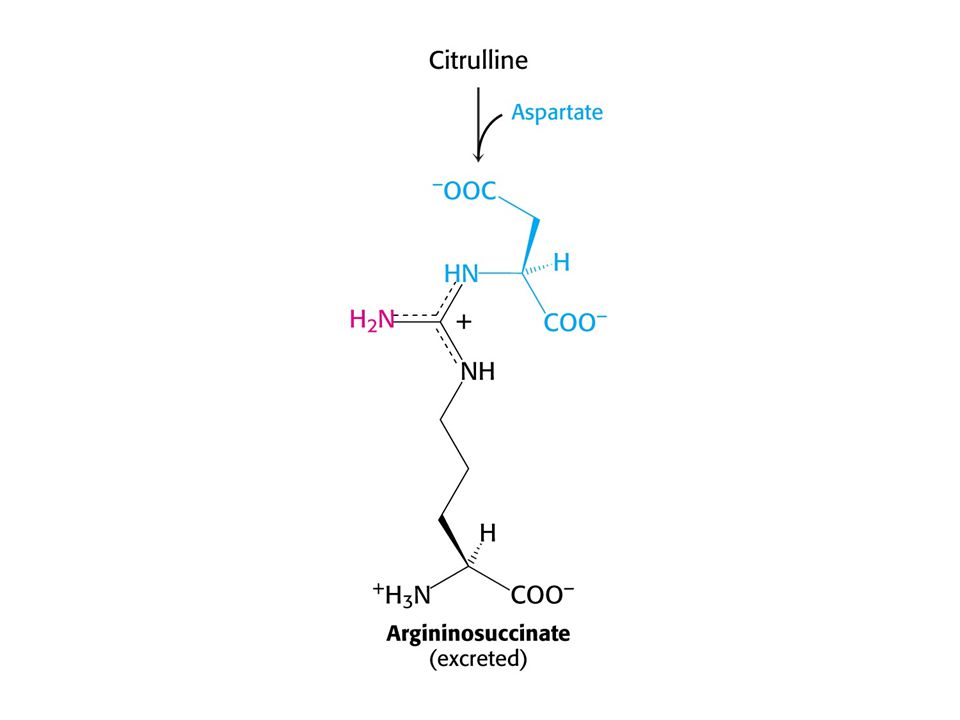
Argininosukcinásová nedostatečnost. Léčí se přídavkem Arg do stravy
Argininosukcinásová nedostatečnost. Léčí se přídavkem Arg do stravy. Dusík se vylučuje ve formě argininosukcinátu

64
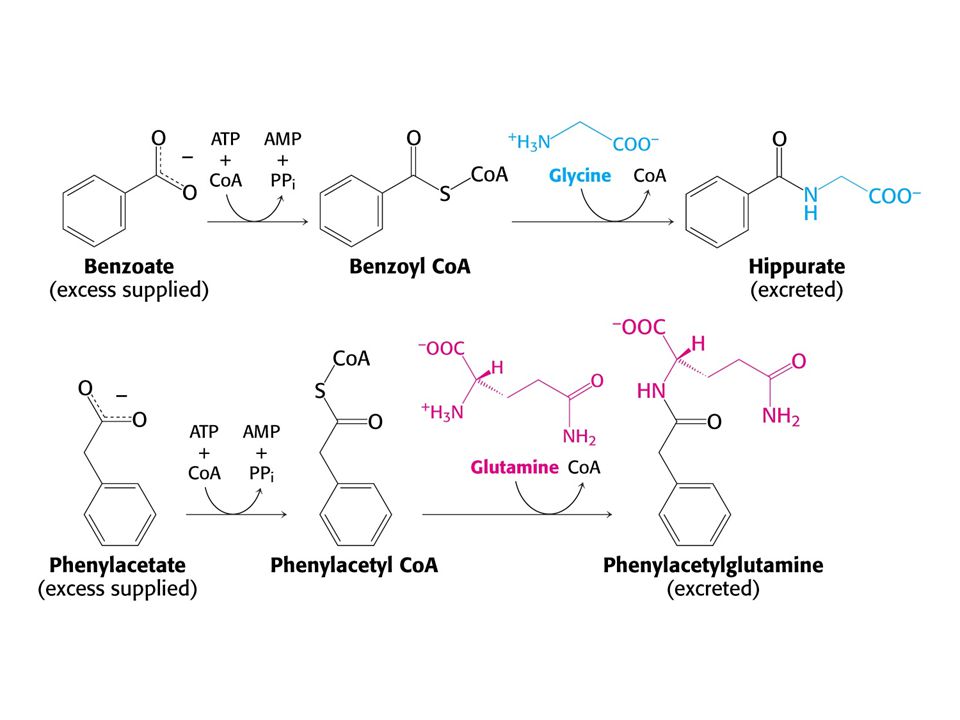
Karbamoylfosfátsynthetasová nedostatečnost.
Nelze podávat citrullin ani argininosukcinát, neboť jejich syntéza je narušena. Nadbytek dusíku se projevuje ve formě Gly a Gln. Podávají se benzoan a fenylacetát. Dusík se vylučuje jako hippurát a fenylacetylglutamin. Je to příklad latentního biochemického procesu, který je aktivován a částečně tak obchází genetický defekt.

68
Močovina není jedinou látkou vedoucí k vyloučení nadbytečného dusíku.
Močovina – ureotelní organismy Amoniak – amonotelní organismy (vodní obratlovci a bezobratlí) Obojí živočichové potřebují k vyloučení dusíku mnoho vody. Močová kyselina a její soli (uráty) – urikotelní organismy (ptáci).
 Obojí živočichové potřebují k vyloučení dusíku mnoho vody. Močová kyselina a její soli (uráty) – urikotelní organismy (ptáci).")
69
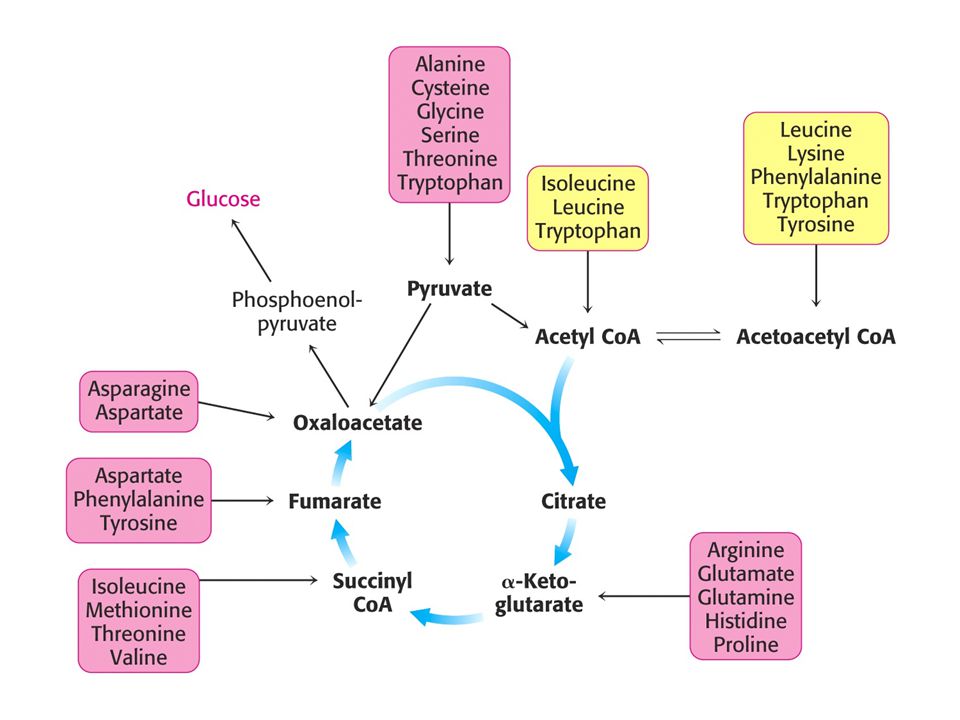
Osud uhlíkaté kostry aminokyselin.
Strategie – převést uhlíkatý skelet aminokyselin na hlavní metabolické meziprodukty, které mohou být převedeny na glukosu nebo plně oxidovány v citrátovém cyklu. Uhlíkatý skelet 20 aminokyselin lze převést na 7 sloučenin: pyruvát, acetylCoA, acetoacetylCoA, a-oxoglutarát, sukcinylCoA, fumarát a oxaloacetát. Aminokyseliny převedené na acetylCoA a acetoacetylCoA se nazývají ketogenní. Lze využít k tvorbě ketolátek nebo mastných kyselin. Aminokyseliny, které lze převést na ostatních pět produktů se nazývají glukogenní. Pouze dvě aminokyseliny jsou výhradně ketogenní: Leu a Lys. Ile, Phe, Trp a Tyr jsou keto i glukogenní. Ostatních 14 je čistě glukogenních.

71
Vstup uhlíkaté kostry aminokyselin do metabolismu.
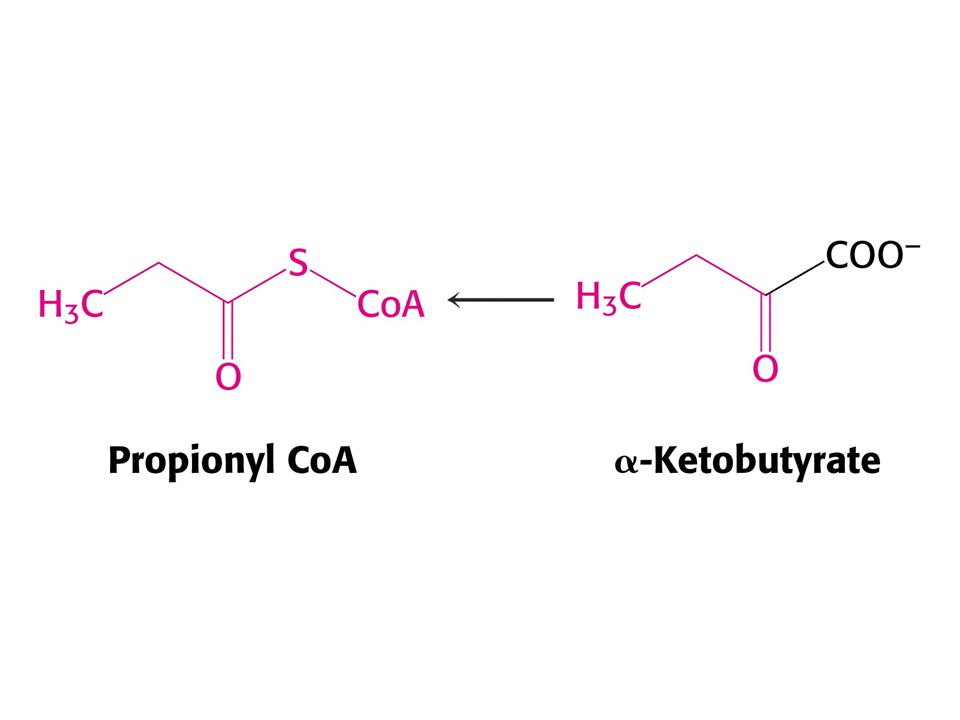
1. Pyruvát 2. Oxaloacetát 3. Alfa-oxoglutarát 4. SukcinylCoA 5. Větvené aminokyseliny vstupují jako acetylCoA, acetoacetát a propionylCoA.

74
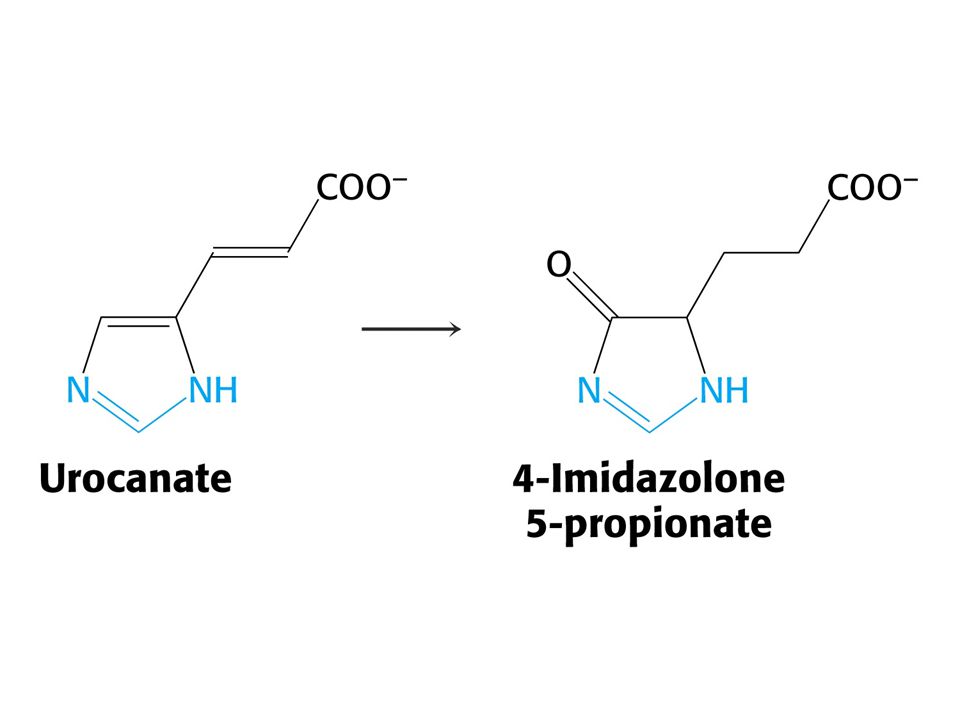
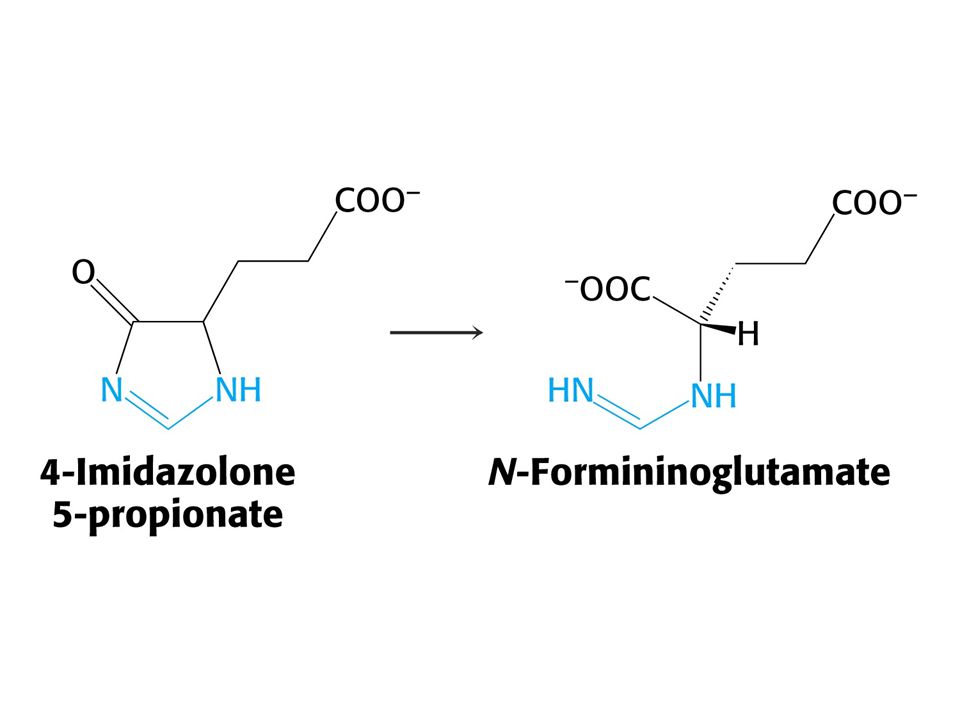
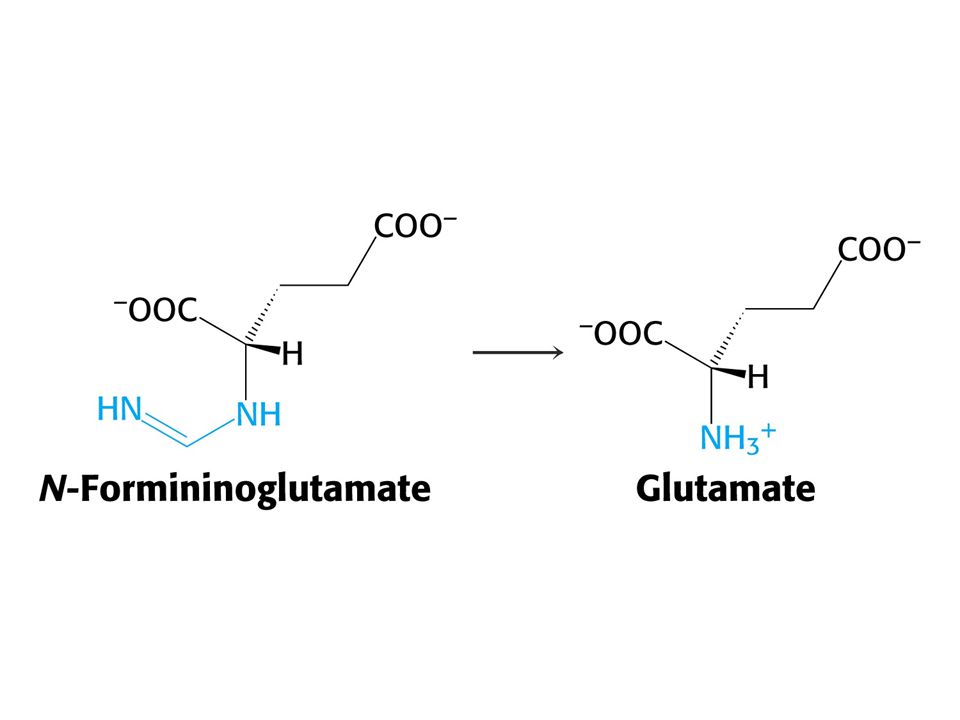
His je odbouráván přes 4-imidazolon-5-propionát na Glu a po transaminaci a-oxoglutarát.

79
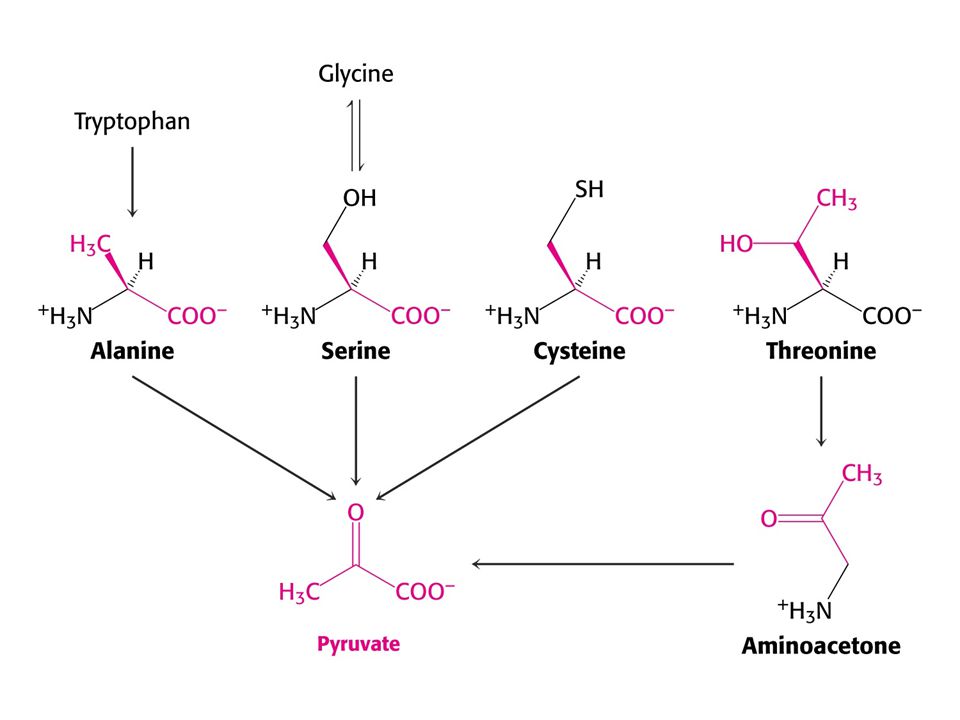
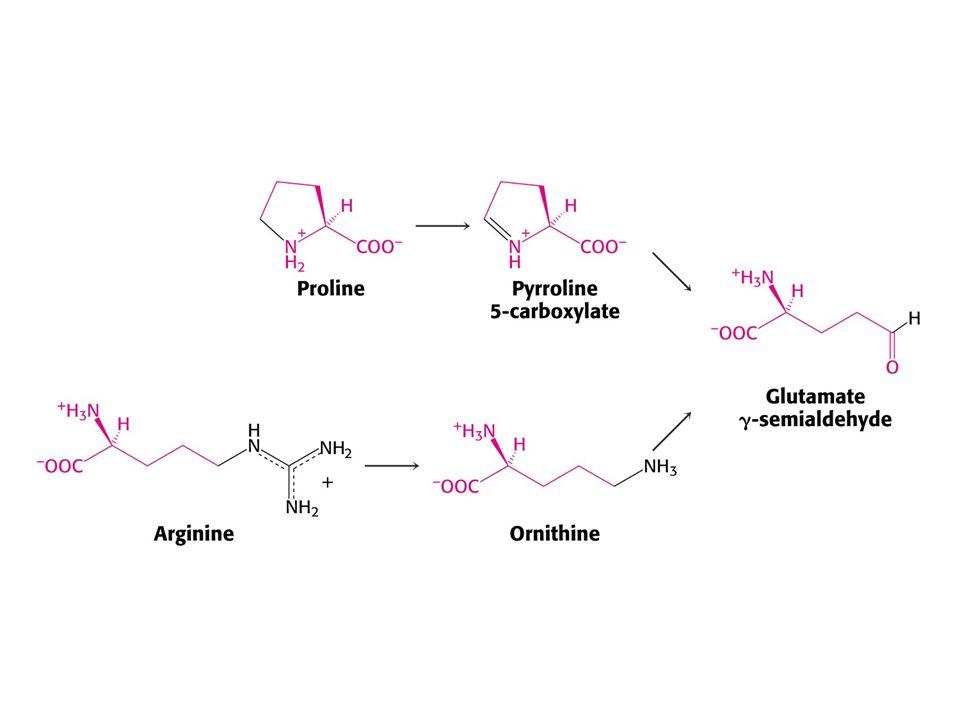
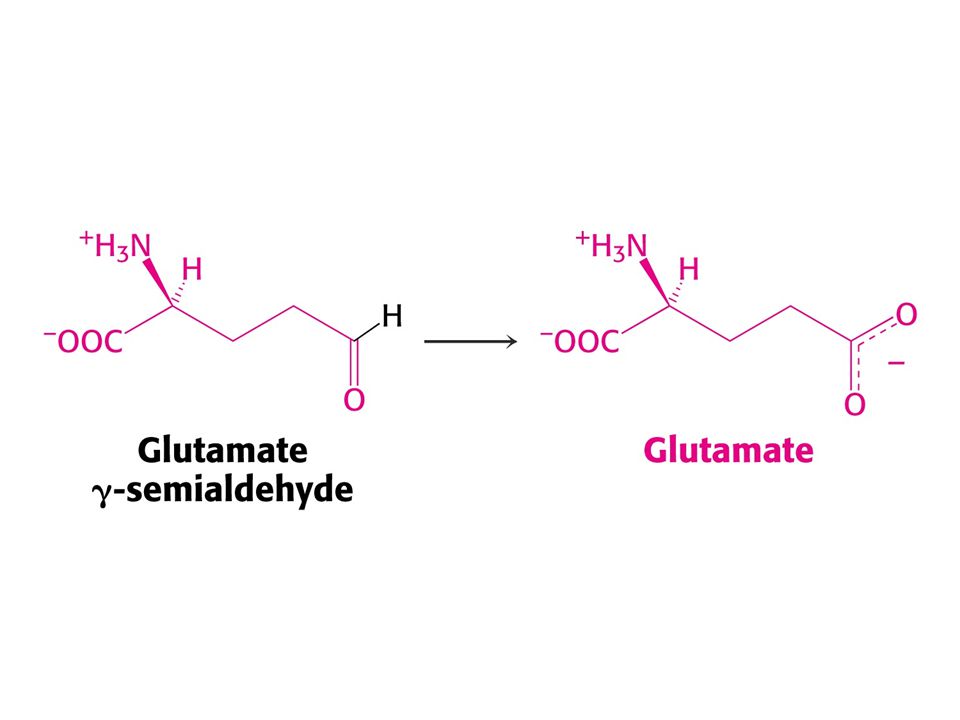
Pro a Arg se odbourávají na Glu

85
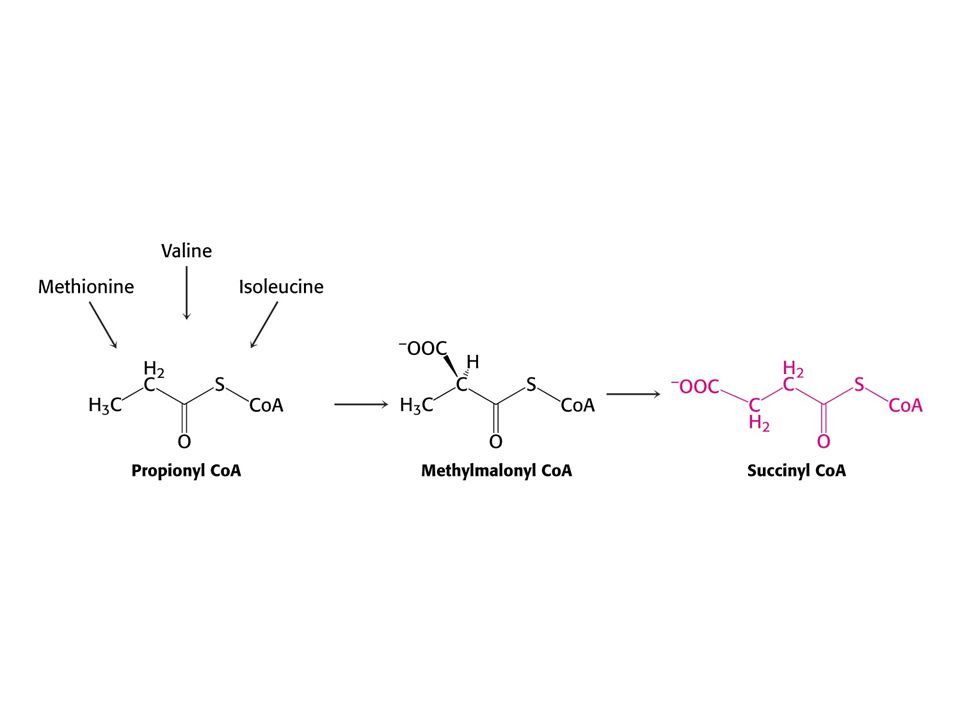
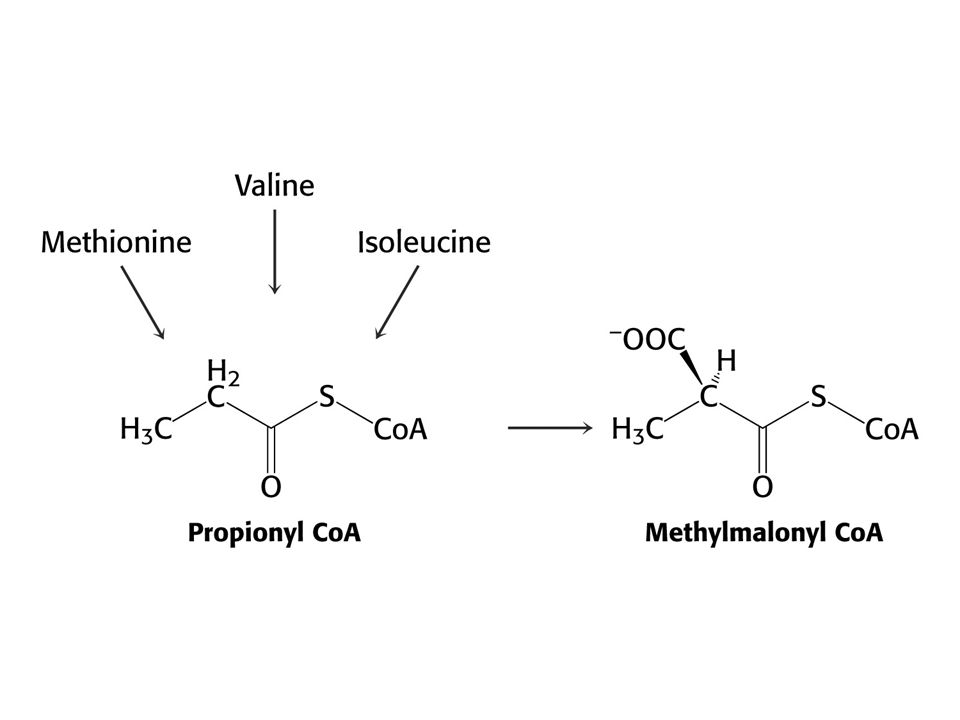
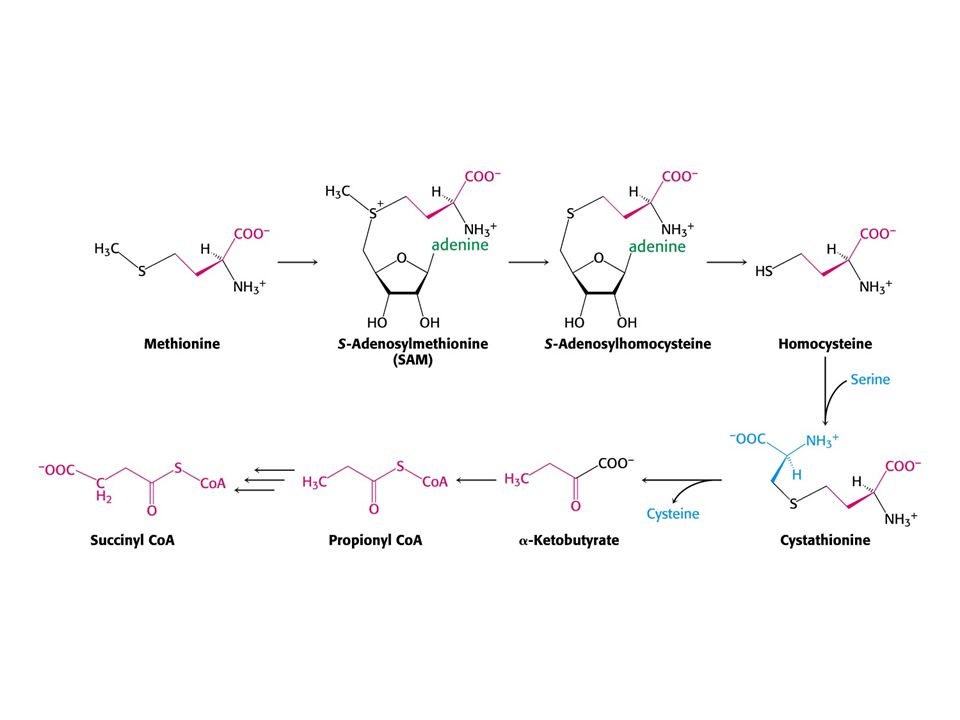
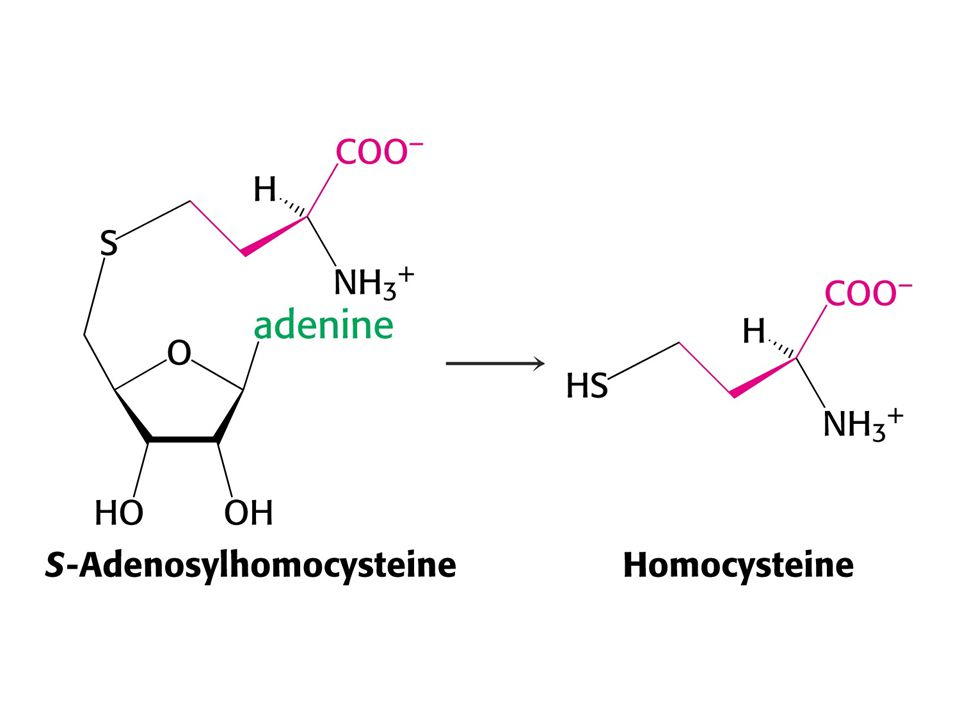
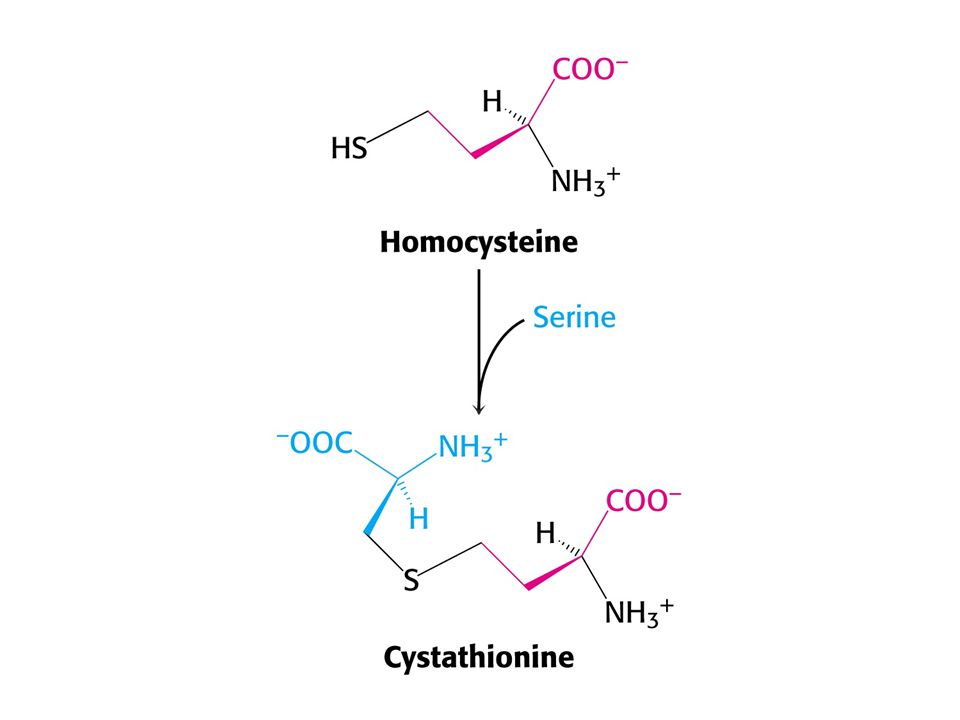
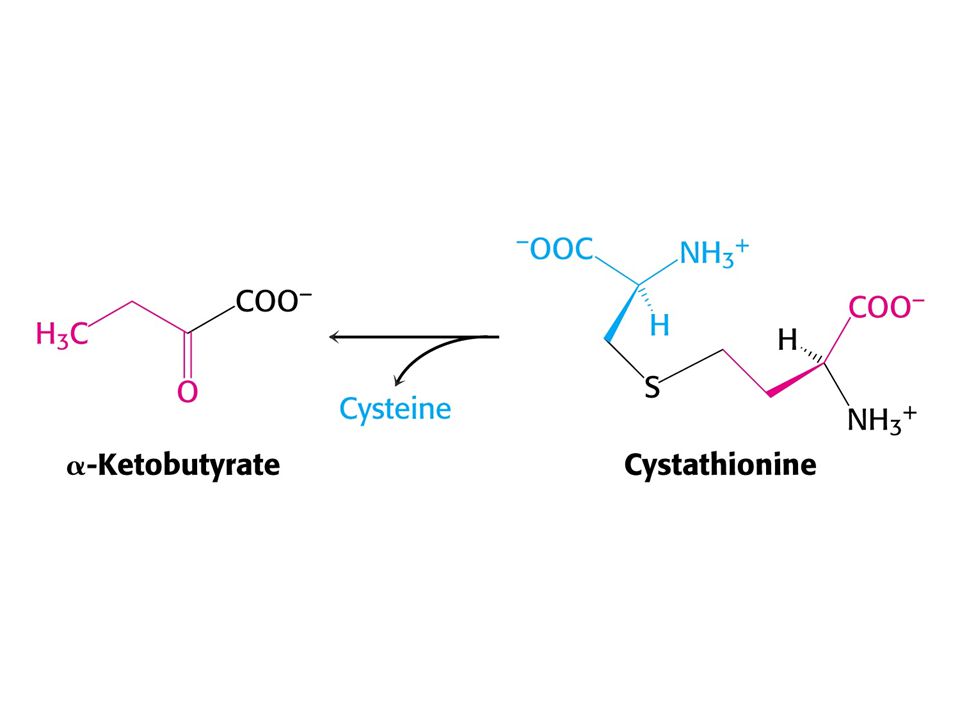
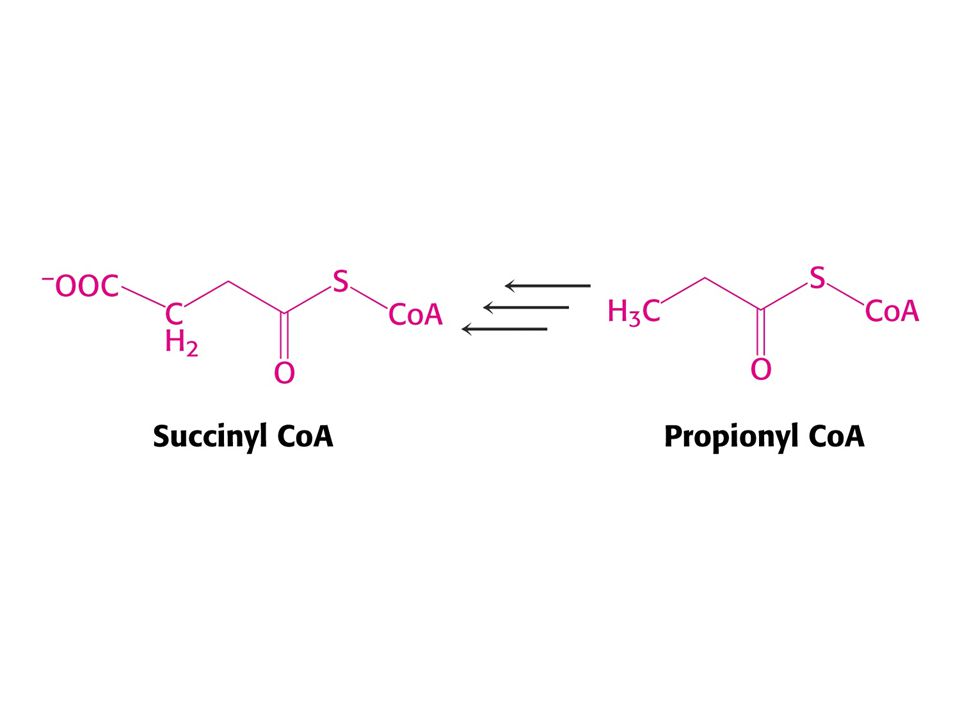
Met je odbouráván v devíti krocích na sukcinylCoA
Met je odbouráván v devíti krocích na sukcinylCoA. V prvním kroku je adenylován za tvorby S-adenosylmethioninu (SAM) což donor methylu v buňce !
 což donor methylu v buňce !")
94
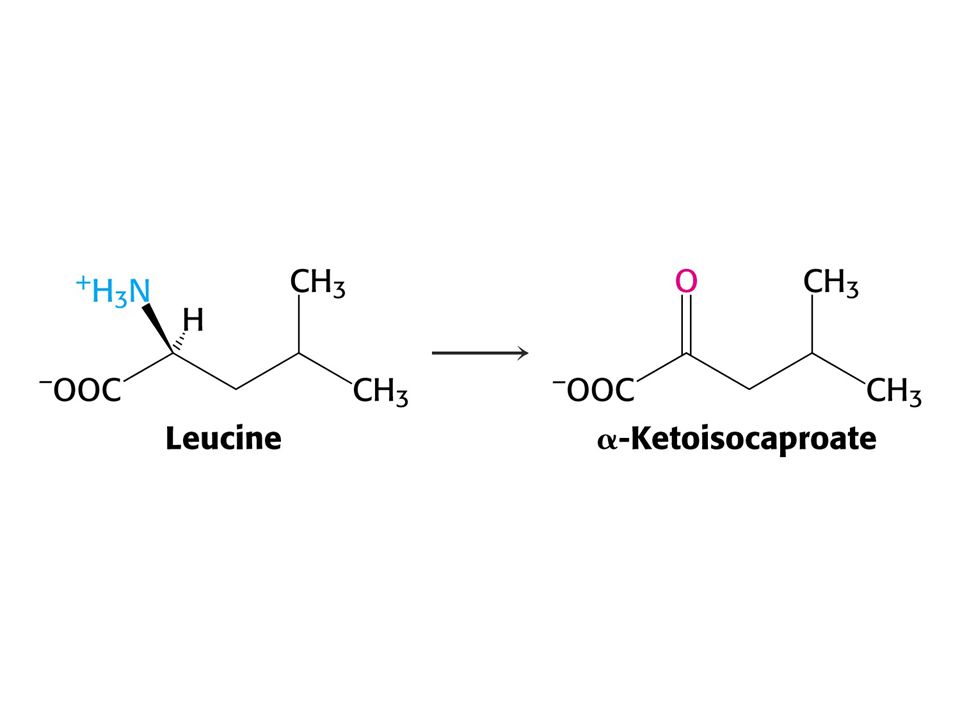
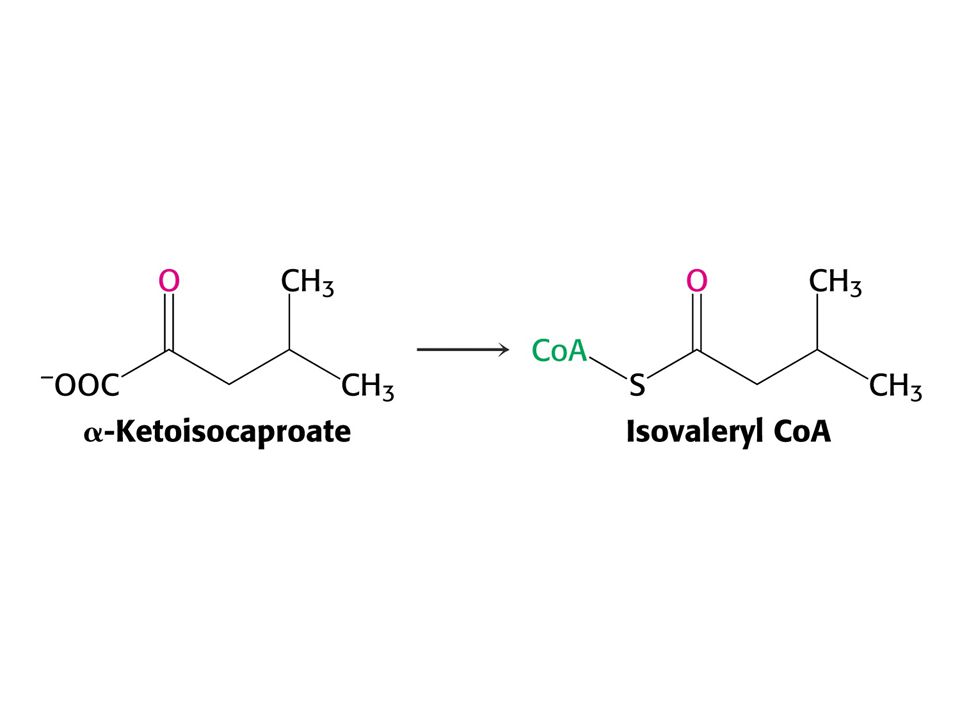
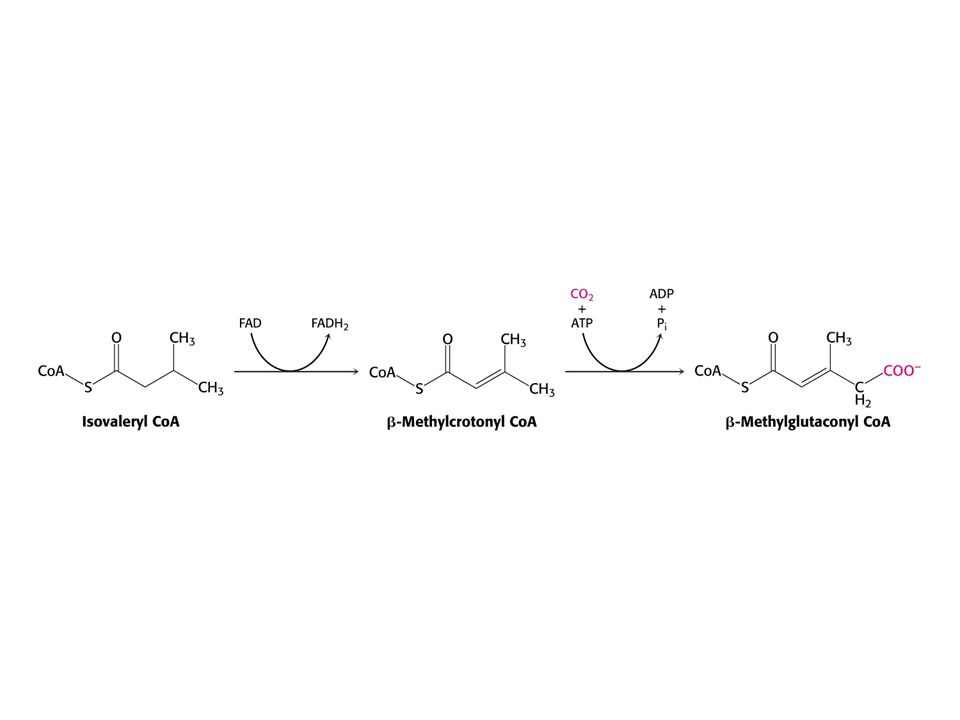
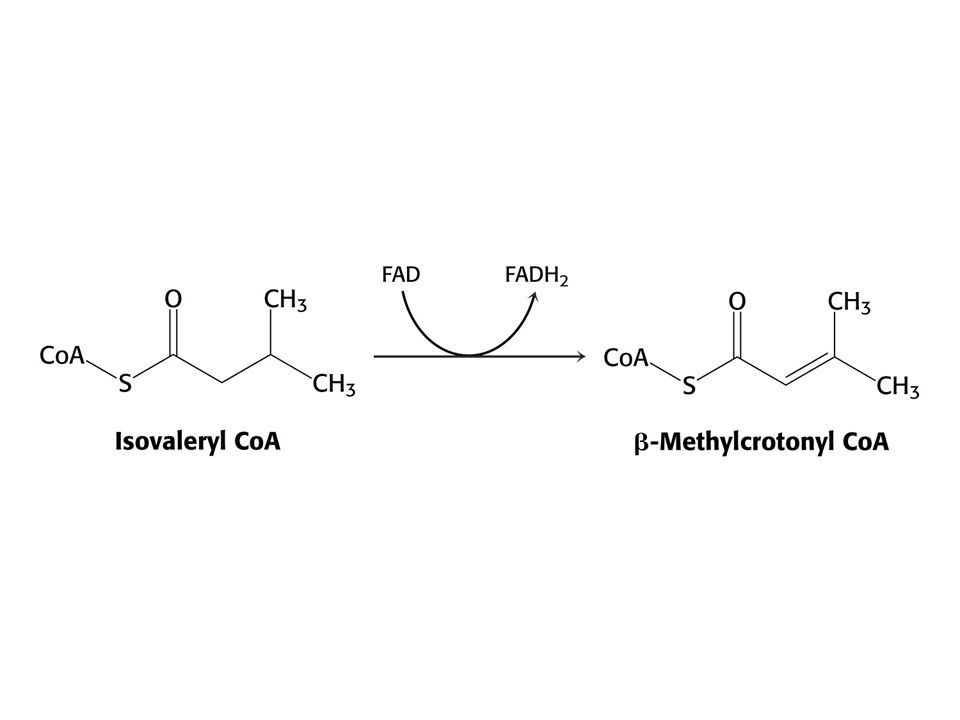
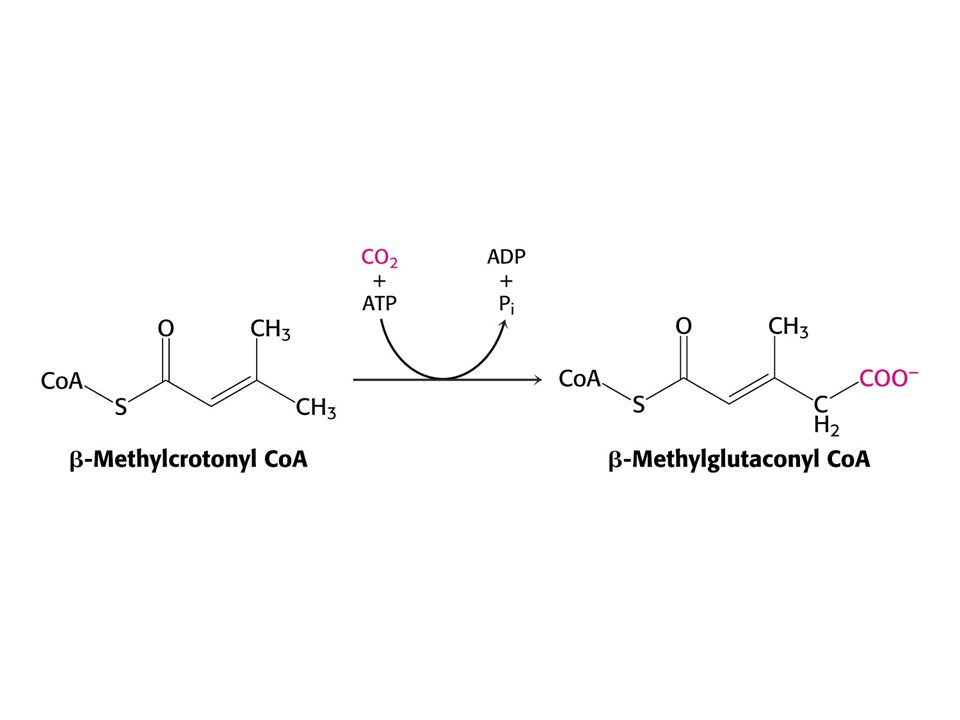
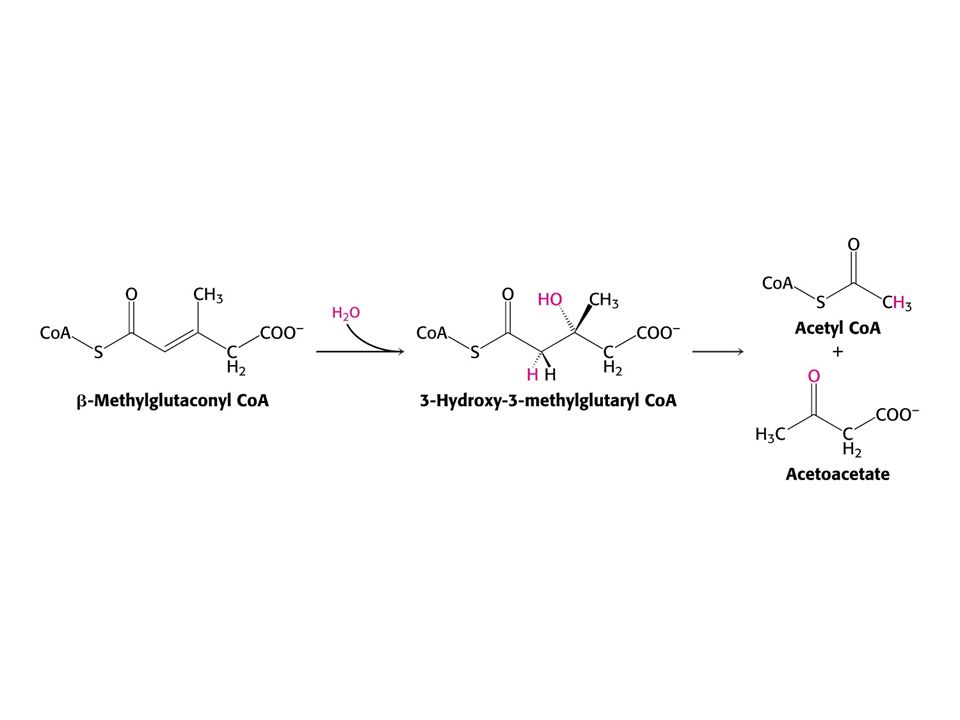
Leu – ketogenní aminokyselina – odbourává se na acetylCoA a acetoacetát.

103
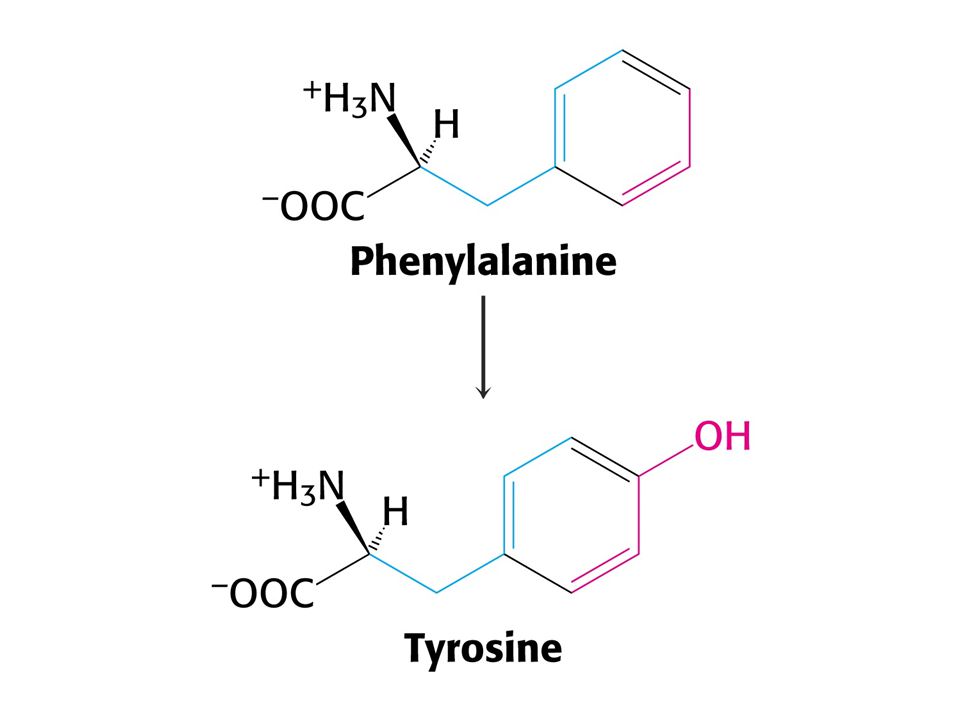
K degradaci aromatických aminokyselin jsou nutné OXYGENASY – molekulární kyslík.Fenylalaninhydroxylasa je monooxygenasa-(mixed-function oxygenase) Jeden atom kyslíku je po reakci v produktu a druhý ve vodě.
 Jeden atom kyslíku je po reakci v produktu a druhý ve vodě.")
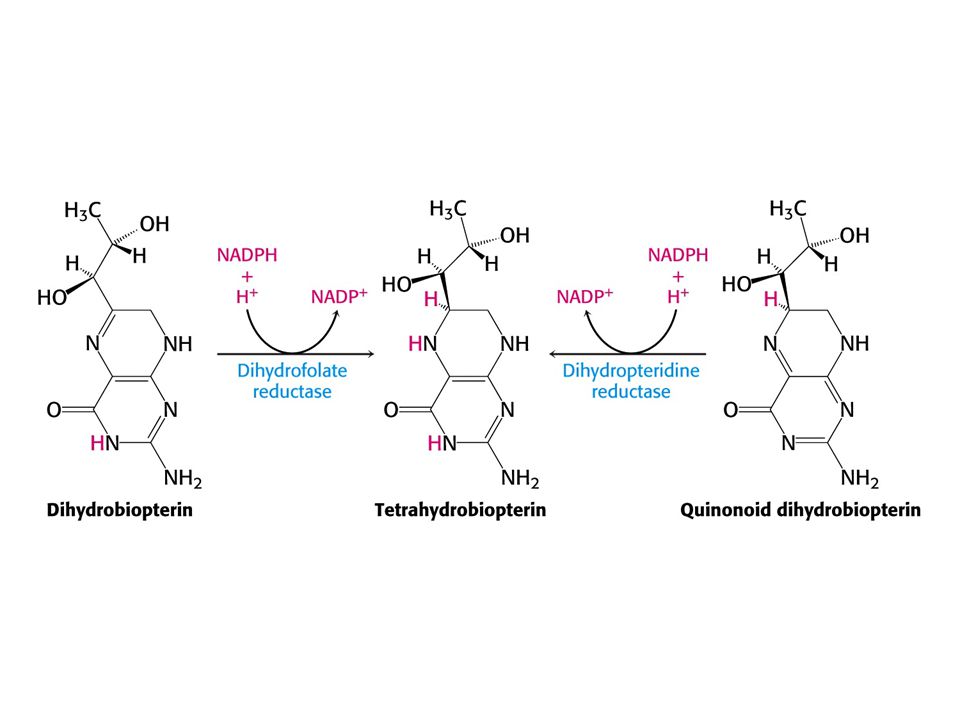
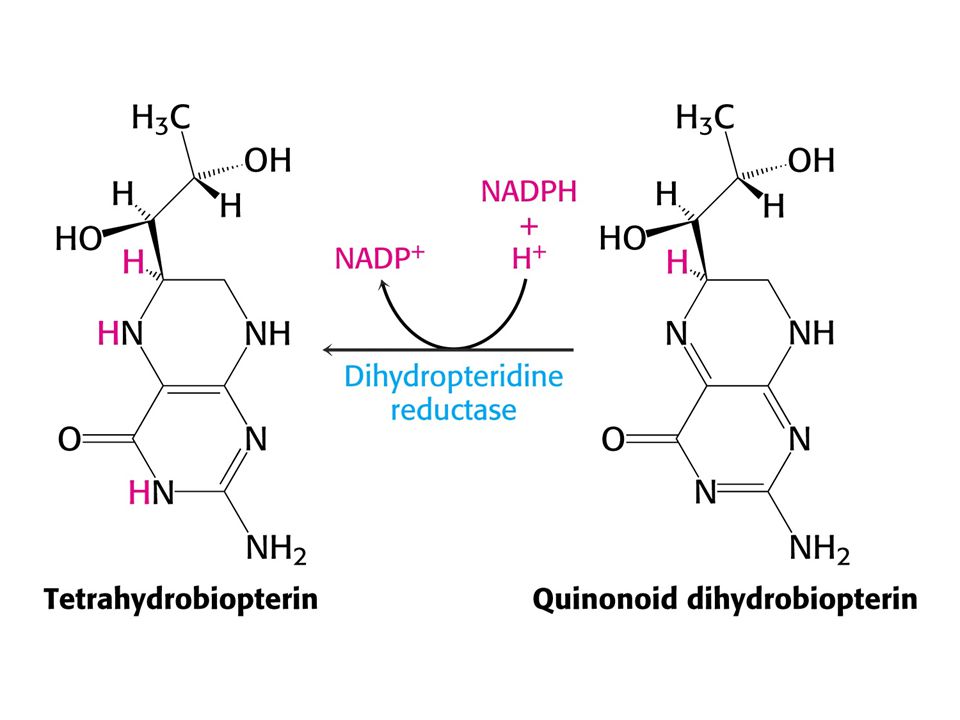
104
Redukčním činidlem je tetrahydrobiopterin, který se tvoří redukcí z kofaktoru biopterinu. Není to vitamin !!

108
Suma reakcí katalyzovaných fenylalaninhydroxylasou a dihydropterinreduktasou:
Fenylalanin + O2 + NADH + H+ = Tyrosin + NAD+ + H2O

110
Další stupně degradace Phe a Tyr začínají transaminací ….

111
Reakce p-hydroxyfenylpyruvátu s O2 za vzniku homogentisátu
Reakce p-hydroxyfenylpyruvátu s O2 za vzniku homogentisátu. Enzym je p–hydroxyfenylhydroxylasa – dioxygenasa. Oba atomy O2 vstupují do produktu.

112
Homogentisátoxygenasa – další dioxygenasa.

115
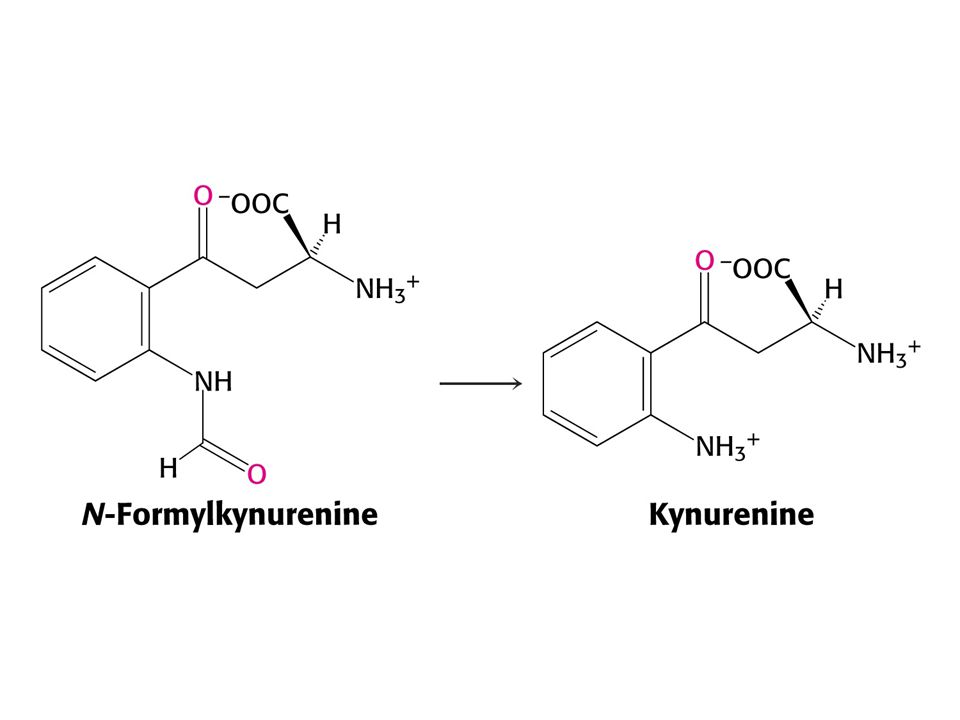
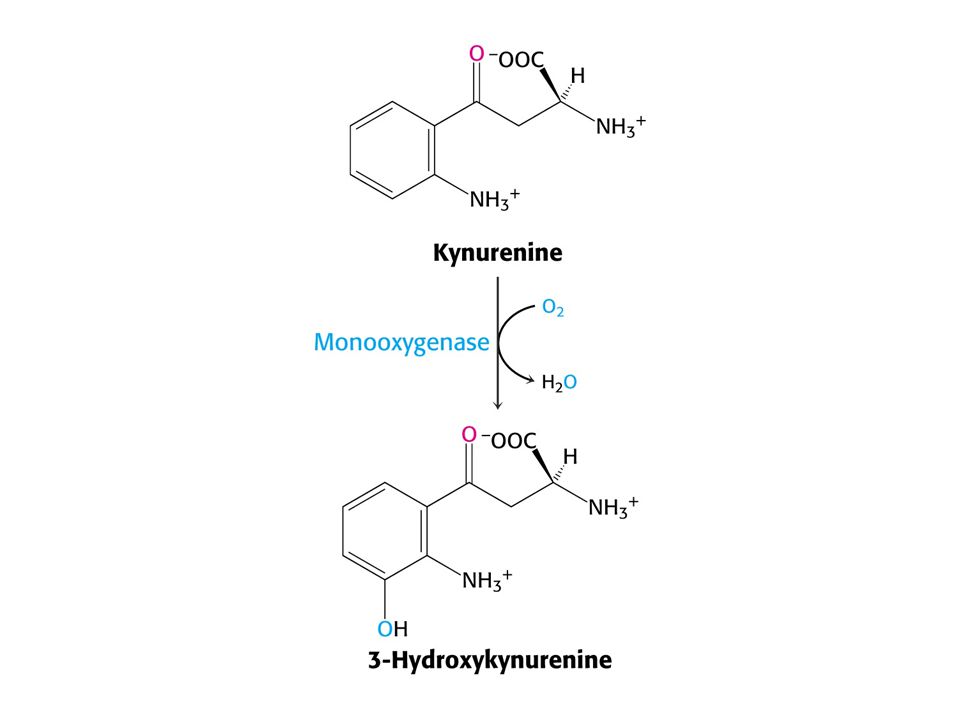
Odbourávání Trp – šestnáct kroků
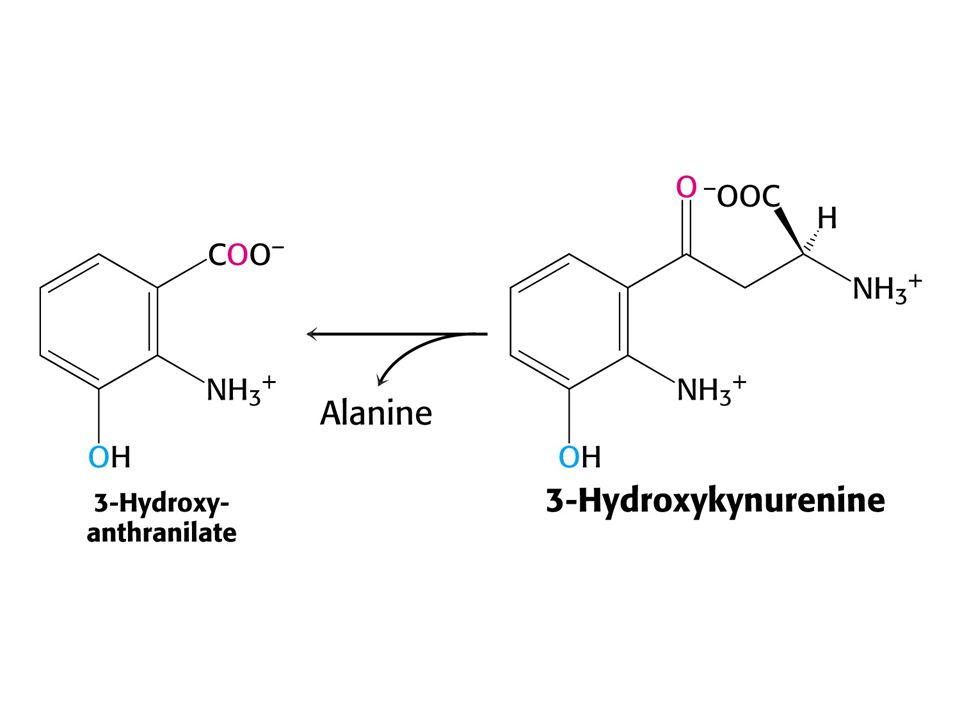
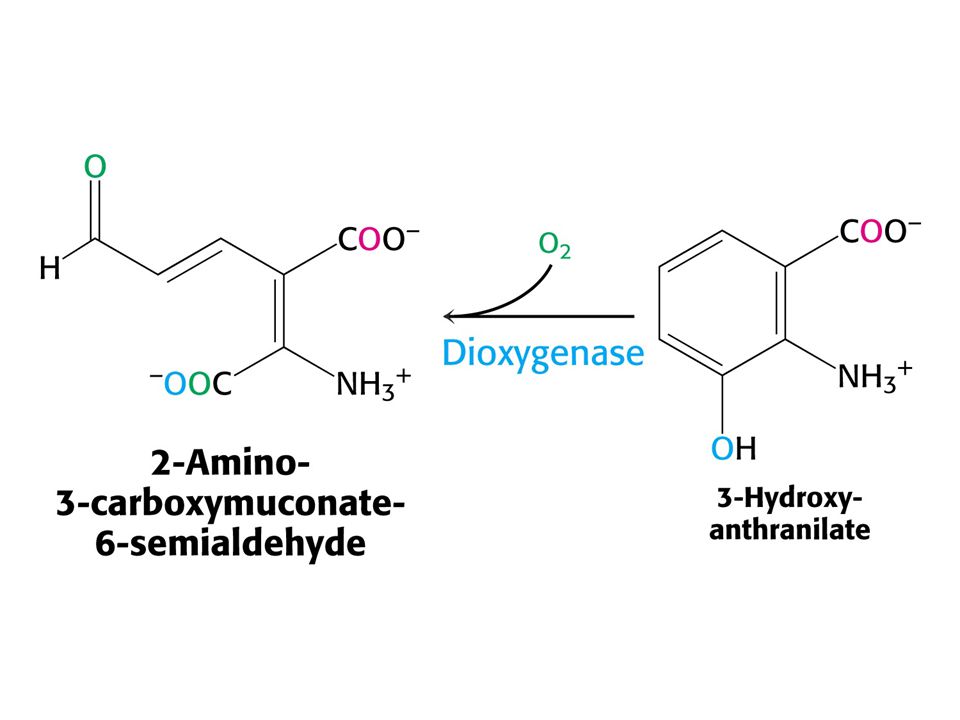
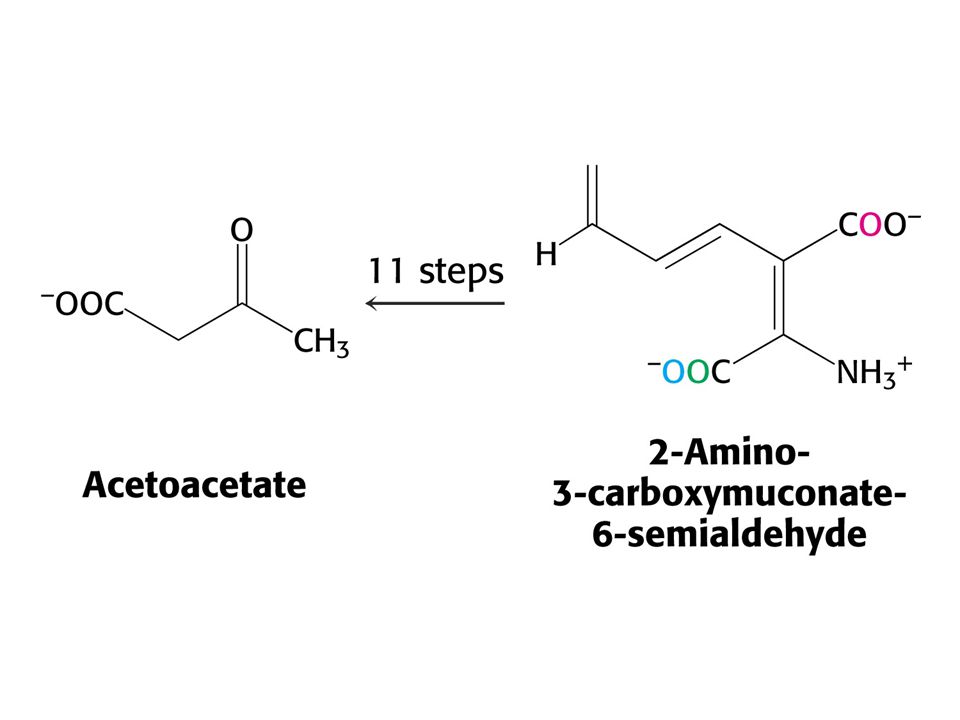
Odbourávání Trp – šestnáct kroků. Tryptofan-2,3-dixygenasa, kynurenin-3-monooxygenasa a další dioxygenasa štěpí 3-hydroxyanthranilát.

122
Vrozené vady metabolismu aminokyselin.
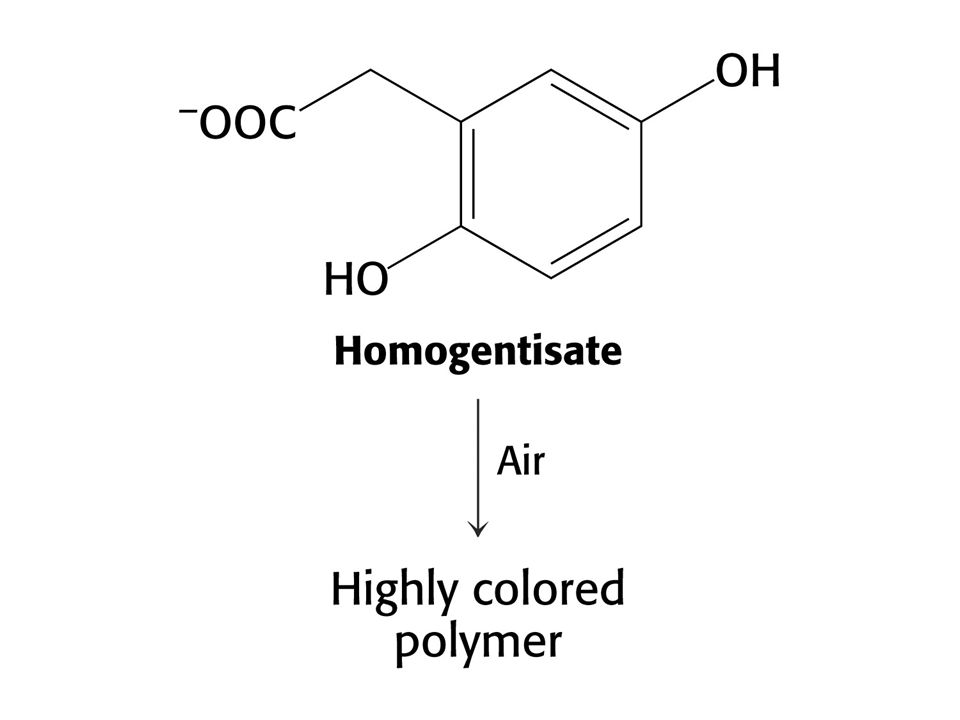
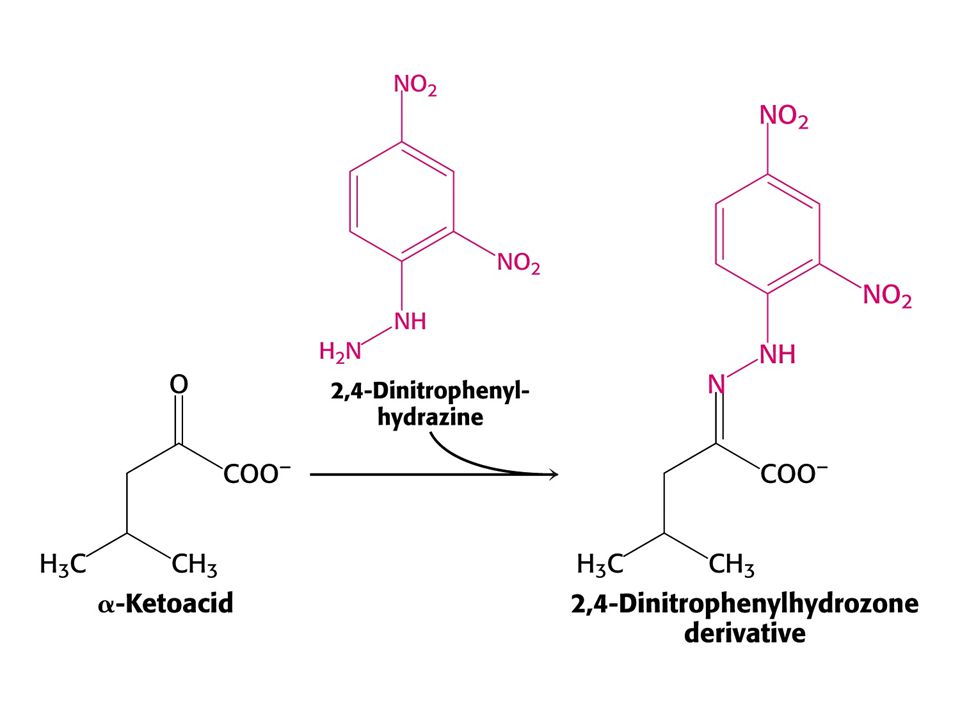
Alkaptonurie – absence homogentisátoxidasy. Homogentisát se vylučuje močí a způsobuje její černání (melaninové látky). Moč s pachem javorového sirupu – je blokována oxidativní dekarboxylace a-oxokyselin z větvených aminokyselin (Val, Leu a Ile). Mentální a fyzická retardace. Nemoc lze zachytit u novorozenců a dodržovat dietu bez větvených aminokyselin. Činidlem je 2,4-dinitrofenylhydrazin.
. Moč s pachem javorového sirupu – je blokována oxidativní dekarboxylace a-oxokyselin z větvených aminokyselin (Val, Leu a Ile). Mentální a fyzická retardace. Nemoc lze zachytit u novorozenců a dodržovat dietu bez větvených aminokyselin. Činidlem je 2,4-dinitrofenylhydrazin.")
125
Fenylketonurie – absence nebo nedostatečnost fenylalaninhydroxylasy nebo méně často jejího tetrabiopterinového kofaktoru. Četnost je 1 : 20 tis. novorozeňat. Hromadí se Phe. Normálně jsou ¾ Phe převedeny na Tyr a zbylá čtvrtina zapojena do proteinů. Při fenylketonurii je hladina Phe 20x vyšší než normálně. Fenylpyruvát se dá detegovat s FeCl3 – vzniká olivově zelené zbarvení. Preferuje se stanovení fenylpyruvátu v krvi. Prenatální diagnóza je možná – gen byl naklonován Therapie: Dieta s nízkým obsahem Phe. Nízký obsah Phe má kasein z mléka. Ten je hydrolyzován a Phe odstraněn adsorpcí.

127
Některá další onemocnění způsobená poruchou metabolismu aminokyselin.

Podobné prezentace