Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Studijní materiál pro bakaláře FTK UP Olomouc
METABOLISMUS Studijní materiál pro bakaláře FTK UP Olomouc Doc. MUDr. Pavel Stejskal, CSc

2
ENERGETICKÝ METABOLISMUS
Energie Získávání energie z okolí, Uvolňování energie v buňce Přenos energie v buňce Transformace energie na potřebné formy energie uvnitř buňky Nevzniká Neztrácí se Různé formy energie se navzájem přeměňují Při každé transformaci energie se část uvolní jako teplo (nevyužitelná energie - není schopna konat práci) Energie schopná konat práci - volná energie ENERGETICKÝ METABOLISMUS
 Energie schopná konat práci - volná energie. ENERGETICKÝ METABOLISMUS.")
3
Energie Neustálý přívod vhodné formy energie z okolí
Tepelná energie musí být odváděna z buňky do okolí Buňka - otevřený systém Při chemických reakcích se tato energie buď uvolňuje exergonické reakce (např. biologická oxidace vodíku) nebo spotřebovává endergonické reakce (např. syntéza polysacharidů, bílkovin nebo nukleových kyselin) Spontánně probíhají pouze reakce exergonické
 nebo spotřebovává. endergonické reakce (např. syntéza polysacharidů, bílkovin nebo. nukleových kyselin) Spontánně probíhají pouze reakce exergonické.")
4
METABOLISMUS látková a energetická přeměna
v buňkách a v živých organizmech s cílem zachovat život Umožňuje živým organismům růst a rozmnožování udržování jejich struktury reagovat na zevní prostředí Celkový metabolismus všechny biochemické procesy v lidském organismu Buněčný metabolismus všechny chemické procesy v buňce Metabolity vznikají a vzájemně se přeměňují při látkové výměně

5
Primární metabolismus závisí život a růst organismu
pro většinu živých organismů podobné (např. metabolismus sacharidů, tuků, aminokyselin a nukleových kyselin) Sekundární metabolismus v jednotlivých skupinách organizmů produkce a odbourávání specifických chemických látek (např. obranné látky, signální molekuly - hormony, pigmenty, stavební molekuly, atd.) Chemické reakce uspořádány do metabolických drah jedna chemická látka transformována do jiné pomocí tzv. enzymů Enzymy umožňují efektivně a rychle řídit systém navzájem napojených termodynamických reakcí regulují metabolické dráhy při reakci na změny v buněčném prostředí nebo na signály z jiných buněk
 Sekundární metabolismus. v jednotlivých skupinách organizmů. produkce a odbourávání specifických chemických látek (např. obranné. látky, signální molekuly - hormony, pigmenty, stavební molekuly, atd.) Chemické reakce uspořádány do metabolických drah. jedna chemická látka transformována do jiné pomocí tzv. enzymů. Enzymy. umožňují efektivně a rychle řídit systém navzájem napojených. termodynamických reakcí. regulují metabolické dráhy při reakci na změny v buněčném prostředí. nebo na signály z jiných buněk.")
6
Anabolismus (výstavbový proces)
soubor syntetických reakcí (biosyntéza) z látek jednodušších vznikají látky složitější (stavební látky – bílkoviny a zásobní látky – glykogen a triglyceridy) Vyžadují dodávání energie, která se při nich spotřebovává Převažují v buňce, která roste a rozmnožuje se Obnovování živé hmoty Vytváření energetických zásob (pro mechanickou práci, transport membránou, atd.) Vytváření látek potřebných k řízení (hormonů, enzymů nebo mediátorů)
 z látek jednodušších vznikají látky složitější. (stavební látky – bílkoviny a zásobní látky – glykogen a triglyceridy) Vyžadují dodávání energie, která se při nich spotřebovává. Převažují v buňce, která roste a rozmnožuje se. Obnovování živé hmoty. Vytváření energetických zásob (pro mechanickou práci, transport. membránou, atd.) Vytváření látek potřebných k řízení (hormonů, enzymů nebo. mediátorů)")
7
Katabolismus (rozkladový proces)
rozklad látek složitějších (zásobních látek nebo přijatých živin) na látky jednodušší a energii Např. oxidativní procesy - uvolňují ze sloučenin volnou energii Energie uvolněná katabolickými pochody Udržení tělesných funkcí Štěpení a využití potravy Termoregulace Pohybová aktivita
 na látky jednodušší a energii. Např. oxidativní procesy - uvolňují ze sloučenin volnou energii. Energie uvolněná katabolickými pochody. Udržení tělesných funkcí. Štěpení a využití potravy. Termoregulace. Pohybová aktivita.")
8
Energetický metabolismus - z chemické energie živin
biologická energie využitelná v organismu Živiny procházejí třemi fázemi chemického zpracování: Ve střevě při trávení - složité živiny na jednoduché vstřebatelné složky Jednoduché cukry Aminokyseliny Glycerol a mastné kyseliny

9
Energetický metabolismus - z chemické energie živin
biologická energie využitelná v organismu Živiny procházejí třemi fázemi chemického zpracování: 2. V cytoplazmě buněk tkání nastává další degradace. Glukóza pyruvát Mastné kyseliny Aminokyseliny Baze nukleových kyselin kyselina acetoctová

10
UTP Glykogen Pi G-fosforyláza laktát UDP G 1-P NAD ADP ATP IZOMERACE LDH NADH G 6-P pyruvát IZOMERACE GL hexokináza F 6-P Pyruvát kináza Pi ATP ATP fruktóza-difosfatáza PFK fosfoglycerát kináza ADP ADP F 1,6-P ATP ADP enoláza P-enol pyruvát Glyceraldehyd 3-P Glyceraldehyd 3-P NADH NAD dtto Glyceraldehyd dehydrogenáza

11
Energetický metabolismus - z chemické energie živin
biologická energie využitelná v organismu Živiny procházejí třemi fázemi chemického zpracování: 2. V cytoplazmě buněk tkání nastává další degradace. Glukóza pyruvát Mastné kyseliny Aminokyseliny Baze nukleových kyselin Nejsnáze a nejrychleji - přeměna glukózy už v cytoplazmě + vznik volné energie kyselina acetoctová

12
acetyl koenzym A (acetyl-CoA) respiračního řetězce
Energetický metabolismus - z chemické energie živin biologická energie využitelná v organismu Živiny procházejí třemi fázemi chemického zpracování: 3. V mitochondriích pyruvát a kyselina acetoctová odbourávány na společný meziprodukt acetyl koenzym A (acetyl-CoA) vstupuje do cyklu kyseliny citrónové respiračního řetězce Úplná oxidace za vzniku energie ve formě adenositrifosfátu (ATP) a konečných produktů (vody a kysličníku uhličitého).
 vstupuje do cyklu kyseliny citrónové. respiračního řetězce. Úplná oxidace za vzniku energie ve formě adenositrifosfátu. (ATP) a konečných produktů (vody a kysličníku uhličitého).")
13
Alfa-ketoglutarát (5C)
Pyruvát (3C) NAD+ CO2 NADH + H+ Acetyl-CoA (2C) Oxalacetát (4C) Citrát (6C) NAD+ NADH + H+ Malát (4C) Izocitrát (6C) NAD+ CO2 Fumarát (4C) NADH + H+ FADH2 Alfa-ketoglutarát (5C) Sukcinát (4C) P FAD Sukcinyl-CoA (4C) CO2 GTP NAD+ NADH + H+ GDP
 NAD+ CO2. NADH + H+ Acetyl-CoA (2C) Oxalacetát (4C) Citrát (6C) NAD+ NADH + H+ Malát (4C) Izocitrát (6C) NAD+ CO2. Fumarát (4C) NADH + H+ FADH2. Alfa-ketoglutarát (5C) Sukcinát (4C) P. FAD. Sukcinyl-CoA (4C) CO2. GTP. NAD+ NADH + H+ GDP.")
14
Usměrnění reakcí a řízení rychlosti metabolických dějů
Katalyzátory - Enzymy umožňují řídit systém navzájem napojených termodynamických reakcí Aktivitu enzymů ovlivňuje přítomnost některých iontů či metabolitů Enzymy jsou velmi specifické katalyzují reakci jediné látky (substrátu) na zcela určitou sloučeninu (produkt) A enzym B
 na zcela určitou sloučeninu (produkt) A. enzym. B.")
15
Reakce přeměn látek na sebe navazují
produkt jedné reakce tvoří substrát reakce druhé METABOLICKÉ DRÁHY Přeměna látek není jen jednou reakcí ale celou řadou reakcí A enzym B enzym C enzym D

16
Metabolické děje jsou jako celek jednosměrné
enzym B enzym C enzym D Metabolické děje jsou jako celek jednosměrné protože se při nich část energie přemění na energii tepelnou kterou nelze zcela převést na jiné formy energie A ENERGIE SE MUSÍ DODAT Z JINÝCH ZDROJŮ! Energie enzym CHYBÍ ENERGIE! B

17
MK + ATP + CoA Acyl-CoA + AMP + PPi KOFAKTOR energie
+ ENZYMY = SLOŽENÉ ENZYMY Kofaktory nezbytné pro funkci daného enzymu přenos atomů, skupin atomů nebo elektronů při chemické reakci katalyzováné enzymem deriváty vitamínů rozpustných ve vodě váží molekuly kovů tvořeny některými stopovými prvky (např.železo nebo zinek) energie enzym acyl CoA syntetáza MK + ATP + CoA Acyl-CoA + AMP + PPi Mg2+ kofaktor
 energie. enzym. acyl CoA syntetáza. MK + ATP + CoA. Acyl-CoA + AMP + PPi. Mg2+ kofaktor.")
18
KOENZYM přenos atomů, jejich skupin, nebo samotných elektronů
mezi odlišnými enzymy Spřažení různých biochemických reakcí Během těchto spřažených reakcí koenzymy regenerují Koenzym, který se např. při první reakci sloučil s vodíkem přechází na jiný enzym v jiné chemické reakci se opět vrací do původního stavu Hladina aktivních koenzymů v buňce je udržována na přibližně stejné úrovni C D A B Enzym A Enzym B K K H+ H+

19
Hlavní katabolické procesy u člověka:
Do katabolických procesů mnoho různých organických látek vystupuje z nich málo produktů Hlavní katabolické procesy u člověka: a) rozkládání živin na své složky; b) glykolýza rozkládající glukózu na pyruvát a následně na acetyl-CoA; c) β-oxidace mastných kyselin - vzniká acetyl-CoA; d) rozklad aminokyselin a bazí nukleových kyselin; e) citrátový cyklus, vstupuje acetyl-CoA – vzniká oxid uhličitý, f) respirační (dýchací) řetězec, vstupují vodíkové atomy získané z rozkladu sacharidů, mastných kyselin, aminokyselin a bazí nukleových kyselin a je získána energie v podobě vysokoenergetických sloučenin. Vodíkové ionty jsou oxidovány na vodu.
 rozkládání živin na své složky; b) glykolýza rozkládající glukózu na pyruvát a následně na acetyl-CoA; c) β-oxidace mastných kyselin - vzniká acetyl-CoA; d) rozklad aminokyselin a bazí nukleových kyselin; e) citrátový cyklus, vstupuje acetyl-CoA – vzniká oxid uhličitý, f) respirační (dýchací) řetězec, vstupují vodíkové atomy získané z rozkladu sacharidů, mastných kyselin, aminokyselin a bazí nukleových kyselin a je získána energie v podobě vysokoenergetických sloučenin. Vodíkové ionty jsou oxidovány na vodu.")
20
Vysokoenergetické (makroergické) fosfátové sloučeniny
Energie uvolněná katabolismem není využitá buňkami přímo tvorba vazeb mezi zbytky kyseliny fosforečné a některými organickými sloučeninami (fosfáty) V těchto jednoduchých sloučeninách velké množství energie Při jejich rozložení (reakce s vodou = hydrolýza) se uvolňuje energie Vysokoenergetické (makroergické) fosfátové sloučeniny nejdůležitějším reprezentantem adenozintrifosfát (ATP)
 V těchto jednoduchých sloučeninách. velké množství energie. Při jejich rozložení (reakce s vodou = hydrolýza) se uvolňuje energie. Vysokoenergetické (makroergické) fosfátové sloučeniny. nejdůležitějším reprezentantem. adenozintrifosfát (ATP)")
21
ATP - hlavní „energetická konzerva“
ATP rozvádí energii v buňce a předává ji tam, kde je jí potřeba. Hromadění molekul ATP v buňce = = zásoba okamžitě použitelné energie Vazba mezi druhým a třetím fosfátem (makroergní fosfátová vazba) relativně velké množství energie (asi 50 kJ) ATP - hlavní „energetická konzerva“
 relativně velké množství energie (asi 50 kJ) ATP - hlavní „energetická konzerva")
22
ATP + H2O = ADP + P + 50 kJ V místě potřeby energie
(např. při svalovém stahu nebo svalovém uvolnění nebo při syntéze řady chemických sloučenin) molekula ATP se rozštěpí (je hydrolyzována) na adenosindifosfát (ADP) + fosfát (kyselina fosforečná H3PO4, značeno jen jako P) uvolněná energie ATP + H2O = ADP + P + 50 kJ
 molekula ATP se rozštěpí (je hydrolyzována) na adenosindifosfát (ADP) + fosfát (kyselina fosforečná H3PO4, značeno jen jako P) uvolněná energie. ATP + H2O = ADP + P + 50 kJ.")
23
Koloběh ATP a ADP - základní buněčná funkce
ADP se vrací do mitochondrií znovu fosforylace na ATP Koloběh ATP a ADP - základní buněčná funkce ATP se může rozložit na ADP + 1 P nebo úplnou oxidací na adenozinmonofosfát (AMP) + 2P.
 + 2P.")
24
ATP se může rozložit na ADP + 1 P
nebo úplnou oxidací na adenozinmonofosfát (AMP) + 2P.
 + 2P.")
25
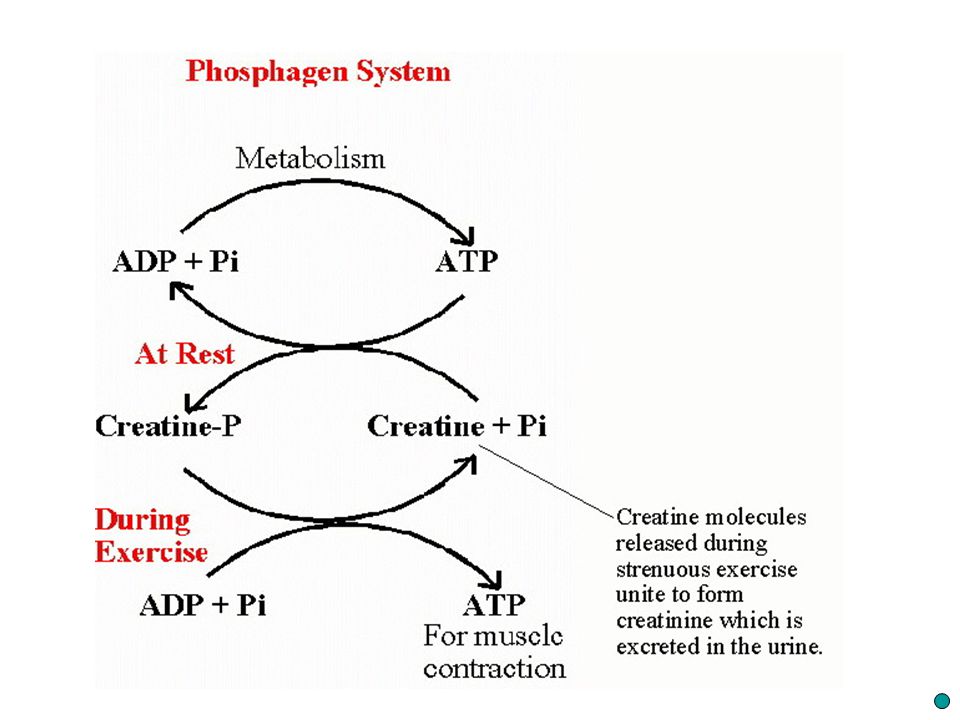
Kreatinfosfát předá svoji fosfátovou skupinu ADP CP + ADP = C + ATP
Zásobní vysokoenergetický substrát kreatinfosfát (CP) Při rozštěpení na kreatin a fosfátový iont (CP = C + P) uvolní se větší množství energie než při rozštěpení vazby ATP Kreatinfosfát předá svoji fosfátovou skupinu ADP CP + ADP = C + ATP Rekonstrukce makroergní vazby ATP
 Při rozštěpení na kreatin a fosfátový iont (CP = C + P) uvolní se větší množství energie než při rozštěpení vazby ATP. Kreatinfosfát předá svoji fosfátovou skupinu ADP. CP + ADP = C + ATP. Rekonstrukce makroergní vazby ATP.")
26
za dostatečného přívodu kyslíku do buněk vysoká
Ve svalu je asi pětkrát víc CP než ATP celá reakce trvá zlomek vteřiny Takto vzniklý kreatin (C) (organická látka přirozeně přítomná v organismu) se v mitochondriích bez nároků na další energii opět slučuje s fosforem a vytváří nový kreatinfosfát (C + P = CP) Tato tvorba kreatinfosfátu je za dostatečného přívodu kyslíku do buněk vysoká Naopak při nedostatku kyslíku je nízká
 (organická látka přirozeně přítomná v organismu) se v mitochondriích bez nároků na další energii. opět slučuje s fosforem. a vytváří nový kreatinfosfát (C + P = CP) Tato tvorba kreatinfosfátu je. za dostatečného přívodu kyslíku do buněk vysoká. Naopak při nedostatku kyslíku je nízká.")
27
(3 – 8 mmol/kg svalů) teoreticky během dvou sekund
ATP a CP tvoří dohromady systém poskytuje v prvních dvou sekundách svalové kontrakce okamžitou energii Množství ATP ve svalech je velmi malé (3 – 8 mmol/kg svalů) Při intenzivní svalové činnosti by teoreticky mohl být všechen ATP spotřebován během dvou sekund
 Při intenzivní svalové činnosti by. teoreticky. mohl být všechen ATP spotřebován. během dvou sekund.")
28
už po 1,3 sekundách ATP a CP systém teoreticky během dvou sekund
Při intenzivní svalové činnosti by teoreticky mohl být všechen ATP spotřebován během dvou sekund Prakticky ATP vyčerpat nelze, CP vyčerpat lze Celý systém ATP- CP disponuje velmi malou energií (asi 5,7 – 6,9 kcal) Proto rychlost degradace CP prudce klesá už po 1,3 sekundách Tím se buňka brání úplnému vyčerpání CP
 Proto rychlost degradace CP prudce klesá. už po 1,3 sekundách. Tím se buňka brání úplnému vyčerpání CP.")
30
10 – 12 kcal využitelné energie Velmi malé množství energie
např. při běhu na 1500 m potřebuje organismus asi 120 kcal Stačí na 150 m Stačí na 150 m

31
nikotinamidadenindinukleotid (NAD)
Většina energie z rozložených složitějších látek získávána pomocí oxidačních reakcí Přenašeči energie - koenzymy uvolňují elektrony v elektronovém transportním systému RESPIRAČNÍ ŘETĚZEC Nejdůležitější koenzym nikotinamidadenindinukleotid (NAD) přijímá protony a elektrony z reakcí při kterých se uvolňuje vodík (tzv. dehydrogenační reakce) Přenašeč energie: NAD+ se redukuje na NADH přitom vzniká energie 3 ATP
 přijímá protony a elektrony z reakcí. při kterých se uvolňuje vodík (tzv. dehydrogenační reakce) Přenašeč energie: NAD+ se redukuje na NADH. přitom vzniká energie 3 ATP.")
32
RESPIRAČNÍ ŘETĚZEC ATP ATP ATP 1/2 O2 NADH2 NAD+ 1 2 3 H20 ADP+Pi

33
RESPIRAČNÍ ŘETĚZEC ATP ATP ATP NAD NADH 3 2 1 H+ ADP+Pi ADP+Pi ADP+Pi

34
Flavinadenindinukleotid (FAD) redukcí vznikají jen dvě molekuly ATP
FADH2 FAD+ ATP ATP 1/2 O2 2 3 H20 ADP+Pi ADP+Pi RESPIRAČNÍ ŘETĚZEC

35
Flavinadenindinukleotid (FAD) redukcí vznikají jen dvě molekuly ATP
FADH2 ATP ATP 2 1 H+ ADP+Pi ADP+Pi RESPIRAČNÍ ŘETĚZEC

36
RESPIRAČNÍ ŘETĚZEC Respirační řetězec
lokalizovaný na vnitřní membráně mitochondrií volná energie jeho elektrochemického potenciálu se využívá k syntéze ATP Respirační řetězec systém specifických enzymů (oxidoreduktáz) ELEKTRONY ATOMOVÝCH VODÍKŮ (odebraných organickým nebo anorganickým substrátům) SE VÁŽÍ NA MOLEKULY KYSLÍKU Oxidoreduktázy respiračních řetězců 4 kotvené proteinové enzymové komplexy (komplexy I, II, III a IV) součásti vnitřní mitochondriální membrány
 ELEKTRONY ATOMOVÝCH VODÍKŮ. (odebraných organickým nebo anorganickým substrátům) SE VÁŽÍ NA MOLEKULY KYSLÍKU. Oxidoreduktázy respiračních řetězců. 4 kotvené proteinové enzymové komplexy. (komplexy I, II, III a IV) součásti vnitřní mitochondriální membrány.")
37
FLAVOPROTEIN-CYTOCHORMOVÝ SYSTÉM přenáší vodík na kyslík
NAD+ NAD+ NAD+ NADH2 NAD+ NADH2 NAD+ NADH2 2H+ 2H+ 2H+ NAD+ 1/2O2+ 2H+ cytochromoxidáza H20 řetězec koenzymů přenáší vodík na kyslík MITOCHONDRIE

38
Nejdůležitější děj aerobního katabolismu a tvoří jeho konečnou fázi.
RESPIRAČNÍ ŘETĚZEC Transport elektronů z koenzymu na koenzym (NAD nebo FAD) pomocí série reakcí Konečným akceptorem těchto elektronů na konci řetězce je O2 Nejdůležitější děj aerobního katabolismu a tvoří jeho konečnou fázi.
 pomocí série reakcí. Konečným akceptorem těchto elektronů. na konci řetězce je O2. Nejdůležitější děj aerobního katabolismu. a tvoří jeho konečnou fázi.")
39
kalorie (cal, malá kalorie, gram kalorie)
Jednotkou energie kalorie (cal, malá kalorie, gram kalorie) množství energie zvyšující teplotu 1 g vody z 15 na 16o C nebo joul (J) 1 cal = 4,18 J; 1 J = 0,239 cal. Násobky těchto jednotek kilokalorie (kcal, velká kalorie, kilogram kalorie) = 1000 cal = 4,18 kJ kilojoul (kJ) = 1000 J = 0,239 kcal
 množství energie zvyšující teplotu 1 g vody z 15 na 16o C. nebo joul (J) 1 cal = 4,18 J; 1 J = 0,239 cal. Násobky těchto jednotek. kilokalorie (kcal, velká kalorie, kilogram kalorie) = 1000 cal = 4,18 kJ. kilojoul (kJ) = 1000 J = 0,239 kcal.")
40
Charakteristika energie poskytnuté různými živinami: SPALNÉ TEPLO
Množstvím energie, které se uvolní dokonalým spálením z jednotkového množství (jednoho gramu) živin = SPALNÉ TEPLO Hodnoty spalného tepla pro jednotlivé živiny: sacharidy = 4,1 kcal/g (17,1 kJ/g) tuky = 9,3 kcal/g (38,9 kJ/g) bílkoviny = 5,3 kcal/g (22,2 kJ/g)
 živin. = SPALNÉ TEPLO. Hodnoty spalného tepla pro jednotlivé živiny: sacharidy = 4,1 kcal/g (17,1 kJ/g) tuky = 9,3 kcal/g (38,9 kJ/g) bílkoviny = 5,3 kcal/g (22,2 kJ/g)")
41
Charakteristika energie poskytnuté různými živinami:
2. Množstvím tepla, které se uvolní, když se k oxidaci jednotlivých živin spotřebuje 1 litr kyslíku = ENERGETICKÝ EKVIVALENT (EE, dříve kalorický ekvivalent) Hodnoty energetického ekvivalentu pro jednotlivé živiny: sacharidy = 5,05 kcal (21,1 kJ) tuky = 4,55 kcal (19,0 kJ) bílkoviny = 4,31 kcal (18,0 kJ) smíšená strava = 4,81 (20,1 kJ)
 Hodnoty energetického ekvivalentu pro jednotlivé živiny: sacharidy = 5,05 kcal (21,1 kJ) tuky = 4,55 kcal (19,0 kJ) bílkoviny = 4,31 kcal (18,0 kJ) smíšená strava = 4,81 (20,1 kJ)")
42
Charakteristika energie poskytnuté různými živinami:
3. Jaké substráty jsou aktuálně metabolizovány = RESPIRAČNÍ KVOCIENT (RQ nebo R) Při oxidaci energetických substrátů spotřebovává se kyslík a vylučuje oxid uhličitý RQ = poměr mezi vydaným CO2 a spotřebovaným O2 za podmínek setrvalého stavu
 Při oxidaci energetických substrátů. spotřebovává se kyslík a vylučuje oxid uhličitý. RQ = poměr mezi vydaným CO2 a spotřebovaným O2. za podmínek setrvalého stavu.")
43
Charakteristika energie poskytnuté různými živinami:
3. RESPIRAČNÍ KVOCIENT Normální hodnoty RQ pro jednotlivé substráty jsou: sacharidy = 1,00 tuky = 0,70 bílkoviny = 0,82 průměrná hodnota běžné potravy = 0,80 Určení RQ při spalování proteinů v těle: Nevzniká pouze oxid uhličitý a voda, ale i jednoduché sloučeniny dusíku (tzv. nebílkovinný dusík, např. močovina, volné aminokyseliny, kyselina močová, kreatin, kreatinin, amoniak, aj.) lidský organismus neumí dále rozložit a získat z nich energii
 lidský organismus neumí dále rozložit a získat z nich energii.")
44
Charakteristika energie poskytnuté různými živinami:
3. RESPIRAČNÍ KVOCIENT Liší se v různých podmínkách: 1. Při hyperventilaci (prohloubené dýchání) RQ stoupá je vydechován ve zvýšené míře oxid uhličitý 2. Při intenzivní námaze RQ stoupá kyselina mléčná (vzniká při intenzivní námaze) se mění na CO2 (viz nárazníkový systém) CO2 ve zvýšené míře vydechován 3. Při acidóze (snížení pH pod 7,36, zvýšení kyselosti krve) RQ stoupá 4. Při alkalóze (zvýšení pH nad 7,44, snížení kyselosti krve) RQ klesá
 RQ stoupá. je vydechován ve zvýšené míře oxid uhličitý. 2. Při intenzivní námaze RQ stoupá. kyselina mléčná (vzniká při intenzivní námaze) se mění na CO2 (viz nárazníkový systém) CO2 ve zvýšené míře vydechován. 3. Při acidóze (snížení pH pod 7,36, zvýšení kyselosti krve) RQ stoupá. 4. Při alkalóze (zvýšení pH nad 7,44, snížení kyselosti krve) RQ klesá.")
45
Energetická spotřeba 1. PŘÍMÁ KALORIMETRIE kalorimetr
můžeme vypočítat 1. PŘÍMÁ KALORIMETRIE (zjištění množství tepla, jež se uvolní při spálení 1g živin) kalorimetr 2. pomocí tzv. TĚŽKÉ VODY (místo vodíku deuterium) Obě metody nákladné a složité přístrojové vybavení význam jen ve výzkumu
 kalorimetr. 2. pomocí tzv. TĚŽKÉ VODY. (místo vodíku deuterium) Obě metody nákladné a složité přístrojové vybavení. význam jen ve výzkumu.")
46
Energetická spotřeba V praxi měření metodou NEPŘÍMÉ KALORIMETRIE
spotřeba živin vypočítána ze spotřeby kyslíku (VO2) a výdeje oxidu uhličitého (VCO2) (viz RQ = VCO2/VO2)
 a výdeje oxidu uhličitého (VCO2) (viz RQ = VCO2/VO2)")
47
Využívání získané energie na
Energetická spotřeba Využívání získané energie na BAZÁLNÍ METABOLIZMUS TRÁVENÍ A VSTŘEBÁVÁNÍ SVALOVOU PRÁCI REGULACI TĚLESNÉ TEPLOTY (termoregulace)
")
48
BAZÁLNÍ METABOLISMUS (základní látková výměna, BMR)
energetická spotřeba nezbytná k udržení základních, životně nezbytných funkcí (srdeční činnost, dýchání, činnost mozku, atd.) Odpovídá minimálnímu množství energie potřebnému k udržení homeostázy (stabilita vnitřního prostředí)
 Odpovídá minimálnímu množství energie. potřebnému k udržení homeostázy. (stabilita vnitřního prostředí)")
49
Faktory ovlivňující rychlost metabolismu
BAZÁLNÍ METABOLISMUS Měříme ráno před probuzením nebo bezprostředně po probuzení před požitím jídla, 12–14 hodin po posledním jídle, v klidu při příjemné pokojové teplotě = Faktory ovlivňující rychlost metabolismu svalová práce specificko-dynamický účinek potravy (nezbytný výdej energie, potřebný pro zpracování živin v těle, nejvyšší bílkoviny) vnější teplota
 vnější teplota.")
50
Zdravý dospělý muž mladšího věku
BAZÁLNÍ METABOLISMUS závisí na povrchu těla (tělesná výška a hmotnosti) věku (věkem se snižuje) pohlaví (žena menší – tuk menší metabolismus než svaly) Zdravý dospělý muž mladšího věku BMR = 40 kcal/m2/hod = 2000 kcal/den . .
 věku (věkem se snižuje) pohlaví (žena menší – tuk menší metabolismus. než svaly) Zdravý dospělý muž mladšího věku. BMR = 40 kcal/m2/hod. = 2000 kcal/den. . .")
51
MET 3,5 ml VO2/kg.min Energetický výdej při pohybové aktivitě
často se vyjadřuje jako násobek klidového metabolismu (REE) MET (metabolický ekvivalent, množné číslo METs) 1 MET = klidová energetická spotřeba 3,5 ml VO2/kg.min Např. VO2 = 38 ml/kg.min = 10,86 METs (téměř jedenáctinásobné zvýšení VO2 proti REE)
 MET. (metabolický ekvivalent, množné číslo METs) 1 MET = klidová energetická spotřeba. 3,5 ml VO2/kg.min. Např. VO2 = 38 ml/kg.min. = 10,86 METs. (téměř jedenáctinásobné zvýšení VO2 proti REE)")
52
Uvolňování nemůže proběhnout najednou velké množství tepla
Všechny buňky uvolňují energii chemických vazeb v podstatě stejným způsobem Uvolňování nemůže proběhnout najednou velké množství tepla zničení buněčné struktury Energie z organických látek = postupné štěpení chemických vazeb

53
(ANAEROBNÍ) GLYKOLÝZA OXIDATIVNÍ FOSFORYLACE
Uvolňování energie 1. z jednoduchého sacharidu (glukózy) rozštěpením uhlíkového řetězce bez přístupu kyslíku (ANAEROBNÍ) GLYKOLÝZA 2. postupnou oxidací vodíku odňatého z rozkládané látky (tj. sacharidu, mastné kyseliny, aminokyseliny) na vodu OXIDATIVNÍ FOSFORYLACE
 rozštěpením uhlíkového řetězce. bez přístupu kyslíku. (ANAEROBNÍ) GLYKOLÝZA. 2. postupnou oxidací vodíku odňatého. z rozkládané látky (tj. sacharidu, mastné kyseliny, aminokyseliny) na vodu. OXIDATIVNÍ FOSFORYLACE.")
54
Glykolýza je metabolická dráha
přeměny glukózy ( 6 C) na dvě molekuly pyruvátu (3 C) za čistého výtěžku dvou molekul ATP a dvou molekul NADH Probíhá nezávisle na kyslíku v cytosolu buněk (cytoplazma) Skládá se z deseti kroků, každý z nich katalyzuje jiný enzym
 na dvě molekuly pyruvátu (3 C) za čistého výtěžku dvou molekul ATP. a dvou molekul NADH. Probíhá nezávisle na kyslíku v cytosolu buněk (cytoplazma) Skládá se z deseti kroků, každý z nich katalyzuje jiný enzym.")
55
UTP Glykogen Pi G-fosforyláza UDP G 1-P ADP ATP IZOMERACE LDH G 6-P pyruvát IZOMERACE GL hexokináza F 6-P Pyruvát kináza Pi ATP ATP fruktóza-difosfatáza PFK fosfoglycerát kináza ADP ADP F 1,6-P ATP ADP enoláza P-enol pyruvát Glyceraldehyd 3-P Glyceraldehyd 3-P NADH NAD dtto Glyceraldehyd dehydrogenáza

56
Při dostatečném množství kyslíku
se pyruvát mění v mitochondriích na acetylkoenzym-A (2 uhlíky) který se slučuje s oxalacetátem (4 uhlíky) na citrát (6 uhlíků)
 který se slučuje s oxalacetátem (4 uhlíky) na citrát (6 uhlíků)")
57
Alfa-ketoglutarát (5C)
Pyruvát (3C) NAD+ CO2 NADH + H+ Acetyl-CoA (2C) Oxalacetát (4C) Citrát (6C) NADH + H+ Malát (4C) Izocitrát (6C) NAD+ CO2 Fumarát (4C) NADH + H+ FADH2 Alfa-ketoglutarát (5C) Sukcinát (4C) P FAD Sukcinyl-CoA (4C) CO2 GTP NAD+ NADH + H+ GDP
 NAD+ CO2. NADH + H+ Acetyl-CoA (2C) Oxalacetát (4C) Citrát (6C) NADH + H+ Malát (4C) Izocitrát (6C) NAD+ CO2. Fumarát (4C) NADH + H+ FADH2. Alfa-ketoglutarát (5C) Sukcinát (4C) P. FAD. Sukcinyl-CoA (4C) CO2. GTP. NAD+ NADH + H+ GDP.")
58
Postupnou dekarboxylací a oxidací citrátu
se uvolňuje CO2 (odpadní produkt) a vodíkové kationy (H+) které redukují koenzymy NAD a FAD Protony nesené redukovanými koenzymy (NADH a FADH) jsou použity v dýchacím řetězci kde oxidativní fosforylací dochází k syntéze ATP a tvorbě H2O
 a vodíkové kationy (H+) které redukují koenzymy NAD a FAD. Protony nesené redukovanými koenzymy. (NADH a FADH) jsou použity v dýchacím řetězci. kde oxidativní fosforylací dochází. k syntéze ATP a tvorbě H2O.")
59
Alfa-ketoglutarát (5C)
Pyruvát (3C) NAD+ CO2 NADH + H+ Acetyl-CoA (2C) Citrát (6C) Malát (4C) Izocitrát (6C) NAD+ CO2 Fumarát (4C) NADH + H+ FADH2 Alfa-ketoglutarát (5C) Sukcinát (4C) P FAD Sukcinyl-CoA (4C) CO2 GTP NAD+ NADH + H+ GDP
 NAD+ CO2. NADH + H+ Acetyl-CoA (2C) Citrát (6C) Malát (4C) Izocitrát (6C) NAD+ CO2. Fumarát (4C) NADH + H+ FADH2. Alfa-ketoglutarát (5C) Sukcinát (4C) P. FAD. Sukcinyl-CoA (4C) CO2. GTP. NAD+ NADH + H+ GDP.")
60
Protože konečným produktem tohoto rozkladu citrátu je opět oxalacetát
(který se kondenzuje s dalším acetyl-CoA) má reakce cyklický charakter nazývá se cyklus kyseliny citrónové (nebo Krebsův cyklus)
 má reakce cyklický charakter. nazývá se cyklus kyseliny citrónové. (nebo Krebsův cyklus)")
61
Alfa-ketoglutarát (5C)
Pyruvát (3C) NAD+ CO2 NADH + H+ Acetyl-CoA (2C) Oxalacetát (4C) Citrát (6C) NAD+ NADH + H+ Malát (4C) Izocitrát (6C) NAD+ CO2 Fumarát (4C) NADH + H+ FADH2 Alfa-ketoglutarát (5C) Sukcinát (4C) P FAD Sukcinyl-CoA (4C) CO2 GTP NAD+ NADH + H+ GDP
 NAD+ CO2. NADH + H+ Acetyl-CoA (2C) Oxalacetát (4C) Citrát (6C) NAD+ NADH + H+ Malát (4C) Izocitrát (6C) NAD+ CO2. Fumarát (4C) NADH + H+ FADH2. Alfa-ketoglutarát (5C) Sukcinát (4C) P. FAD. Sukcinyl-CoA (4C) CO2. GTP. NAD+ NADH + H+ GDP.")
62
15 molekul ATP celkem 38 molekul ATP
Z jedné molekuly pyruvátu vzniká v mitochondriích 15 molekul ATP (přímo v Krebsově cyklu 12 molekul ATP) Při rozkladu jedné molekuly glukózy vznikne dohromady v cytoplazmě a v mitochondriích celkem 38 molekul ATP
 Při rozkladu jedné molekuly glukózy. vznikne dohromady v cytoplazmě a v mitochondriích. celkem 38 molekul ATP.")
63
Z jedné molekuly glykogenu vznikne o jednu molekulu ATP více
(zásobní polysacharid ve svalových a jaterních buňkách) vznikne o jednu molekulu ATP více (tedy 39 ATP)
 vznikne o jednu molekulu ATP více. (tedy 39 ATP)")
64
pyruvát se mění na kyselinu mléčnou disociuje na laktát
Při relativním nedostatku energie a kyslíku pyruvát se mění na kyselinu mléčnou disociuje na laktát

65
UTP Glykogen Pi G-fosforyláza laktát UDP G 1-P NAD ADP ATP IZOMERACE LDH NADH G 6-P pyruvát IZOMERACE GL hexokináza F 6-P Pyruvát kináza Pi ATP ATP fruktóza-difosfatáza PFK fosfoglycerát kináza ADP ADP F 1,6-P ATP ADP enoláza P-enol pyruvát Glyceraldehyd 3-P Glyceraldehyd 3-P NADH NAD dtto Glyceraldehyd dehydrogenáza

66
koordinátor další látkové výměny
Při vysoké produkci může proniknout laktát z buněk do krve a odtud do jiných tkání koordinátor další látkové výměny LAKTÁT

67
a rozložen v mitochondriích (Krebsově cyklu)
Laktát může být ve svalové tkáni, kde byl vytvořen (většinou bílá svalová vlákna) nebo ve tkáni, do které se dostal krví buď zpátky oxidován na pyruvát a rozložen v mitochondriích (Krebsově cyklu) na CO2, H2O a energii,
 nebo ve tkáni, do které se dostal krví. buď zpátky oxidován na pyruvát. a rozložen v mitochondriích (Krebsově cyklu) na CO2, H2O a energii,")
68
Při tělesné práci glukosa s časem klesá pyruvát s časem stoupá laktát
Acetyl-CoA Při tělesné práci

69
a rozložen v mitochondriích (Krebsově cyklu) zásobní glykogen
Laktát může být ve svalové tkáni, kde byl vytvořen (většinou bílá svalová vlákna) nebo ve tkáni, do které se dostal krví buď zpátky oxidován na pyruvát a rozložen v mitochondriích (Krebsově cyklu) na CO2, H2O a energii, b) nebo se z něj může zpětně vytvořit zásobní glykogen (tzv. glukoneogeneze, tj. tvorba glykogenu z nesacharidových zdrojů)
 nebo ve tkáni, do které se dostal krví. buď zpátky oxidován na pyruvát. a rozložen v mitochondriích (Krebsově cyklu) na CO2, H2O a energii, b) nebo se z něj může zpětně vytvořit. zásobní glykogen. (tzv. glukoneogeneze, tj. tvorba glykogenu z nesacharidových zdrojů)")
70
fruktóza-difosfatáza PFK fosfoglycerát kináza ADP ADP F 1,6-P ATP ADP
UTP Glykogen Pi G-fosforyláza laktát UDP G 1-P NAD GLYKONEOGENEZE ADP ATP IZOMERACE LDH NADH G 6-P pyruvát IZOMERACE GL hexokináza F 6-P Pyruvát kináza Pi ATP ATP fruktóza-difosfatáza PFK fosfoglycerát kináza ADP ADP F 1,6-P ATP ADP enoláza P-enol pyruvát Glyceraldehyd 3-P Glyceraldehyd 3-P NADH NAD dtto Glyceraldehyd dehydrogenáza

71
stává se hlavním aerobním substrátem
Při intenzivní tělesné práci se stává laktát preferovaným energetickým zdrojem stává se hlavním aerobním substrátem

72
aerobní využití laktátu převyšuje využití glukózy
glukosa s časem klesá pyruvát s časem stoupá laktát Acetyl-CoA Při tělesné práci Během tělesné práce aerobní využití laktátu převyšuje využití glukózy Laktát se stává hlavním aerobním substrátem

73
stává se hlavním aerobním substrátem
Při intenzivní tělesné práci se stává laktát preferovaným energetickým zdrojem stává se hlavním aerobním substrátem tím se šetří krevní glukóza pro tkáně, které jsou na ní takřka výhradně závislé (především nervové buňky mozku)
")
74
Vzestup laktátu v krvi Preference laktátu jako energetického substrátu (oxidace a glykoneogeneze) G La La La G La G G G La La G G La G G La La

75
Laktát tedy poskytuje energii dvakrát (2 – 3 molekuly ATP)
nejdříve malé množství (2 – 3 molekuly ATP) rozkladem glukózy nebo glykogenu v cytoplazmě, po zpětné přeměně na pyruvát a rozložením v Krebsově cyklu nepoměrně větší množství (36 molekul ATP)
 rozkladem glukózy nebo glykogenu v cytoplazmě, po zpětné přeměně na pyruvát. a rozložením v Krebsově cyklu. nepoměrně větší množství (36 molekul ATP)")
76
glykolýza Nový směr = nová rychlost = aerobní zpracování laktátu
Krátkodobý zážeh = glykolýza

77
stává se hlavním aerobním substrátem
Při intenzivní tělesné práci se stává laktát preferovaným energetickým zdrojem stává se hlavním aerobním substrátem tím se šetří krevní glukóza pro tkáně, které jsou na ní takřka výhradně závislé (především nervové buňky mozku)
")
78
Lidský organismus má k dispozici relativně velké množství energie:
Zásobní cukr - glykogen (muž o hmotnosti 70 kg má asi 500 g glykogenu, z toho 400 g ve svalech a 100 g v játrech – poskytuje energii asi 2500 kcal). Glukóza v krvi (asi 20 g = 100 kcal) Tuk (asi kcal, tj. asi 80% všeho paliva v těle) Proteiny (asi kcal, tj. asi asi 18%) nejsou běžně využitelné
. Glukóza v krvi (asi 20 g = 100 kcal) Tuk (asi kcal, tj. asi 80% všeho paliva. v těle) Proteiny (asi kcal, tj. asi asi 18%) nejsou běžně využitelné.")
79
Svaly v klidu, při lehké práci Mozek spotřebuje v klidu
a v průběhu regenerace spotřebovávají mastné kyseliny (vznikají rozkladem neutrálních tuků - triglyceridy) Mozek spotřebuje v klidu asi % krevní glukózy většinu zbylé glukózy spotřebují v klidu červené krvinky
 Mozek spotřebuje v klidu. asi % krevní glukózy. většinu zbylé glukózy spotřebují v klidu. červené krvinky.")
80
Při tělesné práci se zvyšuje glykogenolýza V průběhu zotavení
a stoupá i spotřeba glukózy ve svalech (vytváří se z jaterního glykogenu) V průběhu zotavení je spotřebovaný jaterní glykogen okamžitě doplňován glukoneogenezí zatímco glykogenolýza je redukovaná
 V průběhu zotavení. je spotřebovaný jaterní glykogen. okamžitě doplňován glukoneogenezí. zatímco glykogenolýza je redukovaná.")
81
Tuky se dělí na mastné kyseliny (MK) triglyceridy (TG) fosfolipidy
steroly

82
Tuky MK jsou uhlíkové sloučeniny
s dlouhým řetězcem a sudým počtem uhlíků Nasycené (bez dvojné vazby mezi uhlíky) Nenasycené (s jednou nebo více dvojnými vazbami) Některé nenasycené MK nezbytná součást potravy (esenciální) (např. kyselina linoleová nebo kyselina linolová)
 Nenasycené (s jednou nebo více dvojnými vazbami) Některé nenasycené MK. nezbytná součást potravy (esenciální) (např. kyselina linoleová nebo kyselina linolová)")
83
Tuky Triglyceridy (TG) jsou sloučeniny glycerolu a tří MK
(nerozpustné ve vodě, ideální zásobárna energie) TG jsou rozkládány v tukových, svalových nebo jaterních buňkách v tenkém střevě v plazmatických lipoproteidech (nosiči cholesterolu a triglyceridů v plazmě) enzymem lipázou na glycerol a tři MK
 TG jsou rozkládány v tukových, svalových nebo jaterních buňkách. v tenkém střevě. v plazmatických lipoproteidech (nosiči cholesterolu. a triglyceridů v plazmě) enzymem lipázou. na glycerol a tři MK.")
84
Tuky MK přeměňují na acyly po prostoupení buněčnou membránou
MK jsou transportovány pomocí albuminu krví Před vstupem do buňky se MK přeměňují na acyly po prostoupení buněčnou membránou se dostávají pomocí karnitinu do mitochondrií

85
Tvorba acyl-CoA MK + ATP + CoA Acyl-CoA + AMP + PPi energie
acyl CoA syntetáza MK + ATP + CoA Acyl-CoA + AMP + PPi Mg2+

86
Acyl karnitin translokáza I II
Cytoplazma Vnitřní membrána Matrix acyl-CoA Karnitin Karnitin acyl-CoA Acyl karnitin translokáza I II acylkarnitin acylkarnitin CoA CoA

87
Tuky acetylkoenzym-A V mitochondriích jsou MK
dekarboxylovány a dehydrogenovány acetylkoenzym-A a vstupují do Krebsova cyklu

88
Acyl-CoA (C = n - 2) Acyl-CoA (C = n) FAD+ FADH2 dehydrogenace NAD+ NADH + H+ Acetyl-CoA C = 2 Acyl-CoA (C = n-2)
")
89
Alfa-ketoglutarát (5C)
Pyruvát (3C) NAD+ CO2 NADH + H+ Acetyl-CoA (2C) Oxalacetát (4C) Citrát (6C) NAD+ NADH + H+ Malát (4C) Izocitrát (6C) NAD+ CO2 Fumarát (4C) NADH + H+ FADH2 Alfa-ketoglutarát (5C) Sukcinát (4C) P FAD Sukcinyl-CoA (4C) CO2 GTP NAD+ NADH + H+ GDP
 NAD+ CO2. NADH + H+ Acetyl-CoA (2C) Oxalacetát (4C) Citrát (6C) NAD+ NADH + H+ Malát (4C) Izocitrát (6C) NAD+ CO2. Fumarát (4C) NADH + H+ FADH2. Alfa-ketoglutarát (5C) Sukcinát (4C) P. FAD. Sukcinyl-CoA (4C) CO2. GTP. NAD+ NADH + H+ GDP.")
90
Tuky Acetylkoenzym-A je energeticky bohatá sloučenina
(tzv. intermediát) na kterou se v mitochondriích rozkládají sacharidy i lipidy
 na kterou se v mitochondriích. rozkládají sacharidy i lipidy.")
91
Tuky 146 molekul ATP několikanásobně větší než ze sacharidů
Celkový energetický zisk z tuků několikanásobně větší než ze sacharidů Má-li MK např. 18 uhlíků (např. nasycená kyselina stearová) získáme oxidativní fosforylací jedné molekuly 146 molekul ATP
 získáme oxidativní fosforylací. jedné molekuly. 146 molekul ATP.")
92
Energetický zisk z beta-oxidace MK
8,5 n - 7 ATP n = počet atomů C Energetický zisk z beta-oxidace MK

93
umožňují transport triglyceridů a cholesterolu krví
Lipoproteiny umožňují transport triglyceridů a cholesterolu krví chylomikrony chylomikronové zbytky lipoproteiny o velmi nízké hustotě (VLDL) lipoproteiny o stření hustotě (IDL) lipoproteiny o nízké hustotě (LDL) lipoproteiny o vysoké hustotě (HDL) Zevní dráha Vnitřní dráha Chylomikrony Chylomikronové zbytky JÁTRA HDL PERIFERIE VLDL IDL LDL
 lipoproteiny o stření hustotě (IDL) lipoproteiny o nízké hustotě (LDL) lipoproteiny o vysoké hustotě (HDL) Zevní dráha. Vnitřní dráha. Chylomikrony. Chylomikronové zbytky. JÁTRA. HDL. PERIFERIE. VLDL. IDL. LDL.")
94
LDL poskytuje tkáním cholesterol CHOLESTEROL
(vyskytuje se jen v živočišné potravě) (syntetizuje se hlavně v játrech) CHOLESTEROL základní a nezbytná součást buněčných membrán vznikají z něj steroidní hormony a žlučové kyseliny Nadbytek cholesterolu v krvi ukládání cholesterolu pod endotelem tepen vznik aterosklerotického plátu (zužuje průsvit tepen)
 (syntetizuje se hlavně v játrech) CHOLESTEROL. základní a nezbytná součást buněčných membrán. vznikají z něj steroidní hormony a žlučové kyseliny. Nadbytek cholesterolu v krvi. ukládání cholesterolu pod endotelem tepen. vznik aterosklerotického plátu (zužuje průsvit tepen)")
95
HDL Poměr mezi LDL a HDL (nad 3,5)
transport cholesterolu z periferie do jater Poměr mezi LDL a HDL určuje převažující směr transportu cholesterolu Při zvýšeném poměru LDL/HDL (nad 3,5) se zvyšuje riziko vzniku aterosklerózy
 se zvyšuje riziko vzniku aterosklerózy.")
96
AMINOKYSELINY (AMK) tzv. esenciální AMK
základní složka bílkovin, peptidů a polypeptidů z 20 základních AMK může být 12 syntetizováno v lidském těle zbylé AMK musí člověk přijmout v potravě tzv. esenciální AMK Zdroje AMK bílkoviny z potravy opotřebované bílkoviny z tkání malé množství vzniká při přeměně sacharidů

97
AMINOKYSELINY (AMK) Bílkoviny Základní stavební látky pro tvorbu
Počet aminokyselin tvořících bílkovinu velmi rozdílný inzulín = 51 AMK protein ze sójových bobů = 4500 AMK Bílkoviny nezbytné pro budování nových buněk reparaci starých buněk a tkání Základní stavební látky pro tvorbu enzymů plazmatických bílkovin protilátek některých hormonů kreatinu atd. Bílkoviny se mohou přeměňovat na sacharidy

98
AMINOKYSELINY (AMK) zisk energie
Část aminokyselin se může odbourávat (deaminovat) na jednodušší látky zisk energie Aminové skupiny se odštěpují ve formě toxického amoniaku v jaterních buňkách se mění na močovinu krví do ledvin vylučuje se močí z těla Uhlíkaté zbytky aminokyselin se začleňují do Krebsova cyklu (podobně jako sacharidy a mastné kyseliny) dekarboxylovány a dehydrogenovány a rozkládají se na CO2, H2O a energii
 na jednodušší látky. zisk energie. Aminové skupiny se odštěpují. ve formě toxického amoniaku. v jaterních buňkách se mění na močovinu. krví do ledvin. vylučuje se močí z těla. Uhlíkaté zbytky aminokyselin. se začleňují do Krebsova cyklu. (podobně jako sacharidy a mastné kyseliny) dekarboxylovány a dehydrogenovány. a rozkládají se na CO2, H2O a energii.")
99
HDL Poměr mezi LDL a HDL (nad 3,5)
transport cholesterolu z periferie do jater Poměr mezi LDL a HDL určuje převažující směr transportu cholesterolu Při zvýšeném poměru LDL/HDL (nad 3,5) se zvyšuje riziko vzniku aterosklerózy
 se zvyšuje riziko vzniku aterosklerózy.")
100
Čerpání energie závisí na
Tělesná práce Čerpání energie závisí na intenzitě zatížení trvání práce druhu zátěže trénovanosti dietě prostředí pohlaví

101
Glykogenolýza v prvních dvou sekundách
Při maximální intenzitě zatížení v prvních dvou sekundách makroergní fosfáty (ATP a CP) Glykogenolýza dosahuje nejvyššího zastoupení kolem 5. sekundy maximální práce Mezi 60. a 75. sekundou maximální práce začíná dominovat aerobní fosforylace Podíl anaerobní glykolýzy a makroergních fosfátů na resyntéze ATP po 4 minutách maximální práce asi 20 %
 Glykogenolýza. dosahuje nejvyššího zastoupení. kolem 5. sekundy maximální práce. Mezi 60. a 75. sekundou maximální práce. začíná dominovat aerobní fosforylace. Podíl anaerobní glykolýzy a makroergních fosfátů. na resyntéze ATP po 4 minutách maximální práce. asi 20 %")
102
Podíl jednotlivých energetických systémů
na hrazení celkového energetického výdeje při různém trvání maximální zátěže. 100 80 ATP-CP 60 40 Příspěvek k celkovému energetickému výdeji (%) 20 50 100 150 200 250 300 Trvání práce (s)
 Trvání práce (s)")
103
Glykogenolýza v prvních dvou sekundách
Při maximální intenzitě zatížení v prvních dvou sekundách makroergní fosfáty (ATP a CP) Glykogenolýza dosahuje nejvyššího zastoupení kolem 5. sekundy maximální práce Mezi 60. a 75. sekundou maximální práce začíná dominovat aerobní fosforylace Podíl anaerobní glykolýzy a makroergních fosfátů na resyntéze ATP po 4 minutách maximální práce asi 20 %
 Glykogenolýza. dosahuje nejvyššího zastoupení. kolem 5. sekundy maximální práce. Mezi 60. a 75. sekundou maximální práce. začíná dominovat aerobní fosforylace. Podíl anaerobní glykolýzy a makroergních fosfátů. na resyntéze ATP po 4 minutách maximální práce. asi 20 %")
104
Podíl jednotlivých energetických systémů
Gastin PB. Energy system interaction and relative contribution during maximal exercise. Sports Med 2001; 31 (10): Podíl jednotlivých energetických systémů na hrazení celkového energetického výdeje při různém trvání maximální zátěže. 100 80 ATP-CP 60 glykolysa 40 Příspěvek k celkovému energetickému výdeji (%) 20 50 100 150 200 250 300 Trvání práce (s)
: Podíl jednotlivých energetických systémů. na hrazení celkového energetického výdeje při různém trvání maximální zátěže ATP-CP. 60. glykolysa. 40. Příspěvek k celkovému energetickému výdeji (%) Trvání práce (s)")
105
Glykogenolýza v prvních dvou sekundách
Při maximální intenzitě zatížení v prvních dvou sekundách makroergní fosfáty (ATP a CP) Glykogenolýza dosahuje nejvyššího zastoupení kolem 5. sekundy maximální práce Mezi 60. a 75. sekundou maximální práce začíná dominovat aerobní fosforylace Podíl anaerobní glykolýzy a makroergních fosfátů na resyntéze ATP po 4 minutách maximální práce asi 20 %
 Glykogenolýza. dosahuje nejvyššího zastoupení. kolem 5. sekundy maximální práce. Mezi 60. a 75. sekundou maximální práce. začíná dominovat aerobní fosforylace. Podíl anaerobní glykolýzy a makroergních fosfátů. na resyntéze ATP po 4 minutách maximální práce. asi 20 %")
106
35-75 s 0-30 30-60 60-90 90-120

107
Glykogenolýza v prvních dvou sekundách
Při maximální intenzitě zatížení v prvních dvou sekundách makroergní fosfáty (ATP a CP) Glykogenolýza dosahuje nejvyššího zastoupení kolem 5. sekundy maximální práce Mezi 60. a 75. sekundou maximální práce začíná dominovat aerobní fosforylace Podíl anaerobní glykolýzy a makroergních fosfátů na resyntéze ATP po 4 minutách maximální práce asi 20 %
 Glykogenolýza. dosahuje nejvyššího zastoupení. kolem 5. sekundy maximální práce. Mezi 60. a 75. sekundou maximální práce. začíná dominovat aerobní fosforylace. Podíl anaerobní glykolýzy a makroergních fosfátů. na resyntéze ATP po 4 minutách maximální práce. asi 20 %")
108
Podíl jednotlivých energetických systémů
Gastin PB. Energy system interaction and relative contribution during maximal exercise. Sports Med 2001; 31 (10): Podíl jednotlivých energetických systémů na hrazení celkového energetického výdeje při různém trvání maximální zátěže. 100 aerobní fosforylace 80 ATP-CP Podíly anaerobní glykolýzy a systému ATP-CP na resyntéze ATP po 4 minutách maximální práce činí jen asi 20 % 60 glykolysa 40 Příspěvek k celkovému energetickému výdeji (%) 20 50 100 150 200 250 300 Trvání práce (s)
: Podíl jednotlivých energetických systémů. na hrazení celkového energetického výdeje při různém trvání maximální zátěže aerobní fosforylace. 80. ATP-CP. Podíly anaerobní glykolýzy a systému ATP-CP na resyntéze ATP po 4 minutách maximální práce činí jen asi 20 % 60. glykolysa. 40. Příspěvek k celkovému energetickému výdeji (%) Trvání práce (s)")
109
Dlouhotrvající práce - využití sacharidů klesá
množství glykogenu ve svalech i v játrech klesá stoupá dostupnost alternativních zdrojů (zejména mastných kyselin a glukózy) Při intenzitě zatížení 60% - 75% VO2 max rozklad glykogenu zejména ve svalových vláknech I. typu (červená vlákna) méně ve vláknech II. typu (bílá vlákna) Při zvyšování intenzity až ke 100 % VO2 max dominuje zapojení vláken II. typu Ke zvýšení glykogenolýzy dochází i v nepracujících svalech uvolněný laktát se stává preferovaným zdrojem glukoneogeneze nebo oxidace v pracujících svalech
 Při intenzitě zatížení 60% - 75% VO2 max. rozklad glykogenu. zejména ve svalových vláknech I. typu (červená vlákna) méně ve vláknech II. typu (bílá vlákna) Při zvyšování intenzity až ke 100 % VO2 max. dominuje zapojení vláken II. typu. Ke zvýšení glykogenolýzy dochází i v nepracujících svalech. uvolněný laktát se stává preferovaným zdrojem. glukoneogeneze nebo oxidace v pracujících svalech.")
Podobné prezentace












>")

.>")




