Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Genetika reverzní a přímá mapování a klasické přístupy ke studiu funkce rostlinných genů

2
Cíle genetického snažení:
pochopení funkce genů Pochopit, jak je prostřednictvím koordinované aktivity genů tvořen komplexní organismus šlechtění Získat rostliny (organismy) s vylepšenými vlastnostmi či novými kombinacemi vlastností
 s vylepšenými vlastnostmi či novými kombinacemi vlastností.")
3
Pojmy Gen (vloha) vztah genotyp-fenotyp (další vlivy)
vztahy mezi geny – genové interakce (epistáze – metabolické a signální dráhy) jde o funkční vztahy mezi produkty genů! Alela (forma genu) vztahy mezi alelami – (ko)dominance vznik alel – mutace Lokus (místo na chromozómu) genová vazba (vzdálenost genů) genetické mapy x fyzické mapy
 jde o funkční vztahy mezi produkty genů! Alela (forma genu) vztahy mezi alelami – (ko)dominance. vznik alel – mutace. Lokus (místo na chromozómu) genová vazba (vzdálenost genů) genetické mapy x fyzické mapy.")
4
Genetické mapy - kompletní sekvence, geny … - markery
- vzdálenosti na genetické a fyzické mapě si neodpovídají (různá pravděpodobnost rekombinace - cM) Jaké sekvence lze čekat ve zvýrazněných oblastech?
 Jaké sekvence lze čekat ve zvýrazněných oblastech")
5
klasická (přímá) a reverzní
Genetika klasická (přímá) a reverzní Přímá – od znaku (fenotypového projevu mutace) k identifikaci příslušného genu Reverzní – od genu k fenotypovému projevu (studium funkce vybraného genu jeho mutagenezí, modulací exprese, atp.) Pro obě cesty je společná příprava a studium mutantů!
 a reverzní. Přímá – od znaku (fenotypového projevu mutace) k identifikaci příslušného genu. Reverzní – od genu k fenotypovému projevu (studium funkce vybraného genu jeho mutagenezí, modulací exprese, atp.) Pro obě cesty je společná příprava a studium mutantů!")
6
Mutageneze - mutanti jsou nástrojem přímé i reverzní genetiky
Mutageneze - mutanti jsou nástrojem přímé i reverzní genetiky přímá: hledáme projev v populaci mutantů reverzní: cílená mutace studovaného genu Klasicky: M. chemická – EMS (ethyl metansulfonát; bodové mutace: 6-O- ethylguanosin páruje s T) M. fyzikální – RTG, gama ... (zpravidla krátké delece) široké spektrum ovlivnění genové funkce (regulace, interakce, …) někdy projev i v heterozygotním stavu (dominantní alela) více odpovídá přirozeným mutacím ALE obtížná a časově náročná identifikace mutovaného genu
 M. fyzikální – RTG, gama ... (zpravidla krátké delece) široké spektrum ovlivnění genové funkce (regulace, interakce, …) někdy projev i v heterozygotním stavu (dominantní alela) více odpovídá přirozeným mutacím. ALE obtížná a časově náročná identifikace mutovaného genu.")
7
Mutageneze Moderně: M. inserční – začlenění úseků DNA
(T-DNA agrobaktéria, transpozónu - náhodné inzerce!) - umožňují relativně přímé a rychlé určení místa začlenění (místo mutace označeno začleněnou DNA = tag) - různé variace pro cílenější izolace genů
 - umožňují relativně přímé a rychlé určení místa začlenění. (místo mutace označeno začleněnou DNA = tag) - různé variace pro cílenější izolace genů.")
8
Izolace genů na základě fenotypového projevu
inzerční mutageneze (výběr na základě změny fenotypu) - inaktivace genu v důsledku začlenění úseku DNA - T-DNA tagging - transposon tagging - recesivní aktivační mutageneze (výběr na základě změny fenotypu) transformace promotorem (enhancerem), který může v místě začlenění aktivovat expresi jinak neaktivního genu promotor a enhancer-trap linie (výběr dle exprese reportéru) - transformace T-DNA s reportérovým genem bez promotoru či s minimálním promotorem původní gen (wild type, nativní)
 - inaktivace genu v důsledku začlenění úseku DNA. - T-DNA tagging. - transposon tagging. - recesivní. aktivační mutageneze (výběr na základě změny fenotypu) transformace promotorem (enhancerem), který může v místě začlenění aktivovat expresi jinak neaktivního genu. promotor a enhancer-trap linie (výběr dle exprese reportéru) - transformace T-DNA s reportérovým genem bez promotoru či s minimálním promotorem. původní gen (wild type, nativní)")
9
Identifikace mutovaného genu
Jak identifikovat mutovaný gen mezi desetitisíci nemutovaných? (či mutovanými bez fenotypového projevu) Bodové mutace a delece: 1) - na základě segregační analýzy (genetické mapy) – viz dále - chromosom walking, sekvenování (zdlouhavé, náročné a nákladné!) viz dále s využitím NGS (rychlejší, nákladné) - bez nutnosti znát výchozí genom!!! - směsné vzorky (zpětné křížení) - porovnávání frekvencí podobných oligomerů Nordström et al. Nature Biotech.2013
 Bodové mutace a delece: 1) - na základě segregační analýzy. (genetické mapy) – viz dále. - chromosom walking, sekvenování. (zdlouhavé, náročné a nákladné!) viz dále. s využitím NGS (rychlejší, nákladné) - bez nutnosti znát výchozí genom!!! - směsné vzorky (zpětné křížení) - porovnávání frekvencí podobných. oligomerů. Nordström et al. Nature Biotech")
10
Identifikace mutovaného genu
u T-DNA nebo transpozónem taggovaných linií sekvenací úseků sousedících s inzercí (málo templátu pro přímé sekvenování!) TAIL PCR (Thermal Asymmetric InterLaced PCR) adaptorová PCR plasmid rescue iPCR
 TAIL PCR (Thermal Asymmetric InterLaced PCR) adaptorová PCR. plasmid rescue. iPCR.")
11
TAIL PCR: Adaptorová PCR:
SP1 SP2 SP3 AP AP SP1 AP AP SP2 AP SP3 AP 1. tři následné PCR (optimalizovaná Ta) s SP1-3 a vždy stejným AP 2. sekvenace produktu SP1-3: specifické primery na vloženou DNA AP: arbitrary (degenerovaný) primer - několik typů, vysoká P nasednutí poblíž vložené DNA Adaptorová PCR: štěpení (E) ligace adaptorů 2-3 PCR (spec. adapt. primer + spec. primery k vložené DNA) 4. sekvenace produktu E E E E SP1 SP2 SP SAP
 s SP1-3 a vždy stejným AP. 2. sekvenace produktu. SP1-3: specifické primery na vloženou DNA. AP: arbitrary (degenerovaný) primer. - několik typů, vysoká P nasednutí poblíž vložené DNA. Adaptorová PCR: štěpení (E) ligace adaptorů. 2-3 PCR (spec. adapt. primer. + spec. primery k vložené DNA) 4. sekvenace produktu. E. E. E. E. SP1 SP2 SP3 SAP.")
12
Plasmid rescue: Inverzní PCR: plasmid štěpení (E)
ori bla/nptIII E E ori bla/nptIII štěpení (E) cirkularizace (ligace) transformace E.coli (ori, R) pomnožení, sekvenace E plasmid Inverzní PCR: E E štěpení (E) cirkularizace (ligace) PCR sekvenace E E
 cirkularizace (ligace) transformace E.coli (ori, R) pomnožení, sekvenace. E. plasmid. Inverzní PCR: E. E. štěpení (E) cirkularizace (ligace) PCR. sekvenace. E. E.")
13
Kolekce inserčních mutantů
Arabidopsis - obří kolekce charakterizovaných inserčních mutantů - snadná transformovatelnost - snadná identifikace a zamapování mutace (malý a známý genom)
")
14
Reverzní genetika 1) analýzy mutantů s inzercí v určitém genu (mutace často knock-out, knock-down, ale i aktivační) veřejně dostupné kolekce mutantů na různých místech genomu (mnohočetné inserce ve většině genů Arabidopsis thaliana) – výběr in silico, objednání semen rostlin s inzercí v určitém genu Gen Gen Gen3 ? číslo linie = místa inzerce T-DNA v jednotlivých mutantních liniích (pro inaktivaci vhodné inzerce do počátečních exonů, 5’UTR, minimálního promotoru - nutné ověřit expresi!)
 – výběr in silico, objednání semen rostlin s inzercí v určitém genu. Gen1 Gen2 Gen číslo linie. = místa inzerce T-DNA v jednotlivých mutantních liniích. (pro inaktivaci vhodné inzerce do počátečních exonů, 5’UTR, minimálního promotoru. - nutné ověřit expresi!)")
15
Hledání inzerčního mutanta Arabidopsis
ve zvoleném genu (př. webového rozhraní) GENY T-DNA
 GENY. T-DNA.")
16
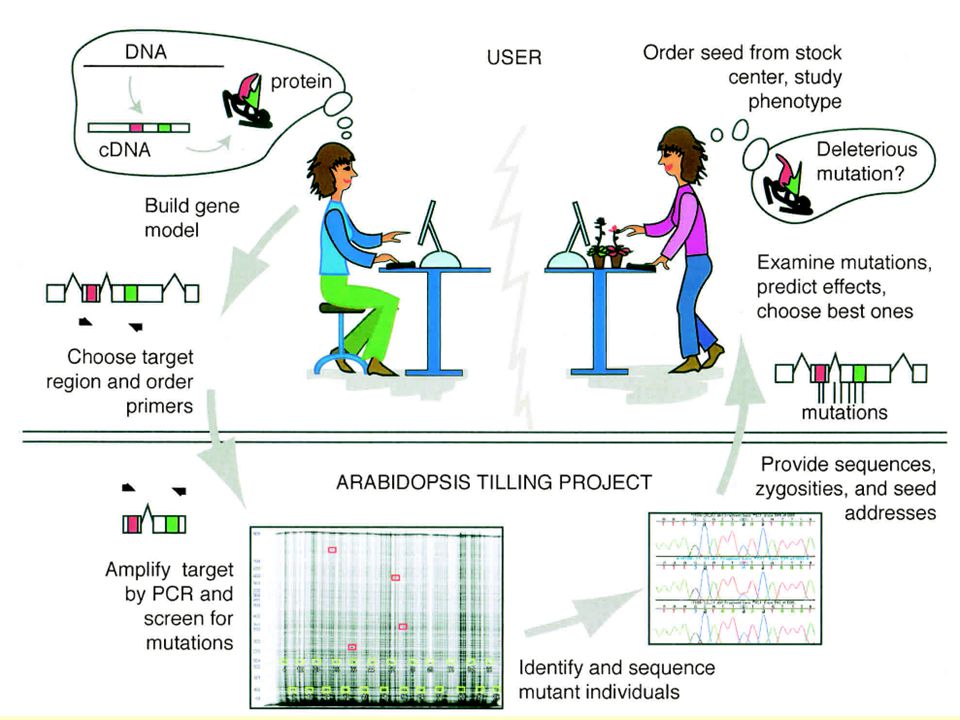
Reverzní genetika 2) TILLING: získání rostlin s náhodnými bodovými mutacemi ve vybraném genu
(Targeting induced local lesions in genomes) Bodové mutace! (potenciálně změny regulace, interakce, …) Princip: - náhodná indukce bodových mutací (EMS) - izolace DNA z velkého počtu linií - hledání linií s mutacemi v cílovém genu pomocí PCR a heteroduplexní analýzy
 Bodové mutace! (potenciálně změny regulace, interakce, …) Princip: - náhodná indukce bodových mutací (EMS) - izolace DNA z velkého počtu linií. - hledání linií s mutacemi v cílovém genu pomocí PCR a heteroduplexní analýzy.")
17
TILLING – detekce mutací
amplifikace cílového fragmentu z DNA mutovaných linií (koncově značené primery) 2. denaturace a reasociace s wt DNA (nekomplementarita v místě mutace) 3. štěpení ss heteroduplexu + elektroforéza + visualizace koncově značených fragmentů
 2. denaturace a reasociace s wt DNA. (nekomplementarita v místě mutace) 3. štěpení ss heteroduplexu + elektroforéza + visualizace koncově značených fragmentů.")
18
TILLING – strategie screeningu

20
Reverzní genetika 3) Cílená bodová modifikace genů (mimo genom)
Inaktivace genu a vnesení další upravené kopie izolace genu (optimálně s nativním promotorem) cílená mutageneze pomocí primerů (ex vivo) izolace homozygotního knock-out mutanta v daném genu (je-li mutace letální, pak heterozygotního mutanta) transformace mutanta upraveným genem
 cílená mutageneze pomocí primerů (ex vivo) izolace homozygotního knock-out mutanta v daném genu. (je-li mutace letální, pak heterozygotního mutanta) transformace mutanta upraveným genem.")
21
Reverzní genetika 4) Cílené modifikace genů v genomu
Oligonucleotide directed mutagenesis Indukce DSB (dvouvláknového zlomu DNA) – lokální mutace ZFN, TALEN zlom v konkrétním místě genomu štěpení chimerickými (fúzními) proteiny s nukleázovou doménou restriktázy Fok I designovatelné DNA vazebné domény: Zn-finger, DNA vazebná doména transcription activator-like effektorů Xanthomonas CRISPR/CAS9 místo štěpení (CAS9) rozpoznáno pomocí guide RNA
 – lokální mutace. ZFN, TALEN. zlom v konkrétním místě genomu štěpení chimerickými (fúzními) proteiny s nukleázovou doménou restriktázy Fok I. designovatelné DNA vazebné domény: Zn-finger, DNA vazebná doména transcription activator-like effektorů Xanthomonas. CRISPR/CAS9. místo štěpení (CAS9) rozpoznáno pomocí guide RNA.")
22
Reverzní genetika 5) Řízená modulace exprese
zvýšení množství proteinu = vnesení genu pod silným (konstitutivním) promotorem (ektopická exprese, overexprese, gain-of-function mutation) snížení množství proteinu pomocí RNAi cílenou mutagenezí (in situ) (umlčení, silencing, knock-down, knock-out, loss-of-function)
 promotorem. (ektopická exprese, overexprese, gain-of-function mutation) snížení množství proteinu. pomocí RNAi. cílenou mutagenezí (in situ) (umlčení, silencing, knock-down, knock-out, loss-of-function)")
23
Snížení množství proteinu
navozením RNA interference: 1) exprese genu v antisense orientaci (za promotor vložen opačně) 2) exprese vlásenkového konstruktu - IR (např. sense-intron-antisense) proti transkribované (PTGS) či promotorové (TGS) sekvenci 3) exprese genu bez terminátoru (dsRNA díky aktivitě RdRP) 4) exprese amiRNA (artificial miRNA) intermolekulární párování 1) + 2) intramolekulární párování RdRP syntéza komplement. vlákna k transkriptu 3) X tvorba sRNA, štěpení DCL = tvorba siRNA, specifická degradace mRNA či cílené metylace DNA promotoru
 exprese genu v antisense orientaci (za promotor vložen opačně) 2) exprese vlásenkového konstruktu - IR (např. sense-intron-antisense) proti transkribované (PTGS) či promotorové (TGS) sekvenci. 3) exprese genu bez terminátoru (dsRNA díky aktivitě RdRP) 4) exprese amiRNA (artificial miRNA) intermolekulární párování. 1) + 2) intramolekulární. párování. RdRP. syntéza komplement. vlákna k transkriptu. 3) X. tvorba sRNA, štěpení DCL = tvorba siRNA, specifická degradace mRNA či cílené metylace DNA promotoru.")
24
Reverzní genetika 5) Analýzy aktivity promotoru
fúze promotoru analyzovaného genu (zpravidla 2 kbp před TSS) s reportérovým genem („transkripční fúze“): Slouží k určení - tkáňové specifity (popř. síla promotoru) - vývojové specifity změn exprese v závislosti na faktorech vnějšího (vnitřního) prostředí Potřeba ověřit i jinými postupy - riziko artefaktů! reportérový gen P gen T Nelze měnit v genomu, vkládá se další (upravená) kopie genu (výměna za reportérový gen či přidání reportérového genu)
 s reportérovým genem („transkripční fúze ): Slouží k určení. - tkáňové specifity (popř. síla promotoru) - vývojové specifity. změn exprese v závislosti na faktorech vnějšího (vnitřního) prostředí. Potřeba ověřit i jinými postupy - riziko artefaktů! reportérový gen. P gen T. Nelze měnit v genomu, vkládá se další (upravená) kopie genu. (výměna za reportérový gen či přidání reportérového genu)")
25
Reportérové geny gen produkt substrát stanovení
kódují enzymy, jejichž aktivitu je možné vizualizovat, či jsou vizualizovatelné přímo samy (fluorescence) stanovení kvantitativní a/nebo kvalitativní použití pro analýzy aktivity promotorů a fúzních proteinů, promotor-trap mutagenezi, … Problémy: pozadí (autofluorescence, enzymová aktivita v pletivu) stabilita proteinu (maskuje rychlé změny v aktivitě promotoru) gen produkt substrát stanovení gusA b-glukuronidáza (E. coli) MUG fluorescence X-gluc histochemicky luc luciferáza (světluška) luciferin luminiscence gfp green fluorescent protein xxx fluorescence (medůza)
 stanovení kvantitativní a/nebo kvalitativní. použití pro analýzy aktivity promotorů a fúzních proteinů, promotor-trap mutagenezi, … Problémy: pozadí (autofluorescence, enzymová aktivita v pletivu) stabilita proteinu (maskuje rychlé změny v aktivitě promotoru) gen produkt substrát stanovení. gusA b-glukuronidáza (E. coli) MUG fluorescence. X-gluc histochemicky. luc luciferáza (světluška) luciferin luminiscence. gfp green fluorescent protein xxx fluorescence. (medůza)")
26
Fluorescenční proteiny GFP, DsRed, mCherry, …
- unikátní nástroje pro vitální barvení buněk a buněčných struktur, - fluorescenční značky kódované genem původ z mořských láčkovců původní geny modifikovány: - fluorescenční vlastnosti, pH, - kódování, sestřih, stabilita, …. Aequoria victoria Barevné varianty GFP a DsRed

27
GUS stanovení kvalitativní (X-gluc5-bromo-4-chloro-3-indolyl glucuronide) po inkubaci substrát štěpen na bezbarvý intermediát, jehož oxidací vzniká modrý nerozpustný precipitát in situ pozadí ve většině pletiv malé produkt málo difunduje - akumuluje se v buňce, kde k expresi genu dochází většina technik pro buňky letální (fixovaný materiál) stanovení kvantitativní (MUG: 4-methylumbelliferyl-beta-D-glucuronide) extrakce GUS enzymu, fluorimetrické stanovení velmi citlivá metoda, malé pozadí
 stanovení kvantitativní (MUG: 4-methylumbelliferyl-beta-D-glucuronide) extrakce GUS enzymu, fluorimetrické stanovení. velmi citlivá metoda, malé pozadí.")
28
Fúze promotoru s reportérovým genem
pro GFP a GUS Arabidopsis thaliana

29
Reverzní genetika 6) Tvorba fúzního proteinu s GFP
- zrušení stop kodónu studovaného genu a připojení GFP genu ve čtecí fázi (translační fúze) – signální sekvence, folding (!)
 – signální sekvence, folding (!)")
30
Tvorba fúzního proteinu s GFP
- studium lokalizace proteinů, proteinových interakcí, - sledování různých pochodů v živé buňce – značení buněčných struktur … Golgiho komplex chromozómy mikrotubuly

31
Přímá genetika: Výběr mutantů na základě fenotypového projevu – vlastností (jejich změn)
Biochemické Barevné fotosynt. barviva – albino, chlorina, variegated pigmenty Vývojové ontogeneze vegetativních či generativních org. regulace shootmeristemless agamous

32
Mutantní screeny - podmínky - ošetření

33
Přímá a reverzní genetika u Arabidopsis
určení místa mutace + Tilling – „prohlížení“ necharakterizované kolekce linií pomocí PCR a reasociace

34
Přímá genetika – identifikace mutovaného genu: stejné fenotypové projevy mohou mít různé příčiny! „existuje mnoho způsobů, jak špatně postavit dům“ - před dalšími analýzami je vhodné testovat, zda mutace nejsou ve stejném genu (cistronu) Cis-trans (komplementační) test – křížení homozygotních mutantů: Co lze očekávat v F1 generaci? F1 mutantní fenotyp = stejný lokus (alelické mutace) F1 wild type fenotyp = 2 geny (v obou mutovaných lokusech jsou rostliny heterozygotní a mutace se neprojeví)
 test. – křížení homozygotních mutantů: Co lze očekávat v F1 generaci F1 mutantní fenotyp = stejný lokus (alelické mutace) F1 wild type fenotyp = 2 geny (v obou mutovaných lokusech jsou rostliny heterozygotní a mutace se neprojeví)")
35
Identifikace mutovaného genu
(u bodových mutantů vybraných na základě fenotypového projevu) či neznámého genu (s fenotypovým projevem) Celogenomovým sekvenováním (mutace - viz dříve) Na základě genetické mapy mapování - (ko)segregační analýza - nalezení přibližné pozice genu v genetické mapě - na základě genové vazby s genetickými markery (znaky, které vykazují polymorfismus = jsou rozdílné mezi rodičovskými genotypy) 2. nalezení konkrétní sekvence (mutace/ genu) - chromosom walking - sekvenace, srovnání s WT sekvencí
 či neznámého genu (s fenotypovým projevem) Celogenomovým sekvenováním (mutace - viz dříve) Na základě genetické mapy. mapování - (ko)segregační analýza. - nalezení přibližné pozice genu v genetické mapě. - na základě genové vazby s genetickými markery. (znaky, které vykazují polymorfismus = jsou rozdílné mezi rodičovskými genotypy) 2. nalezení konkrétní sekvence (mutace/ genu) - chromosom walking. - sekvenace, srovnání s WT sekvencí.")
36
Vazba genů x rekombinace při meiose - crossing-over
1. Cytologic event Parental chromosomes Meiosis Without Crossing-over Gametes 1 2 3 4 Not recombinant Recombinant 2. Genetic result Genotype (heterozygous Aa and Bb ) Locus A Locus B ( same as parental genotype ) ( new ) Síla vazby ~ vzdálenosti, stavu chromatinu = frekvence crossing overu (poměr rekombinantních gamet) Pro mapování je třeba dostatek genetických markerů = polymorfních lokusů.
 Locus A. Locus B. ( same as. parental genotype ) ( new ) Síla vazby ~ vzdálenosti, stavu chromatinu. = frekvence crossing overu (poměr rekombinantních gamet) Pro mapování je třeba dostatek genetických markerů = polymorfních lokusů.")
37
Typy genetických markerů Marker = znak se známou (či snadno dohledatelnou!) polohou v genetické mapě, který vykazuje polymorfismus = je rozdílný mezi rodičovskými genotypy! – výhodné porovnávat např. různé ekotypy (vysoká frekvence odlišností) Morfologické (omezené množství) Molekulární DNA markery – detekovatelné rozdíly v sekvenci DNA (se známou či určitelnou polohou v genomu) ostatní: např. isozymy (limitace - asi jen 30 použitelných enzymatických systémů, stejná aktivita, různá mobilita na elektroforéze – nativní elektroforéza, aktivitní assay)
 Molekulární. DNA markery – detekovatelné rozdíly v sekvenci DNA (se známou či určitelnou polohou v genomu) ostatní: např. isozymy (limitace - asi jen 30 použitelných enzymatických systémů, stejná aktivita, různá mobilita na elektroforéze – nativní elektroforéza, aktivitní assay)")
38
Základní sada genetických markerů
Arabidopsis thaliana 2-3 na každém chromozomálním raménku

39
Genetické mapování lze začít nalezením přibližné polohy mutace v genomu
1) křížení homozygotního mutanta s indikátorovými liniemi s jasným fenotypovým projevem (F1F2) Gene symbol Name Phenotype Location (chr. - cM) an-1 angustifolia narrow leaves, crinkled siliques 1-55.2 ap1-1 apetala no petals 1-99.3 py pyrimidine requiring white leaves, restored by pyrimidine 2-49.1 er-1 erecta compact inflorescence, blunt siliques 2-43.5 hy2-1 long hypocotyl elongated hypocotyl, slender 3-11.5 gl1-1 glabra no trichomes 3-46.2 bp-1 brevipedicellus short pedicels, siliques bent downwards, short plant 4-15.0 cer2-2 eceriferum bright green stems, siliques bent downwards, short plant 4-51.9 ms1-1 male sterile no siliques 5-2.5 tt3-1 transparent testa yellow seeds, no anthocyanin 5-57.4
 křížení homozygotního mutanta s indikátorovými liniemi s jasným fenotypovým projevem (F1F2) Gene symbol. Name. Phenotype. Location (chr. - cM) an-1. angustifolia. narrow leaves, crinkled siliques ap1-1. apetala. no petals py. pyrimidine requiring. white leaves, restored by pyrimidine er-1. erecta. compact inflorescence, blunt siliques hy2-1. long hypocotyl. elongated hypocotyl, slender gl1-1. glabra. no trichomes bp-1. brevipedicellus. short pedicels, siliques bent downwards, short plant cer2-2. eceriferum. bright green stems, siliques bent downwards, short plant ms1-1. male sterile. no siliques tt3-1. transparent testa. yellow seeds, no anthocyanin")
40
Schéma kosegregační analýzy
b A b A b F1 (heterozygot) a B a B a B c c c C C C A b a B C c gamety gamety P1 (homozygot) P2 (homozygot) gamety A a B – úplná vazba! A a C – bez vazby A b A b a B a B C c C c F2 generace F2 - úplná vazba: AB:Ab:aB:ab 2:1:1:0 F2 – bez vazby: AC:Ac:aC:ac = 9:3:3:1 A b A b a B a B a B A b A A A A A A A a A a A a a a a a a a C c c c C C C c c c C C C C c C C C c c
 a. B. a. B. a. B. c. c. c. C. C. C. A. b. a. B. C. c. gamety. gamety. P1. (homozygot) P2. (homozygot) gamety. A a B – úplná vazba! A a C – bez vazby. A. b. A. b. a. B. a. B. C. c. C. c. F2 generace. F2 - úplná vazba: AB:Ab:aB:ab. 2:1:1:0. F2 – bez vazby: AC:Ac:aC:ac = 9:3:3:1. A. b. A. b. a. B. a. B. a. B. A. b. A. A. A. A. A. A. A. a. A. a. A. a. a. a. a. a. a. a. C. c. c. c. C. C. C. c. c. c. C. C. C. C. c. C. C. C. c. c.")
41
Segregace v F2 generaci F1 = XxYy
P: XXyy (pouze gamety Xy) x xxYY (pouze gamety xY) F1 = XxYy méně rekombinantních gamet XY a xy, při absolutní vazbě chybí v případě vazby: gamety XY (0.5) Xy xY xy (0.5) XXYY XY (0.25) XXYy XxYY XxYy XXyy XY Xxyy Xy (0.5) xxYY xxYy xY (0.5) Xy (0. 5) xY (0. 5) xxyy xy (0.25) genotyp fenotyp Vzdálenost genů: 2*0,5/(2*1+2*0,5) = 0,33 = 33 cM (= freq. rekom. gamet) 2*√ freq. xy fenot. A.t. 1 Mbp~4 cM (tj. 1 xy z 2500 rostlin) X,Y bez vazby na stejném raménku Fenotypové štěpení: 9:3:3:1 (XY:Xy:xY:xy) 4,75:2:2:0,25 6.25 % xy % xy Frekv. fenotypu xy:
 x xxYY (pouze gamety xY) F1 = XxYy. méně rekombinantních gamet XY a xy, při absolutní vazbě chybí. v případě vazby: gamety. XY (0.5) Xy. xY. xy (0.5) XXYY. XY (0.25) XXYy. XxYY. XxYy. XXyy. XY. Xxyy. Xy (0.5) xxYY. xxYy. xY (0.5) Xy (0. 5) xY (0. 5) xxyy. xy (0.25) genotyp. fenotyp. Vzdálenost genů: 2*0,5/(2*1+2*0,5) = 0,33 = 33 cM. (= freq. rekom. gamet) 2*√ freq. xy fenot. A.t. 1 Mbp~4 cM. (tj. 1 xy z 2500 rostlin) X,Y bez vazby na stejném raménku. Fenotypové štěpení: 9:3:3:1 (XY:Xy:xY:xy) 4,75:2:2:0, % xy 2.8 % xy. Frekv. fenotypu xy:")
42
Postup genetického mapování
Zkřížit indikátorové linie (xxYY) s homozyg. mutantem (XXyy) - optimálně odlišné ekotypy x = indikátorový marker y = hledaný (mutovaný) gen F1 heterozygot (mnohonásobný) – samoopylení F2-segregace v populaci F2 rostlin (až stovky) spočítat pro každý marker frekvenci fenotypových tříd: XY, Xy, xY, xy snížená frekvence rekombinantních fenotypů (XY , xy) = vazba Následuje mapování v rámci daného raménka: hledání markeru silně kosegregujícího s analyzovanou mutací = lokus v silné genové vazbě (využití markerů se známou pozicí na chromozómu) Alternativně: od samého počátku mapujeme jen s molekulárními markery, jež nemají předem známou pozici křížení homozygotních imbredních linií odlišných ekotypů (typicky markery typu AFLP, RAPD – viz dále)
 s homozyg. mutantem (XXyy) - optimálně odlišné ekotypy. x = indikátorový marker y = hledaný (mutovaný) gen. F1 heterozygot (mnohonásobný) – samoopylení. F2-segregace. v populaci F2 rostlin (až stovky) spočítat pro každý marker frekvenci fenotypových tříd: XY, Xy, xY, xy. snížená frekvence rekombinantních fenotypů (XY , xy) = vazba. Následuje mapování v rámci daného raménka: hledání markeru silně kosegregujícího s analyzovanou mutací. = lokus v silné genové vazbě (využití markerů se známou pozicí na chromozómu) Alternativně: od samého počátku mapujeme jen s molekulárními markery, jež nemají předem známou pozici. křížení homozygotních imbredních linií odlišných ekotypů. (typicky markery typu. AFLP, RAPD – viz dále)")
43
Hledání vhodných molekulárních markerů u Arabidopsis

44
DNA molekulární markery (= proužek na elektroforéze či blotu)
RFLP (Restriction fragment length polymorfism) + Southern RAPD (Random amplified polymorphism detection) AFLP (Amplified fragment length polymorphism) SSR (Simple sequence repeats) SNP (Single nucleotide polymorphism)
 + Southern. RAPD (Random amplified polymorphism detection) AFLP (Amplified fragment length polymorphism) SSR (Simple sequence repeats) SNP (Single nucleotide polymorphism)")
45
Kosegregační analýza s molekulárními markery
nutno křížit odlišné genotypy s vysokou mírou polymorfismu možnost analýzy segregace většího počtu markerů najednou Který z markerů A,B,C,D je ve vazbě se sledovaným lokusem R? r R Fenotyp: fenotyp r fenotyp R

46
Hromadná analýza (bulked segregant analysis)
Je-li vazba silná (o tu nám jde!), je možné analyzovat hromadně. r R fenotyp r fenotyp R
, je možné analyzovat hromadně. r. R. fenotyp r. fenotyp R.")
47
RFLP Restriction fragment length polymorfism

48
RFLP: pro a proti kodominantní markery dobrá reprodukovatelnost
nutno předem charakterizovat všechny markery, t.j. zpravidla je klonovat RFLP polymorfismus není příliš častý drahé a relativně pomalé (hybridizace)
")
49
RAPD Random amplified polymorphism detection - libovolné primery 8-12 nt - nahodilá amplifikace z různých míst genomu

50
RAPD: pro a proti netřeba nic klonovat, stačí dobrá sada primerů
řada markerů současně úspěšný fragment lze snadno klonovat horší reprodukovatelnost mezi pracovišti pouze dominantní markery (z povahy PCR)
")
51
SSR (simple sequence repeats) - detekce polymorfismu v délce mikrosatelitní DNA
problémem je obtížné hledání výchozích markerů (jako u RFLP)
")
52
AFLP Amplified fragment length polymorphism
- restrikce dvěma enzymy - ligace specifických adaptorů - amplifikace s primery nasedajícími na adaptory s přesahem na 3’konci - elektroforéza (fluorescenční znač.) (kombinace určitých koncových trinukleotidů selektivně amplifikuje vždy jen určitou subpopulaci fragmentů) analyzovatelný počet fragmentů!
 (kombinace určitých koncových. trinukleotidů selektivně amplifikuje. vždy jen určitou subpopulaci fragmentů) analyzovatelný počet fragmentů!")
53
AFLP: pro a proti netřeba znát markery, ani předem nic klonovat
řada markerů současně dobrá reprodukovatelnost může být i kodominantní jako RFLP klonování fragmentů problematické (malé množství produktu, fluorescenční či radioaktivní značení)
")
54
Kosegregace s molekulární markery
- následná analýza RFLP, SSR – rovnou známe sekvenci/pozici v genetické mapě RAPD, AFLP - předem neznáme pozici v genomu - pokud kosegregují s hledanou mutací (znakem), pak je můžeme klonovat a sekvenovat (určit pozici)
, pak. je můžeme klonovat a sekvenovat (určit pozici)")
55
Identifikace mutovaného genu kosegregační analýzou
(či třeba genu odolnosti - rezistence) kosegregační analýzou mapování - (ko)segregační analýza nalezení přibližné pozice genu v genetické mapě, na základě genové vazby s genetickými markery 2. nalezení konkrétní sekvence nesoucí mutaci (hledaný gen): - chromosom walking – nalezení a izolace příslušného úseku chromozómu - sekvenace, srovnání s WT sekvencí
 kosegregační analýzou. mapování - (ko)segregační analýza. nalezení přibližné pozice genu v genetické mapě, na základě genové vazby s genetickými markery. 2. nalezení konkrétní sekvence nesoucí mutaci (hledaný gen): - chromosom walking – nalezení a izolace příslušného úseku chromozómu. - sekvenace, srovnání s WT sekvencí.")
56
2) Hledání překryvů na základě hybridizace
Po nalezení optimálně dvou markerů obklopujících mutovaný gen „Chromosome walking“ 1) knihovny velkých fragmentů genomové DNA mutanta (redundantní, náhodné): př. YACs, BACs = yeast (bacterial) arteficial chromosome, ~ 300 (100) kbp cosmidy ( fág, 50 kbp) 2) Hledání překryvů na základě hybridizace koncové sekvence Mutovaný gen X
 knihovny velkých fragmentů genomové DNA mutanta (redundantní, náhodné): př. YACs, BACs. = yeast (bacterial) arteficial chromosome, ~ 300 (100) kbp. cosmidy ( fág, 50 kbp) 2) Hledání překryvů na základě hybridizace. koncové sekvence. Mutovaný gen X.")
57
Marker assisted selection (MAS)
= využití molekulárních markerů ve vazbě s určitým znakem při šlechtění - molekulární marker nahrazuje či doplňuje fenotypový screening - často jednodušší - možnost selekce už ve stádiu semenáčků (ovocné stromy!) - bez vlivu prostředí - někdy (RFLP) možnost rozlišit homo- a heterozygoty podmínka: silná vazba markeru se znakem = přítomnost DNA markeru spolehlivě předpovídá fenotyp
 - bez vlivu prostředí. - někdy (RFLP) možnost rozlišit homo- a heterozygoty. podmínka: silná vazba markeru se znakem = přítomnost DNA markeru spolehlivě předpovídá fenotyp.")
58
Vazba markeru se znakem
Vzdálenost markeru a genu ideálně <5 cM Marker A GenX 5 cM Spolehlivost při selekci S jedním markerem: 1 – rA = ~95% Marker A GenX Marker B 5 cM Se dvěma markery: 1 - 2 rArB = ~99.5% Dva markery zvyšují spolehlivost, ale i pracnost a náklady.

59
Další metody pro identifikaci/izolaci rostlinných genů
Na základě homologie - PCR s degenerovanými primery (či screening knihoven hybridizací) Funkční klonování v kvasinkách: obecný princip: selekce kvasinkových klonů, do nichž byl z knihovny (cDNA) transformován příslušný gen (jen s ním přežijí, dělí se) - hledání homologů na základě funkční komplementace mutací v kvasinkách (u základních pochodů metabolismu, transportu, …) - hledání proteinových interaktorů (genové interakce) kvasinkový 2-hybridní systém
 Funkční klonování v kvasinkách: obecný princip: selekce kvasinkových klonů, do nichž byl z knihovny (cDNA) transformován příslušný gen (jen s ním přežijí, dělí se) - hledání homologů na základě. funkční komplementace mutací v kvasinkách. (u základních pochodů metabolismu, transportu, …) - hledání proteinových interaktorů (genové interakce) kvasinkový 2-hybridní systém.")
60
Kvasinkový dvouhybridní systém - expresi reportérového genu spustí interakce dvou fúzních proteinů = rekonstrukce kompletního transkripčního faktoru transkripce - viz C B DNA vazebná doména TF + protein, jehož interaktory hledáme A Aktivační doména TF (vázající RNA pol.) + fragmenty z knihovny (cDNA) Protein, jehož interaktory hledáme knihovna (cDNA)
")
62
Identifikace mutovaného genu
u T-DNA nebo transpozónem taggovaných linií Fenotyp: Nalezení T-DNA zodpovědné za fenot. projev (je-li jich přítomno více, heterozyg. s reces.m.) (vždy) TAIL PCR degenerovaný primer Transformace Selekce (generace: T1 – přímo po transformaci (T0 – infiltrovaná rostlina), T2 (inbreadní) a další - přítomni i homozygoti s fenotypovým projevem 3) Izolace genomové DNA, Southernova hybridizace 4) (optimálně) výběr linie s jedinou insercí
 (vždy) TAIL PCR. degenerovaný primer. Transformace. Selekce (generace: T1 – přímo po transformaci (T0 – infiltrovaná rostlina), T2 (inbreadní) a další - přítomni i homozygoti s fenotypovým projevem. 3) Izolace genomové DNA, Southernova hybridizace. 4) (optimálně) výběr linie s jedinou insercí.")
63
Přímá a reverzní genetika u Arabidopsis

64
Přímá a reverzní genetika u Arabidopsis

65
Přirozená morfologická variabilita Arabidopsis – ekotypy/accessions

Podobné prezentace


















 RNA>")

