Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Buněčný cyklus Buněčné dělení Abnormality buněčného dělení

2
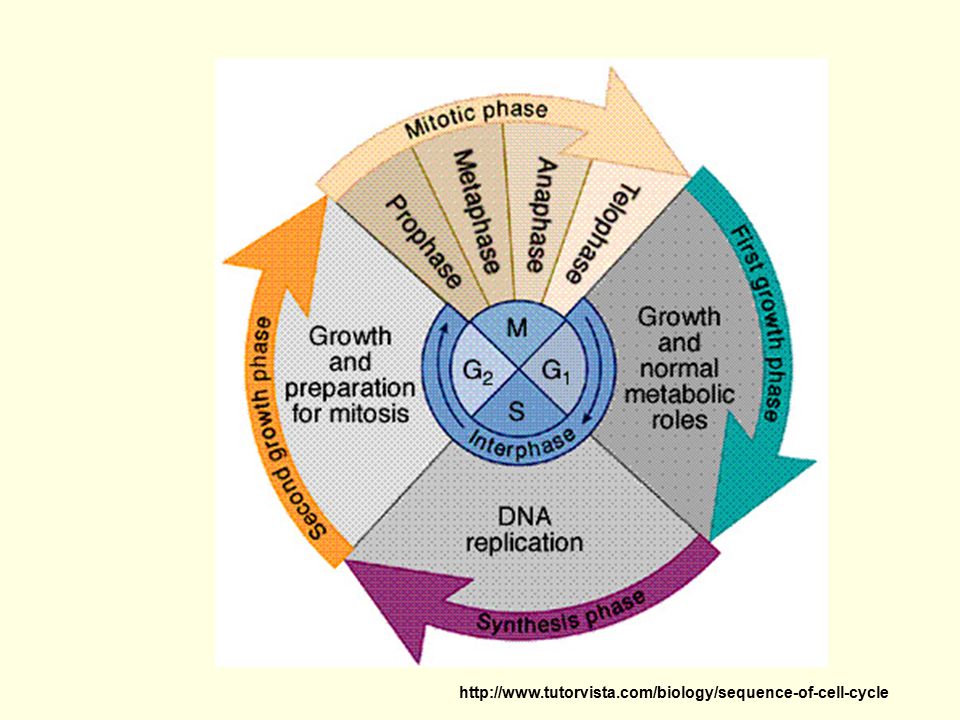
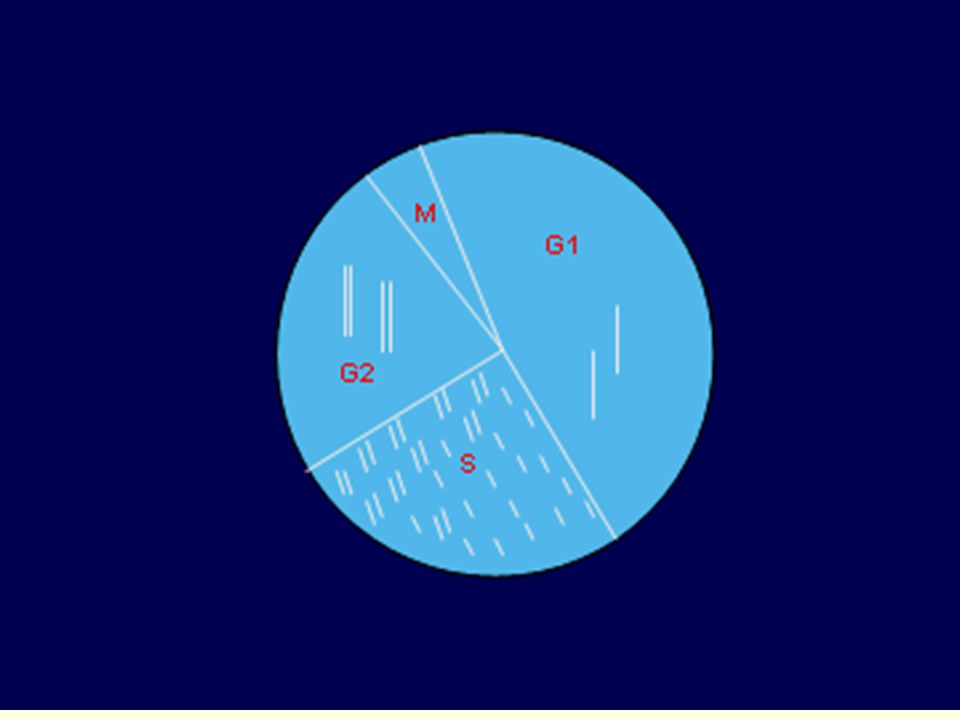
Buněčný cyklus interfáze : G1, S, G2 mitoza
G1 fáze - nejdelší, variabilní část cyklu syntéza RNA, proteinů, doplnění organel (ribozomů, mitochondrií, ER apod.) syntéza nukleotidů, příprava na replikaci kontrolní bod cyklu pro vstup do S fáze
 syntéza nukleotidů, příprava na replikaci kontrolní bod cyklu pro vstup do S fáze.")
3
S fáze - replikace jaderné DNA
mimojaderná DNA (=mitochondriální) se replikuje i mimo S fázi G2 fáze - růst buňky, syntéza proteinů, RNA, tvorba buněčných struktur syntéza rychlejší, transkripce pravděpodobně z obou chromatid kontrolní bod cyklu pro vstup do mitozy Mitoza - spiralizace chromozomů, rozpad jaderné membrány, vytvoření mitotického aparátu- rozdělení chromatid, rozdělení jádra Cytokineze – rozdělení buňky
 se replikuje i mimo S fázi. G2 fáze - růst buňky, syntéza proteinů, RNA, tvorba buněčných struktur. syntéza rychlejší, transkripce pravděpodobně z obou chromatid. kontrolní bod cyklu pro vstup do mitozy. Mitoza - spiralizace chromozomů, rozpad jaderné membrány, vytvoření mitotického aparátu- rozdělení chromatid, rozdělení jádra. Cytokineze – rozdělení buňky.")
6
Dělení somatických buněk = mitoza
= z diploidní mateřské buňky dvě diploidní dceřinné

7
profáze interfáze prometafáze Mitoza 2n 2n metafáze telofáze anafáze

8
Regulace buněčného cyklu
Cyklinový systém: cykliny - cyklická syntéza a odbourávání v průběhu buněčného cyklu cykliny ovlivňují cyklin dependentní kinázy (CDK) = enzymy fosforylující další proteiny trvale přítomny v buňce, aktivace vazbou s cyklinem inaktivace vazbou s inhibitory (p21) komplex cyklin / kináza = fosforylace proteinu = spuštění fáze cyklu
 = enzymy fosforylující další proteiny. trvale přítomny v buňce, aktivace vazbou s cyklinem. inaktivace vazbou s inhibitory (p21) komplex cyklin / kináza = fosforylace proteinu = spuštění fáze cyklu.")
9
Regulační body buněčného cyklu:
G1 - vstup do S – G2 -vstup do M – + další kontrolní body (M - na rozhraní metafáze a anafáze)
")
10
Mechanizmy regulace buněčné proliferace
nadřazeny mechanizmům řízení buněčného cyklu řízení proliferace v: ontogenezi fyziologické obnově buněk reparaci a hojení imunitní odpovědi

11
Exogenní mechanizmy regulace proliferace
- zdroj signálu vně buňky přímé: kontaktem membrán – kontaktní inhibice přímým propojením - „gap junctions“ (kanálky-výměna molekul) zprostředkované : signálními molekulami endokrinní – na velké vzdálenosti – hormony parakrinní – difuze, na krátké vzdálenosti autokrinní – sekretující buňka=cílová buňka synaptická – synapse -neurotransmitery
 zprostředkované : signálními molekulami. endokrinní – na velké vzdálenosti – hormony. parakrinní – difuze, na krátké vzdálenosti. autokrinní – sekretující buňka=cílová buňka. synaptická – synapse -neurotransmitery.")
12
Signální molekuly cytokiny: regulace proliferace, diferenciace –
parakrinní /autokrinní signal. růstové faktory: stimulační signál př. EGF-epidermální růstový faktor IGF – inzulin „like“ růst.faktor PDGF – z trombocytů lymfokiny, monokiny: hemopoetický původ role v imunitních reakcích při zánětu např. IL interleukiny většinou stimulační signál

13
interferony – inhibice proliferace, protivirový účinek
imunomodulátory využití k léčbě nádorů TNF – tumor necrosis faktor regulace buněč.proliferace, genové exprese, zabíjení nádor.buněk hormony

14
Geny regulující buněčné dělení:
Protoonkogeny produkty stimulují buněčné dělení (př. myc, fos, jun aj.) kódují růstové faktory, receptory růstových faktorů, regulační proteiny apod. mutované formy = onkogeny = stálá nebo nadměrná mitotická aktivita
 kódují růstové faktory, receptory růstových faktorů, regulační proteiny apod. mutované formy = onkogeny = stálá nebo nadměrná mitotická aktivita.")
15
Nádorové supresorové geny
produkty inhibují mitotické dělení př. Rb1 gen – mutace u retinoblastomu i jiných tumorů p53 – mutace genu v mnoha nádorech nadřazen dalším genům funkce: zástava cyklu v G1 při poškození DNA a navození reparace nebo apoptózy - programované buněčné smrti

16
Apoptoza = programovaná buněčná smrt
konečné stadium procesu stárnutí buněk odstranění buněk, které již nelze reparovat během embryogeneze-redukce nadbytečných částí Účel: odstranění buněk, které již splnily svou úlohu a mohly by být pro organizmus zhoubné

17
Apoptoza: zánik buňky bez porušení její integrity a integrity organel
rozpad chromatinu, jádra, cytoplazmy – apoptotická tělíska fagocytoza takto změněných buněk bez zánětu nekroza: porušení mebrány buňky, dezintegrace organel, lyze buněk-zánět

18
Meioza

19
Meiotické dělení = dělení pohlavních buněk
2 části: MI, MII

20
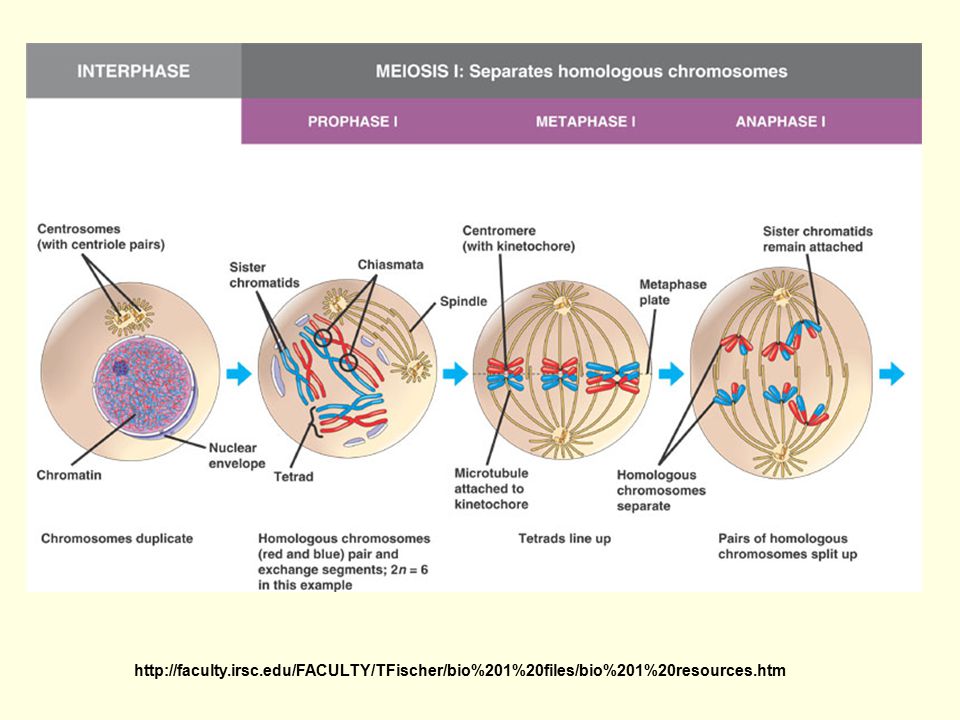
M I = redukční 2n = heterotypické Profáze :
leptotene - počátek spiralizace zygotene - párování (synapse homologů) = bivalenty synaptonemální komplex zajištuje párování homologů párování X a Y pouze konci - sexuální váček
 = bivalenty. synaptonemální komplex zajištuje párování homologů. párování X a Y pouze konci - sexuální váček.")
21
Synaptonemální komplex
Molecular biology of the cell

22
pachytene - patrny sesterské chromatidy = tetrády
crossing over= překřížení a výměna částí nesesterských chromatid homologních chromozomů = rekombinace otcovského a mateřského chrom.materiálu diplotene - separace bivalentů - spojeny v místě crossing overu = chiasmata diakineze - maximální zkrácení chromozomů - terminalizace chiazmat

23
Metafáze: orientace párů chromozomů v ekvatoriální rovině, centromery se neštěpí
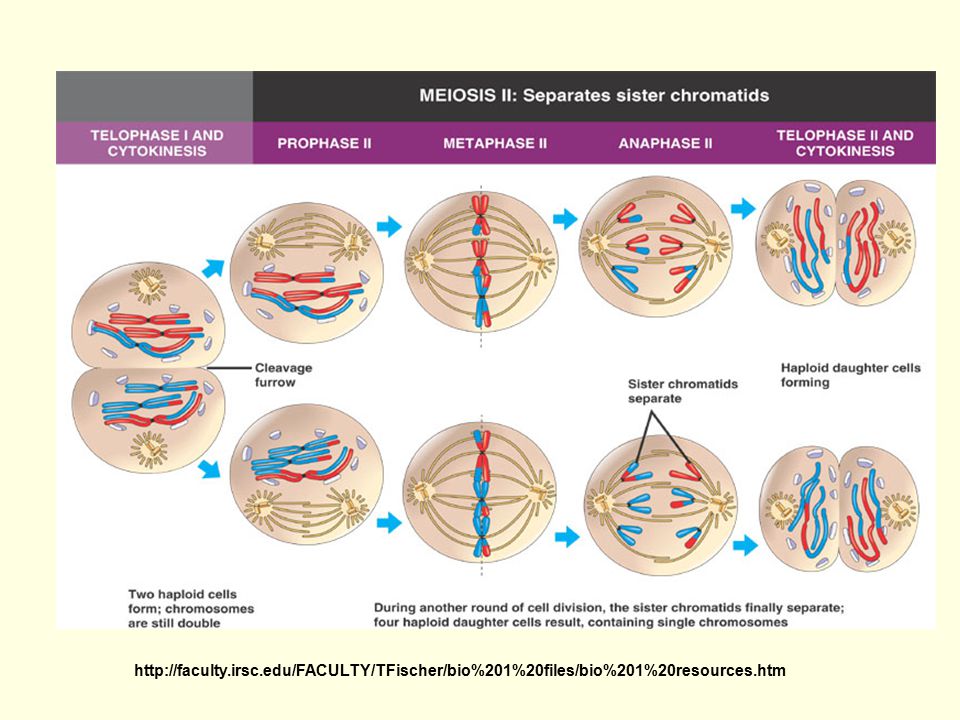
Anafáze: rozchod homologů - náhodný – náhodné kombinace otcovských a mateřských chromozomů Telofáze: haploidní sady na opačných polech Interkineze: bez replikace

24
M II = homeotypické = n ekvační n = mitotické
v metafázi štěpení centromer v anafázi separace chromatid

27
I. Meiotické dělení profáze leptotene zygotene pachytene diplotene
crossing over diakineze anafáze telofáze

28
II.Meiotické dělení anafáze

29
anafáze M I telofáze M I Rozchod chromozomů - náhodný z hlediska rodičovského původu

30
II.Meiotické dělení anafáze telofáze

31
Rozdíly mezi spermiogenezou a oogenezou
Spermiogeneza začíná v době sexuální zralosti z 1 mateřské buňky vzniknou 4 dceřinné Oogeneza začíná již v časném embryonálním období, pak zastavena na konci profáze MI, dále pokračuje v pravidelných intervalech v době sexuální zralosti a plně je dokončeno až po oplození (anafáze a telofáze MII) Z 1 mateřské buňky vznikne 1 vajíčko a 2-3 polová tělíska
 Z 1 mateřské buňky vznikne 1 vajíčko a 2-3 polová tělíska.")
32
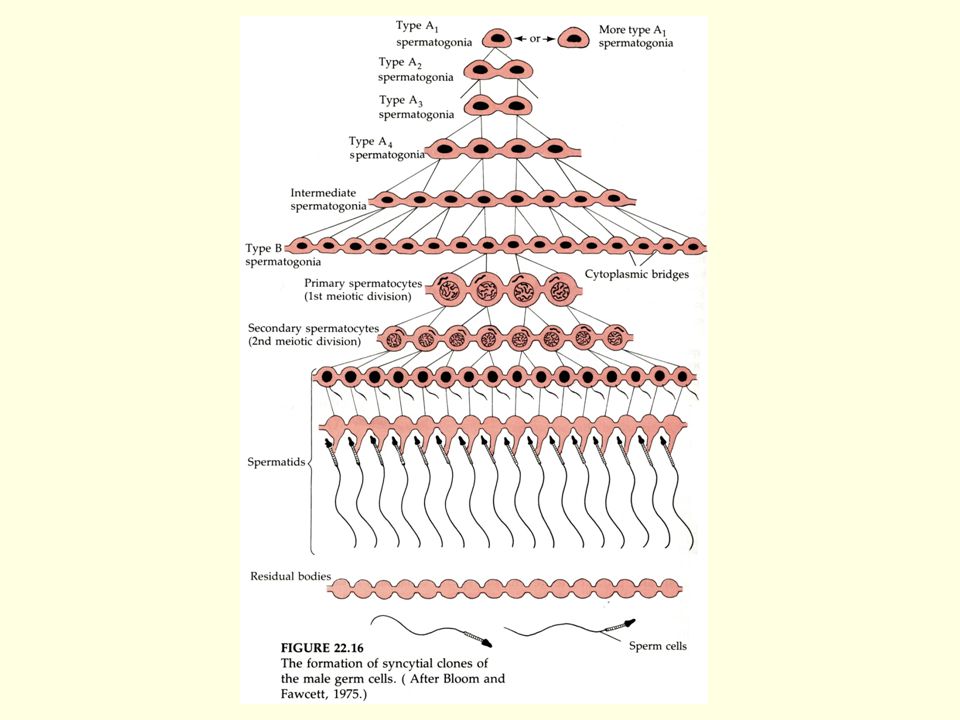
Spermiogeneza – v době sexuální zralosti
spermatogonie Mitotické dělení růst primární spermatocyt M I sekundární spermatocyt meioza M II spermatidy zrání, diferenciace spermie Spermiogeneza – v době sexuální zralosti

34
Oogeneza – začátek v prenatálním období
oogonie Mitotické dělení růst 3.měs.fetál.života dictyotene MI v době porodu primární oocyt M I meioza 1.pol.tělísko Metafáze MII ovulace M II sekundární oocyt 2.pol.tělísko Anafáze,telofáze po oplození Oplození – pronukleus vajíčko dokončí MII- pronukleus zygota Oogeneza – začátek v prenatálním období pokračování v době sexuální dospělosti

35
Spermiogeneza M I M II 2n n n spermatogonie spermatida
primární sekundární spermatocyt spermie

36
Oogeneza a fertilizace
I. meiotické dělení polové tělísko oogonie primární sekundární oocyt n 2n fertilizace a II.meiotické dělení

37
Genetické důsledky meiozy
redukce diploidního počtu chromozomů na haploidní segregace alel v MI , MII náhodný rozchod chromozomů – náhodné kombinace chromozomů v gametách (dle rodičovského původu) zvýšení genetické variability crossing overem (segregující chromozom složen z částí mateřského a otcovského původu)
 zvýšení genetické variability crossing overem (segregující chromozom složen z částí mateřského a otcovského původu)")
38
Chyby v meioze Nondisjunkce Opoždění 1 chromozomu v anafázi
v MI – nerozdělení homologních chromozomů v MII – nerozdělení chromatid Důsledky: pro 1 pár – disomická nebo nullisomická gameta (trisomie-47chrom. + monosomie-45chrom. po oplození) pro celou sadu – diploidní gameta ( triploidie po oplození- 69chrom.) Opoždění 1 chromozomu v anafázi Důsledek: nullisomická gameta – monosomie po oplození (45chrom.)
 pro celou sadu – diploidní gameta. ( triploidie po oplození- 69chrom.) Opoždění 1 chromozomu v anafázi. Důsledek: nullisomická gameta – monosomie po oplození (45chrom.)")
39
Chyby v meioze 46 46 M I 23 24 22 23 M II 23 23 23 23 24 24 22 22 nondisjunkce v M I důsledek: trizomie/monozomie po oplození normální průběh meiozy

40
46 46 M I 23 22 23 23 (X chrom.) M II 24 22 23 23 22 22 23 22 Opoždění chromozomu v anafázi M I nebo M II Důsledek: monozomie po oplození Nondisjunkce v M II Důsledek: trizomie/monozomie po oplození

41
46 46 M I 46 23 23 M II 46 46 23 23 46 Chyby v meiose – nondisjunkce celé sady chromozomů (M I nebo M II) Důsledek: neredukovaná gameta, triploidie po oplození

42
Chyby v mitoze Nondisjunkce ( postzygotická)
Opoždění v anafázi → mozaicismus – 2 nebo více linií buněk s různým karyotypem v jednom organismu ! Endoreduplikace- dělení chromozomů bez dělení buňky → tetraploidie (92 chromozomů)
")
43
46 46 46 46 46 47 45 46 46 46 46 47 47 Nondisjunkce v mitoze – vznik mozaiky - trizomie/normální chrom.vybavení Monozomie letální s vyjímkou gonosomu X

44
46 - X 46 45 46 46 opoždění v anafázi : monozomie a normální buněč.linie v mozaice – letální s vyjímkou gonozomu X

45
Endoreduplikace – dělení chromozomu bez dělení buňky
46 92 tetraploidie

46
Chyby fertilizace Dispermie – oplození vajíčka 2 spermiemi →
triploidie (=69 chromozomů ) = mola částečná Částečná mola -vznik též splynutím normálního haploidního vajíčka s diploidní, neredukovanou spermií (se 46 chromozomy) Chimera – oplození vajíčka a pólového tělíska spermiemi s odlišným gonozomem (XX, XY)→ jedinec je směsí buněk 46,XX a 46,XY
 = mola částečná. Částečná mola -vznik též splynutím normálního haploidního vajíčka s diploidní, neredukovanou spermií (se 46 chromozomy) Chimera – oplození vajíčka a pólového. tělíska spermiemi s odlišným. gonozomem (XX, XY)→ jedinec je směsí buněk 46,XX a 46,XY.")
47
Dispermie- oplození vajíčka 2 spermiemi
Poruchy oplození 23X 23X 23,X 23,X 23Y 46 XX 69 XXY oplození Dispermie- oplození vajíčka 2 spermiemi

48
oplození vajíčka a polového tělíska – vznik chimery
23X 23,X 23,X 23Y 46XX/46,XY oplození vajíčka a polového tělíska – vznik chimery

49
Hydatiformní mola – kompletní
(hypertrofie trofoblastu bez přítomnosti fetální tkáně) karyotyp 46,XX (XY), ale pouze samčí sada chromozomů a) duplikace chromozomů spermie v enukleovaném vajíčku (90%) b) dispermie v enukleovaném vajíčku- (10%)
 karyotyp 46,XX (XY), ale pouze samčí sada chromozomů. a) duplikace chromozomů spermie v enukleovaném vajíčku (90%) b) dispermie v enukleovaném vajíčku- (10%)")
50
hydatiformní mola (úplná) = dispermie nebo duplikace spermie v enukleovaném vajíčku
Case contributed by Nazila Azordegan MD of Tehran, IRAN

51
Mechanizmy vzniku úplné moly
23X 23X 23Y Duplikace chromozomů 46 XX 46 XY Mechanizmy vzniku úplné moly

52
Partenogeneze Ovariální teratom –původ z oocytů po prvním meiotickém dělení- duplikací samičí sady chromozomů (dělení neoplozeného vajíčka) karyotyp buněk benigního tumoru je 46,XX přítomnost různých druhů tkání ( kůže, chrupavka, štítná žláza atd.)
")
53
Partenogeneze u člověka
ovariální teratom = duplikace chromozomů a dělení neoplozeného vajíčka

54
ASISTOVANÁ REPRODUKCE
Úbytek obyvatel na severní polokouli ČR od roku 1994 menší porodnost než úmrtnost - odkládání porodu prvního dítěte - snižování plodnosti mužů ( snížení počtu spermií o 50% za 50 let ) Celosvětové zvyšování počtu mnohočetných těhotenství Ve vyspělých zemích cca 8000 AR / obyvatel ( asi 1,5 % dětí je “ze zkumavky“ ) Počet neplodných párů stoupá
 Celosvětové zvyšování počtu mnohočetných těhotenství. Ve vyspělých zemích cca 8000 AR / obyvatel. ( asi 1,5 % dětí je ze zkumavky ) Počet neplodných párů stoupá.")
55
Nejčastější důvody neplodnosti:
muži – azoospermie – nepřítomnost spermií oligospermie – malý počet spermií azoospermie % aberace gonozomů Yq mikrodelece genu DAZ („deleted in azoospermia“) oligospermie - častěji strukturní aberace autozomů (4%-balancovaná strukturní aberace) mutace CFTR – cystická fibroza ženy – gynekologické: tubární sterilita endometriosa imunologické chrom.abnormality ( balancovaná strukturní aberace)
 oligospermie - častěji strukturní aberace autozomů. (4%-balancovaná strukturní aberace) mutace CFTR – cystická fibroza. ženy – gynekologické: tubární sterilita. endometriosa. imunologické. chrom.abnormality ( balancovaná strukturní aberace)")
56
CFTR gen - cystická fibroza –
u některých mutací je neplodnost muže jediným projevem ( vyšetření heterozygotnosti partnerů ) (možnost chirurgického odběru spermií) Další důvody k AR : genetická choroba ( např. Turnerův syndrom ) stav po protinádorové terapii ( kryokonzervace gamet před začátkem léčby )
 (možnost chirurgického odběru spermií) Další důvody k AR : genetická choroba ( např. Turnerův syndrom ) stav po protinádorové terapii. ( kryokonzervace gamet před začátkem léčby )")
57
Indikace k AR - pouze zdravotní
Matka je žena, která plod donosí a porodí. Nemožnost prenatální adopce. Od 18 let má dítě právo na informace o biologickém rodiči. Uchování informací o dárcích let. Etický problém likvidace zmrazených embryí ( pojem preembryo tj. neimplantované embryo ).
.")
58
Dárcovství gamet Anonymita dárce i příjemce.
Nepřípustnost příbuzeneckého vztahu. Věk od 18 do 40 u mužů a do 35 let u žen. Informace o dárci ( IQ, rasa aj. ) Ochrana proti AIDS – opakované vyšetření po 180 dnech a pak teprve použití zmrazených gamet. Vyšetření karyotypu dárců. V některých zemích je podmínkou dárcovství být rodičem dvou zdravých dětí. Souhlas manželů aj..
 Ochrana proti AIDS – opakované vyšetření po. 180 dnech a pak teprve použití zmrazených. gamet. Vyšetření karyotypu dárců. V některých zemích. je podmínkou dárcovství být rodičem dvou. zdravých dětí. Souhlas manželů aj..")
59
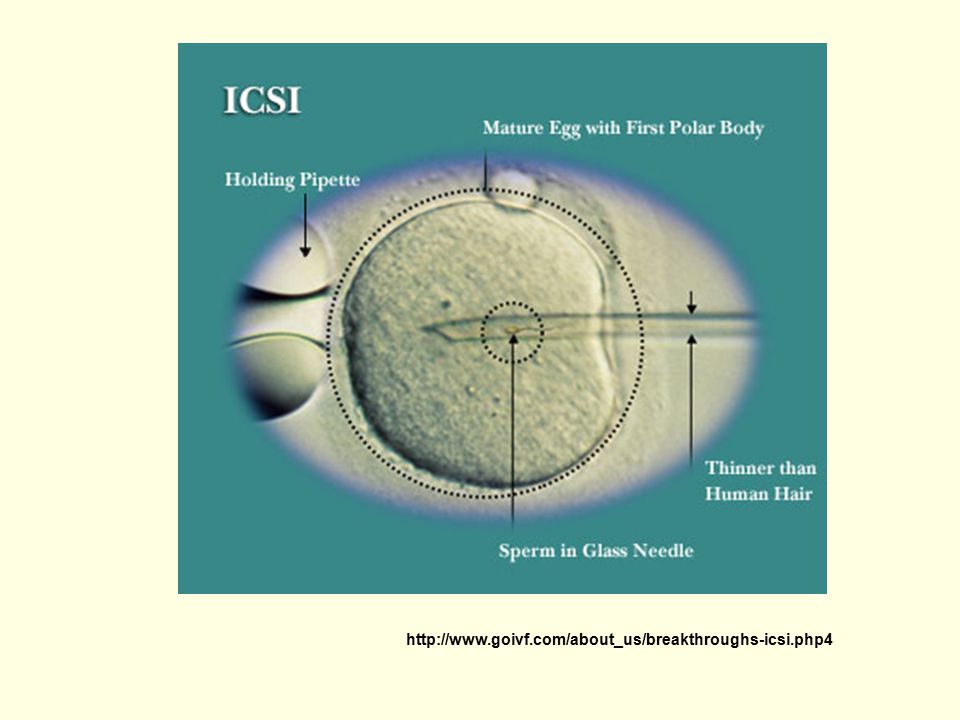
Metody asistované reprodukce
Spermie: dárce nebo biopsie testes u azoospermie IVF (in vitro fertilizace) intracytoplazmatická injekce spermie (ICSI) disekce zona pellucida intracytoplazmatická injekce spermie Nejčastější indikace ICSI : sterilita muže imunologická sterilita (antizonální protilátky)
 intracytoplazmatická injekce spermie (ICSI) disekce zona pellucida. intracytoplazmatická injekce spermie. Nejčastější indikace ICSI : sterilita muže. imunologická sterilita (antizonální protilátky)")
61
Preimplantační diagnoza:
polové tělísko nebo buňky embrya po biopsii (z blastocysty možno odebrat více buněk) Metody vyšetření : PCR = polymerázová řetězová reakce FISH = fluorescenční “in situ“ hybridizace - zjištění numerických chrom. aberací
 Metody vyšetření : PCR = polymerázová řetězová reakce. FISH = fluorescenční in situ hybridizace. - zjištění numerických chrom. aberací.")
62
Po oplození – pólová tělíska a v zygotě 2 prvojádra - pronuclei
Rýhující se zygota

63
Jestli se cítíš malý(á), deprimovaný(á), neužitečný(á), vzpomeň si,
že díky perfektnímu vajíčku a nejrychlejší spermii a jimi nesenými genetickými informacemi jsi absolutně jedinečná a neopakovatelná jednotka tohoto vesmíru!

Podobné prezentace








>")




>")

 = M.>")


>")

