Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
HLA systém (MHC glykoproteiny)
")
2
MHC glykoproteiny I. třídy (major histocompatibility complex)
Funkcí MHC gpI je prezentace peptidových fragmentů, které jsou produkovány buňkou (včetně virových, pokud jsou přítomny), na buněčném povrchu tak, aby byly rozpoznávány T lymfocyty (cytotoxickými CD8) Přítomny na všech jaderných buňkách organismu 3 izotypy klasických lidských MHC gp. ( HLA - A, -B, -C ) 3 izotypy neklasických MHC gp. ( HLA – E, -F, -G; molekuly CD1)
, na buněčném povrchu tak, aby byly rozpoznávány T lymfocyty (cytotoxickými CD8) Přítomny na všech jaderných buňkách organismu. 3 izotypy klasických lidských MHC gp. ( HLA - A, -B, -C ) 3 izotypy neklasických MHC gp. ( HLA – E, -F, -G; molekuly CD1)")
3
Prezentace peptidového fragmentu pomocí MHC gp I
Prezentace peptidového fragmentu pomocí MHC gp I. třídy cytotoxickému T lymfocytu

4
Struktura MHC gp I MHC gp. I. třídy se skládají z transmembránového řetězce a a nekovalentně asociovaného b2mikroglobulinu Řetězec a má 3 domény, 2 N-terminální (a1, a2 – vazebné místo pro peptidy) a 1 C-terminální doménu (a3 – zakotvena v cytoplazmatické membráně) Vazba peptidu je nezbytná pro stabilní konformaci MHC gp a tím zajišťuje jeho dlouhodobou prezentaci na buněčném povrchu
 a 1 C-terminální doménu (a3 – zakotvena v cytoplazmatické membráně) Vazba peptidu je nezbytná pro stabilní konformaci MHC gp a tím zajišťuje jeho dlouhodobou prezentaci na buněčném povrchu.")
5
Vazba peptidů na MHC gp I
MHC gp I váží peptidy o délce 8 až 10 AK Určitá molekula MHC gp váže peptidy sdílející společné strukturní rysy - vazebný motiv (rozhodující jsou AK poblíž konců peptidu) K vazbě endogenních peptidů dochází v endoplazmatickém retikulu během biosyntézy MHC gp. Po vytvoření řetězce a a b2mikrotubulinu dochází v ER k poskládání do správné konformace a k vzájemné asociaci a k asociaci vhodného peptidu, tento komplex je dále zpracován v Golgiho aparátu a pak je prezentován na buněčném povrchu Navázané peptidy pocházejí z proteinů degradovaných proteazómem, který štěpí cytoplasmatické proteiny určené k likvidaci (označené ubiquitinem), peptidové fragmenty jsou transportovány do ER pomocí specifických membránových pump
 K vazbě endogenních peptidů dochází v endoplazmatickém retikulu během biosyntézy MHC gp. Po vytvoření řetězce a a b2mikrotubulinu dochází v ER k poskládání do správné konformace a k vzájemné asociaci a k asociaci vhodného peptidu, tento komplex je dále zpracován v Golgiho aparátu a pak je prezentován na buněčném povrchu. Navázané peptidy pocházejí z proteinů degradovaných proteazómem, který štěpí cytoplasmatické proteiny určené k likvidaci (označené ubiquitinem), peptidové fragmenty jsou transportovány do ER pomocí specifických membránových pump.")
6
Vazba peptidů na MHC gp I

7
Vazba peptidů na MHC gp I

8
Neklasické MHC gp. I HLA – E, -F, -G; molekuly CD1 Strukturně podobné klasickým MHC gp Jsou méně polymorfní Vyskytují se jen na některých buňkách Specializují se na vazbu zvláštních ligandů

9
HLA-E a HLA-G - vyskytují se na buňkách trofoblastu
Komplexy HLA-E a HLA-G s peptidy jsou rozpoznávány inhibičními receptory NK buněk a přispívají k toleranci plodu v děloze

10
MHC glykoproteiny II. třídy
Funkcí MHC gp II je prezentace peptidových fragmentů z proteinů pohlcených buňkou tak, aby byly rozpoznatelné T lymfocyty (pomocnými CD4) Vyskytují se na APC ( dendritické buňky, monocyty, makrofágy, B lymfocyty) 3 izotypy MHC gp II ( DR, DQ, DP )
 Vyskytují se na APC ( dendritické buňky, monocyty, makrofágy, B lymfocyty) 3 izotypy MHC gp II ( DR, DQ, DP )")
11
Struktura MHC gp II MHC gp. II se skládají ze 2 nekovalentně asociovaných transmembránových podjednotek a a b Vazebné místo pro peptid je tvořeno N-terminálními doménami a1 a b1 Vazba peptidu je nezbytná pro stabilní konformaci MHC gp a tím zajišťuje jeho dlouhodobou prezentaci na buněčném povrchu

12
Vazba peptidů na MHC gp II
MHC gp II váží peptidy o délce 15 až 35 AK (ale i delší - vazebné místo pro peptid je na obou koncích otevřené) Určitá molekula MHC gp váže peptidy sdílející společné strukturní rysy - vazebný motiv Po vytvoření řetězce a a b v ER dochází k poskládání do správné konformace a k vzájemné asociaci a k připojení dalšího transmembránového řetězce, tzv. invariantního řetězce, který blokuje vazebné místo pro peptid, tento komplex je dále zpracován v Golgiho aparátu; sekreční váčky oddělené od GA fúzují s endozómy, poté se rozštěpí invariantní řetězce a do vazebného místa MHC gp se naváží peptidové fragmenty endocytovaných proteinů a poté je komplex prezentován na buněčném povrchu
 Určitá molekula MHC gp váže peptidy sdílející společné strukturní rysy - vazebný motiv. Po vytvoření řetězce a a b v ER dochází k poskládání do správné konformace a k vzájemné asociaci a k připojení dalšího transmembránového řetězce, tzv. invariantního řetězce, který blokuje vazebné místo pro peptid, tento komplex je dále zpracován v Golgiho aparátu; sekreční váčky oddělené od GA fúzují s endozómy, poté se rozštěpí invariantní řetězce a do vazebného místa MHC gp se naváží peptidové fragmenty endocytovaných proteinů a poté je komplex prezentován na buněčném povrchu.")
13
Vazba peptidů na MHC gp II

14
Antigenní prezentace

16
Prezentace antigenu T lymfocytům
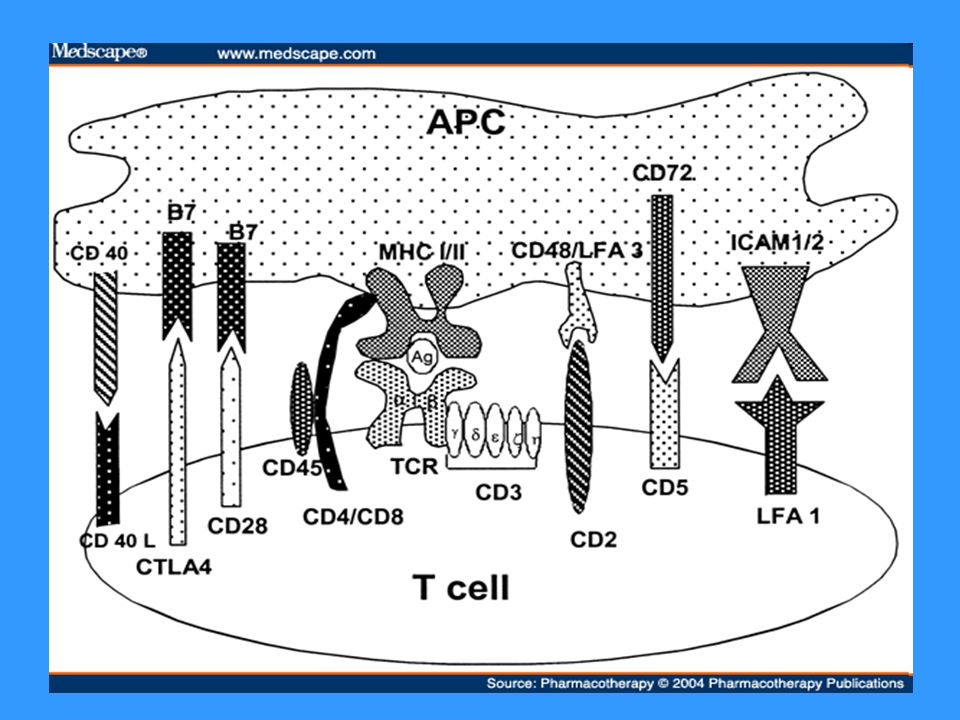
Signál TCR – MHC gp I(II)+Ag peptid (APC) Signál kostimulační CD 28 (Tlymfocyt) – CD 80, CD 86 (APC)
+Ag peptid (APC) Signál kostimulační CD 28 (Tlymfocyt) – CD 80, CD 86 (APC)")
17
Polymorfismus MHC glykoproteinů
HLA komplex se nachází na chromozómu 6 U MHC gp je vysoký polymorfismus, tzn. existují až stovky různých alelických forem jednotlivých izotypů (kromě neklasických MHC gp. I a kromě DR řetězce a) Kodominantní dědičnost alelických forem (jedinec má na povrchu buněk 3 izotypy HLA molekul (HLA-A, -B, -C) většinou ve 2 různých alelických formách) Polymorfismus má ochranný význam na úrovni jedince i na úrovni populace Polymorfismus MHC gp způsobuje komplikace při transplantacích
 Kodominantní dědičnost alelických forem (jedinec má na povrchu buněk 3 izotypy HLA molekul (HLA-A, -B, -C) většinou ve 2 různých alelických formách) Polymorfismus má ochranný význam na úrovni jedince i na úrovni populace. Polymorfismus MHC gp způsobuje komplikace při transplantacích.")
18
HLA typizace = určení HLA antigenů na povrchu lymfocytů Provádí se při předtransplantačním vyšetření a při určení paternity 1) Sérologická typizace mikrolymfocytotoxický test allospecifická séra ( získaná od vícenásobných rodiček do 6 týdnů po porodu, získaná vakcinací dobrovolníků, nebo komerčně připravené sety typizačních sér (monoklonální protilátky)) princip - inkubace lymfocytů s typizačními séry za přítomnosti králičího komplementu, poté je přidáno vitální barvivo, které obarví mrtvé buňky buňky nesoucí určité HLA jsou usmrceny cytotoxickými Ab proti tomuto Ag, procento mrtvých buněk je mírou toxicity séra (síly a titru antileukocytárních protilátek) za pozitivní reakci se považuje více než 10% mrtvých bb. (sérologickou typizaci lze provádět i pomocí průtokové cytometrie)
 Sérologická typizace. mikrolymfocytotoxický test. allospecifická séra ( získaná od vícenásobných rodiček do 6 týdnů po porodu, získaná vakcinací dobrovolníků, nebo komerčně připravené sety typizačních sér (monoklonální protilátky)) princip - inkubace lymfocytů s typizačními séry za přítomnosti králičího komplementu, poté je přidáno vitální barvivo, které obarví mrtvé buňky - buňky nesoucí určité HLA jsou usmrceny cytotoxickými Ab proti tomuto Ag, procento mrtvých buněk je mírou toxicity séra (síly a titru antileukocytárních protilátek) za pozitivní reakci se považuje více než 10% mrtvých bb. (sérologickou typizaci lze provádět i pomocí průtokové cytometrie)")
19
2) Molekulárně genetické metody
Pro typizaci se používají hypervariabilní úseky ve II. exonu genů kódujících HLA II. třídy, pro určení HLA I. třídy se používá polymorfismus v II. a III. exonu kódujících genů 2a) PCR-SSP = polymerázová řetězová reakce se sekvenčními specifickými primery extrahovaná DNA slouží jako substrát v sadě PCR reakcí každá PCR reakce obsahuje primerový pár specifický pro určitou alelu (resp. skupinu alel) pozitivní a negativní reakce se hodnotí elektroforézou každá kombinace alel má svůj specifický elektroforetický obraz
 PCR-SSP. = polymerázová řetězová reakce se sekvenčními specifickými primery. extrahovaná DNA slouží jako substrát v sadě PCR reakcí. každá PCR reakce obsahuje primerový pár specifický pro určitou alelu (resp. skupinu alel) pozitivní a negativní reakce se hodnotí elektroforézou. každá kombinace alel má svůj specifický elektroforetický obraz.")
20
2b) PCR-SSO = PCR reakce se sekvenčně specifickými oligonukleotidy namnoží se hypervariabilní úseky genů kódujících HLA hybridizace s enzymaticky nebo radioaktivně značenými DNA sondami specifickými pro jednotlivé alely 2c) PCR- SBT = sequencing based typing; sekvenování nejpřesnější metodika HLA typizace získáme přesnou sekvenci nukleotidů, kterou porovnáme s databází známých sekvencí HLA alel
 PCR- SBT. = sequencing based typing; sekvenování. nejpřesnější metodika HLA typizace. získáme přesnou sekvenci nukleotidů, kterou porovnáme s databází známých sekvencí HLA alel.")
21
Imunoglobuliny

22
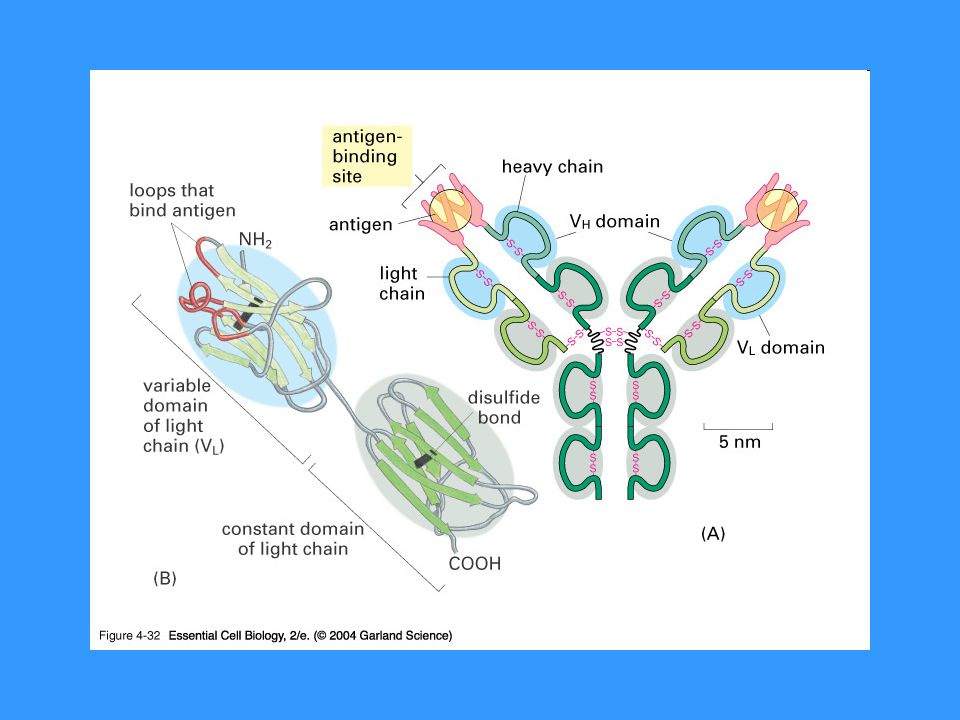
Struktura imunoglobulinů
2 těžké (H) řetězce kovalentně spojeny cystinovými můstky, ke každému H řetězci je cystinovým můstkem připojen lehký (L) řetězec H řetězec se skládá ze 4 až 5 domén (1 variabilní, 3-4 konstantní) L řetězec se skládá ze 2 imunoglobulínových domén (1 variabilní, 1 konstantní) Typy L řetězců - k, l Typy H řetězců – m, d, g (g1-4), a (a1,a2), e
 řetězce kovalentně spojeny cystinovými můstky, ke každému H řetězci je cystinovým můstkem připojen lehký (L) řetězec. H řetězec se skládá ze 4 až 5 domén (1 variabilní, 3-4 konstantní) L řetězec se skládá ze 2 imunoglobulínových domén (1 variabilní, 1 konstantní) Typy L řetězců - k, l. Typy H řetězců – m, d, g (g1-4), a (a1,a2), e.")
24
Variabilní domény L a H řetězce tvoří vazebné místo pro Ag
Pantová oblast = místo, kde jsou těžké řetězce spojeny cystinovými můstky Fc části imunoglobulínů jsou silně glykosilovány, váží se na Fc receptory J řetězec Sekreční komponenta

25
Funkce imunoglobulinů
Neutralizace Ag Aglutinace Ag Aktivace komplementu Opsonizace Aktivace mastocytů prostřednictvím IgE ADCC

26
Funkce imunoglobulinů

27
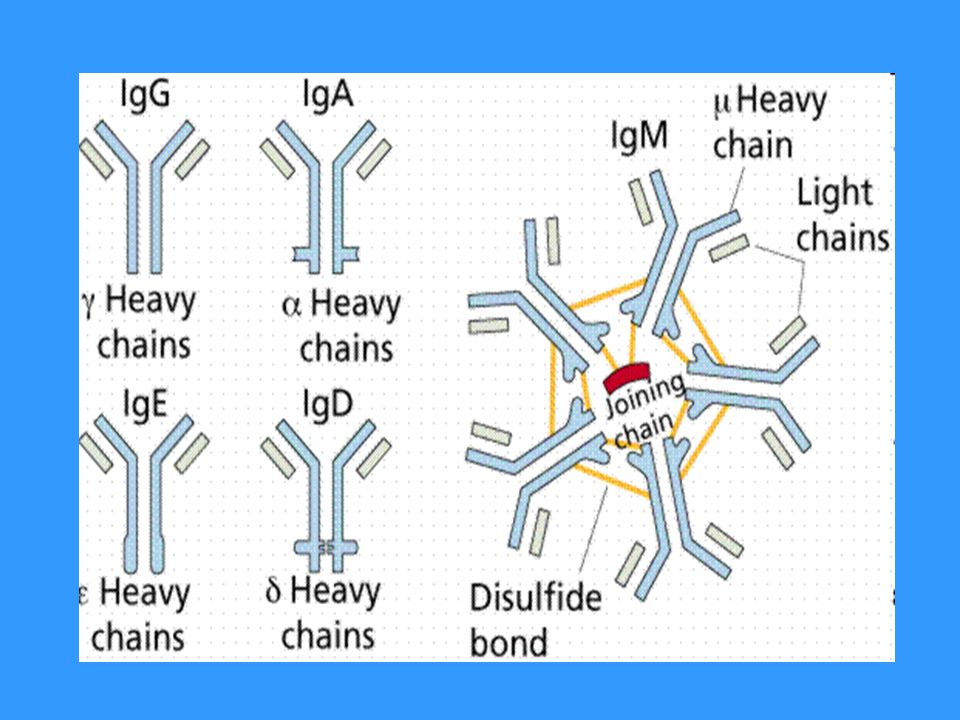
Třídy imunoglobulínů a jejich funkce
Rozlišujeme podle konstantní části H řetězce IgM, IgD, IgG ( IgG1 – IgG4), IgA (IgA1, IgA2), IgE IgM - jako monomer tvoří BCR sekretovaný ve formě pentamerů (10 vazebných míst) první izotyp, který se tvoří po setkání s Ag neutralizace Ag, aktivuje komplement, neváže se na Fc receptory fagocytů (koncentrace 0,9 – 2,5 g/l; biol. poločas 6 dnů) IgD - jako monomer tvoří BCR v séru se nachází ve velmi malé koncentraci - (koncentrace 0,1 g/l; biol. poločas 3 dny)
, IgA (IgA1, IgA2), IgE. IgM - jako monomer tvoří BCR - sekretovaný ve formě pentamerů (10 vazebných míst) - první izotyp, který se tvoří po setkání s Ag - neutralizace Ag, aktivuje komplement, neváže se na Fc receptory fagocytů - (koncentrace 0,9 – 2,5 g/l; biol. poločas 6 dnů) IgD - jako monomer tvoří BCR - v séru se nachází ve velmi malé koncentraci. - (koncentrace 0,1 g/l; biol. poločas 3 dny)")
28
IgG - jednotlivé izotypy IgG1-IgG4 se liší schopností aktivovat komplement a vázat se na Fc receptory fagocytů (opsonizace) - funkce: neutralizace, opsonizace, aktivace komplementu - přestupuje placentou - tvořen při sekundární imunitní odpovědi (koncentrace 8 – 18 g/l; biol. poločas 21 dnů)
")
29
IgA - slizniční IgA - ochrana sliznic, neutralizace, opsonizace, neaktivuje komplement dimér se sekreční komponentou sliny, slzy, mateřské mléko sérový IgA – monomer, dimer či trimer (koncentrace 0,9 – 3,5 g/l; biol. poločas 6 dnů) IgE - uplatňuje se při obraně proti mnohobuněčným parazitům je hlavní příčinou alergických reakcí - (koncentrace 3x10-4 g/l; biol. poločas 2 dny)
")
31
Genetický základ tvorby imunoglobulinů

32
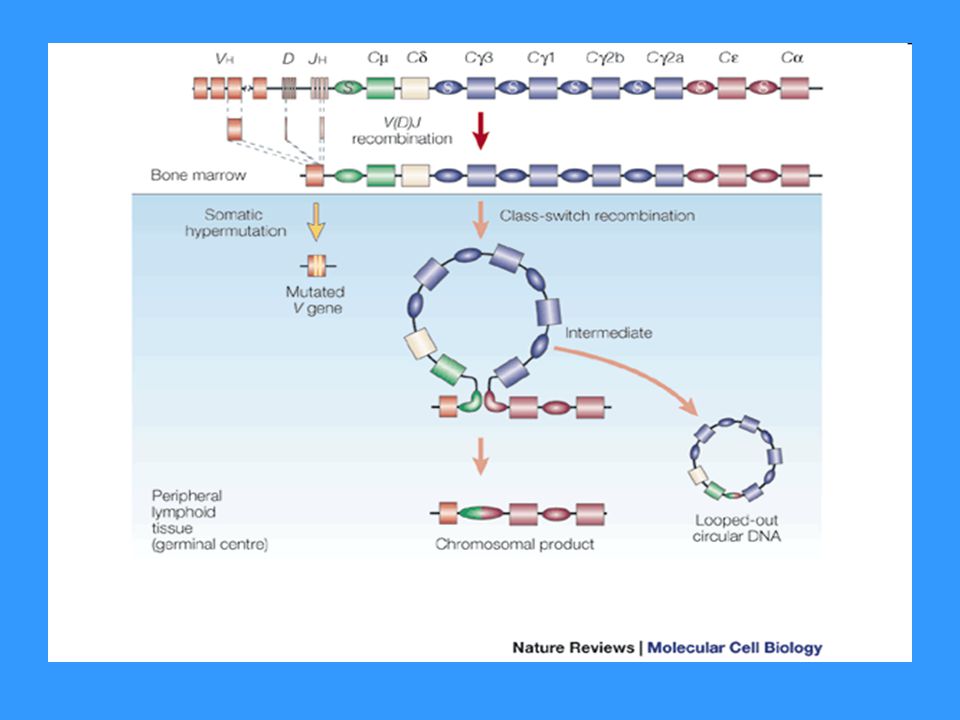
Genové segmenty pro H řetězce - chromozóm 14
V (variabilita) - několik set D (diversita) - asi 50 J (joining) – 9 C konstantní části H řetězce Genové segmenty pro L řetězce - k chromozóm 2 - l chromozóm 22 V (variabilita) J (joining) C konstantní části L řetězce Na koncích V, D, J úseků jsou signální sekvence, které jsou rozpoznávány enzymy rekombinázami, které provádějí přeskupování těchto genů Po stranách C segmentů jsou tzv. switch sekvence (přesmykové), které jsou rozpoznávány enzymy rekombinázami, které provádějí izotypový přesmyk
 - několik set. D (diversita) - asi 50. J (joining) – 9. C konstantní části H řetězce. Genové segmenty pro L řetězce - k chromozóm 2. - l chromozóm 22. V (variabilita) J (joining) C konstantní části L řetězce. Na koncích V, D, J úseků jsou signální sekvence, které jsou rozpoznávány enzymy rekombinázami, které provádějí přeskupování těchto genů. Po stranách C segmentů jsou tzv. switch sekvence (přesmykové), které jsou rozpoznávány enzymy rekombinázami, které provádějí izotypový přesmyk.")
33
Přeskupování genů kódujících H řetězce
D-J přeskupení - vyštěpení úseku IgH mezi některým D a J segmentem (probíhá na obou chromozómech) V-D přeskupení – vyštěpení úseku mezi některým V segmentem a DJ; pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze ( to platí i pro L řetězce) Přepis přeskupeného IgH genu do mRNA, sestřih primárního transkriptu. Jako první se tvoří H řetězce m. Není-li přeskupování úspěšné, B lymfocyt hyne.
 V-D přeskupení – vyštěpení úseku mezi některým V segmentem a DJ; pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze ( to platí i pro L řetězce) Přepis přeskupeného IgH genu do mRNA, sestřih primárního transkriptu. Jako první se tvoří H řetězce m. Není-li přeskupování úspěšné, B lymfocyt hyne.")
34
Přeskupování genů kódujících L řetězce
Nejprve se přeskupují geny kódující L řetězec k, dochází k vyštěpování úseků mezi některým V a J segmentem (souběžně na obou chromozómech), pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze. Jestliže není přeskupování k genů úspěšné, zahájí se přeskupování genů l. Ne všechny H a L řetězce mohou spolu tvořit stabilní dimery. Není-li přeskupování úspěšné, B lymfocyt hyne.
, pokud je přeskupení na některém z chromozómů úspěšné, zastaví se přeskupování na druhém chromozómu – tzv. alelická exkluze. Jestliže není přeskupování k genů úspěšné, zahájí se přeskupování genů l. Ne všechny H a L řetězce mohou spolu tvořit stabilní dimery. Není-li přeskupování úspěšné, B lymfocyt hyne.")
35
Izotypový přesmyk (class switch)
Dochází k němu během terminální diferenciace B lymfocytu po aktivaci Ag na povrchu FDC (nezbytný kostimulační signál přes CD40) Enzymy rekombinázy rozpoznávají switch sekvence lokalizované po stranách C segmentů (tato sekvence není mezi Cm a Cd segmenty – B buňka může před izotypovým přesmykem produkovat IgM i IgD zároveň) a vyštěpují úseky genů Po eliminaci části C oblasti je do mRNA přepsán ten segment, který je nejblíže VDJ segmentu a po sestřihu a translaci vzniká příslušný izotyp H řetězce
 Enzymy rekombinázy rozpoznávají switch sekvence lokalizované po stranách C segmentů (tato sekvence není mezi Cm a Cd segmenty – B buňka může před izotypovým přesmykem produkovat IgM i IgD zároveň) a vyštěpují úseky genů. Po eliminaci části C oblasti je do mRNA přepsán ten segment, který je nejblíže VDJ segmentu a po sestřihu a translaci vzniká příslušný izotyp H řetězce.")
37
Izotypový přesmyk (class switch) - pokračování
Cytokiny regulují k jakému izotypovému přesmyku dojde: IL-4 stimuluje přesmyk na IgE a IgG1, IgG4 TGFb stimuluje přesmyk na IgG2 a IgA

38
Antiidiotypové protilátky
IDIOTOP = každá determinanta variabilní části protilátky, může být součástí paratopu, či může ležet mimo paratop IDIOTYP = soubor idiotopů; souhrn identických vazebných struktur pro Ag na protilátkách stejné specifity Idiotypické struktury protilátek 1. generace jsou rozpoznávány některými B lymfocyty jako antigeny a mohou se proti nim tvořit tzv. antiidiotypové protilátky (protilátky 2. generace; některá vazebná místa mohou připomínat Ag, který vyvolal tvorbu protilátek 1. generace) Proti protilátkám 2. generace se tvoří protilátky 3. generace (anti-antiidiotypové protilátky) Tato idiotypová síť může hrát roli v regulaci protilátkové odpovědi
 Proti protilátkám 2. generace se tvoří protilátky 3. generace (anti-antiidiotypové protilátky) Tato idiotypová síť může hrát roli v regulaci protilátkové odpovědi.")
39
T lymfocyty J. Ochotná

40
T lymfocyty T lymfocyty patří mezi buněčnou složku antigenně specifických mechanismů, při vývoji opouštějí kostní dřeň a migrují do thymu, kde dozrávají Existuje několik různých subpopulací T lymfocytů Podílejí se na regulaci imunitních dějů, při likvidaci virem infikovaných buněk či nádorových buněk Rozpoznávají antigen zpracovaný a prezentovaný APC; prostřednictvím TCR rozpoznávají komplex MHC gp-antigenní peptid T lymfocyty jsou po aktivaci stimulovány k pomnožení a diferenciaci v efektorové buňky, část se diferencuje v paměťové buňky

41
Vývoj T lymfocytů T lymfocyty vznikají v kostní dřeni a pak migrují do thymu, kde dozrávají (abT lymfocyty), konečná diferenciace probíhá až po aktivaci antigenem zpracovaným a prezentovaným APC gdT lymfocyty se mohou vyvíjet i mimo thymus (jde o menšinovou populaci) Pluripotentní hematopoetická kmenová buňka Pro-thymocyt – dostává se z kostní dřeně do thymu, kde se začínají přeskupovat geny pro TCRb, exprimuje na svém povrchu tzv. pre-TCR (složený z b řetězce, pre-TCRa a CD3 komplexu), po té se začínají přeskupovat geny pro TCRa Kortikální thymocyt – exprimuje na svém povrchu TCR (složený z řetězců ab a CD3) a koreceptory CD4 a CD8; v této fázi dochází k selekci autoreaktivních buněk a buněk s nefunkčním TCR Medulární thymocyt (zralá T buňka) – zachovávají si expresi CD4 či CD8, usidlují se v sekundárních lymfoidních orgánech
, konečná diferenciace probíhá až po aktivaci antigenem zpracovaným a prezentovaným APC. gdT lymfocyty se mohou vyvíjet i mimo thymus (jde o menšinovou populaci) Pluripotentní hematopoetická kmenová buňka. Pro-thymocyt – dostává se z kostní dřeně do thymu, kde se začínají přeskupovat geny pro TCRb, exprimuje na svém povrchu tzv. pre-TCR (složený z b řetězce, pre-TCRa a CD3 komplexu), po té se začínají přeskupovat geny pro TCRa. Kortikální thymocyt – exprimuje na svém povrchu TCR (složený z řetězců ab a CD3) a koreceptory CD4 a CD8; v této fázi dochází k selekci autoreaktivních buněk a buněk s nefunkčním TCR. Medulární thymocyt (zralá T buňka) – zachovávají si expresi CD4 či CD8, usidlují se v sekundárních lymfoidních orgánech.")
42
Selekce T lymfocytů Negativní selekce – eliminace autoreaktivních buněk Pokud thymocyt váže svým TCR dostatečně silně komplex MHCgp s normálními peptidy (z autoantigenů) se kterými se setká na povrchu thymových buněk dostane signály vedoucí k apoptotické smrti buňky PAE buňky (peripherial antigen expressing cells) Pozitivní selekce – eliminace buněk s nefunkčním TCR Pozitivně selektovány jsou thymocyty, které s nízkou afinitou rozeznávají MHC gp, ty si pak zachovávají expresi CD4 či CD8 (váže-li příslušný TCR MHC gp I či II) – tyto zralé T bb. (medulární thymocyty) opouštějí thymus a usidlují se v sekundárních lymfoidních orgánech 98% pro-thymocytů v thymu během svého vývoje zahyne
 se kterými se setká na povrchu thymových buněk dostane signály vedoucí k apoptotické smrti buňky. PAE buňky (peripherial antigen expressing cells) Pozitivní selekce – eliminace buněk s nefunkčním TCR. Pozitivně selektovány jsou thymocyty, které s nízkou afinitou rozeznávají MHC gp, ty si pak zachovávají expresi CD4 či CD8 (váže-li příslušný TCR MHC gp I či II) – tyto zralé T bb. (medulární thymocyty) opouštějí thymus a usidlují se v sekundárních lymfoidních orgánech. 98% pro-thymocytů v thymu během svého vývoje zahyne.")
43
Vývoj T lymfocytů

44
Povrchové znaky T lymfocytů
TCR – rozpoznává Ag peptid v komplexu s MHC gp CD3 – součást TCR, účast při přenosu signálu CD4 nebo CD8 – koreceptory, napomáhají k vazbě na MHC gp CD28 – kostimulační receptor; váže CD80, CD86 CTLA-4 (CD152) – inhibiční receptor; váže CD80, CD86
 – inhibiční receptor; váže CD80, CD86.")
46
Subpopulace T lymfocytů
ab-T lymfocyty – mají TCRab, většinový typ (95%), k vývoji potřebují thymus, rozeznávají antigeny v komplexu MHC gp-peptid gd-T lymfocyty – (5%) mohou se vyvíjet i mimo thymus, některé jsou schopny rozpoznat nativní Ag, uplatňují se při obraně kůže a sliznic Intraepiteliální T lymfocyty NK-T lymfocyty – rozeznávají komplexy CD1 molekul s lipidy
, k vývoji potřebují thymus, rozeznávají antigeny v komplexu MHC gp-peptid. gd-T lymfocyty – (5%) mohou se vyvíjet i mimo thymus, některé jsou schopny rozpoznat nativní Ag, uplatňují se při obraně kůže a sliznic. Intraepiteliální T lymfocyty. NK-T lymfocyty – rozeznávají komplexy CD1 molekul s lipidy.")
47
ab-T lymfocyty TH3 – TGFb
Exprimující koreceptor CD4 (koreceptor pro MHC gp II) Jde o prekurzory pomocných T buněk (TH), ty lze rozdělit podle produkce cytokinů na : TH0 – produkují směs cytokinů jako TH1 a TH2 TH1 – IL-2, IFNg (pomoc makrofágům) TH2 – IL-4, IL-5, IL-6, IL-10 (pomoc B lymfocytům) TH3 – TGFb Treg – regulační T lymfocyty, potlačují aktivitu TH1 a částečně fungují jako TS, potlačování autoreaktivních klonů T lymfocytů; produkují IL-10 a TGFb
 Jde o prekurzory pomocných T buněk (TH), ty lze rozdělit podle produkce cytokinů na : TH0 – produkují směs cytokinů jako TH1 a TH2 TH1 – IL-2, IFNg (pomoc makrofágům) TH2 – IL-4, IL-5, IL-6, IL-10 (pomoc B lymfocytům) TH3 – TGFb. Treg – regulační T lymfocyty, potlačují aktivitu TH1 a částečně fungují jako TS, potlačování autoreaktivních klonů T lymfocytů; produkují IL-10 a TGFb.")
48
ab-T lymfocyty Exprimující koreceptor CD8 (koreceptor pro MHC gp I) Jde o prekurzory cytotoxických T buněk (TC), či supresorových T buněk (TS) TC – rozeznávají buňky napadené viry či jinými intracelulárními parazity a některé nádorové buňky TS – potlačují funkce ostatních lymfocytů
 Jde o prekurzory cytotoxických T buněk (TC), či supresorových T buněk (TS) TC – rozeznávají buňky napadené viry či jinými intracelulárními parazity a některé nádorové buňky. TS – potlačují funkce ostatních lymfocytů.")
49
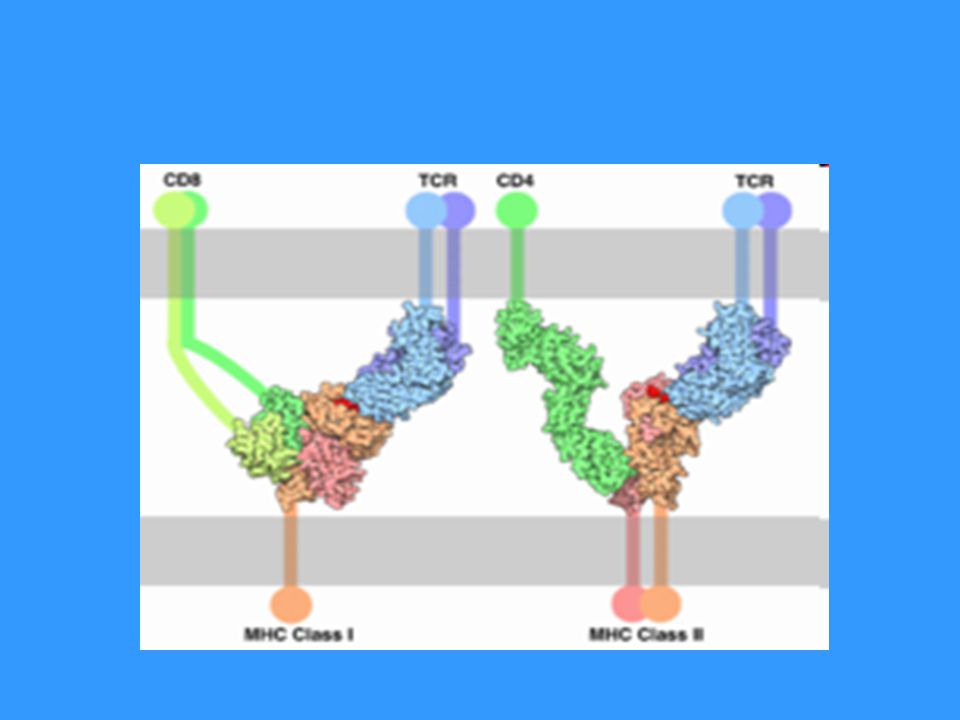
TCR TCR (T cell receptor) se skládá z modulu rozeznávajícího Ag a asociovaného CD3 komplexu, který je nezbytný pro přenos signálu, je propojen s PTK (Src) Modul rozeznávající Ag je tvořen 2 řetězci a a b (g,d), N-terminální části tvoří vazebné místo pro Ag T lymfocyty (ab) rozeznávají komplex MHC gp-Ag peptid na povrchu APC, při rozpoznání spolupracuje TCR s koreceptory CD4 (pomáhá vázat MHC gp II) nebo CD8 (pomáhá vázat MHC gp I) Aktivace přes TCR a CD28 vede k proliferaci a diferenciaci v efektorové buňky CD28 – receptor kostimulačního signálu, váže ligandy CD80 a CD86, které jsou na povrchu APC
 se skládá z modulu rozeznávajícího Ag a asociovaného CD3 komplexu, který je nezbytný pro přenos signálu, je propojen s PTK (Src) Modul rozeznávající Ag je tvořen 2 řetězci a a b (g,d), N-terminální části tvoří vazebné místo pro Ag. T lymfocyty (ab) rozeznávají komplex MHC gp-Ag peptid na povrchu APC, při rozpoznání spolupracuje TCR s koreceptory CD4 (pomáhá vázat MHC gp II) nebo CD8 (pomáhá vázat MHC gp I) Aktivace přes TCR a CD28 vede k proliferaci a diferenciaci v efektorové buňky. CD28 – receptor kostimulačního signálu, váže ligandy CD80 a CD86, které jsou na povrchu APC.")
50
Spolupráce TCR s koreceptory CD4 a CD8

51
Vznik TCR Obdoba s tvorbou BCR
Řetězce b a d - odpovídají genovému komplexu IgH imunoglobulinů V, D, J, C segmenty Řetězce a a g - odpovídají genům pro L řetězce imunoglobulinů V, J, C segmenty Přeskupování genů probíhá podobně jako u BCR a provádějí je shodné rekombinázy u genů pro TCR nedochází k somatickým mutacím a afinitní maturaci

52
TH1 imunitní odpověď

53
TH1 imunitní odpověď – zánětlivá reakce
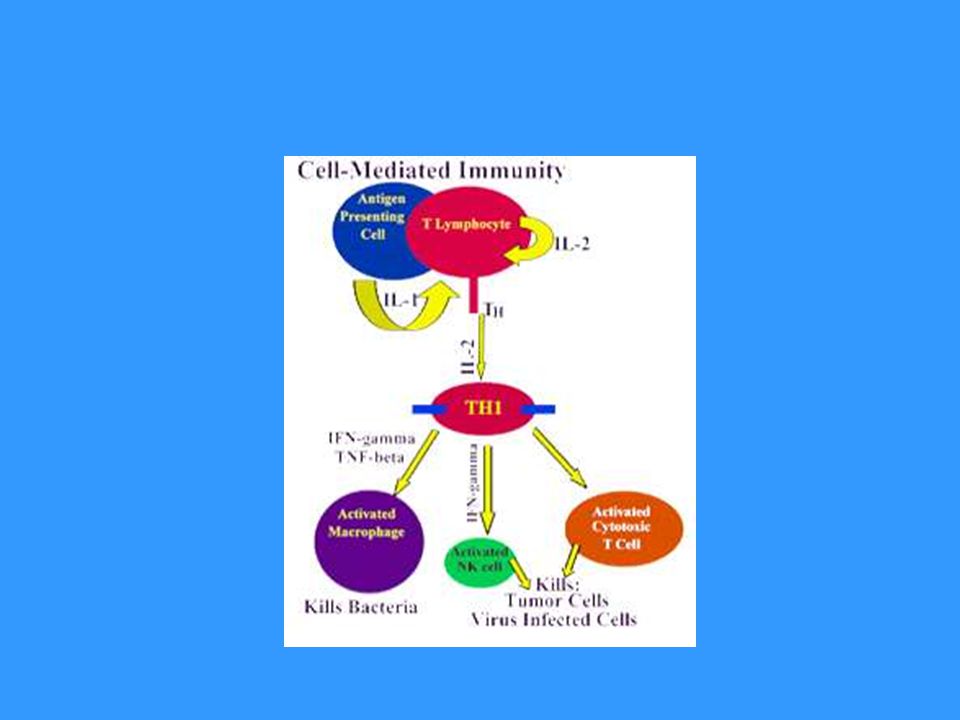
Základní funkcí TH1 buněk je spolupráce s makrofágy a jejich přeměna v aktivované, které jsou schopny produkovat NO, pomocí kterého likvidují své intracelulární parazity Pro přeměnu makrofágů v aktivované jsou nezbytné cytokiny (IFNg) produkované TH1 buňkami Aktivované makrofágy secernují některé cytokiny ( IL-1, TNF…), které napomáhají ke stimulaci T buněk a stimulují lokální zánět, který napomáhá potlačení infekce Vzájemné působení TH1 buněk a makrofágů je základním mechanismem imunopatologické reakce opožděného typu (DTH- delayed type hypersensitivity)
 produkované TH1 buňkami. Aktivované makrofágy secernují některé cytokiny ( IL-1, TNF…), které napomáhají ke stimulaci T buněk a stimulují lokální zánět, který napomáhá potlačení infekce. Vzájemné působení TH1 buněk a makrofágů je základním mechanismem imunopatologické reakce opožděného typu (DTH- delayed type hypersensitivity)")
54
Infikovaný makrofág produkuje fragmenty proteinů pocházející z intracelulárního parazita, některé jsou prezentovány na jeho povrchu pomocí MHC gp II Makrofágy a dendritické buňky stimulované některými mikroorganismy produkují IL-12 Prekurzor TH , který rozpozná infikovaný makrofág a obdrží signály přes TCR, CD 28 a receptor pro IL-12 a další adhezivní a signalizační molekuly proliferuje a diferencuje se na efektorové TH1 buňky, které produkují IFNg a IL-2. IFNg podporuje přeměnu makrofágů v aktivované IL-2 je autokrinní růstový faktor TH1 lymfocytů.

55
Interakce mezi APC a prekurzorem TH

56
Diferenciace TH0

57
Kostimulační signál

59
TH2 imunitní odpověď

60
TH2 imunitní odpověď – pomoc B lymfocytům
Základní funkcí TH2 buněk je spolupráce s B lymfocyty (které byly stimulovány Ag) prostřednictvím cytokinů (IL-4, IL-5, IL-6) a přímého mezibuněčného kontaktu Pro stimulaci B lymfocytů je většinou potřeba spolupráce mezi APC → TH2 buňkou → B lymfocytem V případě tzv. minimálního modelu, pokud se z B lymfocytu stane dobrá APC (CD80, CD86), stačí spolupráce mezi TH2 buňkou → B lymfocytem
 prostřednictvím cytokinů (IL-4, IL-5, IL-6) a přímého mezibuněčného kontaktu. Pro stimulaci B lymfocytů je většinou potřeba spolupráce mezi APC → TH2 buňkou → B lymfocytem. V případě tzv. minimálního modelu, pokud se z B lymfocytu stane dobrá APC (CD80, CD86), stačí spolupráce mezi TH2 buňkou → B lymfocytem.")
61
Prekurzor TH , který rozpozná infikovaný makrofág a dostane signály přes TCR, CD 28, receptor pro IL-4 a receptor pro IL-2 a další adhezivní a signalizační molekuly proliferuje a diferencuje se na efektorové TH2, které poskytují B lymfocytům pomocné signály pomocí sekretovaných cytokinů IL-4, IL-5, IL-6 a prostřednictvím adhezivních molekul CD 40L, které se váží se na kostimulační receptor B lymfocytů CD 40 Interakce mezi CD40 (B lymfocyt) a CD40L (TH2 buňka) je nezbytná pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových buněk IL-4, IL-5, IL-6 : stimulace B lymfocytů
 a CD40L (TH2 buňka) je nezbytná pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových buněk. IL-4, IL-5, IL-6 : stimulace B lymfocytů.")
63
Pomoc B lymfocytům Specifická přímá pomoc B lymfocytům:
TH2 lymfocyt poskytuje pomoc B lymfocytům, které byly stimulovány stejným Ag, který vyvolal vznik TH2 Ke stimulaci sekrece cytokinů TH2 buňce stačí signál přes TCR (signál přes kostimulační receptor CD28 již není nutný) Jeden klon TH2 buněk může poskytovat specifickou pomoc B lymfocytům různých specifit (musí prezentovat příslušné Ag peptidy pomocí MHC gp II, které rozpoznáváTCR)
 Jeden klon TH2 buněk může poskytovat specifickou pomoc B lymfocytům různých specifit (musí prezentovat příslušné Ag peptidy pomocí MHC gp II, které rozpoznáváTCR)")
64
Přímá pomoc B lymfocytům

65
Pomoc B lymfocytům Nepřímá pomoc B lymfocytům („bystander help“):
TH2 lymfocyt poskytuje pomoc B lymfocytům, které byly stimulovány jiným Ag, než který vyvolal vznik TH2 Kontakt mezi TH2 buňkou → B lymfocytem prostřednictvím adhezivních molekul, sekrece cytokinů, vazba CD40-CD40L Nebezpečí aktivace autoreaktivních B lymfocytů

66
Vzájemná regulace aktivit TH1 versus TH2
Zda se prekurzorové TH lymfocyty budou vyvíjet v TH1 či TH2 rozhodne poměr cytokinů IL-12 a IL-4 IL-12 je produkován makrofágy a dendritickými buňkami stimulovanými některými mikroorganismy IL-4 je produkován bazofily a mastocyty Cytokiny produkované TH1 (hlavně IFNg) inhibují vývoj TH2 a stimulují vývoj TH1 (IL-2 stimuluje i TH2) Cytokiny produkované TH2 (IL-4, IL-10) inhibují vývoj TH1 a stimulují vývoj TH2 Vývoj TH3 je stimulován specifickým cytokinovým prostředím IL-4, IL-10, TGFb; TH3 produkují TGFb, spolupracují s B lymfocyty v MALT
 inhibují vývoj TH2 a stimulují vývoj TH1 (IL-2 stimuluje i TH2) Cytokiny produkované TH2 (IL-4, IL-10) inhibují vývoj TH1 a stimulují vývoj TH2. Vývoj TH3 je stimulován specifickým cytokinovým prostředím IL-4, IL-10, TGFb; TH3 produkují TGFb, spolupracují s B lymfocyty v MALT.")
68
Imunitní reakce založená na TC

69
Stimulace cytotoxických T lymfocytů
TC rozeznávají buňky infikované viry či jinými intracelulárními parazity a některé nádorové buňky Prekurzor TC , který rozpozná komplex MHC gp I- antigenní peptid na povrchu APC přes TCR a dostane signály přes CD 28 proliferuje a diferencuje se na klon zralých efektorových cytotoxických buněk (CTL); toto probíhá za pomoci TH1 lymfocytů produkujících IL-2 Efektorové TC jsou rozneseny krevním oběhem do tkání, k aktivaci cytotoxických mechanismů stačí signál přes TCR (signál přes kostimulační receptor CD28 již není nutný)
; toto probíhá za pomoci TH1 lymfocytů produkujících IL-2. Efektorové TC jsou rozneseny krevním oběhem do tkání, k aktivaci cytotoxických mechanismů stačí signál přes TCR (signál přes kostimulační receptor CD28 již není nutný)")
70
Profesionální APC jsou dendritické buňky nebo makrofágy, které jsou infikovány virem, nebo pohltili antigeny z odumřelé virem infikované, nádorové nebo stresované buňky Aby APC mohla aktivovat prekurzor TC ,musí být sama stimulována kontaktem s TH buňkami přes CD 40, poté začne dendritická buňka exprimovat CD 80, CD86 a sekretovat cytokiny (IL-1, IL-12) = přeměna klidové APC v aktivovanou
 = přeměna klidové APC v aktivovanou.")
71
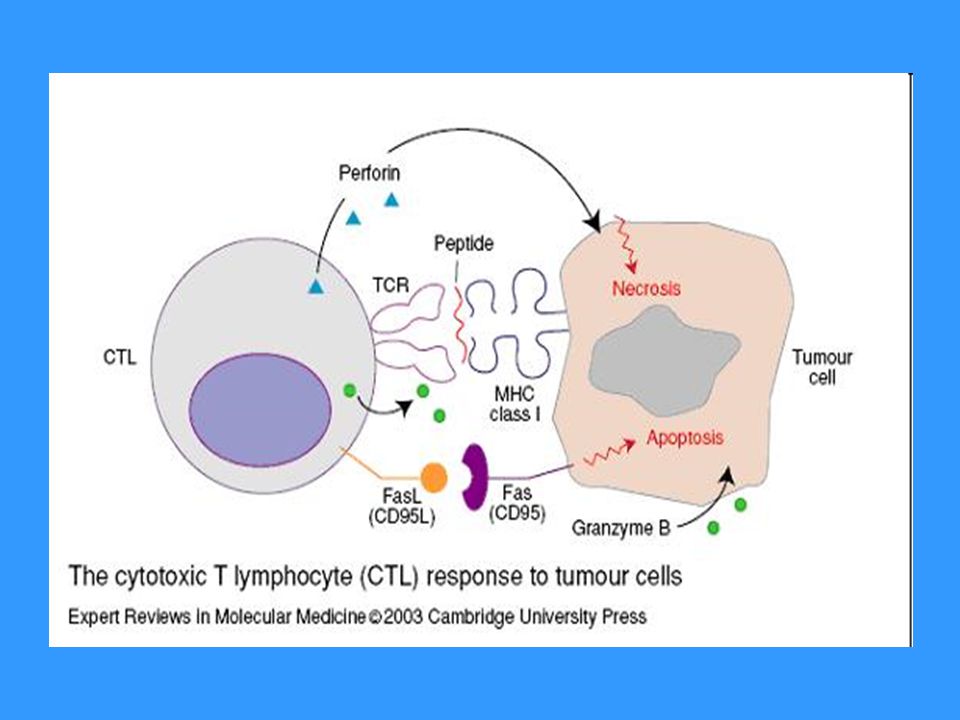
Efektorové mechanismy TC
Cytotoxická granula obsahující perforin a granzymy (perforin vytváří póry v cytoplazmatické membráně napadené buňky,v některých případech může dojít k osmotické lýze napadené buňky, vytvořenými póry se do buňky dostávají granzymy, které aktivují caspázy, což vede k apoptóze napadené buňky Fas-ligand (FasL) – který se váže na apoptotický receptor Fas (CD95) přítomný na povrchu mnoha různých bb. (i na povrchu TC) TNFb
 – který se váže na apoptotický receptor Fas (CD95) přítomný na povrchu mnoha různých bb. (i na povrchu TC) TNFb.")
73
Rozdílná aktivace Tc a NK

74
B lymfocyty

75
B lymfocyty B-lymfocyty (B buňky) jsou buňky zodpovědné především za specifickou, protilátkami zprostředkovanou imunitní odpověď. Mají rovněž velký význam pro imunitní paměť (využívá se při očkování). B-lymfocyty rozpoznávají nativní antigen pomocí BCR (B cell receptor) Příslušný B-lymfocyt, na jehož receptorech došlo k vazbě antigenu, je stimulován k pomnožení a diferenciaci na efektorové neboli plazmatické bb., které produkují velké množství protilátek stejné specifity, jako je BCR (jde vlastně o tentýž protein v rozpustné formě), vážou se tedy na stejný antigen. Z části stimulovaných B-lymfocytů se diferencují paměťové buňky.
 jsou buňky zodpovědné především za specifickou, protilátkami zprostředkovanou imunitní odpověď. Mají rovněž velký význam pro imunitní paměť (využívá se při očkování). B-lymfocyty rozpoznávají nativní antigen pomocí BCR (B cell receptor) Příslušný B-lymfocyt, na jehož receptorech došlo k vazbě antigenu, je stimulován k pomnožení a diferenciaci na efektorové neboli plazmatické bb., které produkují velké množství protilátek stejné specifity, jako je BCR (jde vlastně o tentýž protein v rozpustné formě), vážou se tedy na stejný antigen. Z části stimulovaných B-lymfocytů se diferencují paměťové buňky.")
76
Povrchové znaky B lymfocytů
CD 10 - nezralý B lymfocyt CD 19 - charakteristický povrchový znak B lymfocytů CD 20 - na povrchu Ig-pozitivních B lymfocytů IgM, IgD - BCR MHC gp II.třídy - Ag prezentující molekuly CD 40 – kostimulační receptor

77
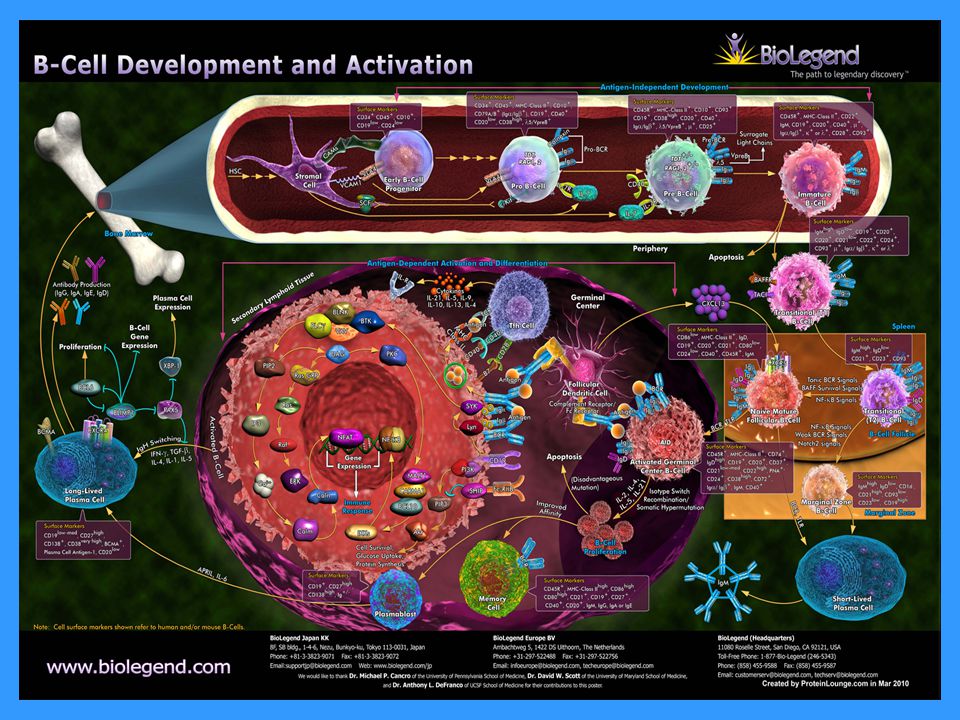
Vývoj B lymfocytů Vývoj B lymfocytů probíhá v kostní dřeni a dokončuje se posetkání s Ag v sekundárních lymfatických orgánech. Pluripotentní hematopoetická kmenová buňka Progenitor B lymfocytu → zahájení rekombinačních procesů, které vedou ke vzniku velkého množství klonů B lymfocytů s individuálně specifickými BCR Pre B lymfocyt → exprese pre-B receptoru (tvořen H(m) řetězcem a náhradním L řetězcem) Nezralý B lymfocyt → exprese povrchového IgM (BCR); v této fázi vývoje dochází k eliminaci autoreaktivních klonů Zralý B lymfocyt → exprese povrchového IgM a IgD (BCR)
 řetězcem a náhradním L řetězcem) Nezralý B lymfocyt → exprese povrchového IgM (BCR); v této fázi vývoje dochází k eliminaci autoreaktivních klonů. Zralý B lymfocyt → exprese povrchového IgM a IgD (BCR)")
78
BCR BCR se skládá z povrchového imunoglobulinu (IgM, IgD – H řetězce jsou transmembránové ; rozeznává Ag) a asociovaných signalizačních molekul (Iga a Igb), které jsou asociovány s cytoplazmatickými protein tyrosin-kinázami (PTK) skupiny Src Po současném navázání Ag na 2 či více BCR dojde k přiblížení PTK, vzájemné fosforylaci a fosforylaci dalších cytoplazmatických proteinů, což vede ke změnám transkripce genů, proliferaci, diferenciaci a sekreci protilátek Signály spuštěné vazbou Ag na BCR mohou být zesíleny spoluprací s CR2, který váže C3dg (opsonin)
 a asociovaných signalizačních molekul (Iga a Igb), které jsou asociovány s cytoplazmatickými protein tyrosin-kinázami (PTK) skupiny Src. Po současném navázání Ag na 2 či více BCR dojde k přiblížení PTK, vzájemné fosforylaci a fosforylaci dalších cytoplazmatických proteinů, což vede ke změnám transkripce genů, proliferaci, diferenciaci a sekreci protilátek. Signály spuštěné vazbou Ag na BCR mohou být zesíleny spoluprací s CR2, který váže C3dg (opsonin)")
79
Eliminace autoreaktivních klonů B lymfocytů
Při náhodném přeskupováním genů, spojovacích nepřesnostech, párování H-L a somatických mutací mohou vzniknout i klony B lymfocytů nesoucí autoreaktivní receptory a produkující autoreaktivní protilátky. Většina autoreaktivních B lymfocytů je eliminována na úrovni nezralých B lymfocytů (v kostní dřeni), jestliže svým BCR váží autoantigen s dostatečnou afinitou, obdrží signál vedoucí k apoptotické smrti. Pokud touto eliminací projdou některé autoreaktivní klony, jejich autoreaktivita se většinou neprojeví, protože k jejich aktivaci chybí příslušné TH lymfocyty, mnohé autoantigeny jsou kryptické, či se vyskytují v malé koncentraci a jsou imunitním systémem ignorovány.
, jestliže svým BCR váží autoantigen s dostatečnou afinitou, obdrží signál vedoucí k apoptotické smrti. Pokud touto eliminací projdou některé autoreaktivní klony, jejich autoreaktivita se většinou neprojeví, protože k jejich aktivaci chybí příslušné TH lymfocyty, mnohé autoantigeny jsou kryptické, či se vyskytují v malé koncentraci a jsou imunitním systémem ignorovány.")
80
Setkání B lymfocytu s Ag v sekundárních lymfatických orgánech

82
Ontogeneze tvorby protilátek
Syntéza specifických protilátek začíná kolem týdne gestace, celková koncentrace IgA a IgM zůstává až do porodu neměřitelná, IgG se začínají tvořit až po porodu B lymfocyty na imunizaci reagují převážně tvorbou IgM, přesmyk na jiné izotypy je pomalejší Pozvolný nárůst tvorby vlastních IgG za poklesu mateřských IgG (kolem 3.-6.měs.)
")
83
Ontogeneze tvorby protilátek
Koncentrace IgM dosahuje hodnot srovnatelných s dospělými v 1.-3.roce života, IgG+A mezi r. Protilátková reakce na polysacharidové antigeny se objevuje až kolem 2.roku života Ve stáří je slabší protilátková odpověď na nové podněty a vyšší produkce autoprotilátek

84
Imunitní odpověď založená na protilátkách

85
Protilátková reakce vyvolaná:
Antigeny nezávislými na T lymfocytech Vyvolávají převážně tvorbu IgM protilátek Jedná se zvláště o bakteriální polysacharidy, lipopolysacharidy a polymerní formy proteinů Antigeny závislými na T lymfocytech Reakce na tyto Ag probíhá ve dvou fázích – primární a sekundární odpověď Tyto reakce mohou být od sebe odděleny či mohou na sebe bezprostředně navazovat Vyvolávají tvorbu paměťových buněk a vznik vysokoafinních protilátek

86
T-nezávislá a T-závislá imunitní odpověď

87
Protilátková reakce vyvolaná antigeny závislými na T lymfocytech
Primární fáze protilátkové reakce - při prvním setkání s Ag Probíhá v sekundárních lymfatických orgánech Stimulace B lymfocytu vazbou Ag na BCR Pohlcení Ag APC a jeho prezentace prostřednictvím MHC gp II prekurzorům TH buněk → vznik klonu antigenně specifických TH2 buněk, které poskytují pomoc příslušným B lymfocytům, což vede k jejich proliferaci, diferenciaci na plazmatické bb (produkují Ab) a na paměťové bb Plazmatické bb jsou rozneseny oběhovým systémem do organismu (zvláště kostní dřeně) Protilátky produkované v primární fázi (za 3-4 dny) jsou IgM a mají nízkou afinitu k Ag, s Ag tvoří imunokomplexy Imunokomplexy jsou zachytávány v sekundárních lymfoidních orgánech na povrchu FDC (folikulárně dendritických bb) – bb prezentující Ag B lymfocytům
 a na paměťové bb. Plazmatické bb jsou rozneseny oběhovým systémem do organismu (zvláště kostní dřeně) Protilátky produkované v primární fázi (za 3-4 dny) jsou IgM a mají nízkou afinitu k Ag, s Ag tvoří imunokomplexy. Imunokomplexy jsou zachytávány v sekundárních lymfoidních orgánech na povrchu FDC (folikulárně dendritických bb) – bb prezentující Ag B lymfocytům.")
88
Protilátková reakce vyvolaná antigeny závislými na T lymfocytech
Sekundární fáze protilátkové reakce – po rozpoznání Ag na FDC Vznikne-li dostatečné množství imunokomplexů na FDC a jsou-li rozpoznány B lymfocyty Germinální reakce - pod vlivem signálů od FDC (Ag) a TH2 bb (CD40L, cytokiny) dochází opět k proliferaci a diferenciaci B lymfocytů doprovázené somatickými mutacemi → vznik klonů B lymfocytů s novými BCR→ přežívají pouze B lymfocyty s BCR s nejvyšší afinitou k Ag = afinitní maturace protilátek Dochází také k izotypovému přesmyku, jaké izotypy vzniknou určuje cytokinové prostředí
 a TH2 bb (CD40L, cytokiny) dochází opět k proliferaci a diferenciaci B lymfocytů doprovázené somatickými mutacemi → vznik klonů B lymfocytů s novými BCR→ přežívají pouze B lymfocyty s BCR s nejvyšší afinitou k Ag = afinitní maturace protilátek. Dochází také k izotypovému přesmyku, jaké izotypy vzniknou určuje cytokinové prostředí.")
89
V sekundární fázi imunitní reakce vznikají protilátky s vyšší afinitou k Ag a s jinými efektorovými vlastnostmi závislými na izotypu, vznikají také paměťové bb pro další setkání s Ag Protilátky v organismu po primární infekci přetrvávají po dlouhou dobu Kontakt mezi CD40 (B lymfocyt) a CD40L (TH2 lymfocyt) je nezbytný pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových bb.
 a CD40L (TH2 lymfocyt) je nezbytný pro zahájení somatických mutací, izotypového přesmyku a vznik paměťových bb.")
91
Protilátková odpověď Primární odpověď
- následuje po prvním kontaktu s antigenem, kdy ještě nejsou přítomny paměťové buňky - tvoří se převážně nízkoafinní IgM, postupně se tvoří ostatní izotypy s vyšší afinitou Sekundární odpověď - následuje po opakovaných setkáních se stejným antigenem, jsou již přítomny paměťové buňky - hladina IgM protilátek je nižší, rychleji se tvoří ostatní izotypy, protilátky mají vyšší afinitu k antigenu, protože jsou aktivovány paměťové buňky

92
Dynamika tvorby protilátek při primární a sekundární odpovědi

Podobné prezentace



















>")



