Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Komplement, antigeny Martin Liška

2
Komplement Soubor nejméně 20 sérových proteinů, působících jako humorální složka nespecifické imunity Složky komplementu jsou v séru přítomny v inaktivní formě Aktivace komplementu má kaskádovitý charakter (= každý protein aktivuje následující) a vede k řadě fyziologických i patofyziologických účinků Proteiny komplementu jsou syntetizovány především v játrech, v menší míře též tkáňovými makrofágy a fibroblasty
 a vede k řadě fyziologických i patofyziologických účinků. Proteiny komplementu jsou syntetizovány především v játrech, v menší míře též tkáňovými makrofágy a fibroblasty.")
3
Komplement - nomenklatura
C1q, C1r, C1s, C2-9 Faktor B, D Properdin Regulační proteiny (C1-inhibitor, faktor I, C4bBP, faktor H, S protein, inaktivátor anafylatoxinu)
")
4
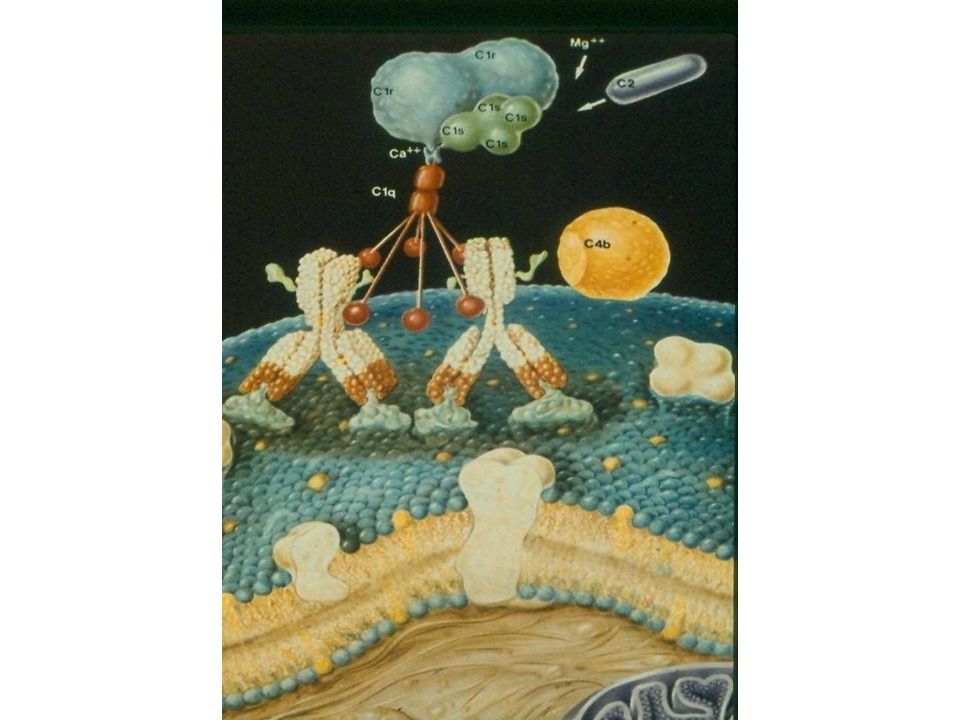
Klasická cesta aktivace komplementu I
Dráha aktivována komplexem antigen-protilátka: na začátku se C1-složka naváže na Fc-fragment Ig (IgG (ale nikoli IgG4) nebo IgM), ovšem s nativními molekulami Ig nereaguje C1-složka – obsahuje 3 polypeptidy (C1q, C1r, C1s); C1q se jako první váže na Ig (C1q je multivalentní, k aktivaci se musí navázat na ≥ 2 monomery Ig → aktivuje proenzym C1r → štěpí proenzym C1s → schopen štěpit C4-složku
 nebo IgM), ovšem s nativními molekulami Ig nereaguje. C1-složka – obsahuje 3 polypeptidy (C1q, C1r, C1s); C1q se jako první váže na Ig (C1q je multivalentní, k aktivaci se musí navázat na ≥ 2 monomery Ig → aktivuje proenzym C1r → štěpí proenzym C1s → schopen štěpit C4-složku.")
6
Klasická cesta aktivace komplementu II
Aktivovaná složka C1s štěpí C4 na C4a (anafylatoxin, odštěpí se) + C4b → naváže se na buněčnou membránu → další složka komplementové kaskády je přístupná působení aktivovaného C1s C4b + C2 + C1s → odštěpí C2a → enzymaticky aktivní molekulární komplex C4b2b (= C3-konvertáza klasické dráhy) Vznik C3-konvertázy je styčným bodem všech drah aktivace komplementu
 + C4b → naváže se na buněčnou membránu → další složka komplementové kaskády je přístupná působení aktivovaného C1s. C4b + C2 + C1s → odštěpí C2a → enzymaticky aktivní molekulární komplex C4b2b (= C3-konvertáza klasické dráhy) Vznik C3-konvertázy je styčným bodem všech drah aktivace komplementu.")
12
Alternativní cesta aktivace komplementu I
Považovaná za primitivní „zkratkový“ mechanismus, který k aktivaci komplementu nepotřebuje C1, C2, C4 Dráha aktivována reakcí komplementu s některými produkty mikroorganismů (polysacharidy – např.lipopolysacharidy G- baktérií, kys.teichoová G+ baktérií, zymosan kvasinek, povrchové složky některých živočišných parazitů) nebo jinými cizorodého materiálu
 nebo jinými cizorodého materiálu.")
13
Alternativní cesta aktivace komplementu II
C3 se samovolně štěpí na C3a + C3b, ale za normálních okolností je inaktivován V tomto případě se však C3b naváže na cizorodý povrch → reakce s B → odštěpení Ba (= působí chemotakticky na neutrofily) → C3bBb (= C3-konvertáza alternativní dráhy) – stabilizována properdinem (P)
 → C3bBb (= C3-konvertáza alternativní dráhy) – stabilizována properdinem (P)")
14
Lektinová cesta aktivace komplementu
Dráha aktivována vazbou MBL (mannose-binding lectin) na povrch mikroorganismu → C4 → C4a (odštěpí se) + C4b → reaguje s C2 → odštěpí se C2b → komplex C4b2a (= C3-konvertáza)
 na povrch mikroorganismu → C4 → C4a (odštěpí se) + C4b → reaguje s C2 → odštěpí se C2b → komplex C4b2a (= C3-konvertáza)")
15
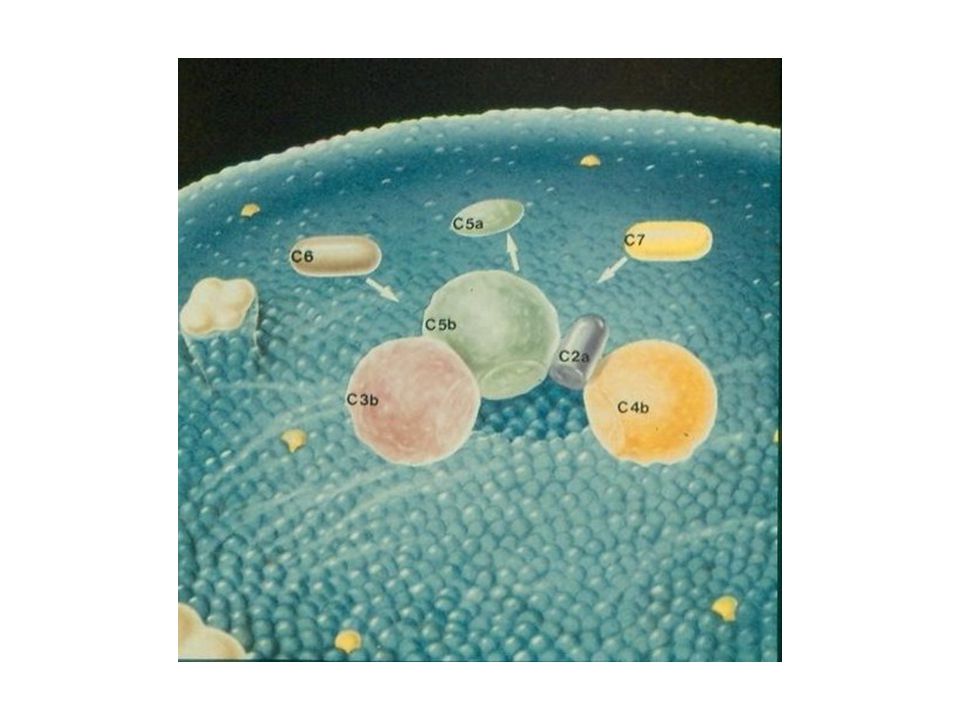
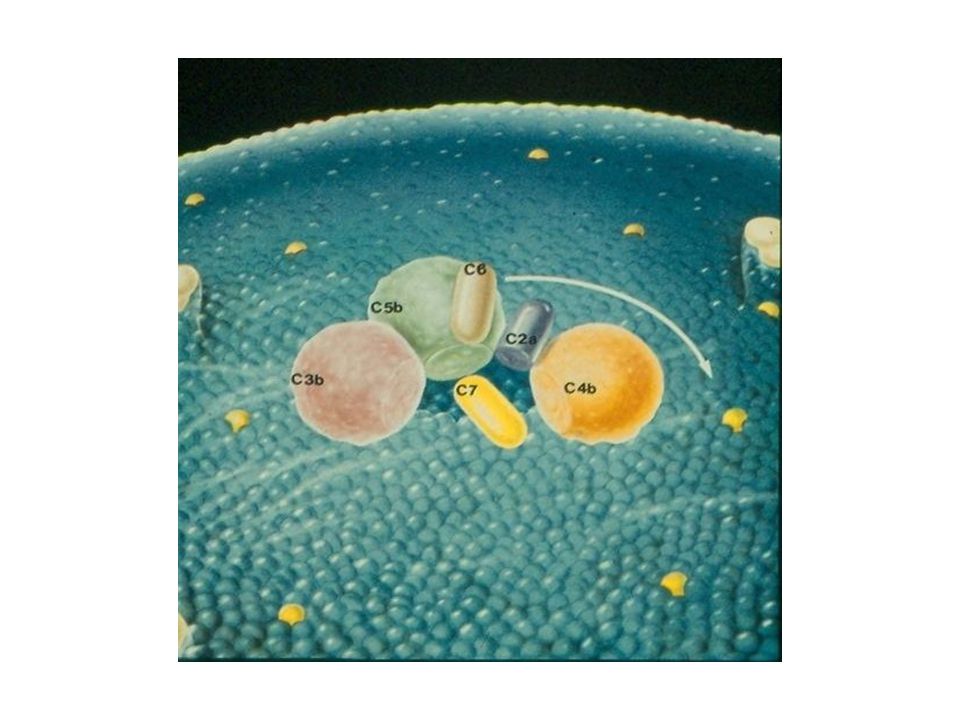
Další průběh komplementové kaskády I
C3-konvertáza působí: a/ vznik štěpů C3a, C4a, C5a - působí jako peptidové mediátory zánětu působí chemotakticky na fagocyty b/ vznik C3b váže se na komplementové receptory a působí opsonizaci mikrobů a imunokomplexů

18
Další průběh komplementové kaskády II
c/vznik MAC (membrane attack complex), složeného z terminálních složek komplementu (C5b, C6, C7, C8, C9), který atakuje membránu buněk (např. mikroorganismů) a působí jejich osmolýzu
, složeného z terminálních složek komplementu (C5b, C6, C7, C8, C9), který atakuje membránu buněk (např. mikroorganismů) a působí jejich osmolýzu.")
23
Komplementové receptory
vážou fragmenty složek komplementu CR1 – na erytrocytech, granulocytech, monocytech a B lymfocytech - význam pro odstraňování IK CR2 – na B lymfocytech a FDC - zajišťuje funkční spojení mezi nespecifickou a specifickou imunitou CR3, CR4 – na fagocytech - účast v opsonizaci

24
Regulační mechanismy komplementu I
Některé proteiny enzymaticky působí na složky komplementu a inaktivují je (faktor I – inaktivuje C3b; inaktivátor anafylatoxinu – inaktivuje anafylatoxiny (C3a, C4a, C5a)) Některé sérové proteiny se vážou na složky komplementu a inhibují je (C1-INH – inhibuje C1, při jeho deficitu → HAE = rekurentní epizody lokálního edému; faktor H – spolu s faktorem I inhibuje C3b; S protein – váže se na C5b67 → zabraňuje vazbě MAC na buněčnou membránu)
) Některé sérové proteiny se vážou na složky komplementu a inhibují je (C1-INH – inhibuje C1, při jeho deficitu → HAE = rekurentní epizody lokálního edému; faktor H – spolu s faktorem I inhibuje C3b; S protein – váže se na C5b67 → zabraňuje vazbě MAC na buněčnou membránu)")
25
Regulační mechanismy komplementu II
Regulační proteiny v buněčné mebráně (DAF (decay-accelerating factor) – stejná funkce jako faktor H → inaktivace C3b a C4b; membránový kofaktorový protein – kofaktor inaktivace C4b a C3b)
 – stejná funkce jako faktor H → inaktivace C3b a C4b; membránový kofaktorový protein – kofaktor inaktivace C4b a C3b)")
26
Funkce komplementu I C3a, C5a – anafylatoxiny (= uvolnění histaminu a dalších vazoaktivních látek z bazofilů a mastocytů, zvýšená permeabilita kapilár) C3b, C4b – opsonizace (váže IK na makrofágy a neutrofily a podporuje tím jejich fagocytózu; váže IK na erytrocyty a podporuje jejich odstraňování v játrech a slezině) C5a – chemotaxe (přitahuje fagocyty do míst zánětu a zvyšuje jejich aktivitu )
 C3b, C4b – opsonizace (váže IK na makrofágy a neutrofily a podporuje tím jejich fagocytózu; váže IK na erytrocyty a podporuje jejich odstraňování v játrech a slezině) C5a – chemotaxe (přitahuje fagocyty do míst zánětu a zvyšuje jejich aktivitu )")
27
Funkce komplementu II C8, C9 – součást MAC Ba – chemotaxe neutrofilů
Bb – aktivace makrofágů

28
Funkce komplementu - přehled
Zánět (degranulace mastocytů, chemotaxe, zvýšená cévní permeabilita, vycestování polymorfonukleárů, kontrakce hladké svaloviny, aktivace polymorfonukleárů, NK-bb. a makrofágů) Odstraňování IK Lýza buněk (G- baktérie, Protozoa, některé viry) Neutralizace virů Opsonizace
 Odstraňování IK. Lýza buněk (G- baktérie, Protozoa, některé viry) Neutralizace virů. Opsonizace.")
29
Komplement – přehled I Alternativní a lektinová cesta aktivace jasně zapadají do systému nespecifické imunity, zatímco klasická cesta aktivace je závislá na specifické imunitní odpovědi (spouští ji reakce antigen-protilátka) Funkce komplementu ve třech bodech: 1/ C3b obaluje mikroby, a tím podporuje jejich vazbu na fagocyty (prostřednictvím receptorů pro C3b)
 Funkce komplementu ve třech bodech: 1/ C3b obaluje mikroby, a tím podporuje jejich vazbu na fagocyty (prostřednictvím receptorů pro C3b)")
30
Komplement – přehled II
2/některé produkty štěpení komplementu jsou chemoatraktanty, přitahujícími neutrofily a monocyty, a podporují zánět v místě aktivace komplementu 3/ tvorba proteinových polymerických komplexů (MAC), vyvolávajících osmolýzu nebo apoptózu mikrobů
, vyvolávajících osmolýzu nebo apoptózu mikrobů.")
31
Antigen Látka, kterou imunitní systém rozpoznává a reaguje na ni
Může pocházet z vnějšího prostředí (exoantigen), ale i z vlastních struktur (autoantigen) Obvykle proteiny nebo polysacharidy (lipidy a nukleové kyseliny jimi mohou být pouze v kombinaci s proteiny nebo polysacharidy)
, ale i z vlastních struktur (autoantigen) Obvykle proteiny nebo polysacharidy (lipidy a nukleové kyseliny jimi mohou být pouze v kombinaci s proteiny nebo polysacharidy)")
32
Hapteny Malé a dobře definované chemické struktury, které nejsou imunogenní ze své podstaty, ale reagují s preformovanými protilátkami, jejichž tvorba je indukována haptenem, spojeným s nosnou molekulou, která imunogenem je Typicky léky (např.penicilinová ATB, hydralazin)
")
33
Epitop Část antigenu, kterou rozpoznává imunitní systém (lymfocyty, Ig) U některých epitopů záleží především na jejich terciární struktuře (konformační epitop), u jiných na složení AMK (lineární epitop) Některé epitopy se nacházejí na povrchu antigenů (povrchové), některé uvnitř (interní) Zkříženě reagující antigeny – sdílí jeden a více stejných nebo podobných epitopů
, u jiných na složení AMK (lineární epitop) Některé epitopy se nacházejí na povrchu antigenů (povrchové), některé uvnitř (interní) Zkříženě reagující antigeny – sdílí jeden a více stejných nebo podobných epitopů.")
34
Reakce antigenu s protilátkou
Vazebná místa protilátek (paratop) tvoří nekovalentní komplexy s korespondujícími místy na molekulách antigenů (epitop) Uplatňují se přitom vodíkové, elektrostatické a hydrofobní vazby, van der Waalsovy síly Komplex antigen-protilátka je reverzibilní
 tvoří nekovalentní komplexy s korespondujícími místy na molekulách antigenů (epitop) Uplatňují se přitom vodíkové, elektrostatické a hydrofobní vazby, van der Waalsovy síly. Komplex antigen-protilátka je reverzibilní.")
35
Typy antigenů z hlediska prezentace antigenů I
1/ thymus dependentní antigeny častější, typicky proteiny specifická humorální imunitní odpověď na antigen vyžaduje spolupráci s T-lymfocyty, jinak není dost efektivní pomoc realizována ve formě cytokinů secernovaných T-lymfocyty

36
Typy antigenů z hlediska prezentace antigenů II
2/ thymus independentní antigeny u malého počtu antigenů může být tvorba protilátek indukována přímo bez spoluúčasti T-lymfocytů zjm.bakteriální lipopolysacharidy a polymerní formy proteinů (např.Haemophilus, Str.pneumoniae)
")
37
Superantigeny Antigenní struktury, schopné vyvolat imunitní odpověď T-lymfocytů vnější vazbou na molekuly MHC (tj.mimo obvyklé místo vazby) Stimulují polyklonálně a masivně Např.bakteriální toxiny (Staph.aureus, Str.pyogenes, Pseud.aeruginosa)
")
38
Sekvestrované antigeny
Antigeny, které jsou za normálních okolností před imunitním systémem ukryty a tudíž je nezná (např.oční čočka, testes) Jsou-li však při poškození „odkryty“, může na ně imunitní systém zareagovat (jedna z teorií vzniku autoimunitních procesů)
 Jsou-li však při poškození „odkryty , může na ně imunitní systém zareagovat (jedna z teorií vzniku autoimunitních procesů)")
39
Imunologicky privilegovaná místa
Některé tkáně bývají při allogenní transplantaci odhojovány mnohem méně (např.CNS, rohovka, gonády). Mechanismy ochrany před imunitním systémem: izolace od imunitního systému (hematoencefalická bariéra); preference Th2- a suprese Th1-reakcí; aktivní ochrana proti efektorovým T-lymfocytům Toto privilegované postavení však neplatí absolutně (viz RS)
. Mechanismy ochrany před imunitním systémem: izolace od imunitního systému (hematoencefalická bariéra); preference Th2- a suprese Th1-reakcí; aktivní ochrana proti efektorovým T-lymfocytům. Toto privilegované postavení však neplatí absolutně (viz RS)")
40
Velikost molekuly antigenů
Molekuly < 5 kDa nemohou vyvolat imunitní odpověď, optimální velikost molekuly antigenu pro vyvolání imunitní odpovědi je cca 40 kDa

41
Stupeň nepříbuznosti I
Antigen musí být pro imunitní systém „cizí“ Čím větší je fylogenetický rozdíl mezi jednotlivci, tím více jsou si, v imunologickém smyslu, „cizí“; na základě toho rozlišujeme následující typy antigenů:

42
Stupeň nepříbuznosti II
Autologní – antigeny stejného jedince (např. kožní štěp přenesený ze stehna na hrudník téhož jedince), takže nejsou cizí Syngenní – antigeny geneticky identických jedinců (např. jednovaječná dvojčata), takže nejsou cizí Allogenní (alloantigeny) – antigeny geneticky odlišných jedinců téhož druhu (např. ledvina matky transplantovaná její dceři), takže jsou cizí Xenogenní (heterogenní) – antigeny pocházející od jedinců různého druhu (např. transplantace opičí ledviny člověku), takže jsou cizí
, takže nejsou cizí. Syngenní – antigeny geneticky identických jedinců (např. jednovaječná dvojčata), takže nejsou cizí. Allogenní (alloantigeny) – antigeny geneticky odlišných jedinců téhož druhu (např. ledvina matky transplantovaná její dceři), takže jsou cizí. Xenogenní (heterogenní) – antigeny pocházející od jedinců různého druhu (např. transplantace opičí ledviny člověku), takže jsou cizí.")
Podobné prezentace






















 Mgr. Martin Šmíd.>")





