Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Patogenita Pathogenita je schopnost pathogena vyvolat infekční onemocnění organismu. Často je zaměňovaná za termín virulence , tento termín mnoho autorů ponechává k popisu relativního stupně poškození způsobeného pathogenem. Virulence je schopnost mikroorganismu napadat krevní oběh.

2
Základní pojmy Symbiosa – soužití dvou typů organismů
Komensalismus – jeden z partnerů má ze spojení určité výhody, ale druhý je neovlivněn. Propionibacterium sp. žije na povrchu kůže z kožního sekretu, ani neškodí, ani neprospívá Mutualismus – prospívá oběma partnerům. E.coli a člověk vitamin K a některé vitaminy B, vstřebávají se v tlustém střevě, hostitel chrání životní prostředí Parasitismus – jeden organismus získává výhody na úkor druhého. Všechny živé organismy mohou sloužit jako hostitelé dalším organismům. Hostitel – makroskopický organismus, parasit – mikroskopický.

3
Patogen – mikroorganismus vyvolávající onemocněnní
Pathogenita – schopnost vyvolávat onemocnění Patogen má schopnost průniku do hostitele a obsadit tam svou „niku“. Přitom často buď přímo nebo nepřímo poškodí buňky hostitele. Dovede prolomit ochranné bariéry hostitele a zaujmout specifická místa.

4
Pathogen je jakýkoli mikroorganismus, který přímou interakcí (infekcí) s jiným organismem, většinou vícebuněčným vyvolá onemocnění tohoto organismu. Pathogen tedy není jakýkoli mikroorganismus, který je náhodně spojen s onemocněním. Např. mikrob, který produkuje TOXIN, který vyvolá onemocnění v nepřítomnosti produkčního mikroorganismu nemůže být považován za pathogena.

5
Claviceps purpurea může být považován za pathogena skupiny obilovin, ale nemůže být označen jako pathogen pro člověka, i když požití ergotových alkaloidů,vyvolává u člověka ergotimus. Oportunistický pathogen je organismus, který je schopen samostatné existence, nebo který je normální součástí mikroflory člověka, ale může se za určitých podmínek stát patogenem. To nastává v případech, kdy normální obranný systém organismu je poškozen a je neúčinný.

6
Definice patogenity Jaké má patogen odlišné vlastnosti, které jej oddělují od komensálních typů? Některé základní determinanty patogenity jsou na mobilních genetických elementech – bakteriofágy a plasmidy.

7
Trojúhelník onemocnění: kompatibilní interakce mezi patogenem a hostitelem končí nemocí při vhodných vnějších podmínkách. Nejsou-li všechny podmínky splněny, nemoc nepropukne.

8
Virulemce Virulence je individuální vlastnost pathogena (např. bakterie nebo viru), která vyjadřuje stupeň pathogenity určitého mikrobiálního kmene ve srovnání s ostatními kmeny daného druhu. Také se dá říci, že jednotlivé kmeny jsou různě virulentní. Virulence se určuje například podle schopnosti mikroba vyvolat onemocnění či v rámci něho usmrtit hostitele. Kmeny, které mají tak nízkou virulenci, které téměř nezpůsobují onemocnění, ačkoliv daný druh patogenní je, se nazývají avirulentní (a daná vlastnost avirulence).
, která vyjadřuje stupeň pathogenity určitého mikrobiálního kmene ve srovnání s ostatními kmeny daného druhu. Také se dá říci, že jednotlivé kmeny jsou různě virulentní. Virulence se určuje například podle schopnosti mikroba vyvolat onemocnění či v rámci něho usmrtit hostitele. Kmeny, které mají tak nízkou virulenci, které téměř nezpůsobují onemocnění, ačkoliv daný druh patogenní je, se nazývají avirulentní (a daná vlastnost avirulence).")
9
Bakteriální virulence
Virulence – porovnatelný stupeň patogenity A. LD50 – míra virulence, počet mikroorganismů nezbytných k usmrcení 50% pokusných zvířat nebo buněk v kultuře. B. ID50 - míra virulence, počet mikroorganismů nezbytných k vyvolání infekce u 50% pokusných zvířat nebo buněk v kultuře. C. LD50 bakterie A u králíka je 1000 organismů, zatímco pro bakterii B 5000 organismů. Která bakterie je více virulentní?

11
Determinanty virulence bakterií
A. adheziny – proteiny bakterií, umožňují bakterii připevnění a kolonizaci hostitelských buněk 1. Pili (fimbrie)
")
12
Adheze Adheze Mnoho bakterií musí nejprve obsadit povrch buněk hostitele. Mnoho bakteriálních a hostitelských molekul, které jsou součástí mechanismu adheze již bylo identifikováno. Často, jsou bakteriální receptory buněk hostitele nezbytnými proteiny pro další funkce.

13
Kolonizace Některé virulentní bakterie produkují speciální proteiny, které jim umožňují kolonizaci částí těla hostitele. Helicobacter pylori je schopen přežít v kyselém prostředí lidského žaludku tvorbou enzymu ureasa. Kolonizace žaludku pak vede k vředům. Virulence různých kmenů Helicobacter pylori značně koreluje se stupněm produkce ureasy.

14
2. Mediátory aktivní invaze
1.invasiny - proteiny bakteriálního povrchu startující invazi do hostitelské buňky 2.indukovaná fagocytosa:L.monocytogenes produkuje internalin reagující s E-cadherinem hostitelské buňky 3.Invasin E. coli EPEC - Hp90 hostitelské buňky

15
invazivita Některé virulentní bakterie tvoří proteiny, které poškozují membrány hostitelských buněk nebo stimulují endocytosu do hostitelských buněk. Tyto virulenční faktory umožňují bakteriím vstup do hostitelských buněk a usnadňují tak vstup do těla přes povrchové vrstvy epithelu.

16
Virulence internalin Listeriolysin, lecitinasa Aktinová vlákna
Peptidoglykan-rezidua glykosaminu s volnými vázanými AMK - rezistence stěny na účinek lysozymu Navázání na povrch (INTERNALIN) Průnik do buňky - vyvolaná fagocytoza Navázání na molekulu cholesterolu - LISTERIOLIZIN O LECITINASA - lyze volna v cytoplasme eukaryotycke bunky. Mnozi a obaluje do malych vlaken aktinu. ( polymerizuji a vytvari " ocas komety ") dosahne cytoplasmaticke membrany, vyvola tvorbu podlouheho objektu, ktery je fagocytovan sousedici bunkou. Bakterie je tak uzavrena vdvojite membrane, ktera je lyzovana diky fosfolipase C. Listeria je znovu volna v cytoplasmaticke membrane nove bunky a muže se znovu množit. Faktory virulence jsou umístěny v lecithinasovém operonu a jejich transkripce ke regulována transkripčním aktivátorem PrfA.
 Průnik do buňky - vyvolaná fagocytoza. Navázání na molekulu cholesterolu - LISTERIOLIZIN O. LECITINASA - lyze. volna v cytoplasme eukaryotycke bunky. Mnozi a obaluje do malych vlaken aktinu. ( polymerizuji a vytvari ocas komety ) dosahne cytoplasmaticke membrany, vyvola tvorbu podlouheho objektu, ktery je fagocytovan sousedici bunkou. Bakterie je tak uzavrena vdvojite membrane, ktera je lyzovana diky fosfolipase C. Listeria je znovu volna v cytoplasmaticke membrane nove bunky a muže se znovu množit. Faktory virulence jsou umístěny v lecithinasovém operonu a jejich transkripce ke regulována transkripčním aktivátorem PrfA.")
17
3. Mediátory pasivní invaze
bakterie se váže na molekuly hostitele k usnadnění invaze fagocytů Legionella pneumophila a Mycobacterium spp. obě se vážou na komplement C3b a fragmenty C3bi - alveolární makrofágy

18
4. Bičíky lineární přívěsky delší nežli „pili“
a) pohyb a průnik do tkání b) umožňují chemotaxi Nejvíce glykosylované bakteriální proteiny jsou flageliny Campylobacter jejuni a Campylobacter coli. fla geny Flagelin z kmene C. jejuni 81–176 byl glykosylován na 19 serinových nebo threoninových zbytcích, a C. coli VC167 na přibližně 16 místech.
 pohyb a průnik do tkání. b) umožňují chemotaxi. Nejvíce glykosylované bakteriální proteiny jsou flageliny Campylobacter jejuni a Campylobacter coli. fla geny. Flagelin z kmene C. jejuni 81–176 byl glykosylován na 19 serinových nebo threoninových zbytcích, a C. coli VC167 na přibližně 16 místech.")
19
Typická patogenní buňka

20
5.Toxiny Mnoho faktorů virulence jsou proteiny bakterií, které otravují hostitelské buňky a působí poškození tkáně. Na př.mnoho toxinů tvoří bakterie přenášené potravinami. Některé pak přetrvávají v potravině i po tepelné úpravě a vyvolávají po konzumaci onemocnění.Některé bakteriální toxiny jsou však inaktivovány po záhřevu.

21
Typy toxinů Endotoxiny a exotoxiny.
Endotoxin je lipopolysacharid (LPS), který je součástí vnější membrány Gramnegativních bakterií. Lipid A složka LPS má toxické vlastnosti. LPS je velmi účinným antigenem a stimuluje účinnou imunitní odpověď hostitele. Jako část této odpovědi jsou uvolňovány cytokiny, které vyvolávají horečku a další symptomy onemocnění. Je-li přítomna vysoká hladina LPS nastává septický šok (nebo endotoxický šok),může vyvolat i smrt.
, který je součástí vnější membrány Gramnegativních bakterií. Lipid A složka LPS má toxické vlastnosti. LPS je velmi účinným antigenem a stimuluje účinnou imunitní odpověď hostitele. Jako část této odpovědi jsou uvolňovány cytokiny, které vyvolávají horečku a další symptomy onemocnění. Je-li přítomna vysoká hladina LPS nastává septický šok (nebo endotoxický šok),může vyvolat i smrt.")
22
5. Toxiny a) Endotoxiny - lipopolysacharidy G- - strukturální protektivní složky pouzdra - toxická aktivita Lipidu A b) Exotoxiny G- a G+ c) Enterotoxiny – Staphylococcus aureus
 Exotoxiny G- a G+ c) Enterotoxiny – Staphylococcus aureus.")
23
Haemophilus influenzae
6. Získání Fe 1. Siderofory - sekretované bakteriemi a) vysoká afinita k Fe b) komplex Fe-siderofor -internalizován c) některé bakterie užívají siderofory jiných druhů - hub 2. Některé bakterie vážou transferin a laktofe rin přímo: Neisseria meningitidis Haemophilus influenzae
 vysoká afinita k Fe. b) komplex Fe-siderofor -internalizován. c) některé bakterie užívají siderofory jiných druhů - hub. 2. Některé bakterie vážou transferin a laktofe- rin přímo: Neisseria meningitidis. Haemophilus influenzae.")
24
Získávání Fe V tělech teplokrevných živočichů (a lidí) je Fe vázané na transportní proteiny: transferin a laktoferin. Koncentrace volného Fe je zanedbatelná a k přežití patogena je nutné Fe získat. E. coli – siderofory – nízkomolekulární chelátory Fe, enterobaktin a aerobaktin. Yersinia yersiniabaktin, kódovaný HPI ( high pathogenity island) Nalezen u všech patogenních variant E. coli
 je Fe vázané na transportní proteiny: transferin a laktoferin. Koncentrace volného Fe je zanedbatelná a k přežití patogena je nutné Fe získat. E. coli – siderofory – nízkomolekulární chelátory Fe, enterobaktin a aerobaktin. Yersinia yersiniabaktin, kódovaný HPI ( high pathogenity island) Nalezen u všech patogenních variant E. coli.")
25
Modelové zobrazení způsobu získávání železa u Proteus mirabilis
Receptory na OM jsou červené Vstup hemu je zelený Údajný systém vstupu Fecitrátu je oranžový Proteiny účastnící se bio-syntézy sideroforů- modré. Fialový je kolicinový receptor. Cytoplasmatické Fe proteiny-bílé, Hnědé -removal

26
Receptory na OM jsou červené
Vstup hemu je zelený Údajný systém vstupu Fecitrátu je oranžový Proteiny účastnící se bio-syntézy sideroforů- modré. Fialový je kolicinový receptor. Cytoplasmatické Fe proteiny-bílé, Hnědé -removal

27
7. Inhibice imunitní odpovědi
Mnoho bakterií produkuje faktory virulence, které inhibují hostitelův imunitní obranný systém. Na př. běžnou bakteriální strategií je tvorba proteinů, které se vážou na protilátky hostitele. Polysacharidové pouzdro Streptococcus pneumoniae inhibuje fagocytosu této bakterie buňkami imunitního systému.

28
Úloha pouzder Pouzdra, tvořená polysacharidy umístěná na vnějších strukturách mnoha bakterií včetně Neisseria meningitidis (příčinným faktorem meningitis). Pouzdra mají významnou roli při úniku působení imunitního systému,neboť inhibují fagocytosu a ochraňují bakterie mimo hostitele.
. Pouzdra mají významnou roli při úniku působení imunitního systému,neboť inhibují fagocytosu a ochraňují bakterie mimo hostitele.")
29
8. Složky modulující imunitní odpověď
1. mnoho z nich je uvolňováno sekrecí III - po kon- taktu s cílovou buňkou 2. Destrukce fagocytosy - injikované proteiny lyzují fagocyt: Yersinia spp., Shigella spp., Salmonella spp., 3. Injikovaný protein znemožní fagocytosu – YopH Yersinia - defosforyluje proteiny fagocytu

30
Sekrece typu III

31
8. Složky chránící před imunitní odpovědí
1. Pouzdro - polysacharid, nebo komplex polysacharidu s proteiny na povrchu bakterie 2. Chrání před aktivací komplementu a fagocytosou 3. Preventivní tvorba C3bBb na povrchu bakterie zmenšuje opsonizaci vyvolanou C3b 4. Lipopolysacharid -LPS znemožňuje tvorbu „membrane attack complex“ MAC, délka řetězce O-antigenu je kritická

32
9. Složky mechanismu umožňující ochranu před protilátkami
1.Staphylococcus aureus - protein A a protein G Streptococcus pyogenes vážou Fe složku protilátky a znemožňují opsonizaci 2. Variace antigenu - proměnlivost povrchových složek :LPS, bičíky, pili, pouzdro, proteiny vnější membrány

33
10. Enzymy 1. IgA proteasa, štěpí IgA a) zabraňuje fagocytose působené Fc receptory protilátky b) nalezena u H. influenzae, A. meningitidis 2. Tkáň poškozující exotoxiny - penetrace 3. Ostatní exotoxiny
 nalezena u H. influenzae, A. meningitidis. 2. Tkáň poškozující exotoxiny - penetrace. 3. Ostatní exotoxiny.")
34
11. Geny determinant virulence jsou buď na chromosomu, plasmidu nebo fágu
Shigella a Salmonella spp. jak chromosom, tak plasmidy 2. Příklady fága: Corynebacterium diphtheriae - difterický toxin Vibrio cholerae - cholera toxin

35
Virulenční faktory Virulenční faktory bakterií jsou specifické proteiny nebo jiné molekuly, tvořené enzymovou činností. Tyto proteiny jsou kódované geny na chromosomální DNA, bakteriofágové DNA nebo na plasmidech. Bakterie užívají quorum sensing pro synchronizaci sekrece těchto molekul. Jsou bezprostředními příčinami úmrtnosti hostitele.

36
Campylobacter jejuni - OMV
Váčky vnější membrány – specifický sekreční systém Proteomika – 151 proteinů, některé z OM, ale také CDT (cytoletální distendní toxin) heterotrimerní toxiny tvořené proteobakteriemi, mají Dnasovou aktivitu. Tyto toxiny spouští zástavu buněčného cyklu v G2/M fázi u specifických buněčných linií, která vede ke zvětšeným nebo „oteklým“ buňkám, podle kterých byl toxin nazván. Postižené buňky odumírají apoptosou. Každý toxin sestává ze 3 rozdílných podjednotek, označených abecedně, v pořadí jak jsou kódující geny umístěny v cdt operonu. Cytoletální distendní toxiny náleží mezi AB toxiny. s aktivní ("A") podjednotkou, která přímo napadá DNA a vazebnou ("B") podjednotkou napomáhající toxinu připojení k cílové buňce. CdtB je aktivní podjednotka a homolog k savčí DNase1, CdtA a CdtC jsou vazebné podjednotky.
 heterotrimerní toxiny tvořené proteobakteriemi, mají Dnasovou aktivitu. Tyto toxiny spouští zástavu buněčného cyklu v G2/M fázi u specifických buněčných linií, která vede ke zvětšeným nebo „oteklým buňkám, podle kterých byl toxin nazván. Postižené buňky odumírají apoptosou. Každý toxin sestává ze 3 rozdílných podjednotek, označených abecedně, v pořadí jak jsou kódující geny umístěny v cdt operonu. Cytoletální distendní toxiny náleží mezi AB toxiny. s aktivní ( A ) podjednotkou, která přímo napadá DNA a vazebnou ( B ) podjednotkou napomáhající toxinu připojení k cílové buňce. CdtB je aktivní podjednotka a homolog k savčí DNase1, CdtA a CdtC jsou vazebné podjednotky.")
37
OMV – váčky vnější membrány
Genom Camp. Jejuni nemá sekreční systémy Jediný analog sekrece typu III jsou bičíky, zajišťují sekreci invazních antigenů. Velikost 10 – 500 nm průměr. Obsahují složky vnější membrány (proteiny, fosfolipidy, lipoologosacharidy, periplasmatické proteiny, a také β-laktamasy, alkalickou proteasu, hemolytickou fosfolipasu C.) Kromě CDT (CdtA, CdtB a CdtC), vazebné proteiny fibronektinu, glykosylační proteiny.
 Kromě CDT (CdtA, CdtB a CdtC), vazebné proteiny fibronektinu, glykosylační proteiny.")
38
DNA sondy založené na genech spojených s virulencí
Cílový gen sonda specifita α–listeriolysin přirozená L. monocytogenes syntetická L. monocytogenes β –listeriolysin přirozená L. monocytogenes Oddálená hyper přirozená L. monocytogenes sensitivita (DTH) L. ivanovii α–listeriolysin – SH aktivovaný hemolysin (60 kDa) Hly listeriolysin O β–listeriolysin – β-hemolysin, 5.2 kb DNA
 L. ivanovii. α–listeriolysin – SH aktivovaný hemolysin (60 kDa) Hly listeriolysin O. β–listeriolysin – β-hemolysin, 5.2 kb DNA.")
39
Problém nemocničních infekcí
V USA tyto infekce patří mezi 10 nejčastějších příčin smrti. Infekce E. coli v nemocnici – když epidemie zvládnuta do 1 týdne nepředstavuje riziko, když měsíc – zemře až 10% dětí. Salmonelosa – mimo nemocnici dnes není smrtelná, při nemocničních infekcích zemře až 1/7 nakažených. Klebsiela – 2× smrtelnější v nemocnici.

40
Válka a virulence Evakuační nemocnice ve Francii za 1. světové války – 360 lůžek pro pacienty s respiračními chorobami, za 3 měsíce tudy prošlo osob. Chřipka – 3 měsíce před a 3 po konci války – 10× větší úmrtnost, během 3 let se vrátila k normálu (0,1%). Celkem na španělskou chřipku zemřelo milionů lidí. Americká Občanská válka – první rok umírala na průjmová onemocnění 3% pacientů, nakonec 20%. (Úmrtnost na malárii byla po celou dobu stejná.) Z Napoleonovy armády 0,5 mil. se vrátilo z Ruska 130 tisíc. Teprve ve 2. světové válce padlo více vojáků v boji než zemřelo na nemoci.
. Celkem na španělskou chřipku zemřelo milionů lidí. Americká Občanská válka – první rok umírala na průjmová onemocnění 3% pacientů, nakonec 20%. (Úmrtnost na malárii byla po celou dobu stejná.) Z Napoleonovy armády 0,5 mil. se vrátilo z Ruska 130 tisíc. Teprve ve 2. světové válce padlo více vojáků v boji než zemřelo na nemoci.")
41
Genetika patogenity Geny podmiňující patogenitu jsou nápadně často součástí PAIS (Pathogenicity Islands) – až 200 kB, často odchylný obsah GC párů, často ohraničeny IS elementy. Přítomnost určitých genů naznačuje možnost horizontálního šíření. Vibrio cholerae – PAI kóduje pilus na který přisedá fág s choleratoxinem. S. aureus – PAI kóduje toxin toxického šoku, může být přenesen do jiné buňky. Geny s možností horizontálního přenosu mají tendenci poškozovat svého nositele, samy mohou včas opustit potápějící se loď.
 – až 200 kB, často odchylný obsah GC párů, často ohraničeny IS elementy. Přítomnost určitých genů naznačuje možnost horizontálního šíření. Vibrio cholerae – PAI kóduje pilus na který přisedá fág s choleratoxinem. S. aureus – PAI kóduje toxin toxického šoku, může být přenesen do jiné buňky. Geny s možností horizontálního přenosu mají tendenci poškozovat svého nositele, samy mohou včas opustit potápějící se loď.")
42
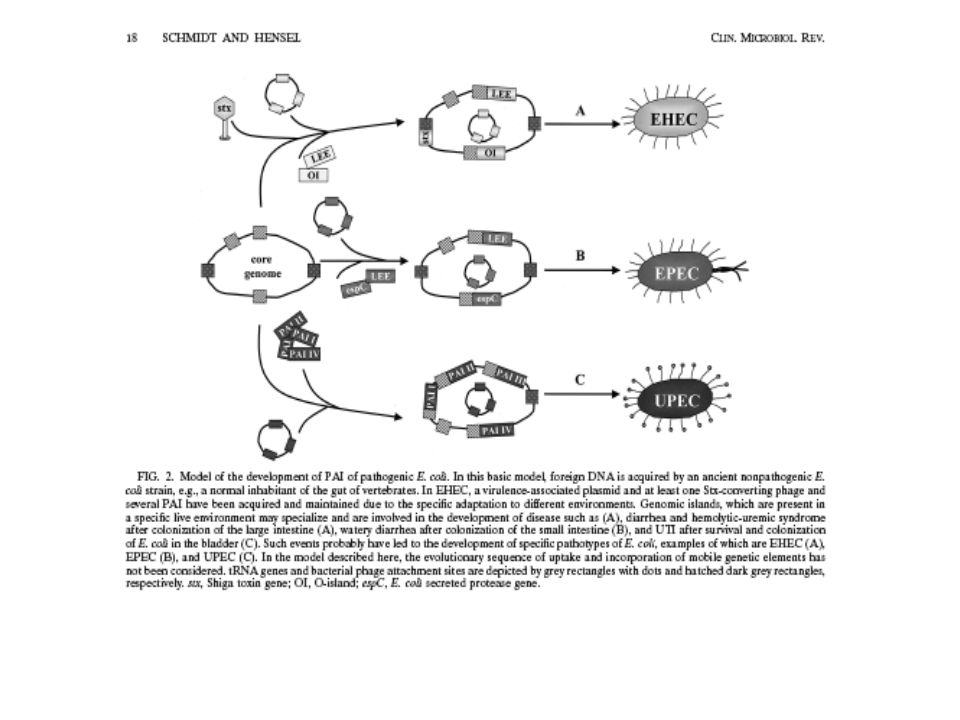
Ostrůvky pathogenity Úseky DNA, obklopené sekvencemi pro tRNA
%GC je relativně nízké při porovnání se zbytkem genomu Obsahují sekvence, které jsou unikátní pro daný organismus Jsou často značně velké (>20kb)
")
43
Pathogenicity islands (PAIs)
Jsou zvláštní třídou genomických ostrovů, které jsou přenášeny horizontálním transferem. Jsou včleněny do genomu pathogeních mikroorganismů , nejsou přítomny v nepathogenních organismech téhož nebo úzce příbuzného druhu.Mohou být také na plasmidech . Obvykle zabírají relativně velké úseky genomu od kb a kódují geny přispívající k virulenci pathogena. Typické vlastnosti jsou adherenční faktory, toxiny, systém na vstup Fe, invazní faktory a sekreční systémy. Ostrovy pathogenity jsou samostatné genetické jednotky obklopené repetitivními sekvencemi, inserčními sekvencemi nebo tRNA geny, které jsou místy pro rekombinace do DNA. Vyskytuje se též geny kryptické mobility, které ukazují původ transdukce. Analogní genomické struktury u rhizobií jsou označovány jako ostrovy symbiosy.

44
Vlastnosti PAI 1. PAIs kódují jeden nebo více faktorů virulence: adheziny, toxiny, invasiny apod.,. 2. Jejich obsah GC se často liší od zbytku genomu. 3. Jsou obklopeny přímými repeticemi: Sekvence bazí na obou koncích jsou totožné. 4. PAIs jsou spojeny s geny pro tRNA, která tak určují cílová místa k integraci do DNA 5. PAIs nesou i další funkční geny např. integrasa, transposasa, nebo části inserčních sekvencí. 6. Representují nestabilní oblasti DNA. Mohou se pohybovat od jednoho tRNA lokusu ke druhému, nebo být deletovány. 7. Site- specific integrace PAIs bývá zprostředkovaná integrasou a rekombinasou. 8. Jsou přenositelné jako samostatné jednotky do nových buněk a tak vytvářet virulenci u původně neškodných kmenů.

45
PAI Jeden druh bakterie může obsahovat více než jeden PAI. (např. Salmonella má minimálně 5). PAIs jsou umístěny převážně v Gram negativních buňkách ,ale není to pravidlo. Některé byly prokázány v Gram-pozitivních buňkách. Jsou u pathogenů, které mají přenesené geny plasmidy, fágy nebo konjugativním transposomem. Jejich přenos je horizontální přenos genů.

46
Velké inserty DNA- pathogenicity islands (Pais)
Tyto úseky DNA nejsou u nepatogenních mikroorganismů EPEC (enteropatogenní E.coli) 35 – 170 kb Celá řada genů virulence Některé geny nalézané na Pais – nápadná homologie Podobně geny na plasmidech virulence
 35 – 170 kb. Celá řada genů virulence. Některé geny nalézané na Pais – nápadná homologie. Podobně geny na plasmidech virulence.")
47
Sekreční systém typu III
Spouští se po kontaktu s hostitelskou buňkou Dopravuje účinné molekuly na povrch hostitelské buňky DNA složení Pais je odlišné od většiny genů na chromosomu Pais – jedinečné sekvence „přímá opakování“ a inserční sekvence IS

49
Šíření Pais – horizontální přenos
Bakteriální populace mají charakter klonů Většina klonů má odlišné kombinace genů Vstup Pais neznamená vznik nového druhu patogenu Varianty stejného druhu patogenu jsou časté – nadbytek serotypů Salmonella

50
Více o „pathogenity islands“
Pathogenicity islands (PAIs) jsou zřetelně odlišnou skupinou „genomových ostrůvků“, které jsou šířeny horizontálním přenosem. Jsou začleněny do genomu pathogenního mikroorganismu a obvykle chybí v nepatogenních příbuzných druzích téhož organismu. Jsou velké od 10 do 200 kB a kódují geny přispívající k virulenci daného pathogena. Typickými příklady jsou geny pro adherenci, toxiny, získávání Fe, invazní faktory a sekreční systémy.
 jsou zřetelně odlišnou skupinou „genomových ostrůvků , které jsou šířeny horizontálním přenosem. Jsou začleněny do genomu pathogenního mikroorganismu a obvykle chybí v nepatogenních příbuzných druzích téhož organismu. Jsou velké od 10 do 200 kB a kódují geny přispívající k virulenci daného pathogena. Typickými příklady jsou geny pro adherenci, toxiny, získávání Fe, invazní faktory a sekreční systémy.")
51
Ostrovy pathogenity - symbiosis island
Ostrovy pathogenity jsou samostatné genetické jednotky ohraničené přímými opakování sekvecí, inserčními sekvencemi nebo tRNA geny, které tvoří místa pro jejich rekombinaci s DNA. Geny pro kryptickou (skrytou) mobilitu mohou být také přítomny a ukazují tak na jejich původ a transdukci. Analogické genomové struktury u rhizobií jsou označovány jako ostrovy symbiosy - symbiosis island
 mobilitu mohou být také přítomny a ukazují tak na jejich původ a transdukci. Analogické genomové struktury u rhizobií jsou označovány jako ostrovy symbiosy - symbiosis island.")
52
Charakteristiky některých Pais u enterobakterií
Uropatogenní PaiI není v normálním α-hemolysin I >170 kb E.coli E.coli pap adhezní fimbrie IS PaiII není v normálním α-hemolysin II kb E.coli prs adhezní fimbrie cytotoxický nekrotický fak.I homolog OmpR Enteropatogenní locus of en- žádné IS tvoří AE lese kb E.coli EPEC terocyte efface sekrece III typu ment LEE S.Typhimurium Salmonella žádné IS sekrece typu III kb pathogenicity invaze do buněk epitelu island SP 1

53
Genomová mapa E.coli O157 H7

54
Přežívání patogenů ve fagocytujících buňkách
Salmonella, Mycobacterium spp.,Legionella pneumophila, Francisella tularensis, Listeria monocytogenes Salmonella typhimurium – do epitelu i makrofágů aktivací CDC-42 makropinocytosa U makrofágů indukuje apoptosu Geny pro vstup a apoptosu- na SpiI Uvnitř buňky jsou kyselém kompartmentu (pH 4,5)
")
55
Makrofág infikovaný S.tyhimurium značenou gfp v promotorové oblasti makrofágem indukovatelné chromosomální sekvence. Extracelulární mikroby jsou barveny protilátkou kojugovanou s fykoerytrinem (červené).
..")
56
Molekulární metody detekce
Identifikace mRNA transkriptů – izolace RNA z intracelulárně narostlých buněk, reversní transkripce –cDNA –templát pro PCR s primery typickými pro hledané geny PCR produkt je porovnán s produktem získaným z buněk standardně kultivovaných Diferenciální exprese genů - L. pneumophila uropatogenní E. coli

57
Bakteriální interakce
MAPK – mitogen activated protein kinase ECM – extracellular matrix – kolageny různých typů, fibronektiny, elastiny, adhezivní glykoproteiny Nové strategie pro prevenci a léčení- patogeny rezistentní ke konvenčním ATB. Antiadhezní terapie – po identifikaci adheze a invazních mechanismů – látky jako rozpustný bakteriální adhezin, nebo analogy receptoru hostitelské buňky, protilátky proti bakteriálním adhezinům ECM – vazebné bakteriální proteiny nebo adheziny – využití pro antiadhezní vakciny.

58
Regulace determinant virulence u St. aureus
Regulační faktory tvoří síť, specifických interakcí s cílovými promotory. Jedná se o dvousložkové regulační systémy: agr, saeAB, arlSR, a lytRS. Jsou citlivé na signály z prostředí a je to histidin kinasa (sensor) a odpovídající regulační protein. Násobné dráhy umožńují bakterii adaptaci na prostředí a tak rozvinout infekci.
 a odpovídající regulační protein. Násobné dráhy umožńují bakterii adaptaci na prostředí a tak rozvinout infekci.")
59
Schema regulace proteinu A u Staphylococcus aureus

60
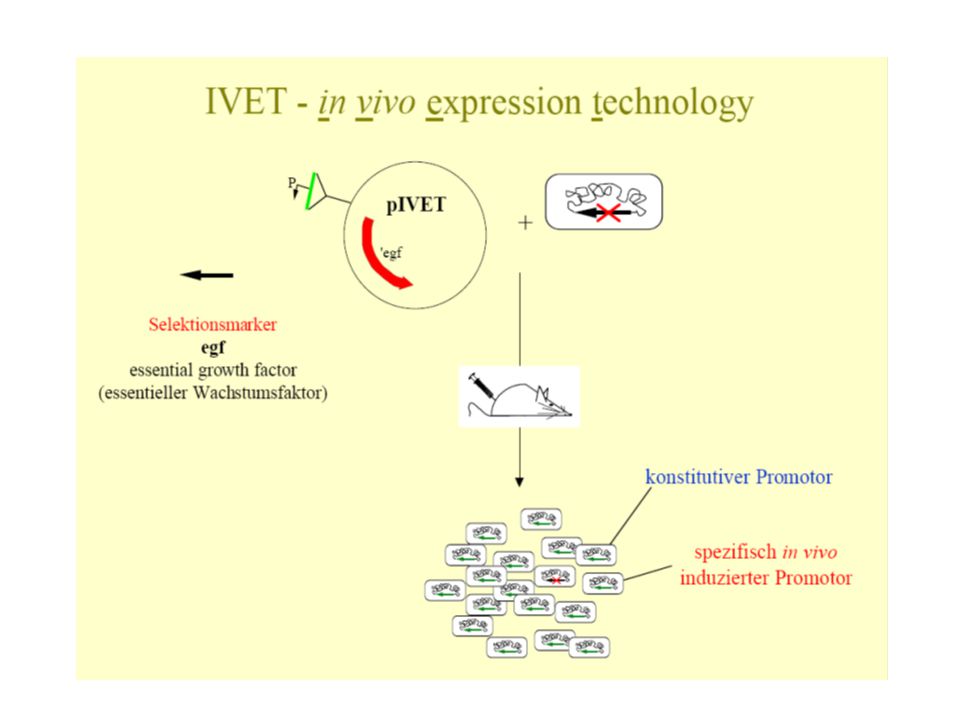
In vivo expression technology - IVET
Celá řada genů je exprimovaná teprve v hostiteli IVET slouží k identifikaci in vivo indukovaných genů – ivi Původní uspořádání (1993) používalo S. enterica Typhimurium, auxotrofní na purin (ΔpurA). Myši tento kmen bez komplementace rychle eliminují. Nahodilé fragmenty genomu „upstream“ naklonovány do bezpromotorového purA-lacZY syntetického operonu. Tato knihovna konjugovaná do Δpur salmonely, kde se homologní replikací tvoří merodiploidy. Buňky injikovány do peritoneální dutiny myši, spolu s defektním kmenem.
 používalo S. enterica Typhimurium, auxotrofní na purin (ΔpurA). Myši tento kmen bez komplementace rychle eliminují. Nahodilé fragmenty genomu „upstream naklonovány do bezpromotorového purA-lacZY syntetického operonu. Tato knihovna konjugovaná do Δpur salmonely, kde se homologní replikací tvoří merodiploidy. Buňky injikovány do peritoneální dutiny myši, spolu s defektním kmenem.")
62
Obrana proti pathogenům
Mimo zřejmý fakt, že virulentnější kmeny způsobí vážnější onemocnění, je důležitý také fakt, že z těch méně virulentních kmenů se vychází při přípravě živých očkovacích látek. Pro tento účel se uměle snižuje virulence určitých druhů a tento proces se nazývá atenuace. Atenuaci prováděl již Louis Pasteur (a to na viru vztekliny), později Calmette a Guérine (vakcína proti bakteriím Mycobacterium, původci TBC) či také Sabin (vakcína proti obrně).
, později Calmette a Guérine (vakcína proti bakteriím Mycobacterium, původci TBC) či také Sabin (vakcína proti obrně).")
Podobné prezentace















 Nespecifická : Fagocytóza granulocytů,monocytů.>")





