Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
John R. Helper & Alfred G. Gilman Zuzana Kauerová 2005/2006
G-PROTEINY John R. Helper & Alfred G. Gilman Zuzana Kauerová 2005/2006

2
OBSAH Buněčná signalizace G proteiny 2.1 Struktura a vlastnosti
2.2 Cholera toxin a pertussis toxin 2.3 Ukotvení v membráně 2.4 Typy G proteinových jednotek Modelové příklady 4.1 Transducin 4.2 Fosfatidylinositolová dráha 5. Buněčné odpovědi 6. Závěr

3
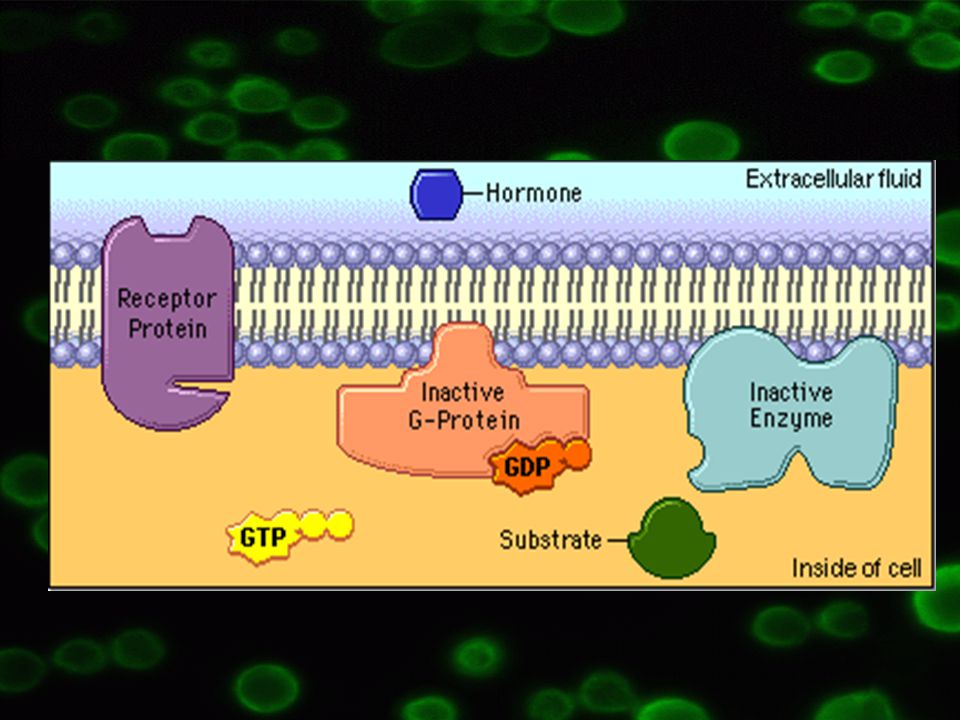
1. Buněčná signalizace Schopnost přijímat a zpracovávat vnější signály vlastní všem buňkám Přenos signálu: Primární messengery = hormony, neurotransmitery, růstové faktory => vazba na specifické receptory na povrchu buňky Výsledkem aktivace efektorového proteinu => mobilizace sekundárních messengerů => iniciace příslušné akce uvnitř buňky

4
2. G proteiny Ve všech eukaryotních buňkách
Heterotrimerní GTP-vazebné a hydrolyzující proteiny Nezbytné pro interakci povrchových receptorů s efektorovými proteiny na plazmatické membráně Nesou transdukující informaci

5
2.1 Struktura a vlastnosti
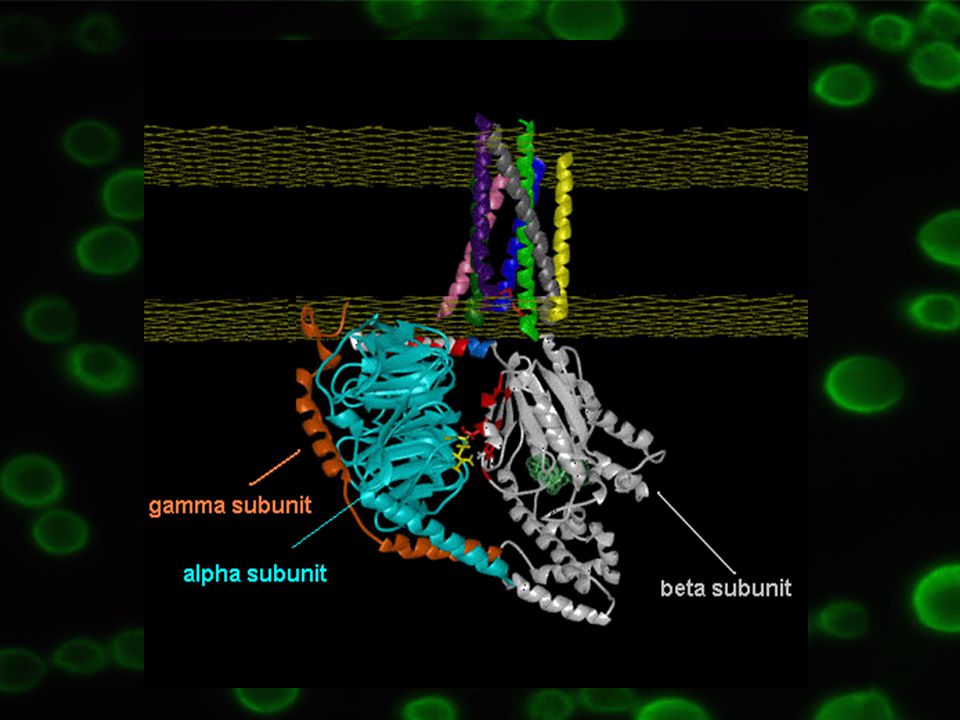
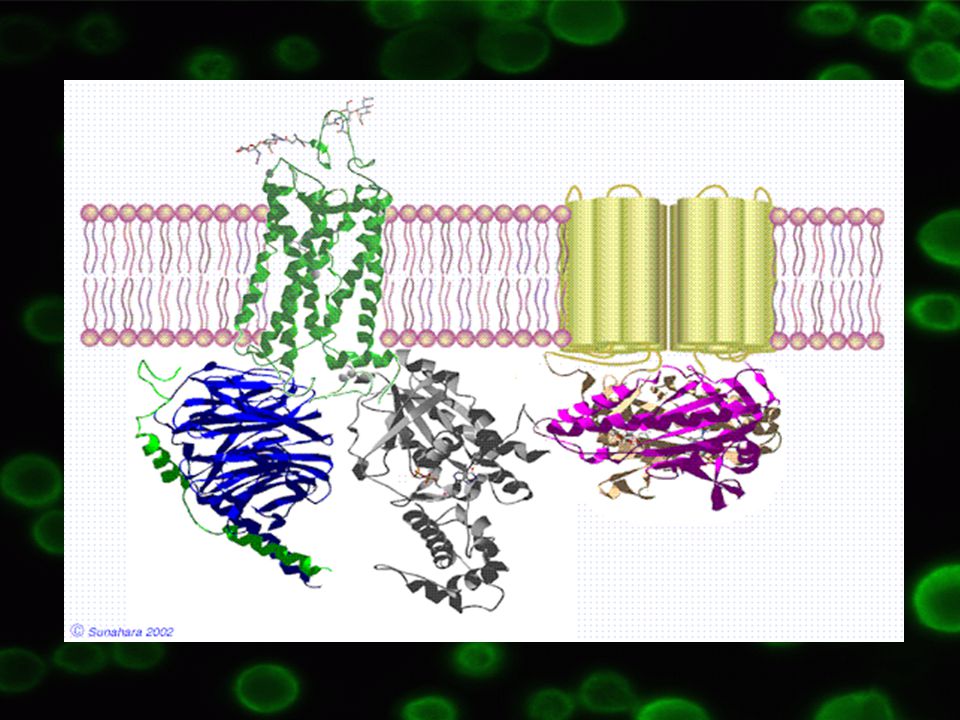
Heterotrimery, tři rozdílné podjednotky: a – 39 – 46 kDA, určuje typ oligomeru b – 37 kDa g – 8 kDa bg komplex těsně vázán k sobě, funguje jako jednotka

7
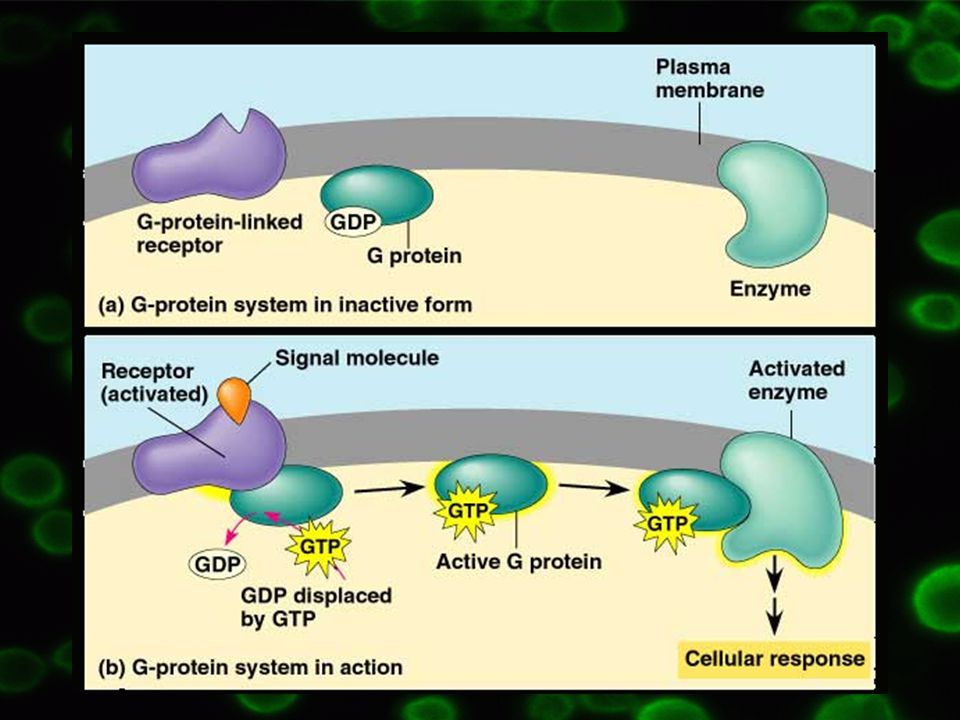
a podjednotka – vysoká afinita ke guanin nukleotidům – GDP/GTP
Vazba GDP na a podjednotku => neaktivní forma, bg komplex těsně navázán Vazba GTP na a podjednotku => aktivní forma, bg komplex oddisociuje => a podjednotka slouží jako efektorový protein Podobná funkce jako GTP = AlF4- společně s Mg2+, interagují s a podjednotkou => aktivace

9
Ukončení procesu odštěpením terminálního fosfátu v GTP => přeměna na GDP => inaktivace, vznik heterotrimeru abg

12
2.2 Cholera toxin a pertussis toxin
Některé a podjednotky mají specifické AMK zbytky, které mohou být kovalentně modifikovány bakteriálními toxiny Cholera toxin – katalyzuje transfer ADP-ribózy NAD na specifický Arg zbytek Pertussis toxin – katalyzuje transfer ADP ribózy NAD na specifický Cys zbytek na C-terminálním konci Následkem = zabránění aktivace G proteinů zprostředkované receptory

13
2.3 Ukotvení v membráně G proteiny ukotveny v plazmatické membráně prostřednictvím konce g podjednotky => myristoylové nebo palmitové zbytky

14
2.4 Typy G proteinových podjednotek

16
3. Modelové příklady GS proteiny – interakce s hormonálními a čichovými receptory => stimulace adenylát cyklázy => urychlení syntézy cAMP GOLF slouží ke spojení čichových receptorů se specifickou formou adenylát cyklázy Gsa reguluje nejméně dva iontové kanály stimulací napěťově řízených Ca2+kanálů v kosterních svalech a inhibicí Na+ kanálů v srdci

17
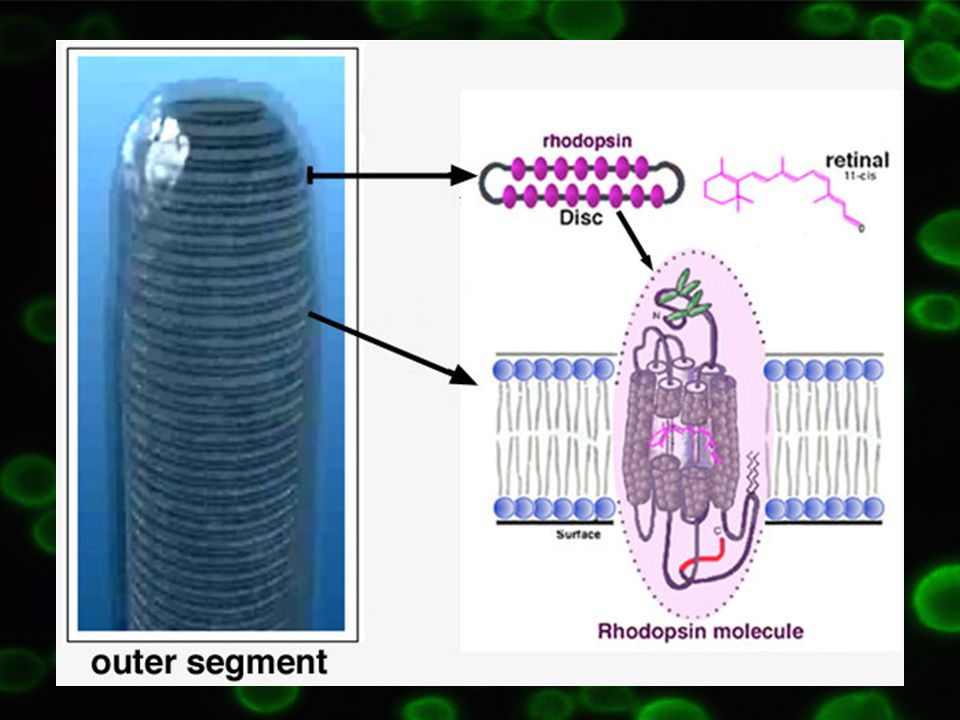
3.1 Transducin Nejvýznamnější model
Lokalizace ve fotoreceptorech na retinálních tyčinkách Základní funkce = vidění

19
Po dopadu fotonů o vhodné vlnové délce na rhodopsin => aktivace transducinu = Gt1 => stimulace cGMP fosfodiesterázy Cytoplazmatická koncentrace cGMP snížena

20
Princip: Fotony => retinal mění konfiguraci z cis na trans => aktivace rhodopsinu => transducin Gt fosforylován => další postup stejně jako u ostatních G proteinů

22
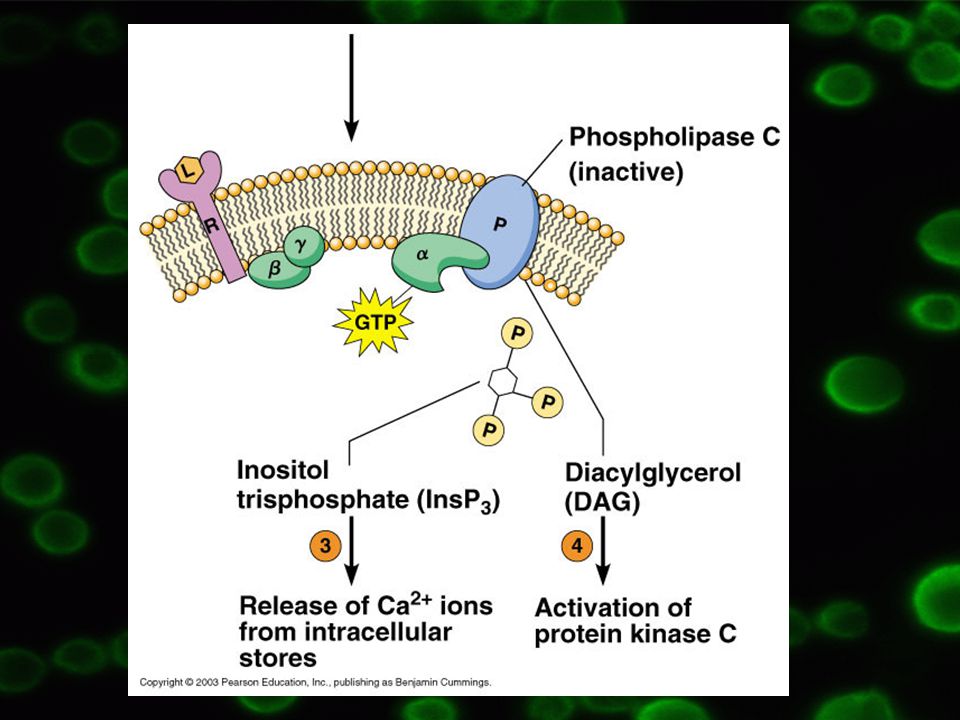
3.2 Fosfatidylinositolová dráha
PIP2 v poloze 4 a 5 fosforylován Součást membrán všech buněk Funkce prekurzoru druhých poslů Vzniká z PI

23
PIP2 může podléhat hydrolýze fosfolipázy C (typ b1)=> vzniká 1,4,5-PIP3 a diacylglycerol = signální funkce, druzí poslové Spřažen s G proteiny typu Q = Gq (izolovány z mozku skotu, krysích jater a erytrocytů krocanů)
")
25
4. Buněčné odpovědi Odpovědi G proteinu v některých orgánech podmiňovány interakcí efektorového proteinu s komplexem bg Různé izoformy adenylát cyklázy (AC), např.: Typ I AC inhibován přímo bg komplexem Typ II a IV aktivován bg komplexem za přítomnosti Gsa Regulátorem odpovědi nikoliv pouze a podjednotka, ale i komplex bg podjednotek Fáze výzkumu
, např.: Typ I AC inhibován přímo bg komplexem. Typ II a IV aktivován bg komplexem za přítomnosti Gsa. Regulátorem odpovědi nikoliv pouze a podjednotka, ale i komplex bg podjednotek. Fáze výzkumu.")
26
5. Závěr Heterotrimerní GTP-vazebné a hydrolyzující proteiny
Tři podjednotky: a a bg vázané do komplexu Funkce v celém organizmu – buněčná signalizace aktivací efektorového proteinu a zprostředkování buněčné odpovědi Nejvýznamnější modely fosfatidylinositolové dráhy a transducinu v očních tyčinkách

27
Děkuji Vám za pozornost

Podobné prezentace




















 vylučované kůrou nadledvinek (aldosteron, kortisol); 2) vylučované pohlavními žlázami (progesteron, testosteron, estradiol)>")
