Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Molekulární mechanismy smyslového vnímání František Duška Prezentace ke stažení na www.duska.eu

2
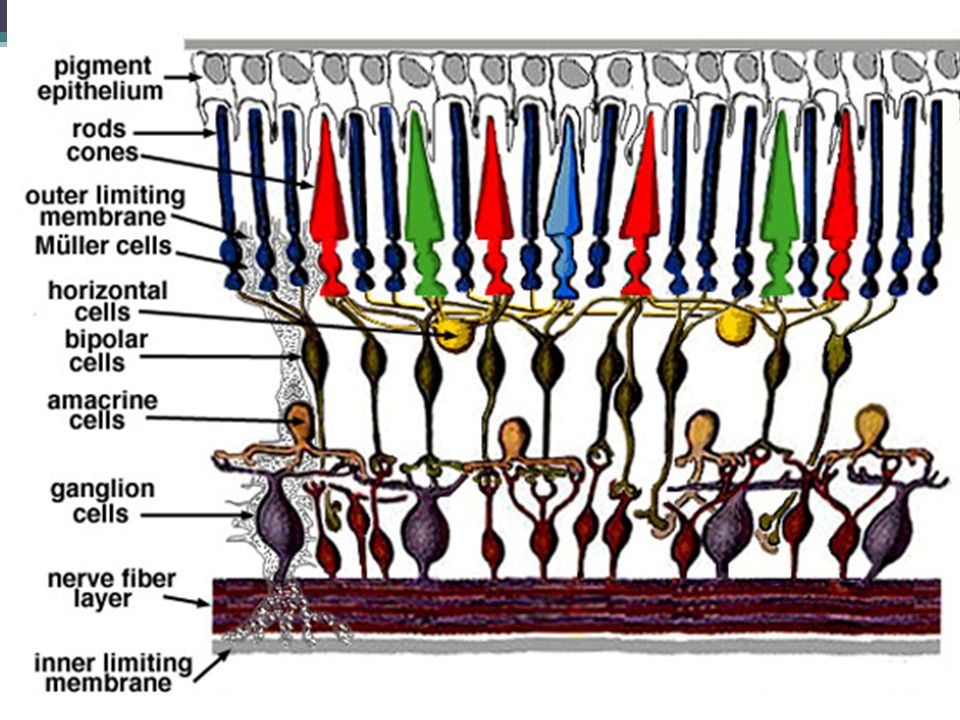
Úvod Doplnění molekulárních mechanismů vzniku akčních potenciálů na smyslových buňkách Smyslové buňky: ▫Primární = modifikované neurony (tyčinky, čípky, čichové buňky) ▫Sekundární = modifikované epitelové buňky ovíjené dendrity neuronů (chuťové bb., vláskové bb.) Akční potenciál je veden dalšími neurony smyslových drah až do mozkové kůry
 ▫Sekundární = modifikované epitelové buňky ovíjené dendrity neuronů (chuťové bb., vláskové bb.) Akční potenciál je veden dalšími neurony smyslových drah až do mozkové kůry")
3
Molekulární mechanismus čití světla Barevné vidění

7
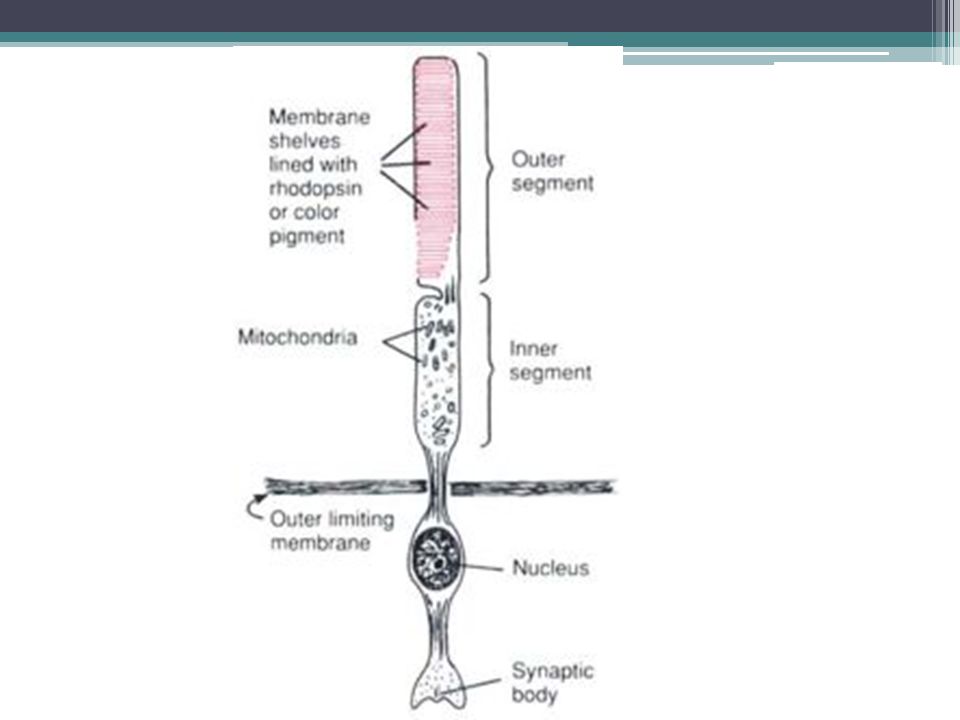
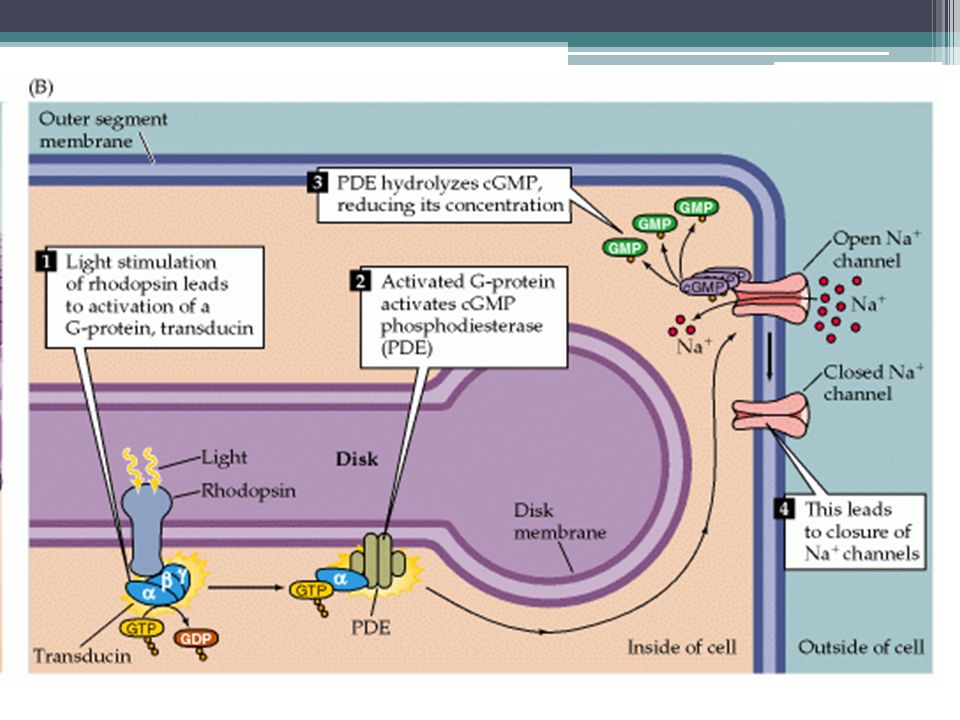
Mechanismus čití světla 1.Světločivný protein = rhodopsin ▫Retinal = kofaktor (odvozený od vit. A) ▫Opsin = apoprotein 2.Dopad fotonu vede k aktivaci rhodopsin, který přenáší signál na transducin 3.Transducin = G-protein, který aktivuje fofsfodiesterázu, která degraduje cGMP 4.Snížení IC koncentrace cGMP uzavře Na+ kanály v membráně, což vede k hyperpolarizaci
 ▫Opsin = apoprotein 2.Dopad fotonu vede k aktivaci rhodopsin, který přenáší signál na transducin 3.Transducin = G-protein, který aktivuje fofsfodiesterázu, která degraduje cGMP 4.Snížení IC koncentrace cGMP uzavře Na+ kanály v membráně, což vede k hyperpolarizaci.")
8
Detaily rhodopsinového cyklu All- trans- retinal All- trans- retinol Pig. epitel 11-cis- retinal Světlo Změna konformace opsinu, aktivace transducinu

9
Terminace fototransdukční kaskády Opsin-kináza fosroryluje rhodopsin, fosforylovaný rhodopsin váže arrestin Inaktivace transducinu spontánní hydrolýzou GTP na GDP+Pi Aktivace guanylátcyklasy (GCA proteinem) znovu na syntetizuje cGMP, což otvírá Na+ kanály
 znovu na syntetizuje cGMP, což otvírá Na+ kanály")
10
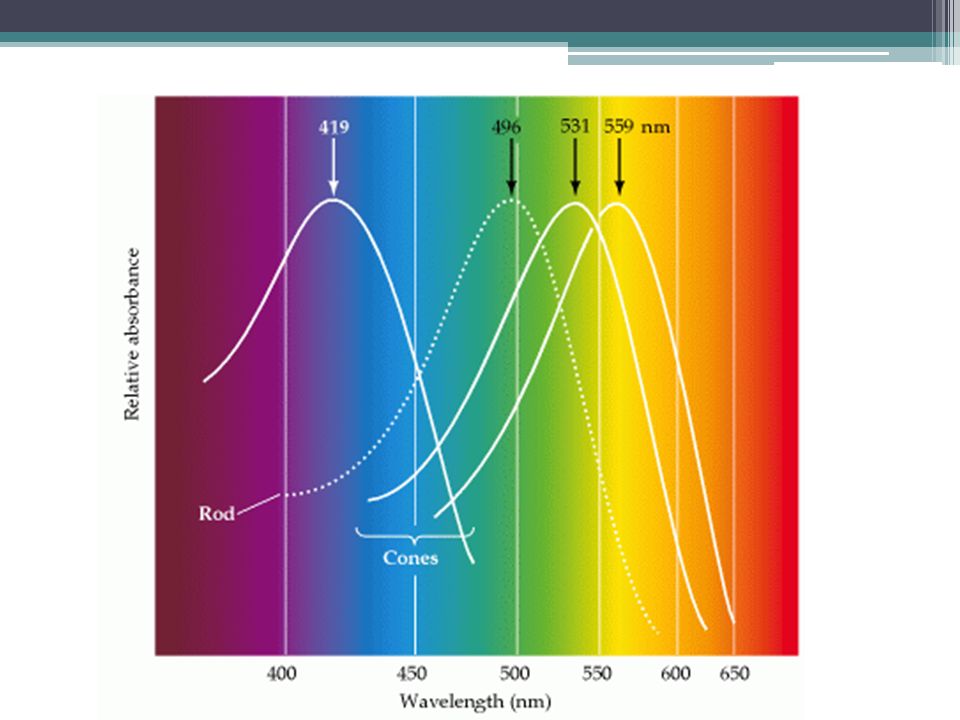
Barevné vidění Čípky Trichromatická teorie (Young-Helmholtz, 1872) 3 pigmenty citlivé na světlo o specifické vlnové délce X-vázaný defekt 1 proteinu = daltonismus (barvoslepost) ▫John Dalton, 1794
 3 pigmenty citlivé na světlo o specifické vlnové délce X-vázaný defekt 1 proteinu = daltonismus (barvoslepost) ▫John Dalton, 1794")
13
Molekulární mechanismy vnímání zvuku

14
Cortiho orgán

15
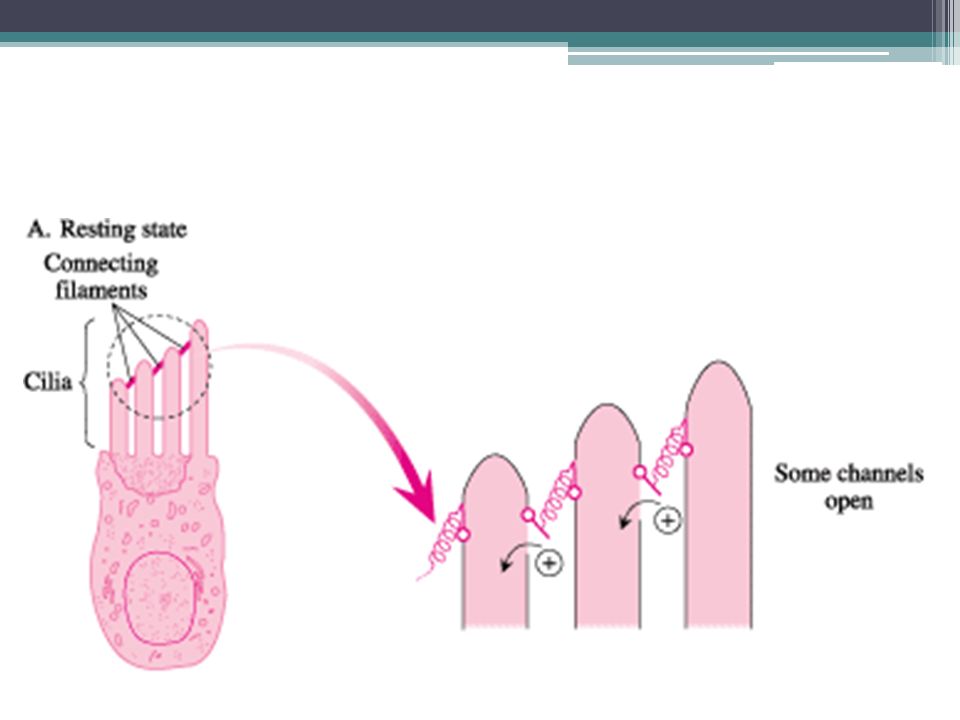
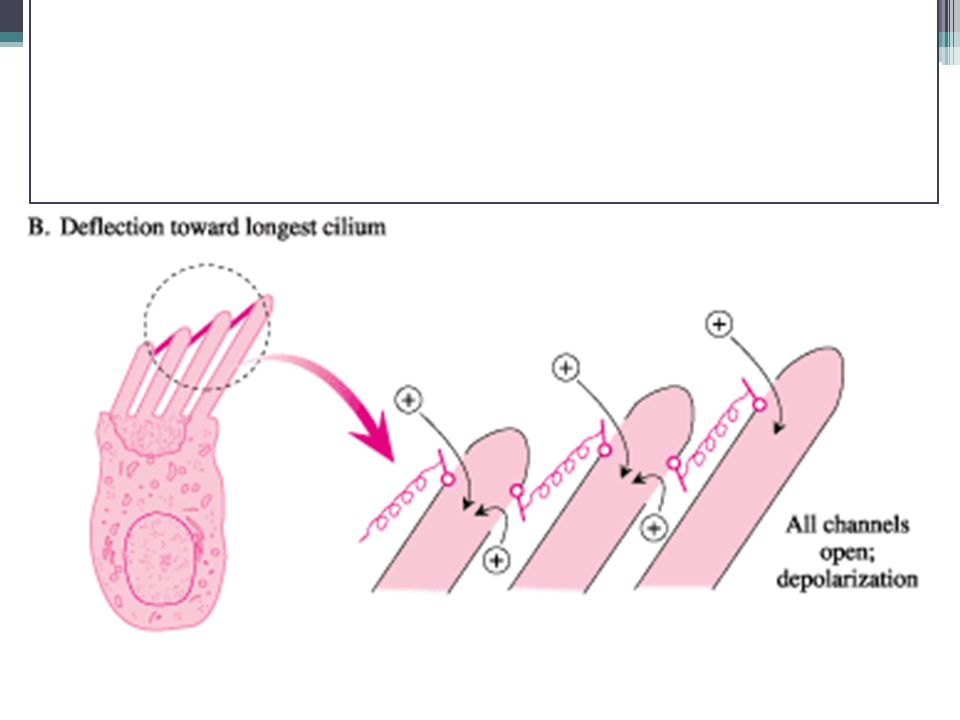
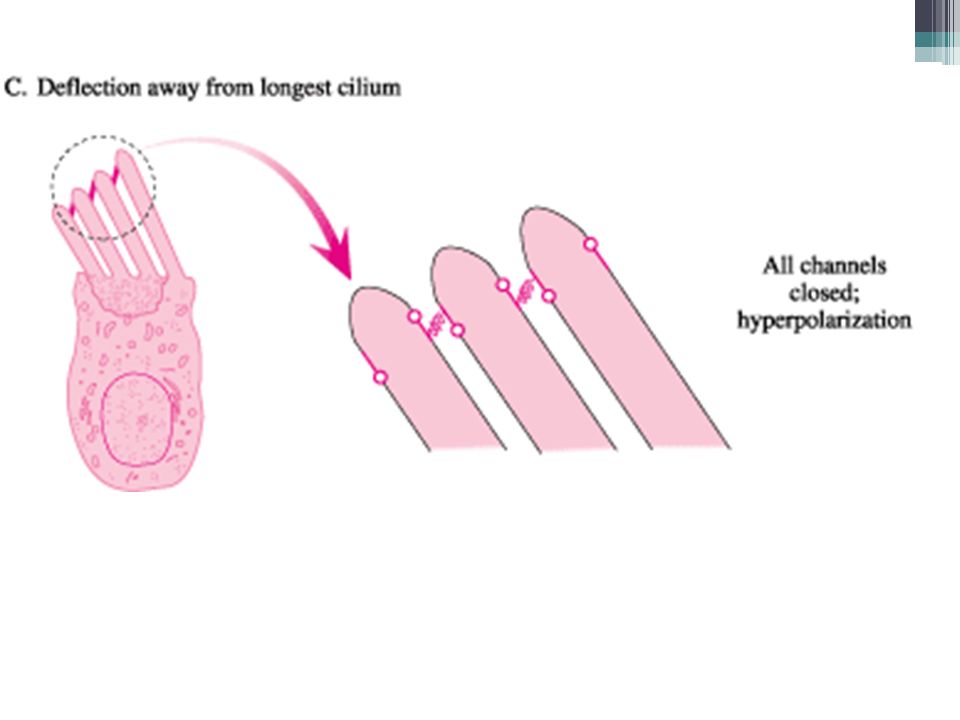
Percepce sluchu Vláskové buňky = v návaznosti na pohyb stereocilií o tektoriální membránu mění akční potenciál membrány To je vnímáno nervovými zakončeními 1. neuronu sluchové dráhy ▫Mechanismus komunikace vlásková b.-neuron není do detailů znám

19
Tok K+ iontů v Cortiho orgánu 1. EC v endolymfě (vysoká koncentrace K+) 2.Po elektrickém gradientu do vláskové b. (=depolarizace) 3.Z vláskové b. do perilymfy (nízká koncentrace K+) 4.Aktivní přečerpávání z perilymfy do endolymfy
 2.Po elektrickém gradientu do vláskové b. (=depolarizace) 3.Z vláskové b. do perilymfy (nízká koncentrace K+) 4.Aktivní přečerpávání z perilymfy do endolymfy.")
20
Chemické smysly (čich a chuť) umožňují rozpoznávat chemické složení okolí
 umožňují rozpoznávat chemické složení okolí")
22
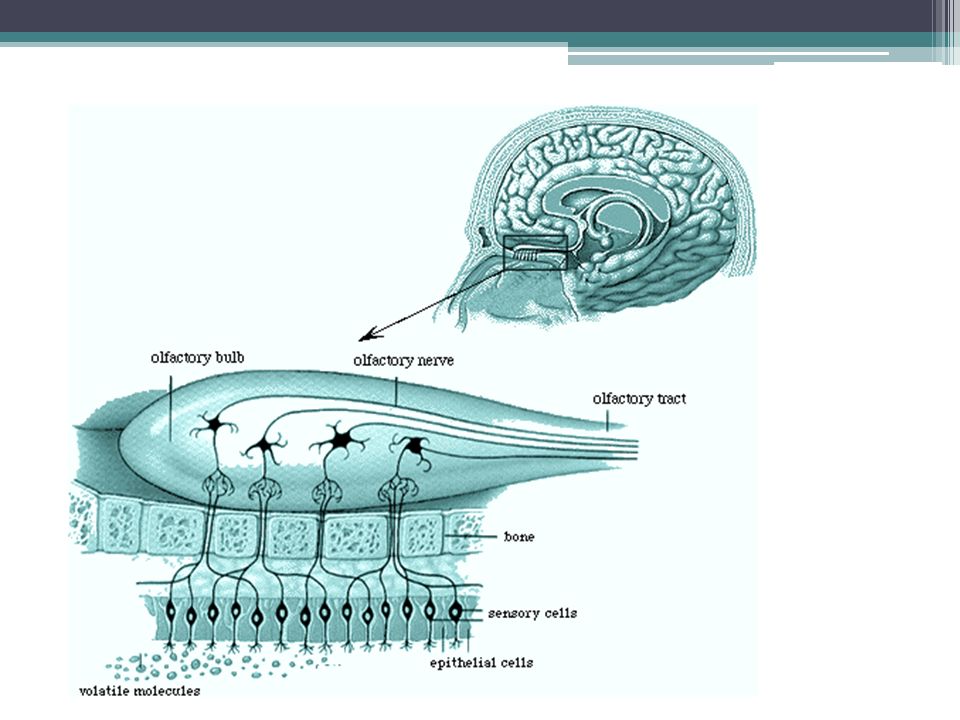
Vnímání čichu Chemické látky rozpuštěné ve vdechovaném vzduchu ▫Hydrofilní se rozpouštějí ve vrstvě hlenu ▫Lipofilní se váží na OBP (odorant binding protein) Vazba na receptor G-olf protein cAMP vstup Na+ a Ca2+ depolarizace a vznik AP ▫Nobelova cena 2004: Linda Buck, Richard Axel Čichová bb = modifikovaný neuron
 Vazba na receptor G-olf protein cAMP vstup Na+ a Ca2+ depolarizace a vznik AP ▫Nobelova cena 2004: Linda Buck, Richard Axel Čichová bb = modifikovaný neuron")
23
Vnímání čichu

24
V evoluci: hořká chuť = toxické substance Sladká + umami = výživné, prospěšné Kyselé, slané = doplňující chuťové modality

25
Chuť. pohárek Senzorické buňky pro 5 hlavních chutí: ▫Hořké ▫Sladké ▫Umami ▫Slané ▫Kyselé Bazální buňky (IV) Nervová zakončení zanořená do membrány senzorických buněk
 Nervová zakončení zanořená do membrány senzorických buněk.")
26
Komunikace Komunikací mezi chuť. rec. bb. a nervovým zakončením je přes purigenní receptory Ligand = ATP Finger et. Al., Science, 2005 Romanov et al. EMBO J, 2007

27
Distribuce čití chuti Tzv. chemotopická mapa čití chutí je mýtem Rozdíly v distribuci čití chuti existují, ale jsou nevýznamné

28
Evoluce chemotopické mapy chutí D.P.Hänig: Zur Psychophysik des Geschmacksinnes. Wundts philosophische Studien 17 (1901), pp. 576–623
, pp. 576–623.")
29
Zpracování chuťových vjemů

30
Hořké Evolučně důležitý smysl pro rozpoznání toxických substancí (např. rostlinné alkaloidy) Rodina T2R (25-30 genů) Podobné receptory v GIT, aktivace vede k nauzey a zvracení (spojené s PYY a CCK) Rosengurt, Am J Physiol, 2006
 Rodina T2R (25-30 genů) Podobné receptory v GIT, aktivace vede k nauzey a zvracení (spojené s PYY a CCK) Rosengurt, Am J Physiol,")
31
Sladké a chuť umami Receptory třídy T1R: ▫T1R1+T1R3 pro umami = chuť aminokyselin (glutamátu) ▫T1R2+T1R3 pro sladké G protein = gustducin… další kaskáda anlogická hořkému
 ▫T1R2+T1R3 pro sladké G protein = gustducin… další kaskáda anlogická hořkému")
32
Kyselé 1.Pokles pH = vzestup koncentrace H + ▫Nižší pokles pH je zapotřebí u slabých kyselin než u silných (citrát vs. HCl) 2.Otevření Ca2+ kanálů (PKD2L1 protein) 3.Depolarizace membrány na základě vstupu Ca2+ 4.Akční potenciál nervových zakončení Richter, J Appl Phys, 2003 Huang et al., Nature, 2006
 2.Otevření Ca2+ kanálů (PKD2L1 protein) 3.Depolarizace membrány na základě vstupu Ca2+ 4.Akční potenciál nervových zakončení Richter, J Appl Phys, 2003 Huang et al., Nature,")
33
„Acid-sensing“ proteiny v chuťových pohárcích Huang et al., Nature, 2006

34
Čítí kyselého: širší souvislosti Receptory pH krve a likvoru v mozkovém kmeni využívají identický mechanismus ▫Viz regulace minutové ventilace pomocí sensingu pCO2 v arteriální krvi: ▫CO2 + H2O = H2CO3 = H + +HCO3-

35
Huang et al., Nature, 2006

36
Centrální chemoreceptory pH Huang et al., Nature, 2006

37
Slané Slané = kationty ▫Na+ ▫Ostatní Influx vede k depolarizaci membrány Modifikováno aldosteronem („salt craving“) DeSimone, Am J Physiol 2006
 DeSimone, Am J Physiol 2006")
38
Chuť - shrnutí Chuťový pohárek obsahuje všech 5 typů bb. Pro čití chuťových modalit Chuťové buňky komunikují s nervovými zakončeními pomocí ATP, které se váže na pourigenní receptory Vznik receptorového potenciálu: ▫Hořké, sladké a umami: receptory s G-proteiny a depolarizace otevřením Ca2+ dep. Na+ kanálů ▫Kyselé a slané: depolarizace vstupem protonů, resp. kationtů

39
Nashledanou u souhrnné zkoušky a hodně štěstí…

Podobné prezentace


 o Mění přijatou.>")














>")


