Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Obezita a její patofyziologické souvislosti

2
Tuková tkáň Na tukovou tkáň se dnes pohlíží jako na aktivní endokrinní orgán a modulátor imunitních funkcí. U obezity nacházíme jak hypertrofii, tak hyperplasii adipocytů. Progresivní infiltrace tukové tkáně makrofágy Sekrece biologicky aktivních proteinů jak adipocyty, tak neadipocytárních buněčných populací v zukové tkáni. Endokrinní a parakrinní modulační funkce.

3
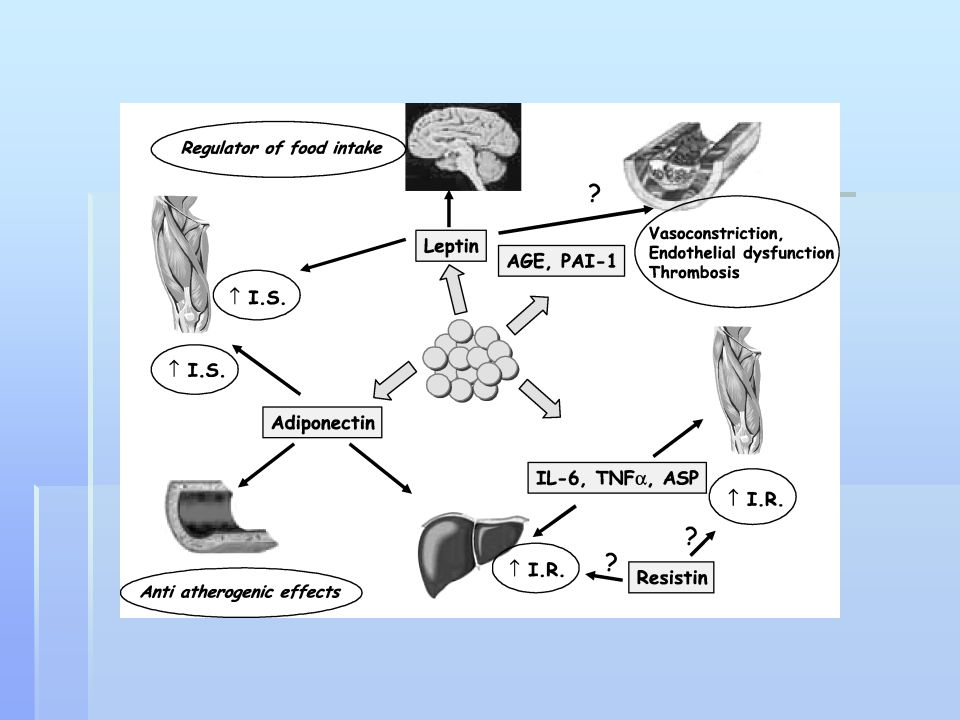
Bílá tuková tkáň Hraje podstatnou roli v regulaci homeostázy energie a glykémie pomocí adipokinů (leptin, adiponektin, TNFα a resistin. S výjimkou adiponektinun jsou sérové hladiny adipokinů u obezity zvýšeny. Adiponectin zvyšuje senzitivitu na inzulín zvýšením tkáňové tukové oxidace , což má za následek redukci cirkulujících hladin mastných kyselin a snížený intracelulární obsah triglyceridů v játrech a příčně pruhovaném svalstvu. Adiponectin inhibuje zánětlivé procesy snížením exprese adhesivních molekul v endoteliílních buňkách a produkce cytokinů v makrofázích. Snížení plasmatických hladin adiponektinu u prediabetických stavů předchází pokles senzitivity na inzulín.

5
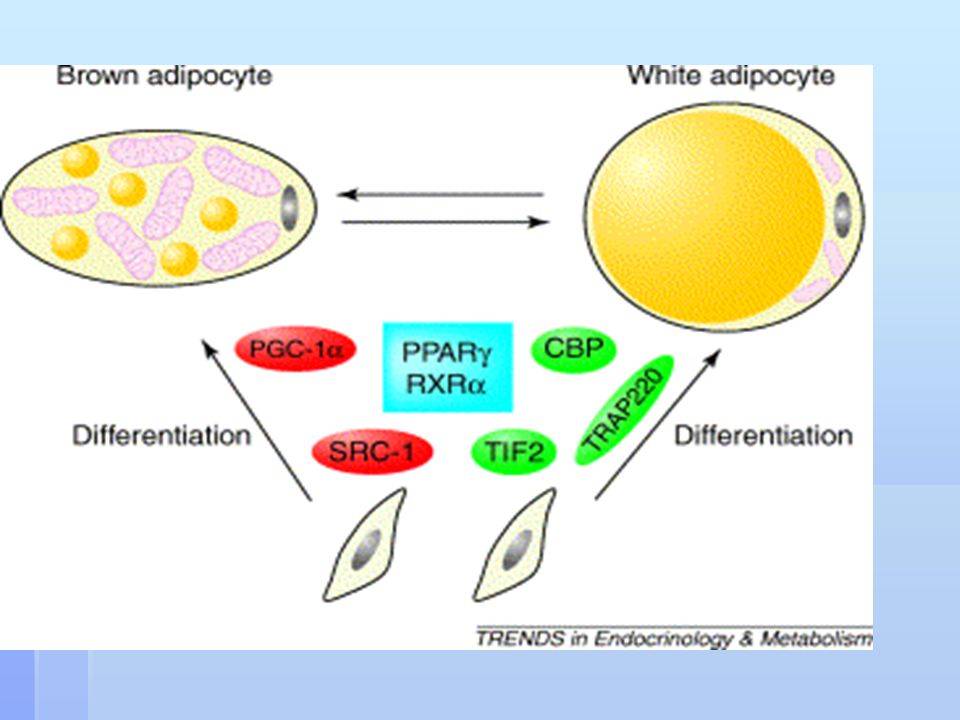
K předchozímu obrázku: role nukleárního receptoru PPAR (peroxisome proliferator-activated receptor ) a jeho koaktivátorů při diferenciaci adipocytů bílé a hnědé tukové tkáně. Preadipocyty podobné fibroblastům se diferencují do adipocytů hnědé nebo bílé tukové tkáně. PPAR vytváří heterodimery s RXR (retinoid X receptor ), které se účastní v diferenciaci obou typů adipocytů. Specifičnost této reakce se zvyšuje pomocí koaktivátorů PPAR. Pro diferenciaci buněk bílé tukové tkáně je potřeba TRAP220 (thyroid hormone receptor-associated protein 220). Koaktivátorem mnoha transkripčních reakcí včetně PPAR je cAMP responsive element binding protein (CREB). Účastní se regulace diferenciace a hypertrofie bílých adipocytů. Vstupuje do interakce s PGC1 (PPAR coactivator 1 ). PGC1 hraje rozhodující roli při vzniku atributů tukové tkáně, jako je vysoký počet mitochondrií a produkce uncoupling protein 1 (UCP-1). PGC1 může podpořit také konverzi bílých diferencovaných preadipocytů do na hnědé. Zvýšená exprese TIF2 (transcriptional intermediary factor 2) v bílém tuku podporuje akumulaci tuků. Zvýšený poměr TIF2 a SRC-1 (steroid receptor coactivator 1) redukuje termogenní kapacitu hnědé tukové tkáně.
, které se účastní v diferenciaci obou typů adipocytů. Specifičnost této reakce se zvyšuje pomocí koaktivátorů PPAR. Pro diferenciaci buněk bílé tukové tkáně je potřeba TRAP220 (thyroid hormone receptor-associated protein 220). Koaktivátorem mnoha transkripčních reakcí včetně PPAR je cAMP responsive element binding protein (CREB). Účastní se regulace diferenciace a hypertrofie bílých adipocytů. Vstupuje do interakce s PGC1 (PPAR coactivator 1 ). PGC1 hraje rozhodující roli při vzniku atributů tukové tkáně, jako je vysoký počet mitochondrií a produkce uncoupling protein 1 (UCP-1). PGC1 může podpořit také konverzi bílých diferencovaných preadipocytů do na hnědé. Zvýšená exprese TIF2 (transcriptional intermediary factor 2) v bílém tuku podporuje akumulaci tuků. Zvýšený poměr TIF2 a SRC-1 (steroid receptor coactivator 1) redukuje termogenní kapacitu hnědé tukové tkáně.")
7
Obezita Zvětšené tukové buňky Dyslipidémie: Vysoké hladiny FFA a VLDL
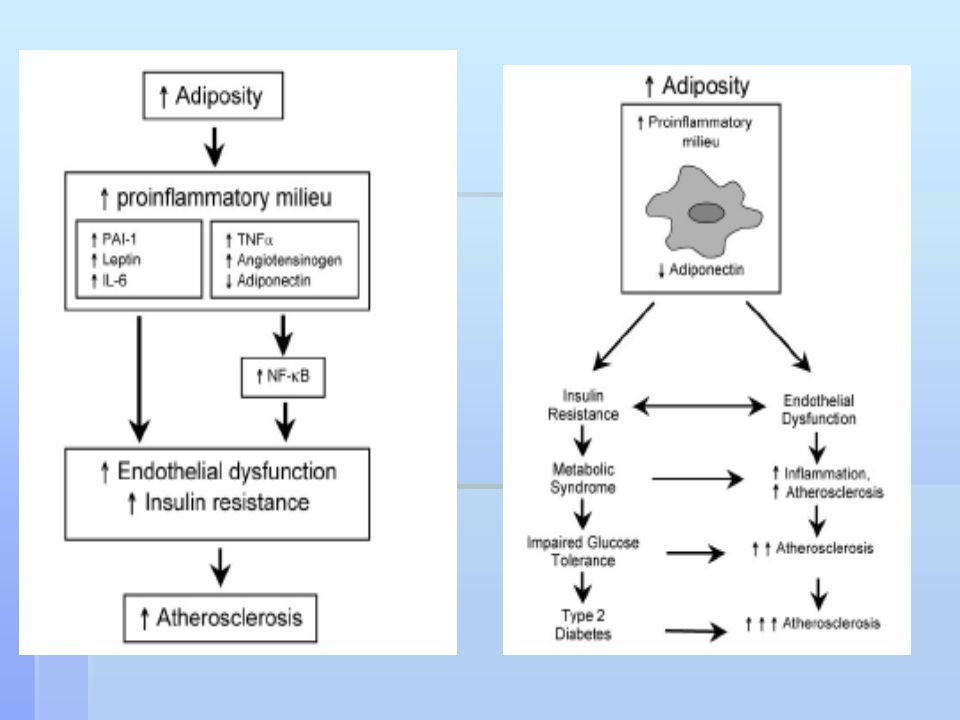
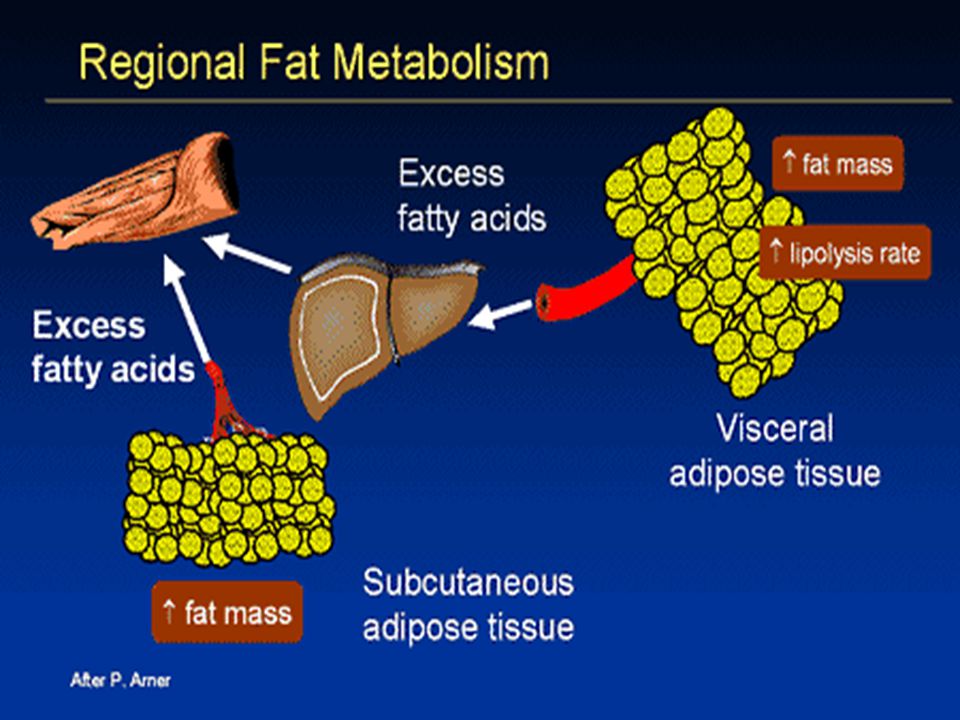
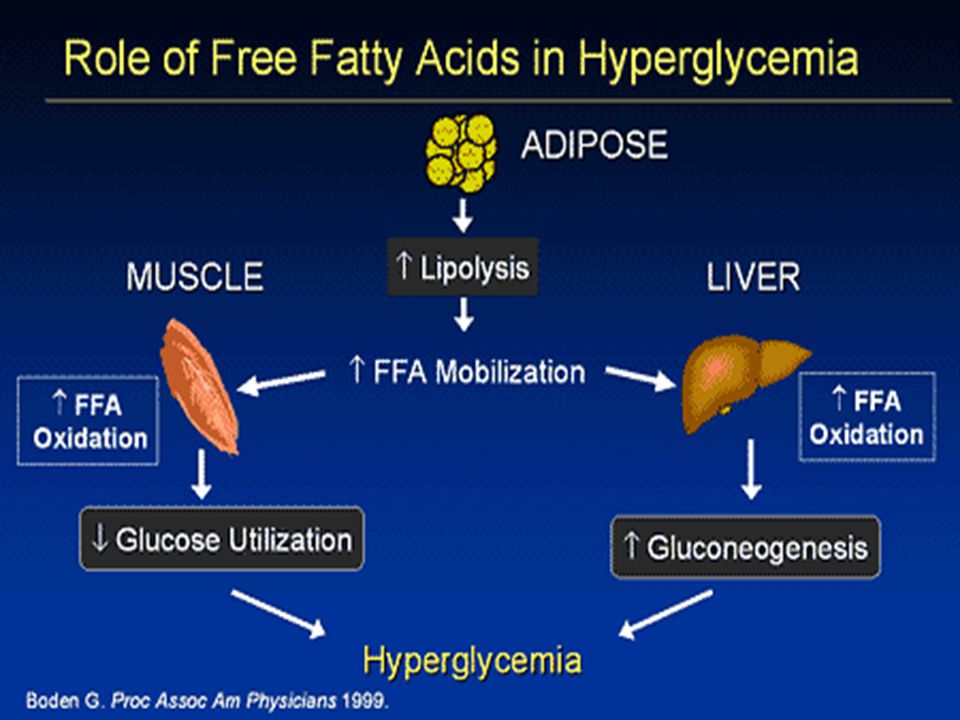
Ektopická depozice tuku v játrech (hepatická steatóza) a ve svalech (intramyocelulární lipidy Korelace mezi akumulací ektopického tuku a inzulínovou rezistencí. FFA mohou indukovat uvolnění prozánětlivých cytokinů (přes NFkapaB), což vede k další podpoře protředí typu „low grade inflammation“, které je indukováno tukovou tkání při obezitě. Saturované FFA působí prozánětlivě na adipocyty a makrofágy prostřednictvím aktivace Toll-like receptoru 4 (TLR4), mediátoru vrozené imunity. The TLR4/NFkapa B cesta ovlivňuje inzilínovou resistenci a vaskulární zánět indukovaný saturovanými FFA a obezitou indukovanou dietou u pokusných zvířat. Chronická aktivace NFkapa B v játrech, kosterním svalu i tukové tkáni je asociována s inzulínovou rezistencí v těchto tkáních. Kromě FFA uvolňuje tuková tkáň do krve četné bioaktivní molekuly (leptin, retinol-binding protein 4 (RBP-4), plasminogen activator inhibitor-1 (PAI-1) a prozánětlivé cytokiny a chemokiny. Adipokiny-interakce různých tkání a orgánů (mozek, játra, kosterní svaly, pankreas, endotel).
 a ve svalech (intramyocelulární lipidy. Korelace mezi akumulací ektopického tuku a inzulínovou rezistencí. FFA mohou indukovat uvolnění prozánětlivých cytokinů (přes NFkapaB), což vede k další podpoře protředí typu „low grade inflammation , které je indukováno tukovou tkání při obezitě. Saturované FFA působí prozánětlivě na adipocyty a makrofágy prostřednictvím aktivace Toll-like receptoru 4 (TLR4), mediátoru vrozené imunity. The TLR4/NFkapa B cesta ovlivňuje inzilínovou resistenci a vaskulární zánět indukovaný saturovanými FFA a obezitou indukovanou dietou u pokusných zvířat. Chronická aktivace NFkapa B v játrech, kosterním svalu i tukové tkáni je asociována s inzulínovou rezistencí v těchto tkáních. Kromě FFA uvolňuje tuková tkáň do krve četné bioaktivní molekuly (leptin, retinol-binding protein 4 (RBP-4), plasminogen activator inhibitor-1 (PAI-1) a prozánětlivé cytokiny a chemokiny. Adipokiny-interakce různých tkání a orgánů (mozek, játra, kosterní svaly, pankreas, endotel).")
8
Obezita Leptin a ciliární neurotrofní faktor (CNTF) centrálně i periferně řídí chuť k jídlu. CNTF suprimuje zánětlivou signalizační kaskádu asociovanou s kumulací lipidů v játrech a příčně pruhovaném svalstvu.

9
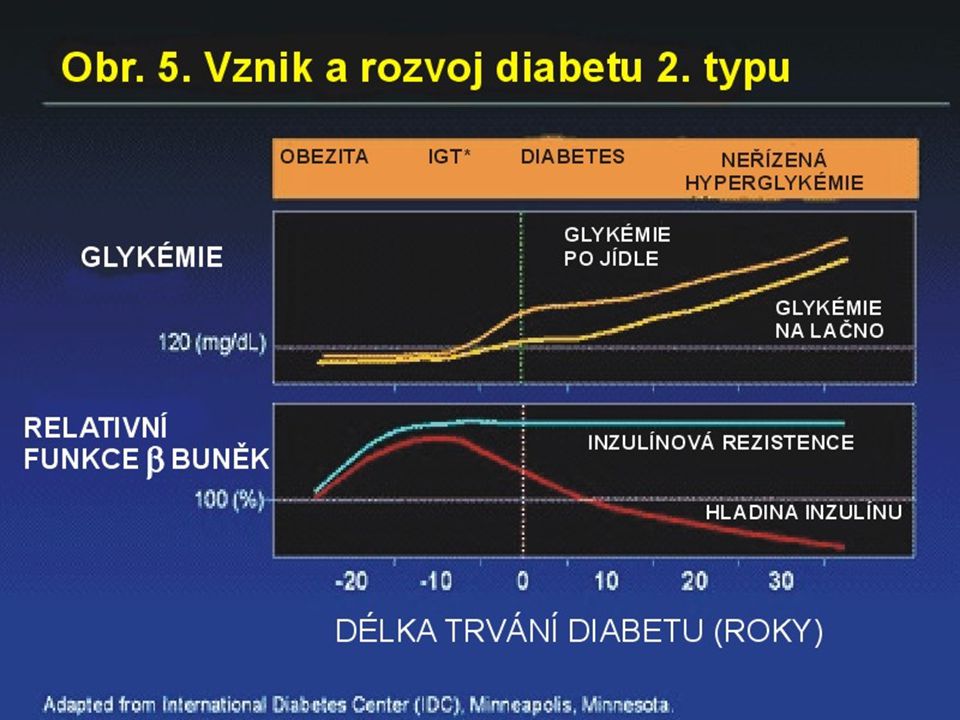
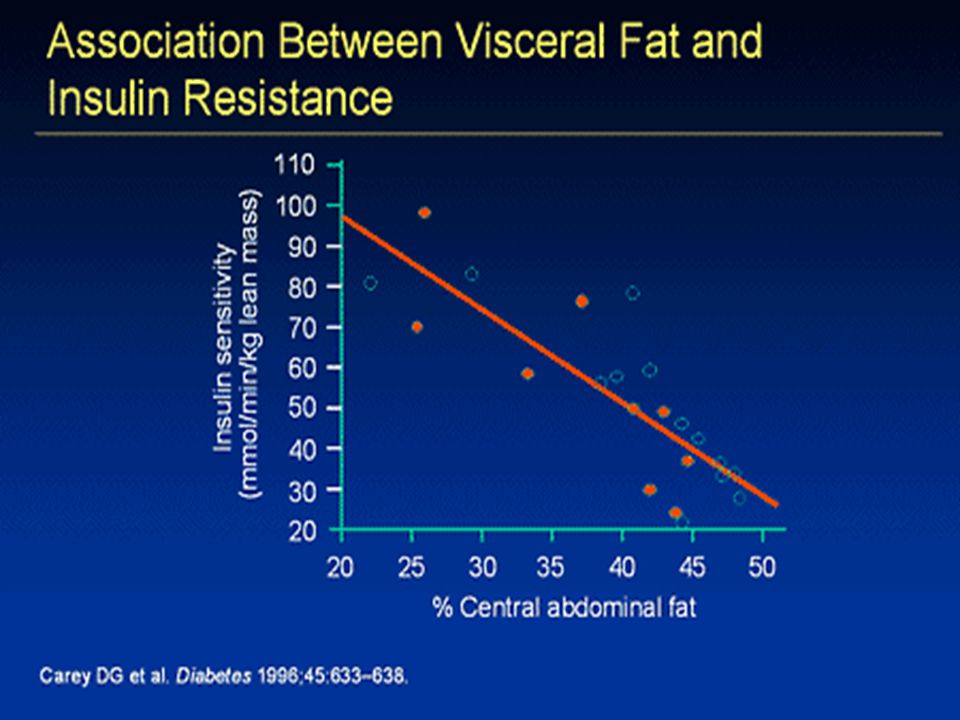
Obezita Nadbytek celkového a viscerálního tuku je asociován s inzulínovou rezistencí, diabetem a metabolickým syndromem. Nadbytek glukokortikoidů vede k centrální obezitě a diabetu. Hladiny systémových glukokortikoidů jsou u osob s typickým DM2T a u pacientů s idiopatickou obezitou normální. Ani koncentrace v portální krvi nejsou rozdílné. Glukokortikoidy mohou být produkovány lokálně prostřednictvím 11 hydroxysteroid dehydrogenázy typu 1 (11 HSD-1).
.")
10
Hypotéza „thrifty genotype“
Selektovány byly alely, které favorizovaly nárůst váhy a skladování tuku, protože byly výhodné v častých obdobích nutriční deprivace V podmínkách dostupnosti jídla a snížené potřeby fyzické aktivity vede taková predispozice k pandemii obezity v rozvinutých zemích Obezita: Zánět nízkého stupně Inzulínová rezistence Rizikové faktory pro četné komplexní nemoci: hypertenze, diabetes mellitus, typ II, syndrom polycystických ovárií, onemocnění GIT včetně rakoviny Geny účastnící se v regulacích nutričního stavu jako kandidátní geny pro obezitu

11
Energetická homeostáza

12
Signaling via LRb a gp130R: podobnosti a rozdíly

13
Metabolické cesty, ve kterých se účastní CNTF jako látka s antiobézními a inzulin senzitizujícími účinky

14
Ciliární neurotrofní faktor (CNTF)
Indukuje ztrátu hmotnosti a zlepšuje glukózovou toleranci (u člověka i hlodavců) CNTF půsoví centrálně indukcí hypothalamické neurogeneze, která má za následek modulaci příjmu potravy a periferně alterací exprese hetatálních genů podobným způsobem jako leptin. CNTF signalizuje přes komplex CNTFRα-IL-6R-gp130beta pro Zvýšení oxidace mastných kyselin redukci insulinové resistence v příčně pruhovaním svalstvu aktivací AMP-activované protein kinázy (AMPK), nezávisle na signalizaci z mozku. Antiobézní efekty CNTF v periferní tkáni jsou následkem přímých efektů na příčně pruhované svalstvo (Watt MJ et al., 2006).
 CNTF půsoví centrálně indukcí hypothalamické neurogeneze, která má za následek modulaci příjmu potravy a periferně alterací exprese hetatálních genů podobným způsobem jako leptin. CNTF signalizuje přes komplex CNTFRα-IL-6R-gp130beta pro. Zvýšení oxidace mastných kyselin. redukci insulinové resistence v příčně pruhovaním svalstvu aktivací AMP-activované protein kinázy (AMPK), nezávisle na signalizaci z mozku. Antiobézní efekty CNTF v periferní tkáni jsou následkem přímých efektů na příčně pruhované svalstvo (Watt MJ et al., 2006).")
15
Model regulace AMPK oxidace mastných kyselin leptinem v příčně pruhovaném svalstvu
Bjorbaek, C. et al. Recent Prog Horm Res 2004;59: Copyright ©2004 The Endocrine Society

16
Adipokiny Adipokines jsou proteiny syntetizované a sekretované v adipocitech. Adypocyty exprimují mnoho receptorů, např. cytokinové receptor (leptin, IL-6, TNF), nukleární receptory (androgenní, estrogenní, progesteronové, glucokortikoidní, pro tyreoidní hormony, PPAR a RXR receptory) i receptory na buněčné membráně (pro insulin, glukagon, růstový hormon, angiotensin I a II, adiponektin). Variabilitu těchto receptorů je potřeba vzít v úvahu, pokud mluvíme o predispozici k obezitě.
, nukleární receptory (androgenní, estrogenní, progesteronové, glucokortikoidní, pro tyreoidní hormony, PPAR a RXR receptory) i receptory na buněčné membráně (pro insulin, glukagon, růstový hormon, angiotensin I a II, adiponektin). Variabilitu těchto receptorů je potřeba vzít v úvahu, pokud mluvíme o predispozici k obezitě.")
17
Adipokiny specificky exprimované a sekretované tukovou tkání
Adipokiny specificky exprimované a sekretované tukovou tkání. Prozánětlivé cytokiny produkované a sekretované tukovou tkání (Cancello R et al., 2006)
")
19
Hypotetický model chronického zánětu a inzulínové rezistence adipocytů
Hypotetický model chronického zánětu a inzulínové rezistence adipocytů. Pokud obezita dosáhne určitého prahu, faktory tvořené v adipocytech indukují aktivaci makrofágů a inflitraci tukové tkáně těmito buňkami.

20
Adipocytokiny Metabolické účinky Adiponectin Leptin IL-6 PAI 1 Adipsin
Inhibition of monocyte adhesion to endothelial cells, macrophage transformation to foam cells, endothelial cell activation. Leptin Satiety signal, inhibits lipogenesis, stimulates lipolysis, improves insulin sensitivity, angiogenic activity. IL-6 Impairs appetite, lost fat tissue with no effect on lean mass, inhibits gluconeogensis, increases hepatic de novo synthesis of fatty acid and cholesterol. PAI 1 Inhibits activity of tissue-type plasminogen activator, an anticlotting factor. Adipsin Stimulates triglyceride storage in adipose cells through stimulation of glucose transport, enhances fatty-acid re-esterification and inhibits lipolysis TNF Stimulates release of FFA by adipocytes, reduces adiponectin synthesis and impaired insulin signalling. Resistin Controversial effects on glucose metabolism. Endothelial dysfunction? Angiotensinogen Acts through vasoactive peptide angiotensin II, correlates significantly with blood pressure. Aromatase Converts androstenedione to estrone driving fat to subcutaneous and breast tissues. 11-Hydroxysteroid dehydrogenase Regenerates metabolically active cortisol from cortisone in humans

22
Funkce leptinu

24
Dvě možná vysvětlení pro vztah mezi leptinem a aterosklerózou: (1) obezita je asociována s hyperleptinémií v důsledku zvětšeného množství tukové tkáně, což vede k podpoře proaterogenních efektrů leptinu, (2) obezita s hyperleptinémií vedou k leptinové rezistenci se sníženým pozitivním protiaterosklerotickým působením leptinu. PON1: paraoxonáza 1, SMC: buňky hladké svaloviny.

25
Regulátor TK Protein akutní fáze

26
Vztah mezi akumulací makrofágů v omentálním bílém tuku a zvýšenou histopatologií jater u obezity. Místní alterace stavu tkáně způsobená zvýšeným uvolňováním FFA a prozánětlivých cytokinů cestou portálního systému.

27
Změny v expresním profilu po několika týdnech diety s vysokou kalorickou
restrikcí

28
Interakce mezi neurotransmitery, NO, leptinem a příjmem potravy

29
Obezita a neuromodulace
Infekce, poranění a zánět jsou charakterizovány negativní energetickou bilancí s redukovaným příjmem potravy, ztrátou hmotnosti, zvýšenou termogenezou a horečkou. Serotonin, dopamin, NPY a leptin mohou ovlivnit syntézu a uvolňování prozánětlivých cytokinů svou schopností ovlivňovat uvolňování acetylcholinu v mozku. Acetylcholin možná působí v mozku nejenom jako neurotransmiter, ale také jako protizánětlivá molekula!!!

30
Obezita, zánět a neuromodulace
NO se v zánětu také účastní. Indukovatelný typ NO produkovaný makrofágy má prozánětlivé účinky (dále stimulované TNF-alpha, IL-1 a IL-6), zatímco endoteliální NO a neuronální NO působí protizánětlivě). Leptin může mít prozánětlivé účinky, protože zvyšuje koncentraci IL-1. Neurotransmitery (serotonin, dopamin a NPY) stimulují protizánětlivé děje modulací uvolňování acetylcholinu, který je schopen snižovat produkci prozánětlivých cytokinů. Úzká vazba existuje také mezi neurotransmitery, cytokiny, dysfunkcí hyperleptinemie způsobí excesivní produkci prozánětlivých pankreatu a inzulinem. Chybění nebo nedostatek inzulinu nebo jeho receptorů a cytokinů.
, zatímco endoteliální NO a neuronální NO působí protizánětlivě). Leptin může mít prozánětlivé účinky, protože zvyšuje koncentraci IL-1. Neurotransmitery (serotonin, dopamin a NPY) stimulují protizánětlivé děje modulací uvolňování acetylcholinu, který je schopen snižovat produkci prozánětlivých cytokinů. Úzká vazba existuje také mezi neurotransmitery, cytokiny, dysfunkcí hyperleptinemie způsobí excesivní produkci prozánětlivých pankreatu a inzulinem. Chybění nebo nedostatek inzulinu nebo jeho receptorů a cytokinů.")
31
Interakce mezi neurotransmitery, NO, leptinem,
inzulinem, zánětem a jejich vztah k obezitě

38
NFLD (non alcoholic fatty liver diseases)-nealkoholická steatóza jater
Častá u stavů spojených s inzulínovou rezistencí (obezita, DMII, dyslipidémie, metabolický syndrom. Prevalence v populaci 20%, v obézní populaci až 75%.

39
NAFLD Periferní inzulínová rezistence může přispívat ke steatóze sníženou antilipolytickou aktivitou insulínu. To vede ke zvýšené produkci FFA a jejich zvýšené distribuci do jater. Hyperinzulinémie vede také ke stimulaci lipogenezi de novo stimulací SREBP-1c. Příznivé efekty léků zvyšujících senzitivitu tkání k inzulínu (thiazolidinediony a metformin).
.")
40
Schéma účasti ChREBP a SREBP-1c v regulaci exprese glykolytických a lipogenních genů na podnět inzulínu a glukózy. Většina lipogenních genů (FAS, ACC) má karbohydrátový responzivní element (ChoRE) pro vazbu komplexu ChREBP–Mlx a sterol responzivní element (SRE) pro vazbu SREBP-1c. Aktivace obou míst vede v podmínkách hyperglykémie a hyperinzulinémie synergicky k indukci lipogenních genů.
 má karbohydrátový responzivní element (ChoRE) pro vazbu komplexu ChREBP–Mlx a sterol responzivní element (SRE) pro vazbu SREBP-1c. Aktivace obou míst vede v podmínkách hyperglykémie a hyperinzulinémie synergicky k indukci lipogenních genů..")
41
Aktivace ChREBP vyžaduje fosforylaci, která je determinována relativní aktivitou proteinové fosfatázy 2A (PP2A), regulované koncentrací X5P( xylulóza-5-fosfát). SREBP-1c je indukován inzulinem.

42
Glykolytické a lipogenní cesty v játrech
Glykolytické a lipogenní cesty v játrech. Hlavní funkcí glykolýzy v játrech je dodávat C z glukózy pro lipogenezu. Kromě jaterní glukokinázy, která je indukována exkluzivně inzulinem, všechny enzymy v tomto schématu jsou indukovány na transkripční úrovni hyperglykémií a hyperinzulinémií. GK: glucokináza; L-PK: jaterní pyruvát kináza, G6PDH: glukózo 6-fosfát dehydrogenáza; 6PGDH: 6-fosfoglukonát dehydrogenáza, ACC: acetyl CoA karboxyláza; FAS: syntetáza mastných kyselin.

43
Patofyziologie „intracelulárního Cushingova stavu“
Patofyziologie „intracelulárního Cushingova stavu“. Influx adipokinů, FFA a kortizolu z abdominální tukové tkáně via vena portae stejně jako hypersekrece inzulínu a LPs uvolněných ze střeva zvyšuje expresi 11β-HSD1, což vede ke zvýšení konverze kortizonu na kortizol v hepatocytech. Tento “intracelulární Cushingův stav” jater může alespoň zčásti vést k podcenění rizika akumulace kortizolu u metabolického syndromu s následným vznikem metabolickcýh nemocí , jako je snížená glukózová tolerance a hyperlipidémie v játrech.

44
NAFLD Alternativní hypotéza: akumulace tuku v játrech může být primárním stimulem pro rozvoj rezistence jater k inzulínu. Studie na lidech s DM II před a po dietě lehce hypokalorické s velmi nízkým obsahem tuků ukázaly až 81% redukci obsahu jaterního tuku. Tento efekt byl spojen se zlepšením jaterního metabolismu glukózy (bazálního i stimulovaného inzulínem), ale bez signifikantních změn ve vychytávání glukózy inzulín dependentními tkáněmi. Změny obsahu tuku ve stravě tedy mohou vést k alteraci obsahu tuků v játrech s alterací senzitivity jater k inzulínu, aniž dojde ke změně senzitivity periferních tkání nebo v hladinách FFA na lačno.
, ale bez signifikantních změn ve vychytávání glukózy inzulín dependentními tkáněmi. Změny obsahu tuku ve stravě tedy mohou vést k alteraci obsahu tuků v játrech s alterací senzitivity jater k inzulínu, aniž dojde ke změně senzitivity periferních tkání nebo v hladinách FFA na lačno.")
45
Perivaskulární adipocyty pracují jako integrovaný orgán v parakrinním a autokrinním režimu. Vysoké plasmatické koncentrace FFA způsobují jejich zvětšení a aktivaci JNK a NFkapaB, které indukují sekreci MCP-1 a tak povolávají monocyty do tkáně. TNF-alfa je sekretován různými buňkami cévní stěny (endotel, buňky hladké svaloviny, aktivované makrofágy) inhibuje aktivaci NO syntázy inzulínem a produkci NO v endoteliálních buňkách, což vede k podpoře vazokonstrikce. Následuje produkce adhezivních molekul jako P- a E-selektiny, VCAM-1 a intercelulární adhezivní molekuly 1 (ICAM-1), což zvyšuje vaskulární permeabilitu a podporuje atrakci cirkulujících prozánětlivých buněk do cévní stěny. TNF-alfa indukovanou produkci adhezivních molekul snižuje adiponektin. Adiponektin také indukuje nárůst koncetrace eNOS a podporuje aktivaci AMPK a inhibici NFkapaB.
 inhibuje aktivaci NO syntázy inzulínem a produkci NO v endoteliálních buňkách, což vede k podpoře vazokonstrikce. Následuje produkce adhezivních molekul jako P- a E-selektiny, VCAM-1 a intercelulární adhezivní molekuly 1 (ICAM-1), což zvyšuje vaskulární permeabilitu a podporuje atrakci cirkulujících prozánětlivých buněk do cévní stěny. TNF-alfa indukovanou produkci adhezivních molekul snižuje adiponektin. Adiponektin také indukuje nárůst koncetrace eNOS a podporuje aktivaci AMPK a inhibici NFkapaB..")
46
Děkuji vám za pozornost

Podobné prezentace










.>")








 vylučované kůrou nadledvinek (aldosteron, kortisol); 2) vylučované pohlavními žlázami (progesteron, testosteron, estradiol)>")
