Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Chromozomální základ dědičnosti
RNDr Z.Polívková Přednáška č.143 – Kurz: struktura buňky

2
Historie studia chromozomů:
r Sutton,Boveri - souvislost chromozomů s dědičností r Painter – počet chromozomů - chybný - 48 r Tjio, Levan – 46 chromozomů u člověka r Lejeune – první chromozomální abnormalita = trisomie chromozomu u pacienta s Downovým syndromem - Jacobs and Strong – karyotyp 47,XXY u mužů s Klinefelterovým syndromem - Ford et al. - monosomie X u žen s Turnerovým syndromem r Patau et al. – trisomie 13 u pacientů s Patauovým syndromem - Edwards et al. – trisomie 18 u pacientů s Edwardsovým syndromem r Steel, Breg - chrom. vyšetření z buněk plodové vody

3
Cytogenetika – studium chromozomů
Klinická cytogenetika – studium chromozomálních abnormalit

4
Ultrastruktura chromozomů:
Složení chromatinu: DNA histony = bazické bílkoviny: H1, H2A, H2B, H3, H4 nehistonové bílkoviny = neutrální n. slabě kyselé celková délka DNA asi 2 m lidské chromozomy obsahují cca strukturních genů = genů kodujících protein = malá část (cca1.5%) genomu
 genomu.")
5
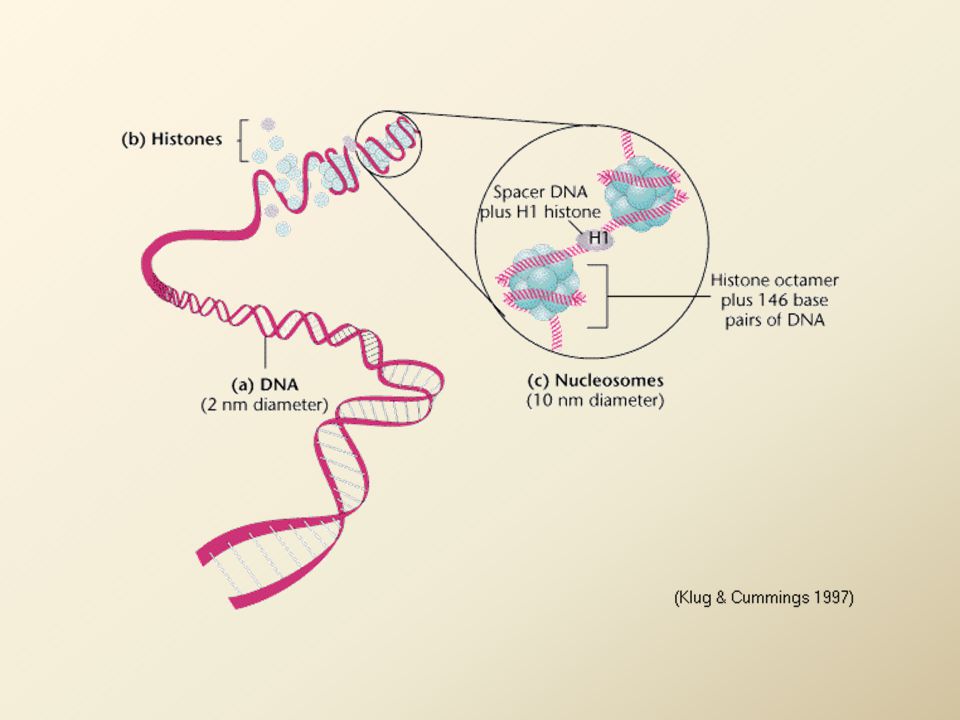
Organizace interfázního chromatinu
Nukleozom: jádro z oktameru histonů H2A, H2B, H3, H4 (liší se obsahem argininu a lyzinu) je obtočeno dvoušroubovicí DNA spojka mezi nukleozomy = vlákno DNA volné nebo asociované s H1 histony (asociace s H1 způsobí utažení vlákna) = kondenzace na 1/10 nativní DNA délky řetězec nukleozomů stočen do solenoidu (1 závit 6 nukleozomů) = základní jednotka chromatinového vlákna
 je obtočeno dvoušroubovicí DNA. spojka mezi nukleozomy = vlákno DNA volné nebo asociované s H1 histony (asociace s H1 způsobí utažení vlákna) = kondenzace na 1/10 nativní DNA délky. řetězec nukleozomů stočen do solenoidu. (1 závit 6 nukleozomů) = základní jednotka chromatinového vlákna.")
8
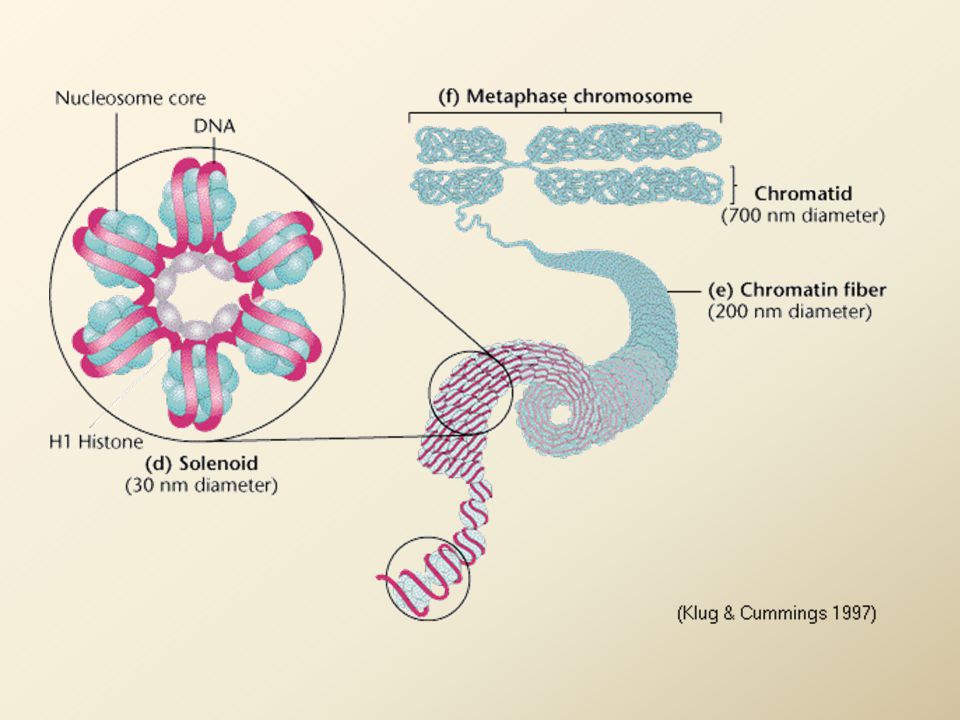
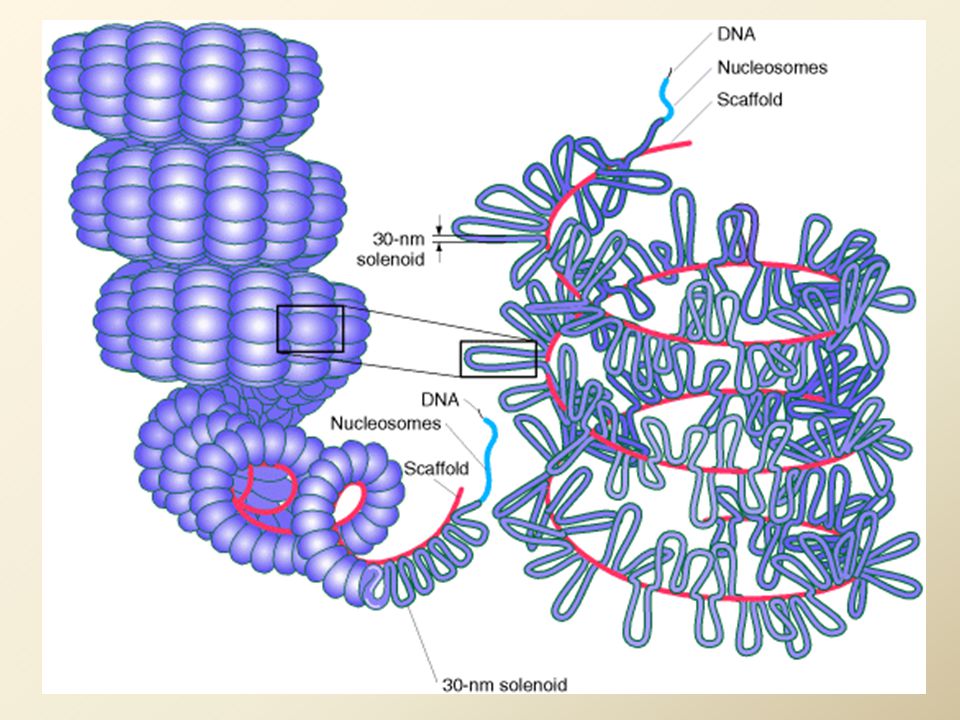
Kondenzace chromatinu do chromozomů
solenoid je skládán do smyček přichycených na kostru z nehistonových proteinů = chromozomy v profázi - zkrácení na 1/3000 délky Chromozomy v metafázi – struktura, tvořená nehistonovými proteiny se smyčkami DNA, je dále spiralizována do struktury chromatidy zkrácení DNA vlákna 1/10000 nativní délky

10
Morfologie lidských chromozomů
metacentrický submetacentrický akrocentrický centromera chromatidy telomera satelit sat. stopka (NOR) p q p = krátké rameno q = dlouhé rameno NOR = oblast organizátoru jadérka (rRNA geny)
 p. q. p = krátké rameno. q = dlouhé rameno. NOR = oblast organizátoru jadérka (rRNA geny)")
11
Centromery Centromera (primární konstrikce) = DNA + histony (větš.α-satelitní DNA = dlouhé tandemně se opakující sekvence) Kinetochor – proteinová struktura v centromeře - zprostředkuje napojení vřeténkových mikrotubulů a pohyb chromozomů v metafázi a v anafázi Narušení centromery → nondisjunkce (chyba v rozdělení chromozomů během dělení

12
Organizátor jadérka = „nucleolus organizer region „= NOR
Jadérko – v jádře, není obaleno membránou - místo transripce a úpravy ribozomálních RNA, místo spojování rRNA a proteinů do dvou ribozomálních podjednotek (v cytoplasmatickém ribozomu podjednotky spojeny) - mizí v průběhu mitozy, zpětně se tvoří v telofázi na specifických místech akrocentrických chromozomů (tj .satelitní stopky chromozomů 13,14,15,21,22) - jadérka fuzují ve větší celek – asociace akrocentrických chromozomů NOR obsahují tandemně se opakující genové klastry rRNA genů - variabilita v délce této oblasti (variabilní počet rRNA genů na každém akrocentrickém chromozomu, cca kopií)
 - mizí v průběhu mitozy, zpětně se tvoří v telofázi na specifických místech akrocentrických chromozomů (tj .satelitní stopky chromozomů 13,14,15,21,22) - jadérka fuzují ve větší celek – asociace akrocentrických chromozomů. NOR obsahují tandemně se opakující genové klastry rRNA genů. - variabilita v délce této oblasti (variabilní počet rRNA genů na každém akrocentrickém chromozomu, cca kopií)")
13
Telomery koncové oblasti chromozomů
opakované sekvence nukleotidů TTAGGG/CCCTAA (několik tisíc opakování) ochrana chromozomů před degradací a fuzemi, stabilizace konců chromozomů (telomery chráněny před účinkem exonuklaáz, které štěpí volné konce DNA) úloha při párování homologních chromozomů v meioze asociace telomer s jaderným obalem Jeho
 ochrana chromozomů před degradací a fuzemi, stabilizace konců chromozomů (telomery chráněny před účinkem exonuklaáz, které štěpí volné konce DNA) úloha při párování homologních chromozomů v meioze. asociace telomer s jaderným obalem. Jeho.")
14
Replikace telomer Enzym telomeráza zajišťuje replikaci telomer = ribonukleoproteinový komplex s funkcí reverzní transkriptázy – syntetizuje DNA dle templátu RNA) vysoká aktivita telomerázy v embryonálních a v nádorových buňkách, aktivita též v kmenových buňkách nízká n. nedetegovatelná aktivita v somatických buňkách zkracování telomerických sekvencí při každém dělení buňky = limitující faktor počtu dělení - souvislost s buněčným stárnutím (Hayflickův limit – buňka umírá po prodělání určitého počtu dělení – zkrácení telomer na neúnosnou míru) abnormální aktivita telomerázy prokázána v nádorových buňkách – nádorové buňky „nesmrtelné“
 vysoká aktivita telomerázy v embryonálních a v nádorových buňkách, aktivita též v kmenových buňkách. nízká n. nedetegovatelná aktivita v somatických buňkách. zkracování telomerických sekvencí při každém dělení buňky = limitující faktor počtu dělení - souvislost s buněčným stárnutím (Hayflickův limit – buňka umírá po prodělání určitého počtu dělení – zkrácení telomer na neúnosnou míru) abnormální aktivita telomerázy prokázána v nádorových buňkách – nádorové buňky „nesmrtelné")
15
Detekce telomerických sekvencí metodou FISH (= fluorescenční in situ hybridizace - telomerické sondy) Vpravo nahoře - myší chromozomy (telocentrické)
")
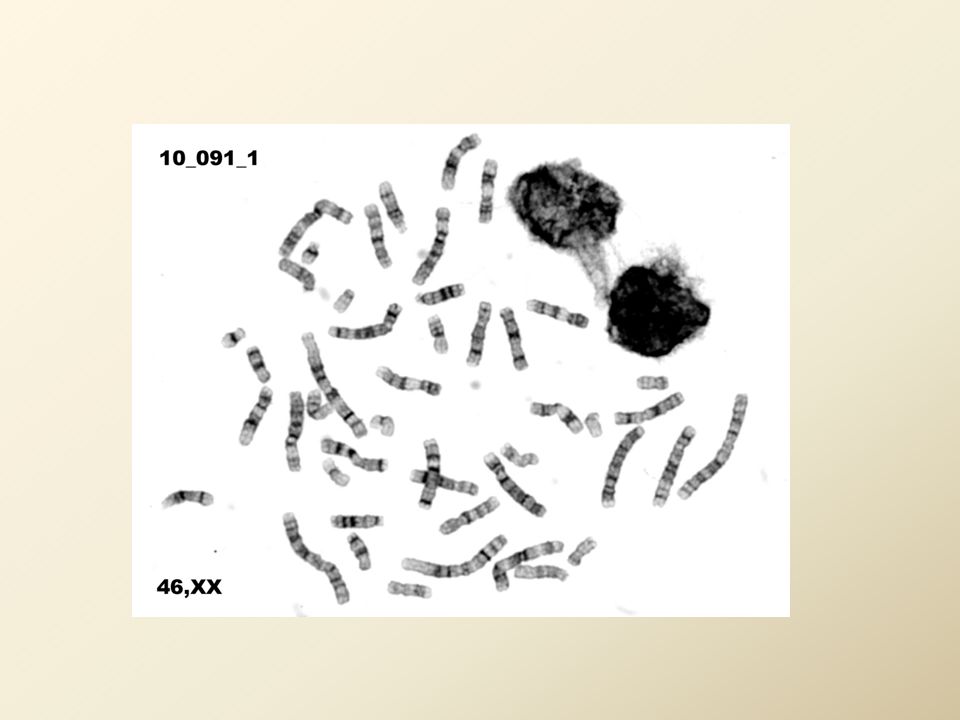
16
Lidské chromozomy 22 párů autozomů 1 pár gonozomů (heterochromozómů) Karyotyp: muž 46, XY, žena 46, XX Chromatin – obsahuje bazické proteiny (histony), DNA, nehistonové proteiny, malé množství RNA Euchromatin despiralizován v interfázi spiralizován v mitoze obsahuje strukturní geny

17
Heterochromatin repetitivní sekvence (konstitutivní heterochromatin)
není transkribován do mRNA (inaktivní) částečně kondenzován (spiralizován) v interfázi pozdě se replikuje tendence shlukovat se a přiléhat k jadernému obalu Konstitutivní (stálý) heterochromatin – v centromerách všech chromozomů bloky heterochromatinu na chrom. 1q, 9q, 16q a Yq (Y- chromatin) obsahuje tandemně se opakující sekvence (tzv.satelitní DNA) Délková variabilita heterochromatinových částí – vznik nerovnoměrným crossing-overem
 částečně kondenzován (spiralizován) v interfázi. pozdě se replikuje. tendence shlukovat se a přiléhat k jadernému obalu. Konstitutivní (stálý) heterochromatin – v centromerách všech chromozomů. bloky heterochromatinu na chrom. 1q, 9q, 16q a Yq (Y- chromatin) obsahuje tandemně se opakující sekvence (tzv.satelitní DNA) Délková variabilita heterochromatinových částí – vznik nerovnoměrným crossing-overem.")
18
Fakultativní heterochromatin (reverzibilní) = strukturně euchromatin, ale chová se jako heterochromatin (potenciálně transkribovatelný, ale je neaktivní Př. Jeden ze dvou X chromozomů u ženy a všech samic savců = geneticky inaktivní, pozdě se replikuje (replikace na konci S fáze) – barvitelný jako sex chromatin= Barrovo tělísko
 – barvitelný jako sex chromatin= Barrovo tělísko.")
20
Karyotyp ženy 46,XX – G pruhy

21
Karyotyp muže - 46,XY – G pruhy

22
Heterochromatin Konstitutitvní heterochromatin bohatý na satelitní DNA (tandemně se opakující DNA sekvence) Stabilita: konstitutivní heterochromatin je stabilní, fakultativní je reversibilní Barvitelnost: konstitutivní heterochromatin se tmavě barví technikou C-pruhování (silná denaturace zejména euchromatinových částí- barví se světle, rezistentní heterochromatin je tmavý) Polymorfismus: konstitutivní heterochromatin je polymorfní (variabilní) ve velikosti i lokalizaci
 Polymorfismus: konstitutivní heterochromatin je polymorfní (variabilní) ve velikosti i lokalizaci.")
23
Vlastnosti heterochromatinu:
1. je silně kondenzován (oba typy: konstitutivní i fakultativní) 2. pozdě se replikuje (oba typy: konstitutivní i fakultativní, inaktivní X se replikuje na konci S fáze) 3. je metylován (cytosiny) 4. Histony heterochromatinu jsou hypoacetylovány (hyperacetylované histony jsou v aktivním chromatinu)
 2. pozdě se replikuje (oba typy: konstitutivní i fakultativní, inaktivní X se replikuje na konci S fáze) 3. je metylován (cytosiny) 4. Histony heterochromatinu jsou hypoacetylovány. (hyperacetylované histony jsou v aktivním chromatinu)")
24
Modifikace histonů, DNA metylace a kondenzace chromozomů
Acetylace histonů odstraní pozitivní náboj histonů – tím uvolní vazbu histonů s elektronegativní DNA = otevřený chromatin (aktivní) Deacetylace histonů obnoví pozitivní náboj histonů – to vede k vazbě mezi DNA a histony (a kondenzaci chromatinových struktur – inaktivní chromatin- nepřístupný pro transkripční faktory) Četné transkripční faktory mají funkci buď Histon Acetyl Transferázy (HAT) v aktivace transkripce nebo Histon De-Acetylázy (HDAC) - potlačení transkripce HDAC – complex mnoha proteinů, obsahuje metyl-cytozin vazebný protein (MeCP1, MeCP2), který se selektivně váže na metylovanou DNA - HDAC je zacílena na metylovanou DNA Další modifikace: př. fosforylace histonu H1 – chrom. kondenzace
 Deacetylace histonů obnoví pozitivní náboj histonů – to vede k vazbě mezi DNA a histony (a kondenzaci chromatinových struktur – inaktivní chromatin- nepřístupný pro transkripční faktory) Četné transkripční faktory mají funkci buď Histon Acetyl Transferázy (HAT) v aktivace transkripce nebo Histon De-Acetylázy (HDAC) - potlačení transkripce. HDAC – complex mnoha proteinů, obsahuje metyl-cytozin vazebný protein (MeCP1, MeCP2), který se selektivně váže na metylovanou DNA - HDAC je zacílena na metylovanou DNA. Další modifikace: př. fosforylace histonu H1 – chrom. kondenzace.")
25
5. Histony v heterochromatinu jsou metylovány (na lysinu 9)
- metylace histonů vytvoří vazebné místo pro chromatin specifický protein HP1 – má úlohu v organizaci heterochromatinu 6. Heterochromatin je transkripčně neaktivní konstitutivní heterochromatin neobsahuje žádné geny fakultativní: geny nejsou transkribovány 7. Heterochromatin se neúčastní genetické rekombinace polymorfismus délky heterochromatinových oblastí způsobuje obtíže při homologním párování 8. Tendence k agregaci v interfázi agregace krátkých ramen akrocentrů – „ nucleolus organiser region = NOR“ – organizátor jadérka 9.Úloha jaderných RNA při vzniku fakultativního heterochromatinu – (př:X inaktivace – mRNA – produkt genu XIST)
")
26
Funkce heterochromatinu:
1. Heterochromatin a euchromatin v různých doménách heterochromatin se nachází na periferii jádra napojen na jadernou membránu aktivní chromatin – centrální uložení v jádře – dovoluje maximální efektivitu replikace a transkripce, chráněn heterochromatinem? 2. Centromerický heterochromatin – úloha při funkci centromery, kohezi sesterských chromatid, normální separaci chromatid v mitóze 3. Úloha v epigenetické regulaci genové exprese Např. v průběhu diferenciace se určité aktivní geny přesunují do domény heterochromatinu – stávají se neaktivními

27
Fakultativní heterochromatin: X-inaktivace, hypotéza Lyonové (Lyon 1961)
U samic savců je pouze jeden X aktivní, druhý (a případně všechny další) je inaktivní (metylace) = inaktivní X je kondenzován v interfázi = je barvitelný Inaktivace začíná v časném embryonálním vývoji (pravděpodobně ve stádiu 1000 – 2000 buněč. embrya) Inaktivace je náhodná (z hlediska rodičovského původu X ), žena = mozaika buněk s inaktivním otcovským a mateřským X Inaktivace je stabilní v liniích dceřinných buněk Inaktivace je reversibilní - v oogenezi jsou oba chromozomy X aktivní
 je inaktivní (metylace) = inaktivní X je kondenzován v interfázi = je barvitelný. Inaktivace začíná v časném embryonálním vývoji (pravděpodobně ve stádiu 1000 – 2000 buněč. embrya) Inaktivace je náhodná (z hlediska rodičovského původu X ), žena = mozaika buněk s inaktivním otcovským a mateřským X. Inaktivace je stabilní v liniích dceřinných buněk. Inaktivace je reversibilní - v oogenezi jsou oba chromozomy X aktivní.")
28
Inaktivace je neúplná některé geny na Xp, i na Xq nejsou inaktivovány strukturně abnormální X – inaktivace je nenáhodná : - balancované (vyvážené) chomozomální abnormality (přestavba, při které žádný materiál nepřebývá,ani nechybí) – je-li X/A (X/autozomální) balancovaná translokace, přednostně se inaktivuje normální X - nebalancované chromozomální abnormality (chromozomální materiál chybí nebo je navíc) – abnormální X je přednostně inaktivní tato nenáhodná inaktivace je důsledkem selekce
 chomozomální abnormality (přestavba, při které žádný materiál nepřebývá,ani nechybí) – je-li X/A (X/autozomální) balancovaná translokace, přednostně se inaktivuje normální X. - nebalancované chromozomální abnormality (chromozomální materiál chybí nebo je navíc) – abnormální X je přednostně inaktivní. tato nenáhodná inaktivace je důsledkem selekce.")
29
Inaktivační centrum v lokusu Xq133,
gen XIST- jeho produkt (mRNA) řídí inaktivaci (v cis poloze), je exprimován jen na inaktivním chromozomu Metoda vyšetření sex chromatinu (skríningová) = orientační ke stanovení pohlaví u sportovců (Sex-chromatin , X-chromatin = Barrovo tělísko)
 řídí inaktivaci. (v cis poloze), je exprimován jen na inaktivním chromozomu. Metoda vyšetření sex chromatinu (skríningová) = orientační ke stanovení pohlaví u sportovců. (Sex-chromatin , X-chromatin = Barrovo tělísko)")
30
Barrovo tělísko (Barr and Bertram, 1949)
")
31
XM XP X inaktivace

32
Nebalancovaná aberace – terminální delece Xp

33
Nenáhodná X inaktivace (detegováno metodou inkorporace BUDr):
unbalanced aberration - terminal deletion of Xp Abnormální X je inaktivní – pozdě se replikuje (světlý)
")
34
Nebalancovaná aberace – prsténcový chromozom X

35
Nenáhodná X inaktivace (detegováno metodou inkorporace BUDr):
Abnormální X (ring X) je inaktivní – pozdě se replikuje (světlý)
 je inaktivní – pozdě se replikuje (světlý)")
36
Balancovaná aberace – reciproká X/A translokace

37
Nenáhodná X inaktivace (detegováno metodou inkorporace BUDr):
Balancovaná aberace – X/A reciproká translokace Normální X je inaktivní – pozdě se replikuje (světlý)
")
38
Thompson &Thompson: Klinická genetika,6. vyd.
Kap. 2: Chromozomální podstata dědičnosti: podkapitoly: Lidské chromozomy, Buněčný cyklus somatických buněk, Lidský karyotyp Kap.3: Lidský genom, struktura a funkce chromozomů:podkapitola Struktura lidských chromozomů Kap. 10 (část): Chromozom X –podkapitola X inaktivace + doplnění informací z prezentace Prezentace: klíč k zápisu: genetika
: Chromozom X –podkapitola X inaktivace. + doplnění informací z prezentace. Prezentace: id=191. klíč k zápisu: genetika.")
Podobné prezentace








>")




>")



>")

