Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
F1190 Úvod do biofyziky Masarykova Univerzita Podzimní semestr 2012 18.11.2014 Dynamika Prof. Jiří Kozelka, Biofyzikální Laboratoř, Ústav fyziky kondenzovaných látek, PřF MU, Kotlářská 2, kozelka.jiri@gmail.comkozelka.jiri@gmail.com Doporučená doplňující četba: Molecular Modelling: Principles and Applications, second edition Andrew R. Leach Pearson Education EMAPearson Education EMA, 2001 ISBN 0-582-38210-6

2
Jednotlivé konformace se liší stabilitou. Stabilitu konformace k (danou Kartézskými souřadnicemi x ki, y ki, z ki všech atomů i) určuje její potenciální energie E: Biologické makromolekuly nejsou rigidní. Jejich struktury vykazují různé stupně volnosti: - Vazby a vazebné úhly vibrují kolem rovnovážných poloh - Jednoduché vazby umožňují rotaci a tím konformační změny - Cyklické podjednotky (např. cukry) přecházejí mezi energeticky výhodnými konformacemi (pro ribózu: N a S) Konformační dynamika makromolekul
 určuje její potenciální energie E: Biologické makromolekuly nejsou rigidní. Jejich struktury vykazují různé stupně volnosti: - Vazby a vazebné úhly vibrují kolem rovnovážných poloh - Jednoduché vazby umožňují rotaci a tím konformační změny - Cyklické podjednotky (např. cukry) přecházejí mezi energeticky výhodnými konformacemi (pro ribózu: N a S) Konformační dynamika makromolekul.")
3
Force-field parameters (fix) Structural variables {f(x ki, y ki, z ki )} Jednotlivé konformace se liší stabilitou. Stabilitu konformace k (danou Kartézskými souřadnicemi x ki, y ki, z ki všech atomů i) určuje její potenciální energie E: E(x ki, y ki, z ki ) deformace vazebdeformace vazeb. úhlů deformace torzních úhlů nevazebné kontakty Lennard-JonesCoulomb
 určuje její potenciální energie E: E(x ki, y ki, z ki ) deformace vazebdeformace vazeb. úhlů deformace torzních úhlů nevazebné kontakty Lennard-JonesCoulomb.")
7
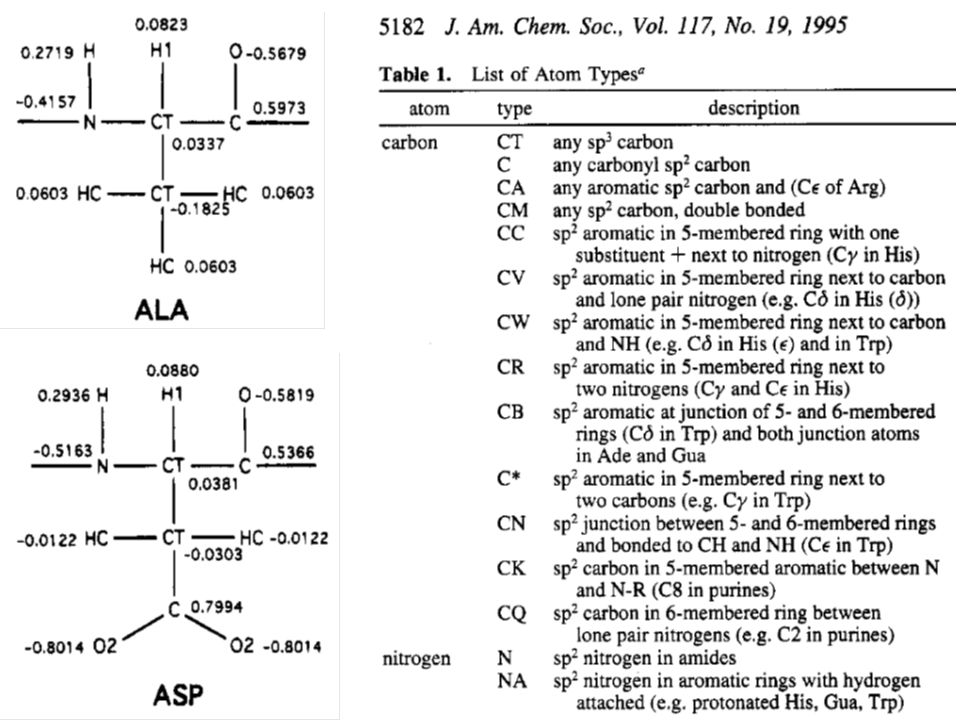
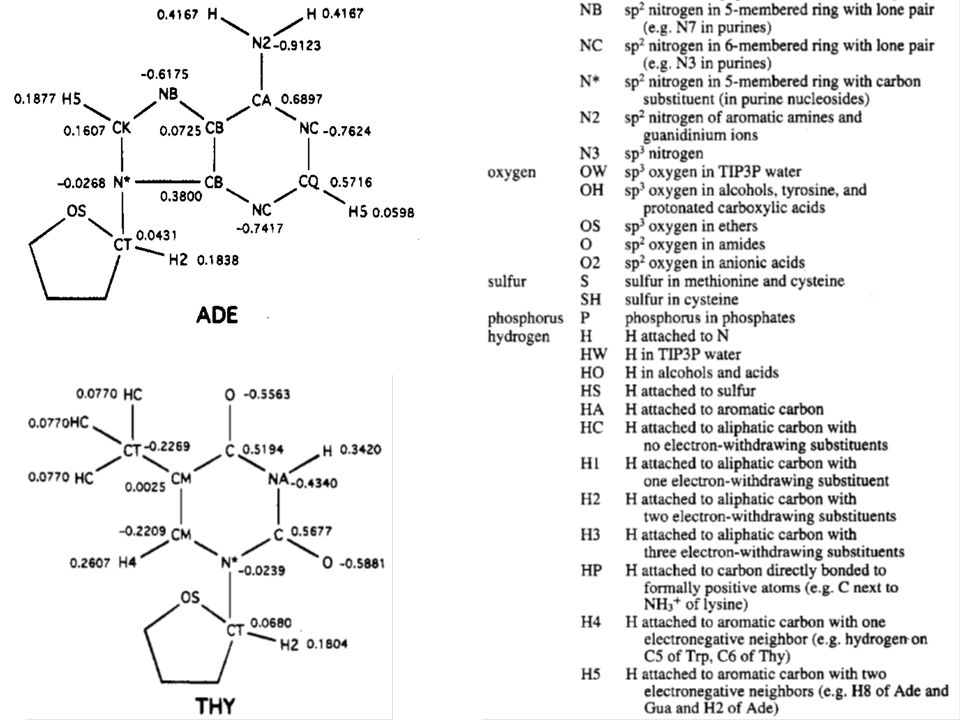
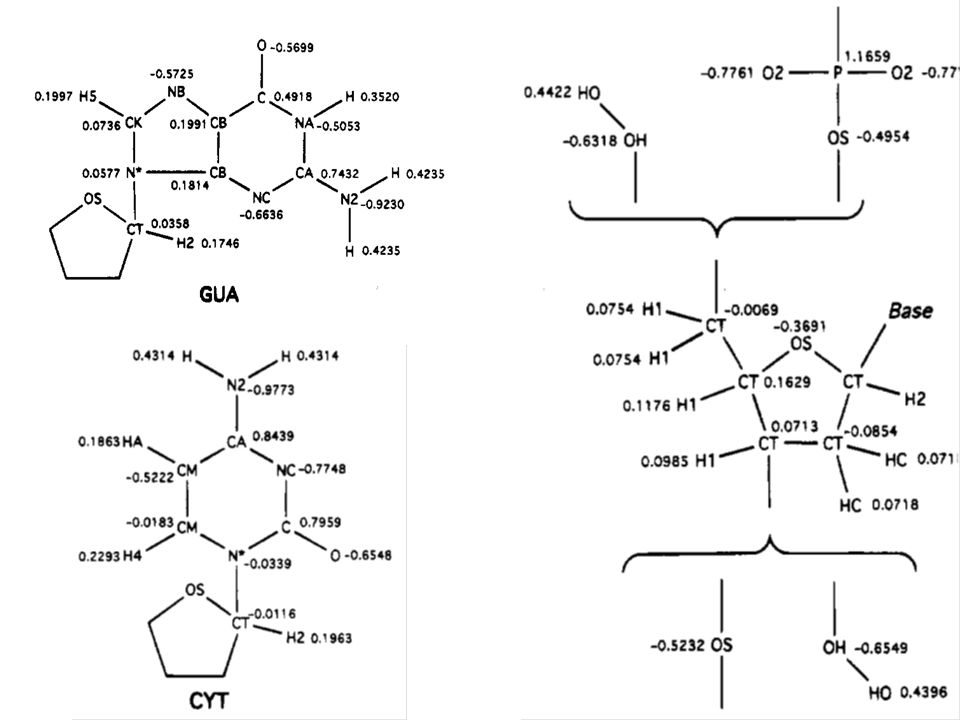
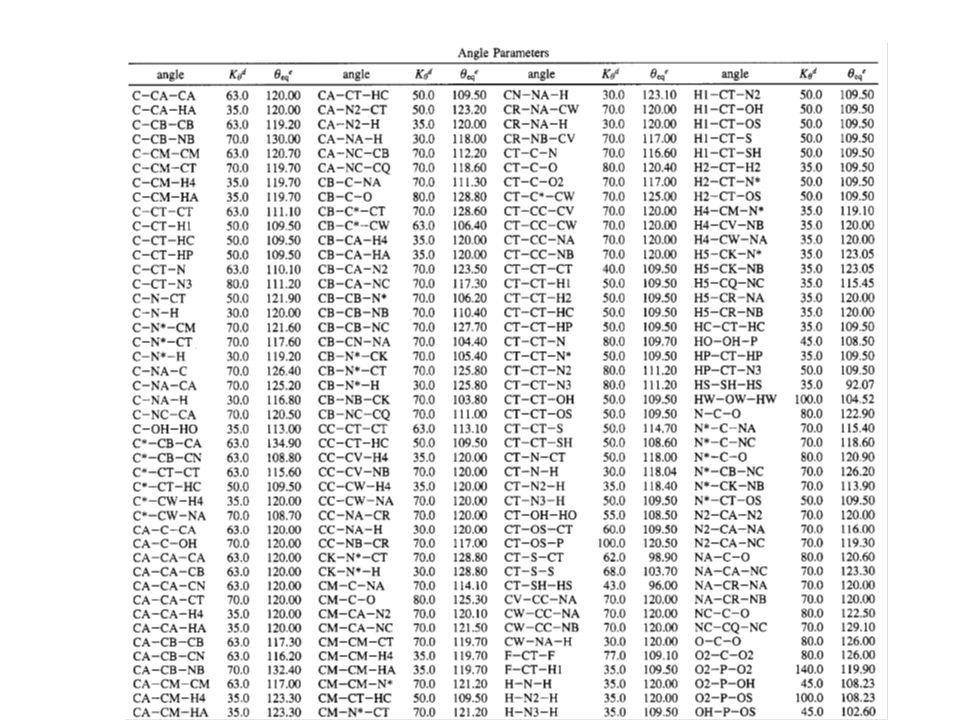
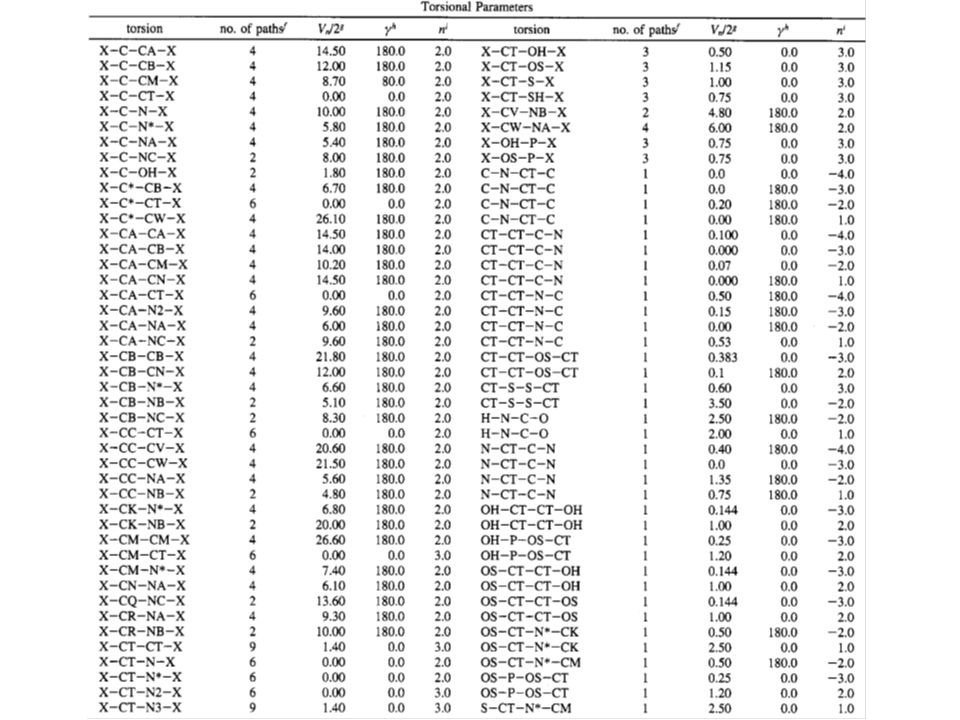
Parametry silového pole programu AMBER (1995) k modelování proteinů a nukleových kyselin
 k modelování proteinů a nukleových kyselin")
10
Methods of (energy) minimization 1. Methods without calculation of energy gradient 1.1. Grid search Energy is calculated for a grid of points spanning the conformational space energy minimum point of lowest energy found by grid search How to refine the search for the energy minimum?- new grid search with an finer grid around the lowest energy point - calculation of the energy gradient and determination of step size using a quadratic line-search (see Section 2)
.")
11
Jak určíme délku kroku? Tzv. line search: 1. Zkusmo posuneme souřadnice o vektory -s 1 f a -s 2 f. 2. Počátečním bodem a těmito dvěma novými body proložíme parabolu a posuneme strukturu do jejího minima. 3. Nový gradient je pak kolmý na předešlý f (k+1) f k = 0 Co je gradient? Gradient funkce je vektor, jehož složky tvoří jednotlivé parciální derivace funkce podle jejích souřadnic např. f(x,y) = x 2 + 2y 2 grad(f(x,y)) = (2x, 4y) ff Při metodě nestrmějšího spádu posunujeme souřadnice ve směru opačném gradientu. 2. Methods using the energy gradient 2.1. Method of the steepest descent- metoda nejstrmějšího spádu
 f k = 0 Co je gradient. Gradient funkce je vektor, jehož složky tvoří jednotlivé parciální derivace funkce podle jejích souřadnic např. f(x,y) = x 2 + 2y 2 grad(f(x,y)) = (2x, 4y) ff Při metodě nestrmějšího spádu posunujeme souřadnice ve směru opačném gradientu. 2. Methods using the energy gradient 2.1. Method of the steepest descent- metoda nejstrmějšího spádu.")
12
2.2. Conjugated gradient method- metoda konjugovaného gradientu K určení vektoru, o který se posunou souřadnice atomů, je použit jednak gradient energie, jednak vektor posunu v předešlém kroku. Tato metoda bere v úvahu zakřivení funkce energie na souřadnicích (tedy parciální 2. derivace), aniž bychom tyto 2. derivace explicitně počítali. 3. Metody využívající 2. parciální derivace funkce energie na souřadnicích. Umožňují optimální určení posunových vektorů, takže k dosažení energetického minima je zapotřebí méně kroků. Výpočet druhých derivací je však sám výpočetně náročný. Tyto algoritmy se většinou používají ke zjemnění modelů, které jsou již blízko energetickému minimu.
, aniž bychom tyto 2. derivace explicitně počítali. 3. Metody využívající 2. parciální derivace funkce energie na souřadnicích. Umožňují optimální určení posunových vektorů, takže k dosažení energetického minima je zapotřebí méně kroků. Výpočet druhých derivací je však sám výpočetně náročný. Tyto algoritmy se většinou používají ke zjemnění modelů, které jsou již blízko energetickému minimu..")
13
3.Molecular dynamics simulations Are based on the partitioning of the motion into infinitesimal time steps dt. The force acting on individual atoms is calculated as the sum of pair interactions with each of the other atoms. The acceleration of each atom is then defined from Newtons equation of motion, a i =F i /m i, and is considered constant during the interval dt. The most well- known algorithm for the calculation of coordinates is the Verlet algorithm.

24
Biological time scale Bond vibrations1 fs(10 -15 s) Sugar repuckering1 ps(10 -12 s) DNA bending 1 ns(10 -9 s) Domain movement1 s(10 -6 s) Base pair opening1 ms(10 -3 s) Transcription2.5 ms / nucleotide Protein synthesis6.5 ms / amino acid Protein folding~ 10 s RNA lifetime~ 300 s MD simulations spectroscopic methods (UV, CD, NMR)
 Sugar repuckering1 ps( s) DNA bending 1 ns(10 -9 s) Domain movement1 s(10 -6 s) Base pair opening1 ms(10 -3 s) Transcription2.5 ms / nucleotide Protein synthesis6.5 ms / amino acid Protein folding~ 10 s RNA lifetime~ 300 s MD simulations spectroscopic methods (UV, CD, NMR)")
25
Analýza molekulových simulací Příklad: simulace DNA Cukr-fosfátová páteř DNA je tvořena výlučně jednoduchými vazbami. Torzní úhly podle nich jsou označovány (obrázek). Rotací podle jednoduchých vazeb přechází DNA mezi 2-3 stabilními konformacemi oddělenými energetickými bariérami. Takové konformační přechody můžeme přehledně sledovat vynesením jednotlivých torzních úhlů jako funkce času. Vzdálenosti mezi jednotlivými atomy vodíku můžeme měřit pomocí nukleární magnetické rezonance (NMR), kvantifikací tzv. nukleárního Overhauserova efektu (NOE). Porovnáním změřených hodnot s hodnotami ve strukturním modelu můžeme hodnotit kvalitu našeho modelu. Měření NOE je ovšem pomalé- trvá zhruba několik ms až ms. Rychlejší konformační přechody (např. přechody N-S u deoxyribózy, ohýbání DNA) nemůžeme sledovat- měříme časový průměr.
. Rotací podle jednoduchých vazeb přechází DNA mezi 2-3 stabilními konformacemi oddělenými energetickými bariérami. Takové konformační přechody můžeme přehledně sledovat vynesením jednotlivých torzních úhlů jako funkce času. Vzdálenosti mezi jednotlivými atomy vodíku můžeme měřit pomocí nukleární magnetické rezonance (NMR), kvantifikací tzv. nukleárního Overhauserova efektu (NOE). Porovnáním změřených hodnot s hodnotami ve strukturním modelu můžeme hodnotit kvalitu našeho modelu. Měření NOE je ovšem pomalé- trvá zhruba několik ms až ms. Rychlejší konformační přechody (např. přechody N-S u deoxyribózy, ohýbání DNA) nemůžeme sledovat- měříme časový průměr..")
26
Příklad: Molekulární simulace DNA-oligonukleotidu d(GCCGGGTCGC)-d(GCGACCCGGC) po reakci s protinádorovým lékem cisplatina (cis-[PtCl 2 (NH 3 ) 2 ]), kde hlavním produktem je přemostění atomů N7 guaninů G4 a G5 atomem platiny: 1. Reakce 10 molů oligonukleotidu s cis-[PtCl 2 (NH 3 ) 2 2. Oddělení hlavního produktu od vedlejších pomocí HPLC- získán 1 mmol čistého hl. produktu 3. Studium produktu v 0.5 mL D 2 O/H 2 O (koncentrace: 2 mM) pomocí NMR 4. Molekulové simulace produktu v H 2 O s využitím dat z NMR ke zjemnění modelu. Délka simulovaného času: 20 ns
![Příklad: Molekulární simulace DNA-oligonukleotidu d(GCCGGGTCGC)-d(GCGACCCGGC) po reakci s protinádorovým lékem cisplatina (cis-[PtCl 2 (NH 3 ) 2 ]), kde hlavním produktem je přemostění atomů N7 guaninů G4 a G5 atomem platiny: 1.](http://images.slideplayer.cz/12/4077073/slides/slide_26.jpg "Reakce 10 molů oligonukleotidu s cis-[PtCl 2 (NH 3 ) 2 2. Oddělení hlavního produktu od vedlejších pomocí HPLC- získán 1 mmol čistého hl. produktu 3. Studium produktu v 0.5 mL D 2 O/H 2 O (koncentrace: 2 mM) pomocí NMR 4. Molekulové simulace produktu v H 2 O s využitím dat z NMR ke zjemnění modelu. Délka simulovaného času: 20 ns.")
27
Torzní úhly a jsou silně korelovány. Stabilní konformace jsou: 180° („trans“); -80° („gauche - “) Tato konformace se nazývá BI- DNA -80° („gauche - “); 180° („trans“) Tato konformace se nazývá BII- DNA BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI [°] [°]
; -80° („gauche - ) Tato konformace se nazývá BI- DNA -80° („gauche - ); 180° („trans ) Tato konformace se nazývá BII- DNA BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI BII BI [°] [°].")
28
[°] Konformaci deoxyribózy lze vyčíst z torzního úhlu : 80° charakterizuje konformaci N = 110°-150° charakterizuje konformaci S Pro B-DNA je typická konformace S, s občasnými přechody do N. Přemostění G4 a G5 platinou má za následek preferenci G4 a C3 pro konformaci N.
![ [°] Konformaci deoxyribózy lze vyčíst z torzního úhlu : 80° charakterizuje konformaci N = 110°-150° charakterizuje konformaci S Pro B-DNA je typická konformace S, s občasnými přechody do N.](http://images.slideplayer.cz/12/4077073/slides/slide_28.jpg "Přemostění G4 a G5 platinou má za následek preferenci G4 a C3 pro konformaci N..")
30
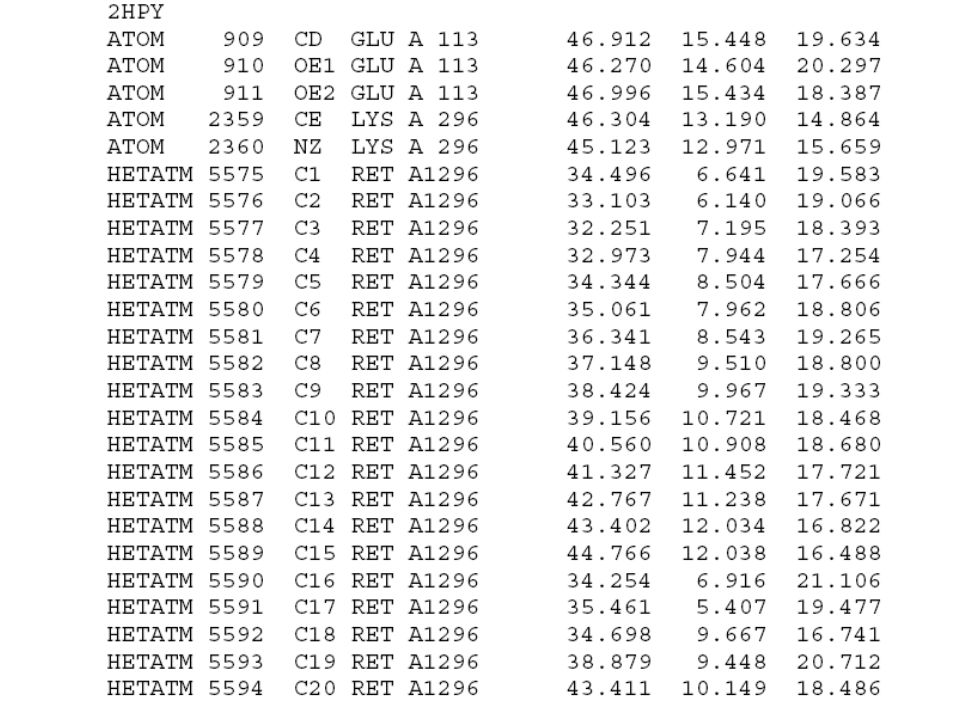
Opsin + 11-cis-retinal = „oční pigment“ („visual pigment“) Lidská sítnice obsahuje 4 různé oční pigmenty: Rhodopsin v tyčinkách, max = 500 nm (krystalová struktura známa) Jodopsin v čípcích Červený, max = 557 nm Zelený, max = 530 nm Modrý, max = 425 nm (struktura jodopsinů je podobná struktuře rhodopsinu, ale zatím přesně neurčena) Většina mechanických studií byla zatím věnována rhodopsinu.
 Lidská sítnice obsahuje 4 různé oční pigmenty: Rhodopsin v tyčinkách, max = 500 nm (krystalová struktura známa) Jodopsin v čípcích Červený, max = 557 nm Zelený, max = 530 nm Modrý, max = 425 nm (struktura jodopsinů je podobná struktuře rhodopsinu, ale zatím přesně neurčena) Většina mechanických studií byla zatím věnována rhodopsinu.")
Podobné prezentace














.>")





 MM/MD.>")


>")
>")







