Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Vnitrobuněčné oddíly Třídění a transport proteinů

2
Kompartmenty živočišné buňky

3
Kompartmenty živočišné buňky - TEM

4
Prokaryotická buňka Eukaryotická buňka

5
Typická rostlinná buňka
Golgiho aparát centrální vakuola vesikul drsné ER ribosomy (na ER) mikrofilamenta ribosomy (volné) hladké ER mitochondrie DNA+nucleoplasma chloroplast jadérko jádro jaderná membrána plasmatická membrána mikrotubuly buněčná stěna
 mikrofilamenta. ribosomy (volné) hladké ER. mitochondrie. DNA+nucleoplasma. chloroplast. jadérko. jádro. jaderná membrána. plasmatická membrána. mikrotubuly. buněčná stěna.")
6
Živočišná buňka Rostlinná buňka

7
Funkce hlavních vnitrobuněčných oddílů
Jádro – obsahuje buněčný genom, syntéza DNA and RNA Cytosol – syntéza proteinů, glykolýza, metabolické dráhy pro syntézu aminokyselin, nukleotidů, etc Endoplasmatické retikulum – syntéza membránových a sekrečních proteinů, syntéza většiny lipidů Golgiho aparát – kovalentní modifikace proteinů z ER, třídění proteinů a lipidů pro sekreci a transport do jiných částí buňky Mitochondrie – syntéza ATP oxidační fosforylací Chloroplasty – syntéza ATP a fixace uhlíku fotosyntézou Lyzosomy – odbourávání látek uvnitř buňky Endosomy – třídění materiálu z endocytózy a GA Peroxisomy – oxidace toxických molekul, beta oxidace mastných kyselin

8
Tři základní typy vnitrobuněčného transportu proteinů
Transport jaderným pórem (Gated transport) Transmembránový transport Vesikulární transport
 Transmembránový transport. Vesikulární transport.")
9
Mapa transportu proteinů
Vznik a správná funkce vnitřních kompartmentů zcela závisí na přesném cílení proteinů.

10
Třídění proteinů – signální sekvence

11
Jednoduchý experiment dokazuje, že většina třídících signálů se skládá z krátkého úseku aminokyselinové sekvence nazývané “signální sekvence”.

12
GFP

13
Cytoplasmatický Jaderný

14
Actin-GFP PAX-GFP

15
Signální sekvence

16
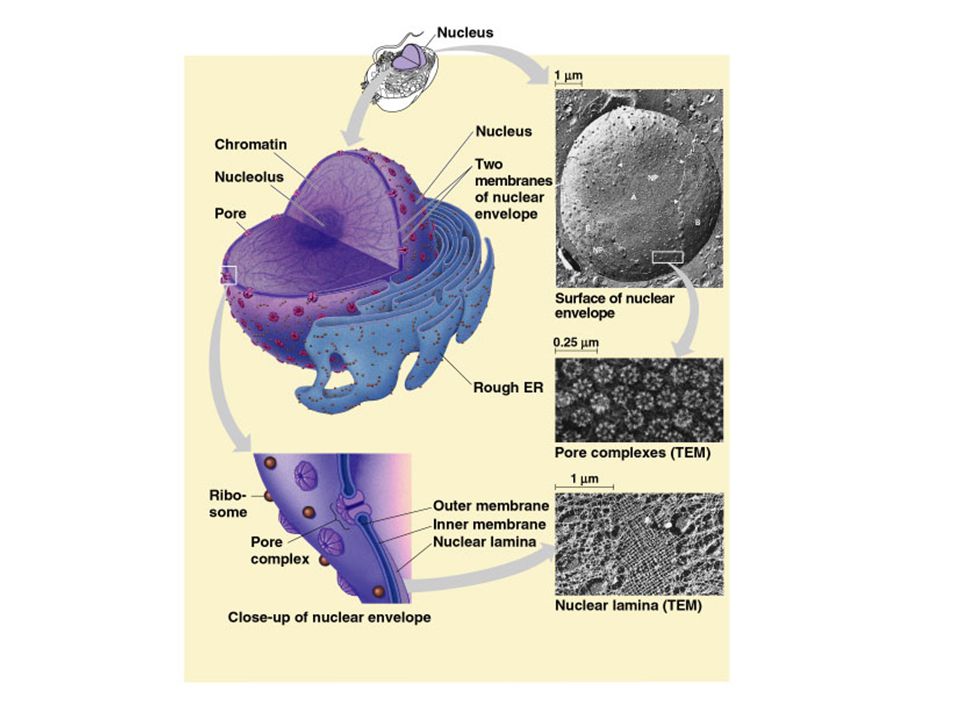
Jaderný import a export
Jaderný obal se skládá ze dvou koncentrických lipidových dvouvrstev. Perinukleární prostor je kontinuální s lumenem ER. Přes komplex jaderného póru probíhá obousměrný transport.

18
Jaderný pór Komplex jaderného póru je vodní kanál, který umožňuje difuzi malých molekul a proteinů do 60kD. Transport těchto malých molekul je pasivní.

19
Molekuly větší než 9 nm nebo 60 kDa musí mít jaderný lokalizační signál (NLS) a jsou aktivně transportovány do jádra.
 a jsou aktivně transportovány do jádra.")
20
Transport velkých proteinů do jádra vyžaduje signální sekvenci nazývanou jaderný lokalizační signál (NLS). NLS může být umístěn kdekoliv v primární sekvenci proteinu. Obvykle je bohatý na arginin a lysin a je poměrně krátký.

21
Mechanismus jaderného transportu

22
NLS směruje proteiny k transportu přes komplex jaderného póru
NLS směruje proteiny k transportu přes komplex jaderného póru. Proteiny si během transportu přes jaderný pór zachovávají svou terciální a kvarterní strukturu. Jsou-li částice koloidního zlata potaženy NLS, můžeme vidět částice procházet jadernými póry. Maximální velikost částice zlata, která může být transportována je 26 nm. Jelikož zlaté částice nemohou být stlačeny, otvor jaderného póru musí být schopen se rozšířit.

23
Protein Ran (monomerická GTPáza) je hnací silou transportu přes komplex jaderného póru
Ran-GAP se nachází výhradně v cytoplasmě = Ran-GDP se hromadí vně jádra. Ran-GEF se nachází uvnitř jádra = Ran-GTP se hromadí uvnitř jádra. Ran: Monomerická GTPáza Ran-GAP: GTPázu-aktivující protein Ran-GEF: Guaninový výměnný faktor

24
Receptory jaderného importu (Nuclear Import Receptors) rozpoznávají NLS.
 rozpoznávají NLS.")
25
Transport proteinů do mitochondrií a chloroplastů
Organely specializované na syntézu ATP. Většina proteinů je kódovaná jaderným genomem a syntetizovaná v cytoplasmě. Proteiny musí být transportovány do jednoho z kompartmentů nebo membrán těchto organel.

26
Mitochondriální (chloroplastové) proteiny
 proteiny")
27
Mitochondriální signální sekvence:
amfipatický alpha-helix, pozitivně nabitý Proteinové transportéty mitochondriálních membrán

28
Import proteinů do mitochondriální matrix je řízen N-terminální signální sekvencí.
Signální sekvence na N-konci naváže protein na TOM komplex na vnější mitochondriální membráně. TOM komplex je současně receptor pro signální sekvenci a translokátor. TOM komplex s navázaným proteinem se přemístí do tzv. membránového kontaktního místa. Protein prochází přes TOM transportér do transportéru komplexu TIM na vnitřní mitochondriální membráně. Protein je transportován do mitochondriální matrix a signální sekvence (=signální peptid) je enzymaticky odštěpena tzv. signální peptidázou.
 je enzymaticky odštěpena tzv. signální peptidázou.")
29
Energetika mitochondriálního transportu: hydrolýza ATP a elektrochemický protonový gradient.
Hydrolýza ATP – chaperonové proteiny v cytoplasmě Elektrochemický protonový gradient na vnitřní mitochondriální membráně Hydrolýza ATP – chaperonové proteiny v mitochondriální matrix

30
Transport do chloroplastů je podobný transportu do mitochondrií
Transport do chloroplastů je podobný transportu do mitochondrií. Vyjímka – tylakoidy (třetí membrána), potřeba druhé signální sekvence U chloroplastů je protonový gradient na tylakoidní membráně a podílí se zde na transportu do tylakoidů. Transport přes chloroplastovou vnitřní membránu (nemá elektrochemický gradient) využívá energii z hydrolýzy GTP a ATP.
, potřeba druhé signální sekvence. U chloroplastů je protonový gradient na tylakoidní membráně a podílí se zde na transportu do tylakoidů. Transport přes chloroplastovou vnitřní membránu (nemá elektrochemický gradient) využívá energii z hydrolýzy GTP a ATP.")
31
Srovnání jaderného a mitochondriálního importu

32
Peroxisomy obsahují oxidativní enzymy, odbourávání toxických molekul, beta oxidace mastných kyselin, syntéza prekursoru myelinu, krátká signální sekvence

33
Endoplasmatické retikulum

34
drsné ER hladké ER Drsné ER -ribosomy
Lumen ER tvoří kontinuální prostor s perinukleárním prostorem. hladké ER

35
Funkce endoplasmatického retikula
Počátek nově syntetizovaných proteinů určených pro Golgi, Endosomy, Lyzosomy, Sekreční vesikuly a Plasmatickou membránu Vytváří orientaci proteinů v membráně Místo syntézy fosfolipidů a cholesterolu Počáteční místo pro N-glykosylaci proteinů Hromadění Ca++ - sarkoplasmatické retikulum ve svalech

36
Volné a vázané ribosomy

37
Signální sekvence ~20 aminokyselin, bohatá na hydrofobní aminokyseliny, nejčastěji na N-konci
SRP-signál rozpoznávající protein (signal recognition particle) SRP receptor Proteinový translokátor Translace a translokace probíhá současně - ko-translační transport Energie pro transport je získána z vlastního procesu translace – jak polypeptid roste, je protlačován přes proteinový translokátor
 SRP receptor. Proteinový translokátor. Translace a translokace probíhá současně - ko-translační transport. Energie pro transport je získána z vlastního procesu translace – jak polypeptid roste, je protlačován přes proteinový translokátor.")
38
Signální sekvence (signální peptid) sekrečních proteinů je odštěpena signální peptidázou.
 sekrečních proteinů je odštěpena signální peptidázou.")
39
Translokace proteinů přes membránu ER

40
Transmembránový protein

41
Ko-translační transport musí být schopen vytvářet řadu různých konfigurací.
For both single-pass and multipass transmembrane proteins, some types will have the N-terminus projecting into the cytosol and others will have the C-terminus projecting into the cytosol.

42
Membránové proteiny s více transmembránovými doménami

43
Things can get pretty complicated!

44
Membránové proteiny ukotvené přes glykolipidy - ER

45
N-glykosylace proteinů (N-linked glycosylation)
Dolichol N-linked glycosylation. Glykosylace – signál pro rozlišení správně složených proteinů

46
Krevní skupiny ABO jsou určeny dvěmi glykosyltransferázami

47
Skládání proteinů v ER

48
Nesprávně složené proteiny jsou exportovány z ER do cytoplasmy a degradovány v proteasomu.

49
Fosfolipidy jsou syntetizovány na cytoplasmatické straně membrány ER

50
Fosfolipidové translokátory

51
Transfer lipidů do jiných organel
Jaderná membrána – laterální difuze Organely sekreční dráhy – vesikulární transport Mitochondrie, chloroplasty a peroxisomy – proteinový transport (Phospholipid exchange proteins)
")
52
Intracelulární vezikulární transport

53
Golgiho aparát hraje centrální roli ve vnitrobuněčném vesikulárním transportu

54
Vesikulární transport přepravuje proteiny a membrány mezi jednotlivými buněčnými kompartmenty
Vesikulární transport zachovává membránovou orientaci

55
Proteiny jsou předávány z ER do Golgiho aparátu.

56
TEM Golgiho komplexu

57
Golgiho aparát – trojrozměrná rekonstrukce
Proteiny z ER vstupují do Golgiho aparátu na cis straně

58
Golgiho aparát má dvě hlavní funkce:
1. Modifikuje N-vázané oligosacharidy a přidává O-vázané oligosacharidy. 2. Třídí proteiny – po odchodu z trans Golgi komplexu jsou doručeny do správných buněčných kompartmentů v transportních vesikulech.

59
Modifikace N-vázaných oligosacharidů je prováděna řadou enzymů Golgiho aparátu.
1. Třídění proteinů 2. Ochrana proteinů před proteázami 3. Buněčná adheze (selectiny) 4. Imunitní systém
 4. Imunitní systém.")
60
Na tvorbě (pučení) vesikulů se podílejí tři typu plášťových proteinů (coat proteins)
 vesikulů se podílejí tři typu plášťových proteinů (coat proteins)")
61
Klathrinové váčky

62
Klathrin se navazuje přes adaptiny na receptory v donorové membráně
Klathrin se navazuje přes adaptiny na receptory v donorové membráně. Receptory vážou specifický náklad. Klathrin se sestavuje do kulovitého útvaru a vtahuje s sebou část donorové membrány. Dynamin odškrtí váček od membrány. Požadovaná energie: Dynamin – hydrolýza GTP Chaperonové proteiny - hydrolýza ATP

63
Cílení transportních vesikulů: komplementární sady SNARE proteinů (v-SNARE a t-SNARE).
.")
64
Správně složené proteiny jsou transportovány v COPII vesikulech do cis-Golgiho komplexu.
COPII plášť je odstraněn a vesikuly vzájemně fúzují – vesikulární tubulární cluster. VTC je transportován podél mikrotubulů za účasti motorových proteinů. VTC fúzuje s cis-Golgi komplexem.

65
Není zcela jasné, jak proteiny postupují přes Golgiho komplex.
1. Model: Stacionární kompartmenty a vesikulární transport mezi nimi. 2. Model Velké pohybující se kompartmenty, které postupně dozrávají do TGN, zpětný návrat enzymů pomocí vesikulárního transportu.

66
Konečnou destinací pro některé proteiny z TGN je lyzosom
Konečnou destinací pro některé proteiny z TGN je lyzosom. Mezi ně patří především kyselé hydrolázy. Lyzosomy představují „žaludek“ buňky. jedna membrána kyselé hydrolázy degradace makromolekul H+ ATPázová pumpa pH ~5

67
Makromolekuly degradované v lyzosomech: endocytoza, fagocytoza nebo autofagie.

68
Endocytoza vs. Exocytoza
Základní typy endocytozy: Fagocytoza – velké částice a buňky Pinocytoza – malá množsví extracelulární tekutiny Receptorová endocytoza – seletivní transport membránových receptorů a jejich navázaných ligandů Endocytoza odebírá značné množství plasmatické membrány. Odebrané složky plasmatické membrány jsou nahrazovány procesem nazývaným exocytoza.

69
Fagocytoza – bílé krvinky: Makrofágy, Neutrofily a Dendritické buňky
Receptory v plasmatické membráně (fosfatidylserin)
")
70
Pinocytoza – klathrinové váčky

71
Endocytická dráha - od plasmatické membrány po lyzosom
časný endosom multivesikulární tělísko pozdní endosom lyzosom

72
Transcytosa – transport proteinů přes epitel
Transport protilátek z mateřského mléka přes střevní epitel. Kyselé pH ve střevě – vazba protilátky na Fc receptor Neutrální pH extracelulární tekutiny – disociace protilátky z receptoru

73
Exocytoza

74
The End

Podobné prezentace
 CYTOLOGIE (stavba buněk) b) HISTOLOGIE.>")










>")







