Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Expresní systémy živé systémy využívající rekombinantních DNA technologií pro produkci bioorganických látek (především proteinů)
")
2
Heterologní expresní systémy
Bakteriální Kvasinkové Hmyzí buňky Savčí buňky Transgenní rostliny

3
požadavky na růstové medium minimální komplexní medium
charakteristika E. coli kvasinky hmyzí buňky savčí buňky buněčný růst rapidní (30min) rapidní (90min) pomalý (18-24h) pomalý (24 h) požadavky na růstové medium minimální komplexní medium cena růstového media nízká vysoká množství exprimovaného proteinu velké malé až velké malé nebo střední extracelulární exprese sekrece do inkluzních tělísek sekrece do media posttranslační modifikace skládání proteinů obvykle nutné dodatečné složení v některých případech nutné dodatečné složení řádně složené proteiny N-glykosylace - vysoký obsah manosy jednoduché, bez sialové kyseliny komplexní O-glykosylace + fosforylace acetylace acylace γ-karboxylace
 rapidní (90min) pomalý (18-24h) pomalý (24 h) požadavky na růstové medium. minimální. komplexní medium. cena růstového media. nízká. vysoká. množství exprimovaného proteinu. velké. malé až velké. malé nebo střední. extracelulární exprese. sekrece do inkluzních tělísek. sekrece do media. posttranslační modifikace. skládání proteinů. obvykle nutné dodatečné složení. v některých případech nutné dodatečné složení. řádně složené proteiny. N-glykosylace. - vysoký obsah manosy. jednoduché, bez sialové kyseliny. komplexní. O-glykosylace. + fosforylace. acetylace. acylace. γ-karboxylace.")
4
Bakteriální expresní systémy
Escherichia coli rychlá produkce nejvýkonnější (až 0.5 g na 1 litr kultury) levné a jednoduché na manipulaci chybí posttranslační úprava proteinů (neaktivní produkty eukaryotních genů) přirozeně neprobíhá sekrece do média preferují jiný genetický kód než vyšší eukaryota
 levné a jednoduché na manipulaci. chybí posttranslační úprava proteinů. (neaktivní produkty eukaryotních genů) přirozeně neprobíhá sekrece do média. preferují jiný genetický kód než vyšší eukaryota.")
5
rekombinatní genetická informace je vklonována do vhodného expresního plasmidu, nedochází k integraci do genomu TA klonování TA TOPO klonování

6
rekombinační klonování
místně specifický rekombinatní systém bakteriofága lambda – att attB x attP ↔ attL x attR (“x” znamená rekombinaci). vzájemná rekombinace mezi nimi probíhá podle daných pravidel obě rekombinace jsou katalyzovány proteiny kódované jak αDNA tak bakteriální. selekce antibiotikum a gen ccdB mezi rekombinantními místy, protein ccdB inhibuje bakteriální DNA gyrasu a způsobuje smrt buněk nesoucí prázdný vektor
. vzájemná rekombinace mezi nimi probíhá podle daných pravidel. obě rekombinace jsou katalyzovány proteiny kódované jak αDNA tak bakteriální. selekce antibiotikum a gen ccdB mezi rekombinantními místy, protein ccdB inhibuje bakteriální DNA gyrasu a způsobuje smrt buněk nesoucí prázdný vektor.")
7
rekombinační klonování
LR reakce BP reakce se účastní integrasa a ekscionasa (αDNA) se účastní IHF a integrasa a IHF (integration host factor) z bakterie rekombinační klonování
 se účastní IHF a integrasa. a IHF (integration host factor) z bakterie. rekombinační klonování.")
8
Invitrogen dodává všechny typy vektorů kompatibilních pro rekombinantní klonování
Reakce trvá 1 hodinu při laboratorní teplotě zachovávání čtecích rámců (ORF), žádné složité plánování
, žádné složité plánování.")
9
N-terminal 6xHis tag umožňuje velice účinnou purifikaci proteinu pomocí metal-chelatační chromatografie popř. detekci pomocí Anti-HisG protilátky EK rozpoznávací sekvence pro specifickou enterokinasu, odštěpuje His-tag T7 transcription termination region silný terminační systém T7 bacteriofága gen pro rezistenci k ampicilinu umožňuje selekci plasmidu v E. coli pUC origin zajišťuje vysokou replikaci plasmidu a růst E. coli C-terminal V5 epitope tag umožňuje detekci fúzního proteinu pomocí Anti-V5 protilátky T7 promoter přesná exprese heterogenního proteinu Ribosome binding site TOPO klonovací místo pro PCR produkt Xpress™ epitop (Asp-Leu-Tyr-Asp-Asp-Asp-Asp-Lys) umožňuje detekci fúzního proteinu pomocí Anti-Xpress™ protilátky
 umožňuje detekci fúzního proteinu pomocí Anti-Xpress™ protilátky.")
10
exprimovaný protein „“ „“ detekce izolace izolace detekce
reverse primerTAG „“ signální peptid exprimovaný protein V5 epitop his tag ATGforward primer detekce izolace reverse primerTAG „“ his tag x-press signální peptid exprimovaný protein forward primer izolace detekce

11
usnadnění izolace rekombinantního proteinu koncové značky
nejpoužívanější N a C-terminální značky (tagy): His-tag pro metal chelatační chromatografii (Ni) FLAG epitope - tag DYKDDDDK (Sigma; specifická protilátka) CBP - calmodulin binding peptide (26 AK) CBD - cellulose binding domain
: His-tag pro metal chelatační chromatografii (Ni) FLAG epitope - tag DYKDDDDK. (Sigma; specifická protilátka) CBP - calmodulin binding peptide. (26 AK) CBD - cellulose binding domain.")
12
TRANSFORMACE baktérii a kvasinek přímý přenos genetické informace z okolí do organismu
buňka schopná přijmout DNA (plasmid) se nazývá KOMPETENTNÍ přirozeně kompetentní jsou některé kmeny Bacillus subtilis, Hemorheae influenze atd. všechny ostatní se mohou transformovat po uvedení do kompetentního stavu dvěma způsoby: A) ELEKTROPORACE B) CHEMICKÁ METODA buňky se pořádně promyjí diH2O smíchají s plasmidovou DNA rozpuštěnou také v diH2O a vloží do elektroporátoru buňky se ošetří roztokem rubidné a vápenaté soli, které způsobují větší permeabilitu membrány DNA se smísí s těmito buňkami a provede se tzv. HEAT SHOCK (45 sec. 42°C) EFEKTIVITA NÁROČNOST
 se nazývá KOMPETENTNÍ. přirozeně kompetentní jsou některé kmeny Bacillus subtilis, Hemorheae influenze atd. všechny ostatní se mohou transformovat po uvedení do kompetentního stavu dvěma způsoby: A) ELEKTROPORACE B) CHEMICKÁ METODA. buňky se pořádně promyjí diH2O. smíchají s plasmidovou DNA rozpuštěnou. také v diH2O a vloží do elektroporátoru. buňky se ošetří roztokem rubidné a vápenaté soli, které způsobují větší permeabilitu membrány. DNA se smísí s těmito buňkami a provede se tzv. HEAT SHOCK (45 sec. 42°C) EFEKTIVITA NÁROČNOST ")
13
Který kmen E.coli zvolit?
tonA mutace chrání bakterii před napadením T1 a T5 fágem, chrání tak vaše klony lacZ.M15 částečná delece wild-typového lacZ genu, po vložení plasmidu dochází k tzv.α- komplementaci potřebné pro blue/white screening na miskách s X-gal endA1 deficience endonukleasy I zaručuje kvalitní izolaci plasmidové DNA lacIq produkuje lacZ represor negativně regulující transkripci z lacZ promotoru; zrušení přídavkem IPTG mcrA, mcrBC, a mrr mutace v těchto genech zaručuje možnost klonování i methylované genomové DNA recA1 zabraňuje rekombinaci mezi plasmidovou a bakteriální DNA F´ episom je potřebný pro produkci ssDNA kóduje protein tvořící tzv pilus na vnější membráně E.coli

14
regulace exprese pod T7 promotorem
exprese naklonovaného genu je kontrolována velice silným promotorem z bakteriofága T7, který původně řídí expresi genu 10 pro obalový protein pro expresi je nutno dodat do hostitelským buněk T7 RNA polymerasu a to buď infekcí bakteriofágem, nebo její indukovanou expresi. v sytému pCR®T7 TOPO® TA Expression je exprese T7 RNA polymerasy indukována lacZ promotorem pomocí IPTG a tento systém je uložen v genomu hostitelských buněk

15
kmeny E.coli vhodné pro expresi - BL21(DE3) nebo BL21(DE3)LysS
 nebo BL21(DE3)LysS")
16
před indukci IPTG probíhá bazální exprese T7 RNA polymerasy, pokud je exprimovaný produkt toxický pro bakterii, nedojde k selekci, selektují se pouze mutované klony, které neprodukují rekombinantní protein kmen E.coli BL21(DE3) nese v genomu T7 RNA polymerasový gen pod lacZ promotorem, tento konstrukt je vložen do genu pro integrasu, jehož inaktivaci se zabrání lyzi, vyštěpení fágové částice v nepřítomnosti pomocného fága. Přirozený lac represor, jehož gen je taktéž vložený genomu bakterie, brání expresi bez přítomnosti induktoru (IPTG) někdy ovšem i přesto dochází k bazální expresi T7 RNA polymerasy a pokud je pod T7 promoter vložen gen produkující toxický produkt pro E.coli může docházet k redukci růstu, smrti baktérie či nestabilitě plasmidu. Kmen BL21(DE3)LysS navíc obsahuje T7 lysozym (produkovaný genem LysS), uložený na speciálním vektoru s nízkou expresí a nezávislou selekci na chloramfenikol. T7 lysozym je schopen se vázat na T7 RNA polymerasu a inhibovat bazální transkripci, exprese indukovaná IPTG je daleko silnější a T7 RNA polymerasa se dostane z této inhibice T7 lysozym je bifunkční enzym, který má navíc vlastní lytickou funkci, naštěpuje bakteriální peptidoglykanovou stěnu a usnadňuje tak následnou izolaci exprimovaného proteinu.
 nese v genomu T7 RNA polymerasový gen pod lacZ promotorem, tento konstrukt je vložen do genu pro integrasu, jehož inaktivaci se zabrání lyzi, vyštěpení fágové částice v nepřítomnosti pomocného fága. Přirozený lac represor, jehož gen je taktéž vložený genomu bakterie, brání expresi bez přítomnosti induktoru (IPTG) někdy ovšem i přesto dochází k bazální expresi T7 RNA polymerasy a pokud je pod T7 promoter vložen gen produkující toxický produkt pro E.coli může docházet k redukci růstu, smrti baktérie či nestabilitě plasmidu. Kmen BL21(DE3)LysS navíc obsahuje T7 lysozym (produkovaný genem LysS), uložený na speciálním vektoru s nízkou expresí a nezávislou selekci na chloramfenikol. T7 lysozym je schopen se vázat na T7 RNA polymerasu a inhibovat bazální transkripci, exprese indukovaná IPTG je daleko silnější a T7 RNA polymerasa se dostane z této inhibice. T7 lysozym je bifunkční enzym, který má navíc vlastní lytickou funkci, naštěpuje bakteriální peptidoglykanovou stěnu a usnadňuje tak následnou izolaci exprimovaného proteinu.")
17
Jaké geny lze v E.coli exprimovat?
většinu z prokaryotických organismů eukaryotní geny jejichž produkty nepodléhají posttranslačním modifikacím většina cytosolárních proteinů (není glykosylovaná) geny kódované chloroplastovou nebo mitochondriální DNA (podobný genetický kód, evoluční příbuznost) všechny geny jejichž produkty nepotřebujeme v aktivní formě
 geny kódované chloroplastovou nebo mitochondriální DNA. (podobný genetický kód, evoluční příbuznost) všechny geny jejichž produkty nepotřebujeme v aktivní formě.")
18
odchylky v genetickém kódu? snižují výtěžek heterologní exprese
výjimky: jiná preference: kodón pro arginin (6 různých): CGU AGA CGA AGG CGG CGC E. coli Arabidopsis th. H. sapiens AGA 2.2% AGA 18.9% AGA 11.9% AGG 1.6% AGG 11.0% AGG 12.1%
: CGU AGA. CGA AGG. CGG. CGC. E. coli Arabidopsis th. H. sapiens. AGA 2.2% AGA 18.9% AGA 11.9% AGG 1.6% AGG 11.0% AGG 12.1%")
19
mutace místně cílená (site-directed mutagenesis)
Stratagene gen, či sekvenci kterou budeme chtít mutovat, je třeba naklonovat do vhodného vektoru navržení dvou komplementárních primerů, v místě kde chceme mutovat, nesoucí tuto mutaci M G A L L W L původní sekvence ’ ATG GGA GCT CTA TTA ACC TTA 3’ forward primer ’ TAC CCT CGA GAT AAT TCG AAT 5’ reverse primer ’ ATG GGA GCT CTA TTA AGC TTA 3’ M G A L L S L PCR s těmito primery na templátový plasmid a s Pfu polymerasou (se samoopravnou funkcí) vytváří se nové cirkulární DNA nesoucí mutaci, jsou k sobě komplementární a drží u sebe, mají přerušení v místě konce primerů (tzv. nick) ošetření restrikční endonukleasou DpnI (štěpí pouze methylovanou DNA, tedy templátový plasmid) transformace do bakterie a namnožení mutovaného plasmidu
 vytváří se nové cirkulární DNA nesoucí mutaci, jsou k sobě komplementární a drží u sebe, mají přerušení v místě konce primerů (tzv. nick) ošetření restrikční endonukleasou DpnI (štěpí pouze methylovanou DNA, tedy templátový plasmid) transformace do bakterie a namnožení mutovaného plasmidu.")
20
stabilizace exprimovaného proteinu
2004 SUMO peptide – Small Ubiquitin like MOdifier ochrana před proteolýzou zvyšování rozpustnosti proteinu zvyšuje množství exprimovaného proteinu Sumo proteasa

21
testování exprese - optimalizace
po transformaci expresního plasmidu do vhodných buněk se namnoží prvotní kultura, selektuje se na vhodném antibiotiku (1.den) prvotní kultura se pak vhodně naředí čistým médiem na OD – 0.5 (2.den) indukuje se exprese přídavekem IPTG ( mM) do kultury (2.den) kultura se inkubuje na třepačce (aerace) při 18-37°C (2.-5. den) a odebírají se vzorky ve kterých se detekuje exprimovaný protein. teplota: 25-18°C – pomalý růst, pomalá exprese, protein je vylučován do cytosolu 37°C - intenzivní růst, mohutná exprese, pokud je ale protein toxický pro bakterii (většina) je ukládán do tzv. INCLUSION BODIES mikrotělíska zůstávající v bakteriální cytoplasmě – ztížená izolace!!! E.coli nikdy nesekretuje protein do média!
 prvotní kultura se pak vhodně naředí čistým médiem na OD – 0.5 (2.den) indukuje se exprese přídavekem IPTG ( mM) do kultury (2.den) kultura se inkubuje na třepačce (aerace) při 18-37°C (2.-5. den) a odebírají se vzorky ve kterých se detekuje exprimovaný protein. teplota: 25-18°C – pomalý růst, pomalá exprese, protein je vylučován do cytosolu. 37°C - intenzivní růst, mohutná exprese, pokud je ale protein toxický pro bakterii (většina) je ukládán do tzv. INCLUSION BODIES mikrotělíska zůstávající v bakteriální cytoplasmě – ztížená izolace!!! E.coli nikdy nesekretuje protein do média!")
22
izolace proteinu z bakteriální kultury:
pokud je protein ukládán do inkluzních tělisek, rozbití buněk tepelným šokem, případně sonifikací nebo lysozymem pokud je ukládán do inkluzních tělísek, oddělení nerozpustné frakce a denaturace 9M močovinou nebo guanidium chloridem poté je nutno protein renaturovat - SLOŽITÉ!!!! imidazol bakt.extrakt marker 0.5mM 200mM 100mM wash II wash I IB IB 1M

23
exprese proteinů ve velkém měřítku: FERMENTORY
regulace: teplota pH obsah kyslíku (případně jiných plynů) přesné dávkování
 přesné dávkování.")
24
problémy a nedostatky exprese v E.coli
příčina řešení buňky umírají, nedaří se selekce toxický produkt, vysoká bazální exprese slabší promotor, kontrola bazální exprese, snížení teploty kultivace nerozpustný produkt (ukládá se do inkluzních tělísek) redukce disulfidických můstků v redukčním prostředí cytoplasmy transport do periplasmy snížení teploty snížení exprese fúze s hydrofilní značkou neaktivní protein redukce v cytoplasmě afinitní značka ovlivňuje aktivitu změna typu a polohy značky zvýšení osmotického tlaku média (1M sorbitol) žádný protein preference jiného genetického kódu dodání raritních tRNA silnější promotor zvýšení počtu kopií plasmidů
 redukce disulfidických můstků v redukčním prostředí cytoplasmy. transport do periplasmy. snížení teploty. snížení exprese. fúze s hydrofilní značkou. neaktivní protein. redukce v cytoplasmě. afinitní značka ovlivňuje aktivitu. změna typu a polohy značky. zvýšení osmotického tlaku média (1M sorbitol) žádný protein. preference jiného genetického kódu. dodání raritních tRNA. silnější promotor. zvýšení počtu kopií plasmidů.")
25
Bacillus subtilis alternativní prokaryotické expresní systémy
gram pozitivní půdní bakterie není lidský patogen má vyvinutý sekreční systém neprodukuje žádné endotoxiny (rek. proteiny se dají využít v medicíně)
")
26
Bacillus subtilis alternativní prokaryotické expresní systémy
laboratorní a průmyslově využívané kmeny B.subtilis mají tyto mutace: delece genu produkujícího tenzidy (sfrC) delece genu produkujícího červený pigment delece genu pro exogenní proteasy
 delece genu produkujícího červený pigment. delece genu pro exogenní proteasy.")
27
Bacillus subtilis alternativní prokaryotické expresní systémy
Bacillus subtilis je přirozeně kompetentní (má systém přenášející DNA přes buněčnou membránu do buňky)
")
28
integrace do genomu pomocí homologní rekombinace
amyE gen kóduje neesenciální alfa-amylasu

29
Bacillus subtilis alternativní prokaryotické expresní systémy
využití pro průmyslovou produkci proteas (prací prášky) a amylas (sladovnictví) a hlavně průmyslově nejdůležitější zdroj kyseliny hyaluronové (polysacharid)
 a amylas (sladovnictví) a hlavně průmyslově nejdůležitější zdroj kyseliny hyaluronové (polysacharid)")
30
operon genů pro syntézu hyaluronové kyseliny z rodu Streptococcus vklonován do genomu Bacillus

31
silný sekreční mechanismus, žádné kontaminující proteiny Streptomyces lividans - půdní bakterie Caulobacter crescentus - vodní baktérie Staphylococcus carnosus - nepatogenní produkce proteinů se zabudovanými radioisotopy 13C, 15N a deuteriem Anabaena sp. - sinice

32
využití rekombinantních bakterií v potravinářství
Biotech chymosin enzym používaný pro srážení mléka v sýr kvasinkový gen transformovaný do bakterie biotechnologie nahrazuje chymosin izolovaný z poražených telat bST (bovine somatotropin) hormon zvyšující u krav produkci mléka gen z genomu krávy naklonován do baktérie přidává se do krmiva v kravínech dříve se používal hormon pracně izolovaný z hypofýzy poražených krav Source: Rent Mother Nature
 hormon zvyšující u krav produkci mléka. gen z genomu krávy naklonován do baktérie. přidává se do krmiva v kravínech. dříve se používal hormon pracně izolovaný z hypofýzy poražených krav. Source: Rent Mother Nature.")
33
využití rekombinantních bakterií v potravinářství

34
využití rekombinantních bakterií ve farmacii lidský inzulín (Humulin®)
2002 – 250 miliónů lidí využívalo léčiv a vakcín produkovaných mikroorganismy lidský inzulín (Humulin®) inzulín – polypeptid (51 ak) inzulín pro léčbu diabetiků byl extrahován z pankreasu prasat a krav prasečí inzulín se liší pouze dvěmi ak někteří diabetici však produkovali protilátky proti živočišnému insulinu lidský inzulín se začal syntetizovat uměle (drahé) 1982 poprvé připraven pomocí rDNA technologie od devadesátých let se produkuje ve velkém a levně lidský insulin pomocí transgenních E.coli nebo kvasinek (např. Humulin®)
 inzulín – polypeptid (51 ak) inzulín pro léčbu diabetiků byl extrahován z pankreasu prasat a krav. prasečí inzulín se liší pouze dvěmi ak. někteří diabetici však produkovali protilátky proti živočišnému insulinu. lidský inzulín se začal syntetizovat uměle (drahé) 1982 poprvé připraven pomocí rDNA technologie. od devadesátých let se produkuje ve velkém a levně lidský insulin pomocí transgenních E.coli nebo kvasinek (např. Humulin®)")
35
využití rekombinantních bakterií ve farmacii
lidský růstový hormon (HGH) produkovaný hypofýzou je důležitý regulátor vývoje člověka děti s vrozenou deficienci genu pro HGH trpí dwarfismem (zakrslostí) pravidelné injekce toho hormonu mohou obnovit normální růst živočišný GH je pro léčbu značně neúčinný HGH se také izoloval z hypofýz lidských mrtvol byl zaznamenán zvýšeny výskyt Creutzfeldt-Jakobsovy choroby (kopurifikace prionu) velice drahé a velká spotřeba mrtvol (např. na izolaci 5 mg hormonu je třeba půl milionů jehněčích mozku) od devadesátých let produkován pomocí rekombinantních bakterií litr bakteriální kultury vyprodukuje 5 mg GH za 15 hodin
 produkovaný hypofýzou je důležitý regulátor vývoje člověka. děti s vrozenou deficienci genu pro HGH trpí dwarfismem (zakrslostí) pravidelné injekce toho hormonu mohou obnovit normální růst. živočišný GH je pro léčbu značně neúčinný. HGH se také izoloval z hypofýz lidských mrtvol. byl zaznamenán zvýšeny výskyt Creutzfeldt-Jakobsovy choroby (kopurifikace prionu) velice drahé a velká spotřeba mrtvol (např. na izolaci 5 mg hormonu je třeba půl milionů jehněčích mozku) od devadesátých let produkován pomocí rekombinantních bakterií. litr bakteriální kultury vyprodukuje 5 mg GH za 15 hodin.")
36
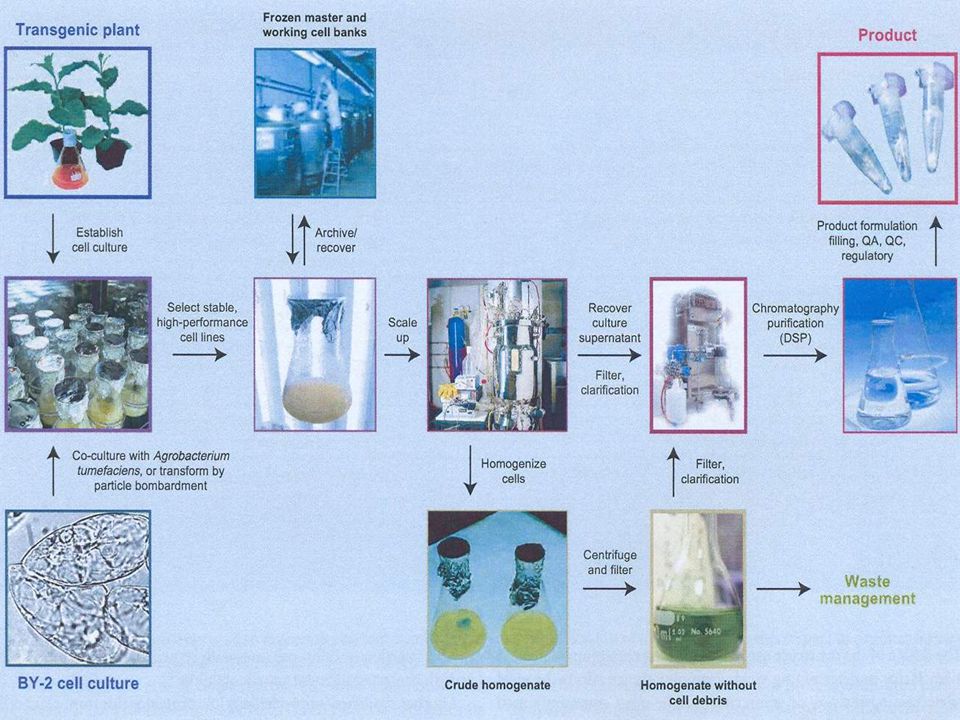
exprese v rostlinách BY-2 tobacco cells
rostlinné buněčné linie se pro produkci látek nepoužívají BY-2 tobacco cells odvozené z dřeně Nicotiana tabacum cv. Bright Yellow-2 v Japonsku (1974) rychle rostou (12 hod.), netvoří kalusy snadno absorbují různé sloučeniny – studium metabolismu snadná synchronizace – studium buněčného cyklu snadná transformace Agrobacteriem
 rychle rostou (12 hod.), netvoří kalusy. snadno absorbují různé sloučeniny – studium metabolismu. snadná synchronizace – studium buněčného cyklu. snadná transformace Agrobacteriem.")
38
TAXOL Hellwig S. et al. Plant cell cultures for the production of recombinant proteins. Nature Biotechnology 22(11), 2004,
, 2004,")
39
molekulární farmářství
rostlinné – jedlé vakcíny jako transgeny se používají oslabené toxiny původců mnohých chorob stimulace mukózního imunitního systému v epitelu trávícího traktu (účinná imunizace) velice levné a efektivní značné uplatnění v rozvojových zemích, pojídáním čerstvých plodů se získá imunita vůči nemocem, které v těchto zemích způsobují milióny úmrtí. vše ve formě testů v rostlinách tabáku a bramboru – zatím úspěšné zubní kaz – protein spaA ze Streptococcus mutans hepatitida B – povrchový antigen HBsAg viru cholera – termolabilní enterotoxin B z Vibrio cholerae 1 g brambor vyrobí 30µg tohoto toxinu brambory se uvaří a toxin denaturuje testy na myši prokázaly vysokou účinnost a neškodnost 1997 poprvé testováno na člověku hlavní snaha vědců je produkce těchto vakcín v plodinách rozšířených ve třetím světě jako je banánovník
 velice levné a efektivní. značné uplatnění v rozvojových zemích, pojídáním čerstvých plodů se získá imunita vůči nemocem, které v těchto zemích způsobují milióny úmrtí. vše ve formě testů v rostlinách tabáku a bramboru – zatím úspěšné. zubní kaz – protein spaA ze Streptococcus mutans. hepatitida B – povrchový antigen HBsAg viru. cholera – termolabilní enterotoxin B z Vibrio cholerae. 1 g brambor vyrobí 30µg tohoto toxinu brambory se uvaří a toxin denaturuje testy na myši prokázaly vysokou účinnost a neškodnost poprvé testováno na člověku. hlavní snaha vědců je produkce těchto vakcín v plodinách rozšířených ve třetím světě jako je banánovník.")
40
molekulární farmářství
plantibodies v rostlinách připravené živočišné protilátky nebo části protilátek v rostlinách lze produkovat i monoklonální protilátky protilátky jsou vylučovány do mezibuněčného prostoru mohou sloužit přímo v rostlině (jako ochrana proti patogenům) nebo mohou být extrahovány a použity v diagnostice či medicíně výrazně se sníží náklady a čas na tvorbu protilátek (hybridomové buňky)
 nebo mohou být extrahovány a použity v diagnostice či medicíně. výrazně se sníží náklady a čas na tvorbu protilátek (hybridomové buňky)")
41
molekulární farmářství
plantibodies hlavní problém – rozdíly v N-glykosylaci proteinů (protilátek) u živočichů a rostlin plantibodies mají jinak glykosylovanou strukturu a po aplikaci do zvířete vyvolávají nechtěnou imunitní odpověď řešení: společně s geny pro Ab je do rostliny vnesen gen pro lidskou β-1,4-galaktosyltransferasu – rostlina pak produkovala „polidštěné protilátky“ jako transgen stačí vložit pouze malou variabilní oblast tzv. Fv domény z lehkého a těžkého řetězce spojenou krátkým peptidem scFv – single chain variable fragment
 u živočichů a rostlin. plantibodies mají jinak glykosylovanou strukturu a po aplikaci do zvířete vyvolávají nechtěnou imunitní odpověď. řešení: společně s geny pro Ab je do rostliny vnesen gen pro lidskou β-1,4-galaktosyltransferasu – rostlina pak produkovala „polidštěné protilátky jako transgen stačí vložit pouze malou variabilní oblast tzv. Fv domény z lehkého a těžkého řetězce spojenou krátkým peptidem. scFv – single chain variable fragment.")
42
non-hodgkinský lymfom – rakovina lymfatických uzlin (B-lymfocytů)
nádorové buňky produkují specifické protilátky (zachycené na svém povrchu, liší se od zdravých) tabák infikován TMV s vloženou části genu pro scFV myši tabák produkuje scFv nádorových buněk
 tabák infikován TMV s vloženou části genu pro scFV myši. tabák produkuje scFv nádorových buněk.")
43
tabák produkuje funkční antigen, který po aplikaci myši produkuje protilátky proti nádorovým buňkám
imunitní odpověď nevznikla na mezidruhovou odlišnost „ produkt rostliny – odlišná glykosylace, špatně ustřižený signální peptid 80% myší přežilo antigen vytvořen během 6 týdnů velice jednoduchá izolace – RYCHLE A LEVNĚ léčba (vakcína) dělaná pacientovi přímo na míru
 dělaná pacientovi přímo na míru.")
44
v současné době klinicky testované plantibodies
antigen rostlina typ rekomb. Protilátky aplikace povrchový antigen Streptokoka tabák sekreční IgA CaroRx™ terapeutická povrchová aplikace Herpes simplex virus sója, rýže IgG sperma kukuřice antikoncepce ve formě gelu non-Hodginský lymfom scFv personalizovaná vakcína virus vztekliny intravenosně CEA - cancer embryonic antigen tabák, rýže, pšenice, rajče scFv, diabodies, chimerické protilátky diagnostická Stoger et al., Recent progress in plantibody technology, Curr. Pharm Design 11, 2005

45
molekulární farmářství biodegradovatelné plasty
některé bakterie jsou schopny uchovávat uhlík a energii ve formě osmoticky inertních polymerů (polyester k. hydroxymaselné PHB) tyto jsou pak v přírodě rychle degradovatelné tři geny z baktérií byly vloženy do rostlin Arabidopsis 3-ketotiolasa NADH-dep. acetoacetyl-CoA-reduktasa PHB-polymerasa syntéza PHB navazuje na syntézu mastných kyselin, jejíž část probíhá v chloroplastech pokud byly tyto geny vloženy do jaderného genomu pod 35S promotorem docházelo k nadměrné produkci polyhydroxybutyrátu v listech (PHB tvořil 0.14% sušiny) pokud byly produkty transgenů cíleny pomocí signálních peptidů do chloroplastů zvýšila se produkce až 100x
 tyto jsou pak v přírodě rychle degradovatelné. tři geny z baktérií byly vloženy do rostlin Arabidopsis. 3-ketotiolasa. NADH-dep. acetoacetyl-CoA-reduktasa. PHB-polymerasa. syntéza PHB navazuje na syntézu mastných kyselin, jejíž část probíhá v chloroplastech. pokud byly tyto geny vloženy do jaderného genomu pod 35S promotorem docházelo k nadměrné produkci polyhydroxybutyrátu v listech (PHB tvořil 0.14% sušiny) pokud byly produkty transgenů cíleny pomocí signálních peptidů do chloroplastů zvýšila se produkce až 100x.")
47
molekulární farmářství biodegradovatelné plasty
Arabidopsis slouží jako modelová rostlina předpokládá se že, tento model bude přenesen do nějaké rostliny, která produkuje velké hlízy nebo semena a není konzumována ani člověkem ani zvířaty (skočec Ricinus communis) transgenní řepka vyprodukuje až 10 g PHB na rostlinu fa MONSANTO zatím tento projekt pozastavila, jelikož to stále ještě není ekonomicky výhodné (výroba plastů z fosilních zdrojů je daleko levnější)
 transgenní řepka vyprodukuje až 10 g PHB na rostlinu. fa MONSANTO zatím tento projekt pozastavila, jelikož to stále ještě není ekonomicky výhodné (výroba plastů z fosilních zdrojů je daleko levnější)")
48
7-metylxantin N-metyltransferasa a theobromin N-metyltransferasa
další využívané transgeny bezkofeinová káva 7-metylxantin N-metyltransferasa a theobromin N-metyltransferasa JAPONSKO 2003 exprese genů inhibována metodou RNAi druh Cofea canephora transformován Agrobaktériem transformované rostliny stejné jako wild type Cofea canephora se pěstuje na Madagaskaru a má 2x větší obsah kofeinu než nejrozšířenější Cofea arabica v mladých listech: theobromin % kofein %

49
YEAST TWO HYBRID SYSTEM
HYBRIDNÍ EXPRESNÍ KNIHOVNY systém pro studium interakce protein - protein, bait vektor (návnada) obsahuje gen pro protein, který studujeme a fish (prey) vektor (obsahuje danou cDNA expresní knihovnu) PRO STUDIUM PROTEIN-PROTEIN INTERAKCÍ – význam pro studium regulačních a signálních drah naklonovaná cDNA knihovna z libovolného euk.organismu konstitutivní promotor kvasinkový chromozom
 obsahuje gen pro protein, který studujeme a fish (prey) vektor (obsahuje danou cDNA expresní knihovnu) PRO STUDIUM PROTEIN-PROTEIN INTERAKCÍ – význam pro studium regulačních a signálních drah. naklonovaná. cDNA. knihovna. z libovolného. euk.organismu. konstitutivní. promotor. kvasinkový. chromozom.")
50
oddělení molekulární biologie katedry biochemie, PřF UP
různé expresní vektory s HIS-tagy na C a N konci pro expresi v E.coli různé expresní kmeny E.coli expresní vektory s HIS-tagy na C a N konci odvozené ze systému pPICZ a pGAPZ (Invitrogen) pro expresi v Pichia pastoris expresní vektory pro expresi v Saccharomyces cerevisiae – replikační i pro integraci expresní kmeny kvasinek Pichia pastoris a Saccharomyces cerevisiae různé vektory pro transformaci jednoděložných i dvouděložných rostlin
 pro expresi v Pichia pastoris. expresní vektory pro expresi v Saccharomyces cerevisiae – replikační i pro integraci. expresní kmeny kvasinek Pichia pastoris a Saccharomyces cerevisiae. různé vektory pro transformaci jednoděložných i dvouděložných rostlin.")
Podobné prezentace









 Nespecifická : Fagocytóza granulocytů,monocytů.>")










