Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
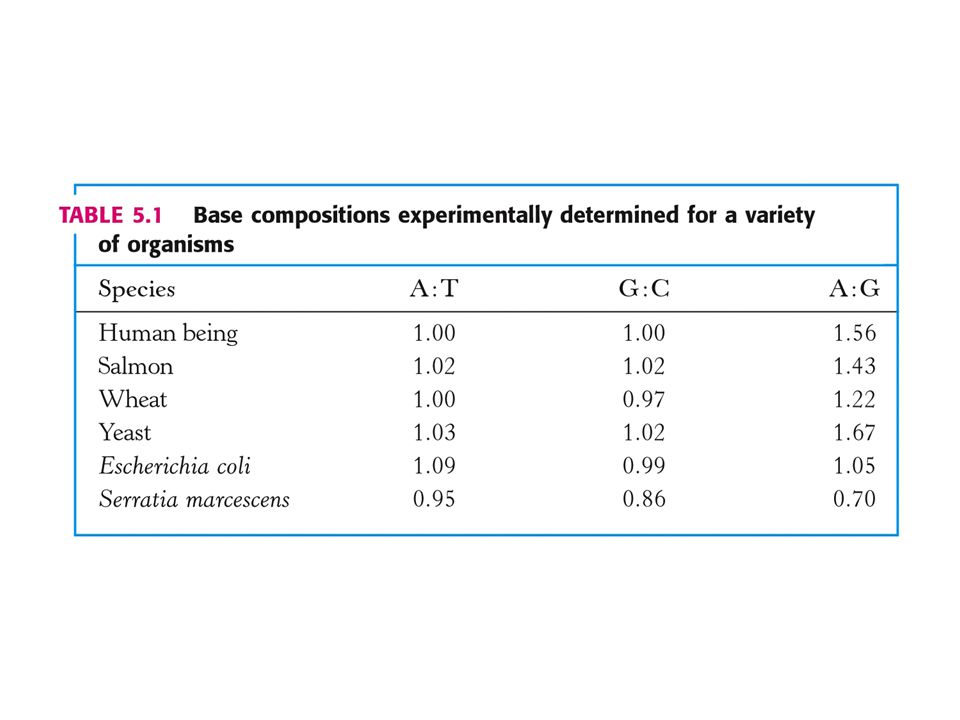
DNA, RNA – genetická informace.
1 Složení a struktura nukleových kyselin. 2. Dvojitá helikální struktura 3. Replikace DNA – polymerasy 4. Exprese genů – přenos informací uložených v DNA na funkční molekuly (proteosyntéza) 5. Kódování aminokyselin – triplety bází 6. Eukaryotní geny
 5. Kódování aminokyselin – triplety bází. 6. Eukaryotní geny.")
2
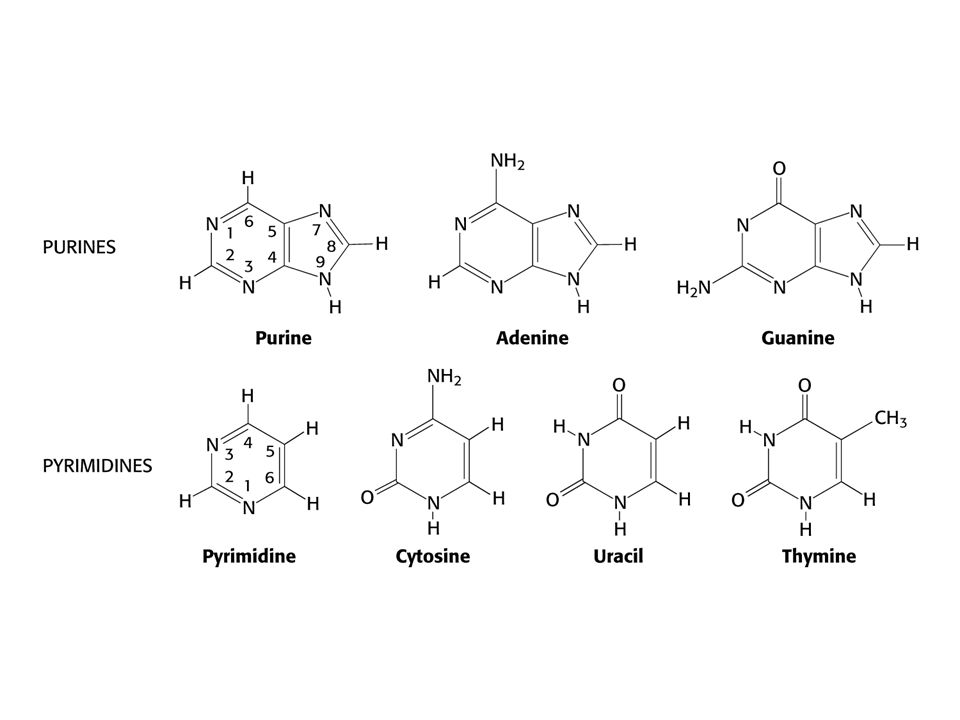
Polymerní struktura nukleových kyselin

3
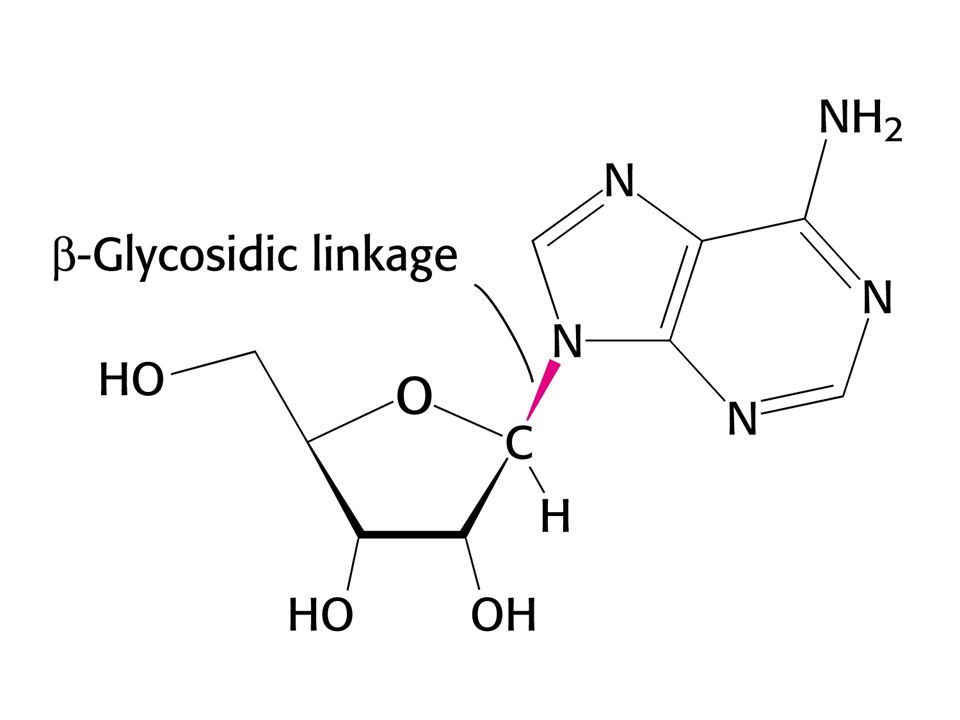
Ribosa a deoxyribosa

10
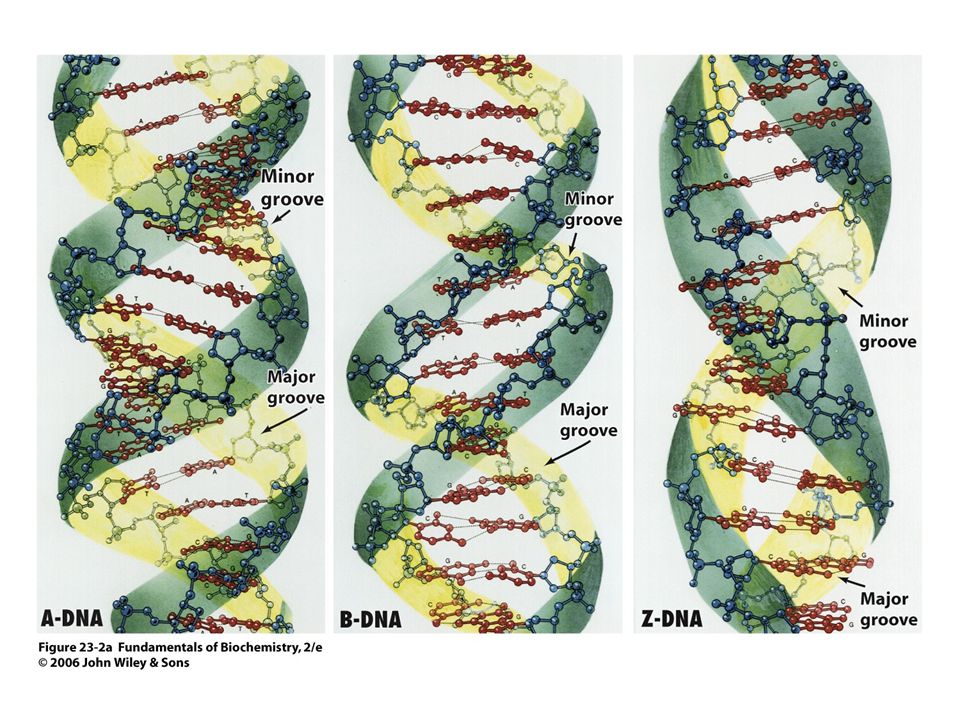
Struktura řetězce DNA

11
Rentgenový difrakční snímek hydratované DNA
Rentgenový difrakční snímek hydratované DNA. Kříž uprostřed je znakem helikální struktury. Tmavé obloučky jsou naskládané nukleotidové báze

12
Watson-Crickův model dvojité helix DNA. Axiální pohled.
Jeden polynukleotidový řetězec je modrý a druhý červený. Purinové a pyrimidinové báze jsou světlejší. Struktura se opakuje (axiálně) po 34 Ă, což odpovídá 10 nukleotidům.
 po 34 Ă, což odpovídá 10 nukleotidům.")
13
Pohled shora na DNA – radiálně.

14
Prvky Watson-Crickova modelu DNA dedukované z difrakčních dat:
a) Dva helikální polynukleotidové řetězce se obtáčí kolem společné osy. Řetězce se vinou protisměrně. b) Řetězec fosfátů a sacharidů leží na vnější straně, purinové a pyrimidinové báze směřují dovnitř helixu. c) Báze jsou téměř kolmé k helikální ose a jsou vzájemně vzdáleny o 3, 4 A . Helikální struktura se opakuje po deseti bázích (otočka 34 A). Rotace je 36o na bázi = 360o na otočku. d) Průměr helixu je 20 A.
 Dva helikální polynukleotidové řetězce se obtáčí kolem společné osy. Řetězce se vinou protisměrně. b) Řetězec fosfátů a sacharidů leží na vnější straně, purinové a pyrimidinové báze směřují dovnitř helixu. c) Báze jsou téměř kolmé k helikální ose a jsou vzájemně vzdáleny o 3, 4 A . Helikální struktura se opakuje po deseti bázích (otočka 34 A). Rotace je 36o na bázi = 360o na otočku. d) Průměr helixu je 20 A.")
19
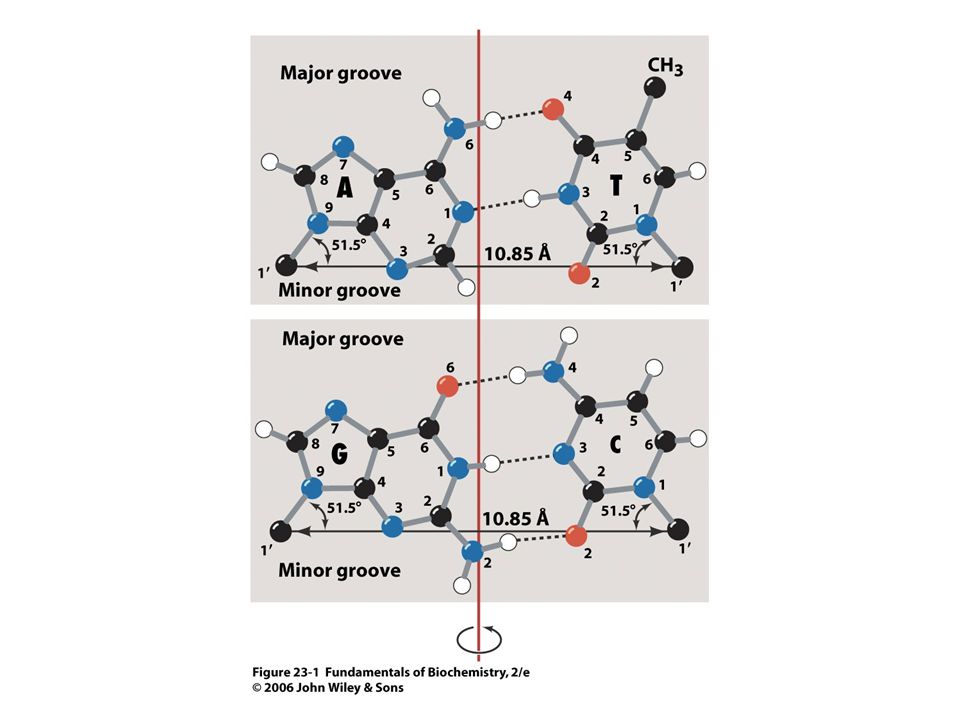
Struktura párů bazí dle Watsona a Cricka

21
Axiální pohled na DNA. Páry bazí jsou na sebe naskládány.

22
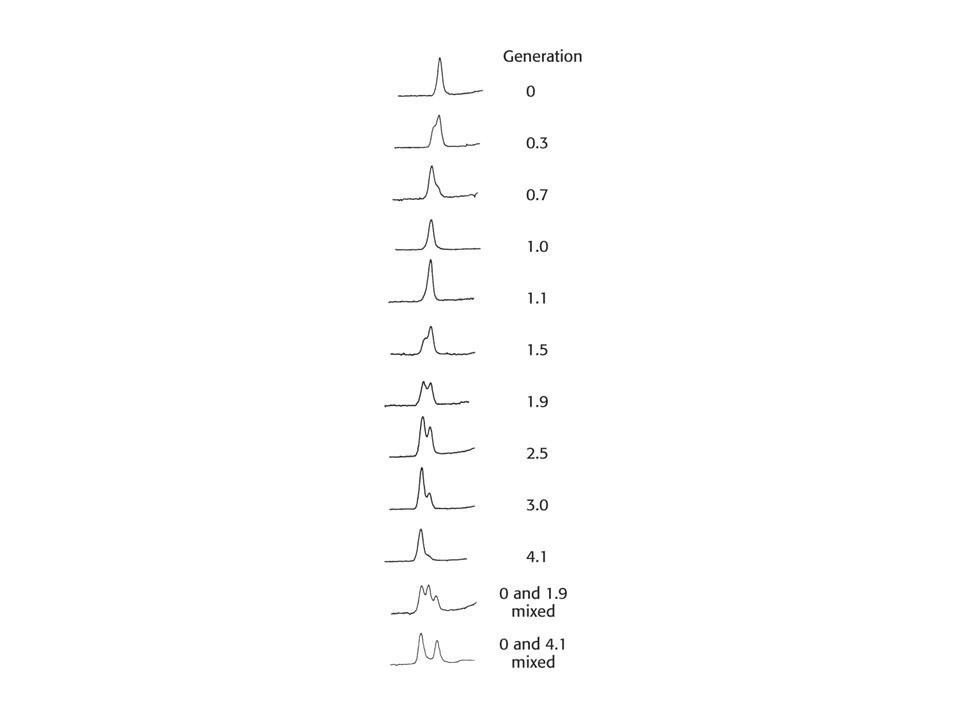
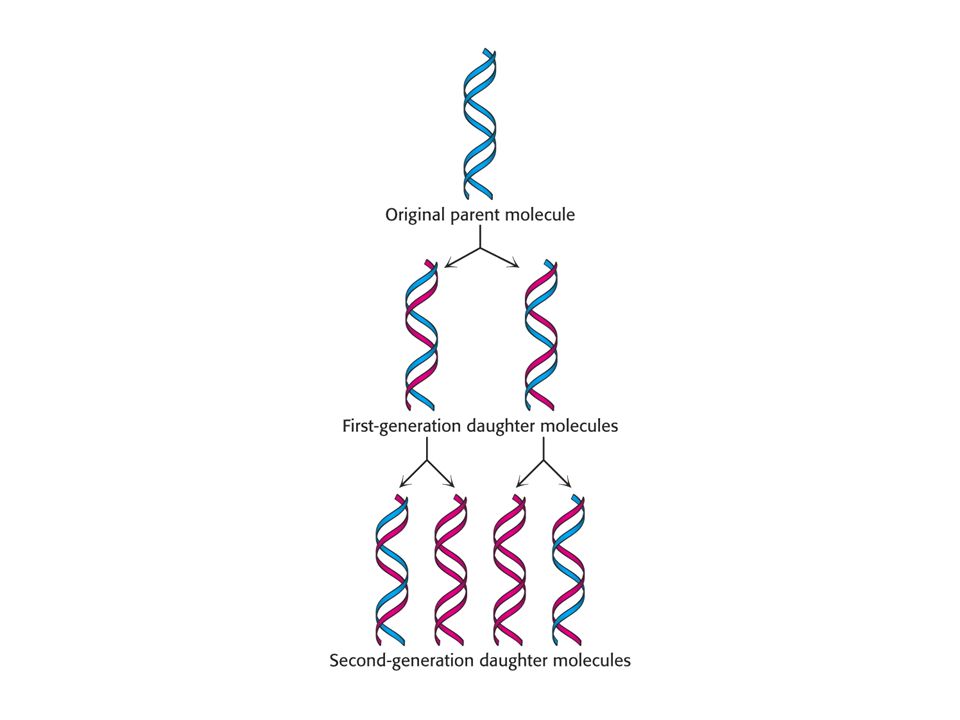
Semikonzervativní replikace DNA – pokus M. Messelsona a F
Semikonzervativní replikace DNA – pokus M. Messelsona a F. Stahla (1958). Hypotéza: oddělením obou řetězců vzniknou dva jednovláknové templáty na kterých se syntetizuje systémem párování bazí stejná dvoušroubovice jako rodičovská. Označili rodičovskou DNA 15N (těžký izotop dusíku). Získali je růstem E. coli na médiu, kde jediným zdrojem dusíku byl 15NH4Cl. Poté byly baktérie rychle převedy do média 14N. Otázka: Jaké bude rozdělení 14N a 15N v molekulách DNA po úspěšném kole replikace ?
. Hypotéza: oddělením obou řetězců vzniknou dva jednovláknové templáty na kterých se syntetizuje systémem párování bazí stejná dvoušroubovice jako rodičovská. Označili rodičovskou DNA 15N (těžký izotop dusíku). Získali je růstem E. coli na médiu, kde jediným zdrojem dusíku byl 15NH4Cl. Poté byly baktérie rychle převedy do média 14N. Otázka: Jaké bude rozdělení 14N a 15N. v molekulách DNA po úspěšném kole replikace")
23
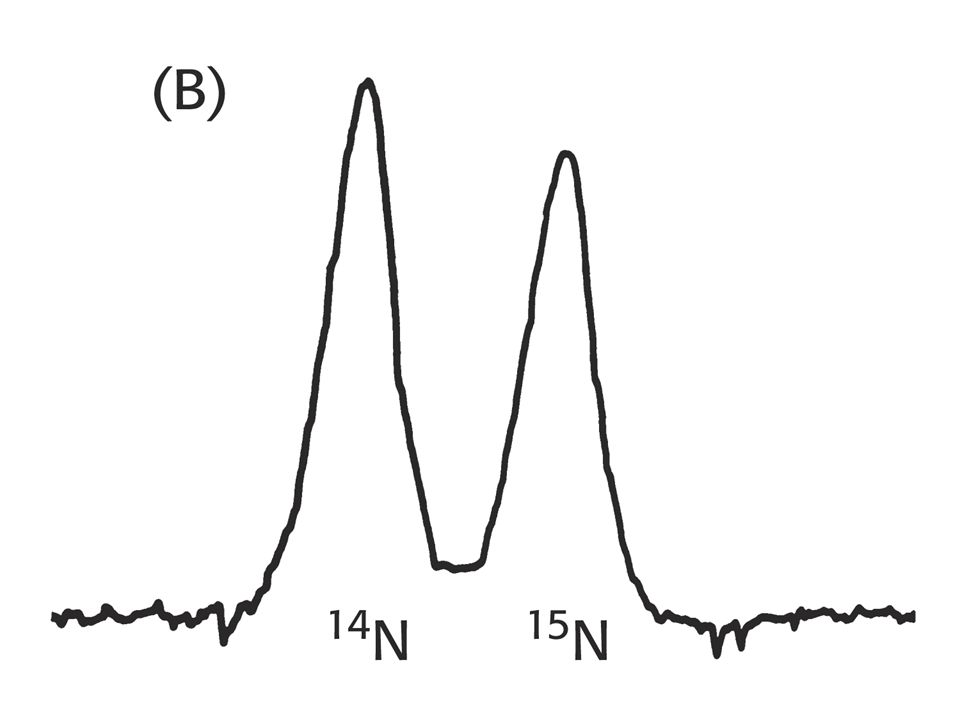
Rozdělení izotopů dusíku bylo řešeno pomocí techniky „sedimentace v rovnovážném hustotním gradientu“. Sedimentace se uskutečnila v hustotním gradientu CsCl, který má stejnou hustotu jako DNA (1, 7 g/cm3 - rozsah gradientu 1, 66 až 1, 76 g/cm3). Molekuly DNA se při centrifugaci shromáždily v místě stejné hustoty jakou mají ony samy. Genomová DNA tvoří proužek (detekce UV světlem). Směs obou DNA (lehké a těžké dává jasně oddělené proužky, přestože se liší hustotou o 1 %. DNA byla extrahována z baktérie v různých časových intervalech.
. Molekuly DNA se při centrifugaci shromáždily v místě stejné hustoty jakou mají ony samy. Genomová DNA tvoří proužek (detekce UV světlem). Směs obou DNA (lehké a těžké dává jasně oddělené proužky, přestože se liší hustotou. o 1 %. DNA byla extrahována z baktérie v různých časových intervalech.")
26
Důkaz semikonzervativní replikace
Důkaz semikonzervativní replikace. Po první generaci, byly všechny molekuly DNA hybridy obsahující stejné množství obou izotopů dusíku.

29
Hypochromismus. Jednovláknová DNA absorbuje světlo efektivněji než dvojvláknová. Absorbance DNA při 260 nm vzroste, když se dvojváknová DNA podrobí tavení.

30
Teplota tání DNA (Tm) je teplota, při které se polovina dvojvláknové DNA rozpojí.
 je teplota, při které se polovina dvojvláknové DNA rozpojí.")
31
Elektronový snímek kruhové mitochondriální DNA. Relaxovaná molekula.

32
Supercoiled form (spirála tvořená propletenými spirálami DNA)
Supercoiled form (spirála tvořená propletenými spirálami DNA). Výhody: a) Kompaktnější tvar, b) zabraňuje interakcím mezi DNA a dalšími molekulami.
. Výhody: a) Kompaktnější tvar, b) zabraňuje interakcím mezi DNA a dalšími molekulami.")
33
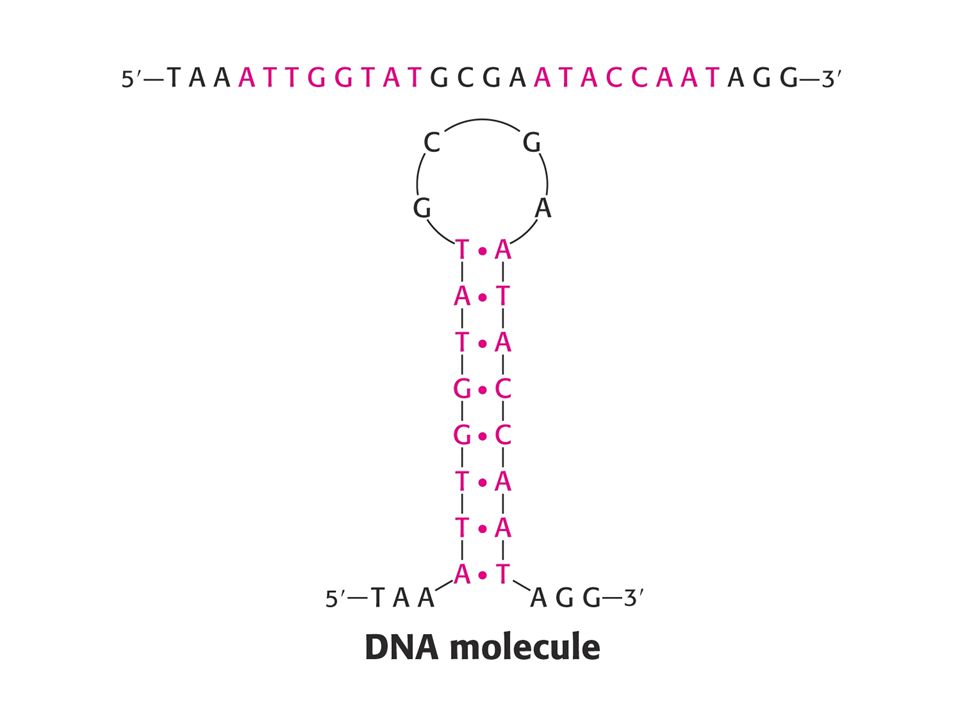
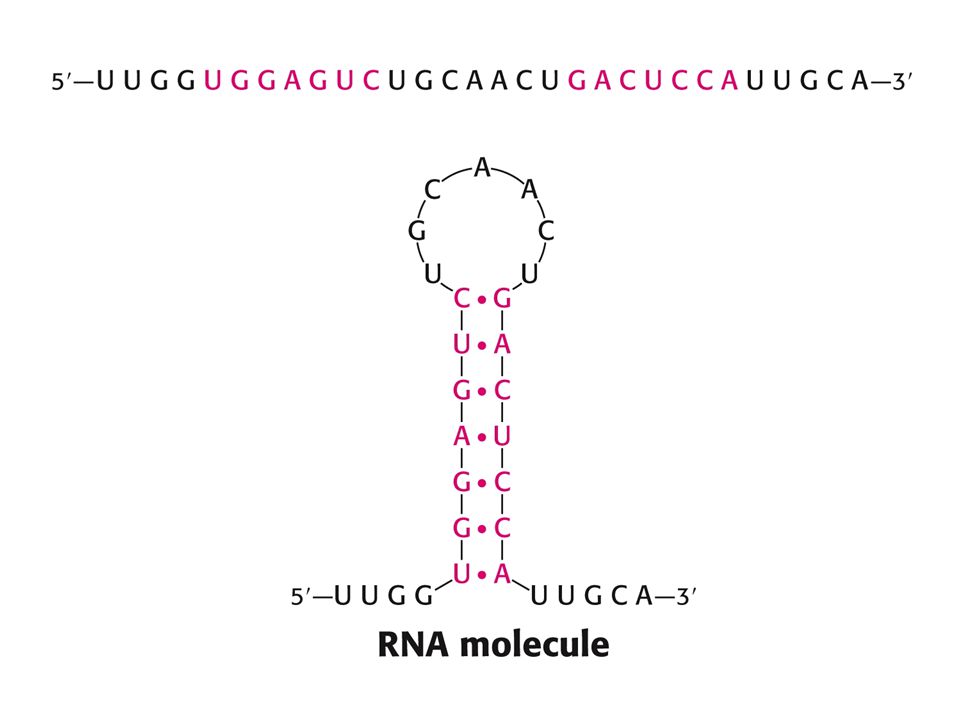
Struktura nukleových kyselin – stonek se smyčkou. (Stem-loop)
")
36
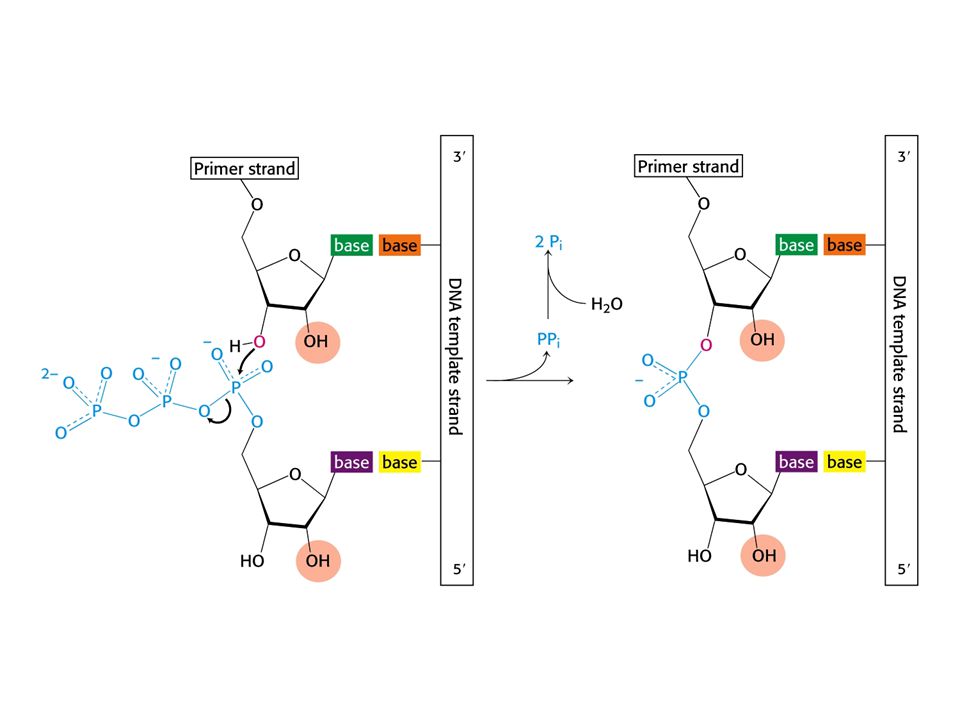
DNApolymerasa – katalyzuje krok po kroku připojení deoxyribonukleotidu k řetězci DNA (nutný DNA templát)
")
37
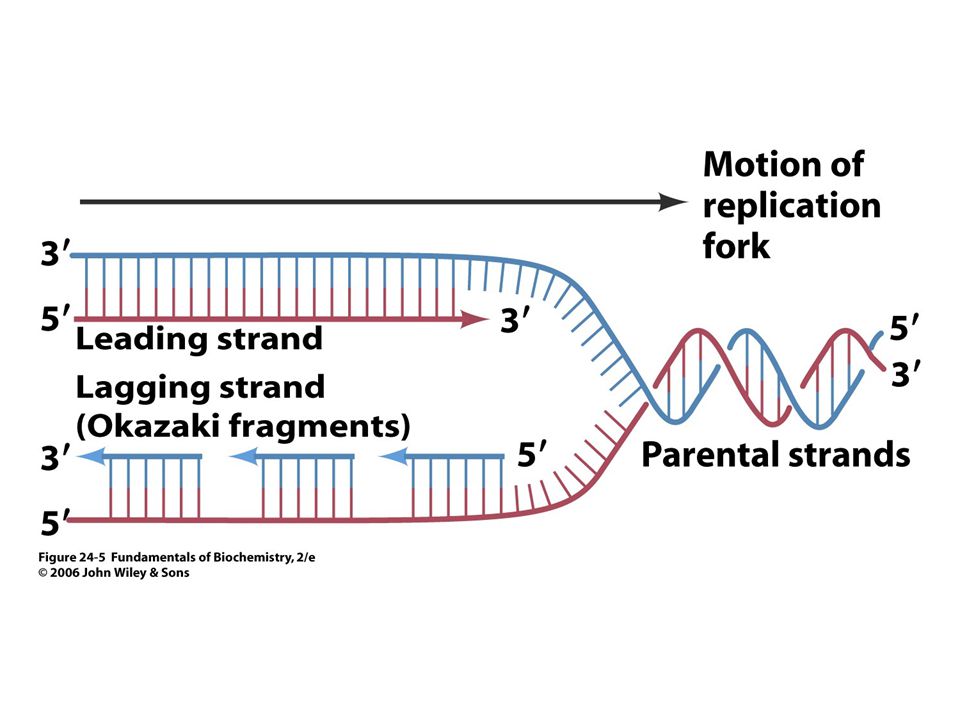
Replikace DNA katalyzovaná DNApolymerasou.

39
Tok informace z RNA na DNA u retrovirů
Tok informace z RNA na DNA u retrovirů. Genom RNA retroviru je převeden na DNA enzymem reverzní transkriptasa, který je do buňky vnesen virem.

40
Typy RNA. a) Messenger RNA(mRNA)(informační)-templát při syntéze proteinů nebo traslokaci. U E.coli je produkována mRNA pro každý gen nebo skupinu genů. U eukaryot je produkována mRNA pro každý gen. mRNA je heterogenní třída molekul. Délka u E. coli 1, 2 kilobazí (kb) b) Transfer RNA (tRNA) – přenáší aminokyseliny v aktivované formě na ribosomy, kde dochází ke tvorbě peptidového řetězce. Každá z dvaceti proteinogenních aminokyselin má svoji tRNA. tRNA obsahuje asi 75 nukleotidů, hmotnost cca 25 kd. c) Ribosomální RNA (rRNA) hlavní složka ribosomů hrající roli strukturní i katalytickou při syntéze proteinů. V E.coli jsou tři typy rRNA dělené podle sedimentačního koeficientu: 23S, 16S a 5S. Každá z těchto rRNA je v každém ribosomu.
 Messenger RNA(mRNA)(informační)-templát při syntéze proteinů nebo traslokaci. U E.coli je produkována mRNA pro každý gen nebo skupinu genů. U eukaryot je produkována mRNA pro každý gen. mRNA je heterogenní třída molekul. Délka u E. coli 1, 2 kilobazí (kb) b) Transfer RNA (tRNA) – přenáší aminokyseliny v aktivované formě na ribosomy, kde dochází ke tvorbě peptidového řetězce. Každá z dvaceti proteinogenních aminokyselin má svoji tRNA. tRNA obsahuje asi 75 nukleotidů, hmotnost cca 25 kd. c) Ribosomální RNA (rRNA) hlavní složka ribosomů hrající roli strukturní i katalytickou při syntéze proteinů. V E.coli jsou tři typy rRNA dělené podle sedimentačního koeficientu: 23S, 16S a 5S. Každá z těchto rRNA je v každém ribosomu.")
41
Eukaryotní buňky obsahují ještě další malé RNA (small nuclear RNA, snRNA). Např. v cytoplasmě se uplaňují při vnášení nově syntetizovaných proteinů do intracelulárních a extracelulárních prostorů (kompartmentů). Všechny RNA jsou syntetizovány RNApolymerasou. Syntéza RNA z DNA templátu se nazývá transkripce.

42
Komponenty katalýzy RNApolymerasy:
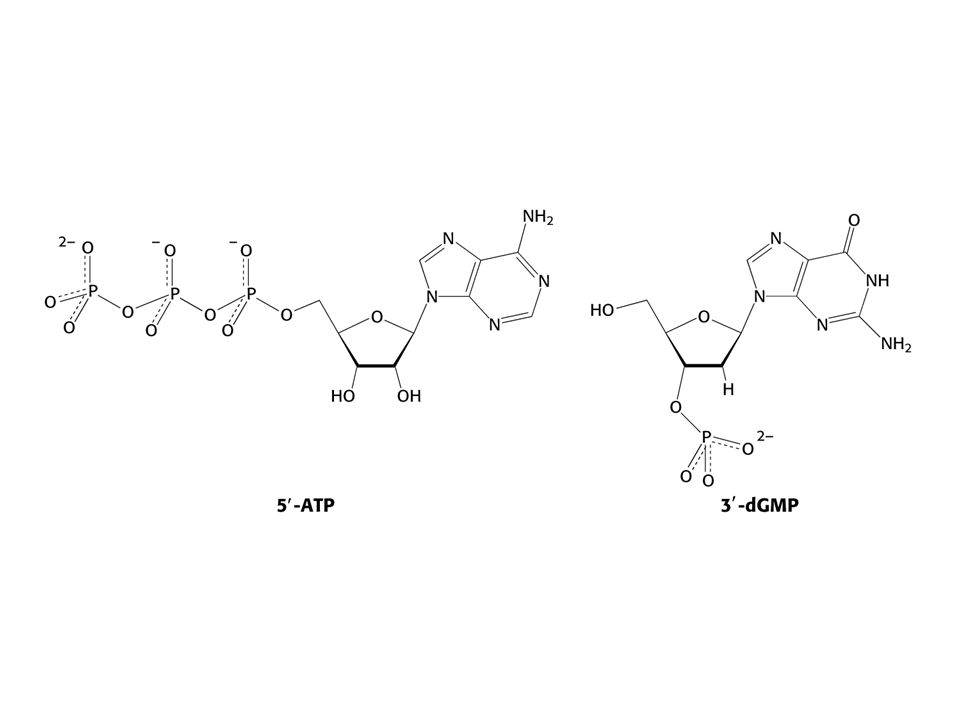
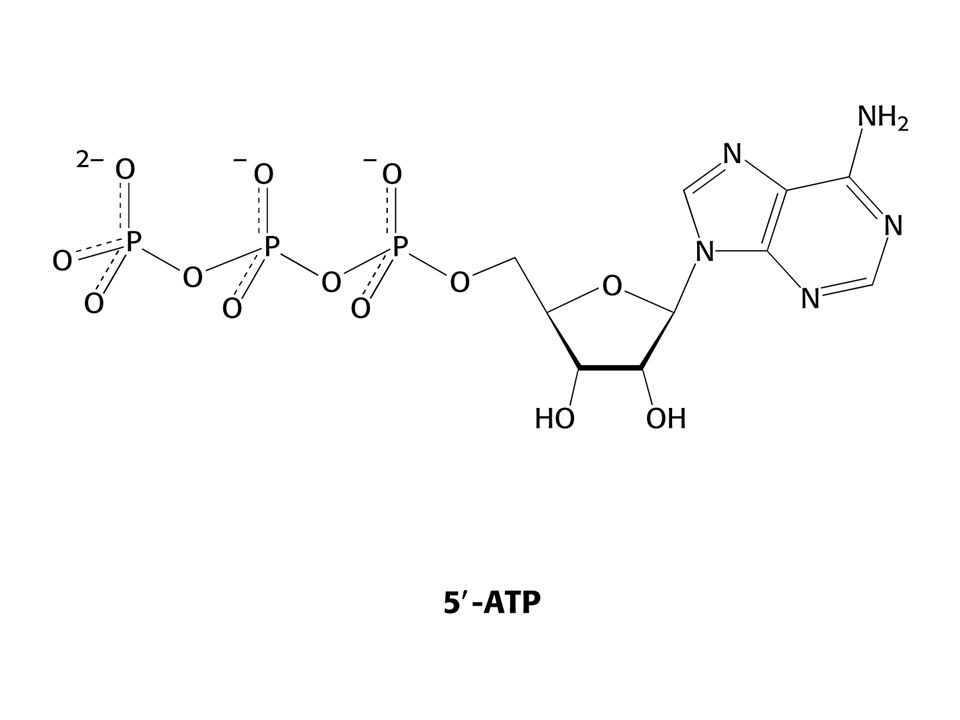
a) Preferovaným templátem je dvojšroubovice DNA, jednovláknová může sloužit také. V žádném případe ne- hybridy DNA-RNA. b) Čtyři ribonukleosidtrifosfáty – ATP, GTP,UTP a CTP. c) Divalentní kovové ionty – Mg2+ a Mn2+. Směr syntézy RNA je 5´ 3´ Elongace řetězce – 3´OH je nukleofiem atakujícím první fosfát na ribose ribonukleosidtrifosfátu. Reakci pohání hydrolýza PPi. Na rozdíl od replikace DNA není přítomen a nutný primer. RNApolymerasa nemá nukleasovou aktivitu jakou má DNApolymerasa, nedovede tedy vyštěpit vadné nukleotidy. V savčích buňkách, na rozdíl od E. coli, je řada RNApolymeras.
 Preferovaným templátem je dvojšroubovice DNA, jednovláknová může sloužit také. V žádném případe ne- hybridy DNA-RNA. b) Čtyři ribonukleosidtrifosfáty – ATP, GTP,UTP a CTP. c) Divalentní kovové ionty – Mg2+ a Mn2+. Směr syntézy RNA je 5´ 3´ Elongace řetězce – 3´OH je nukleofiem atakujícím první fosfát na ribose ribonukleosidtrifosfátu. Reakci pohání hydrolýza PPi. Na rozdíl od replikace DNA není přítomen a nutný primer. RNApolymerasa nemá nukleasovou aktivitu jakou má DNApolymerasa, nedovede tedy vyštěpit vadné nukleotidy. V savčích buňkách, na rozdíl od E. coli, je řada RNApolymeras.")
43
Typy RNA v E. coli.

45
RNApolymerasa přebírá instrukce pro pořadí ribonukleotidů od DNA templátu !!
Složení a pořadí bází nově syntetizované RNA je komplementem DNA templátu.

46
Transkripce startuje u promotorových míst.

47
Transkripce startuje u promotorových míst.

48
Terminační místo na nově syntetizované mRNA E. coli
Terminační místo na nově syntetizované mRNA E. coli. Po čtyřech U následuje“hairpin“struktura.

49
mRNA u eukaryot je modifikována po traskripci
mRNA u eukaryot je modifikována po traskripci. Nukleotidová „čepička“ je přidána na 5´konec a poly (A) je přidán na 3´ konec.
 je přidán na 3´ konec.")
50
Připojení aminokyseliny k molekule tRNA
Připojení aminokyseliny k molekule tRNA. Aminokyselina je esterifikována s 3´OH skupinou terminálního adenosinu tRNA.

51
Transfer RNA je adaptérová molekula při syntéze proteinů.
Transferová RNA má dvě hlavní místa: aminoacylové (připojení aminokyseliny). Templátové rozpoznatelné místo (template-recognition site). Aminokyselina se připojuje na tRNA enzymem – aminoacyl-tRNAsynthetasou. Templátové rozpoznatelné místo je tvořeno třemi bázemi – nazváno antikodon. Antikodon na tRNA rozpozná komplementární místo tří bází na mRNA nazvané kodon.
. Templátové rozpoznatelné místo (template-recognition site). Aminokyselina se připojuje na tRNA enzymem – aminoacyl-tRNAsynthetasou. Templátové rozpoznatelné místo je tvořeno třemi bázemi – nazváno antikodon. Antikodon na tRNA rozpozná komplementární místo tří bází na mRNA nazvané kodon.")
52
Symbolická struktura aminoacyl-tRNA

53
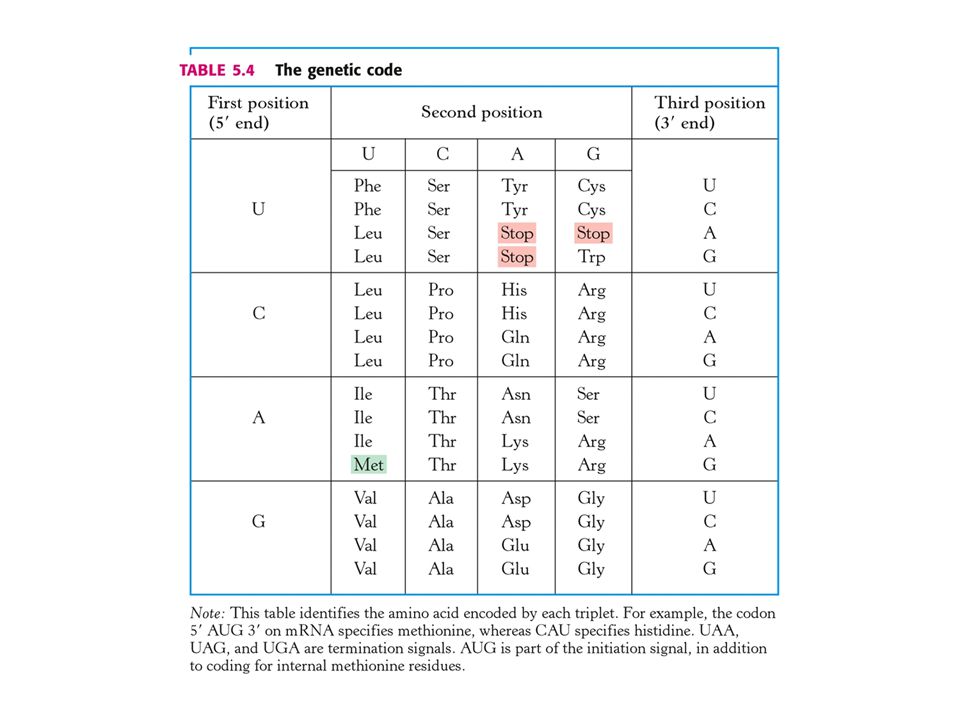
Genetický kód. Genetický kód je vztah mezi sekvencemi bází DNA (nebo jejího transkriptu mRNA) a sekvencí aminokyselin v proteinu. Genetický kód byl postulován v roce 1961 F. Crickem a S. Brennerem. 1) Tři nukleotidy (kodon) kódují jednu aminokyselinu. 2) V genetickém kódu se kodony nepřekrývají. 3) Triplety (kodony) nejsou od sebe odděleny tečkou. 4. Genetický kód je degenerovaný. Máme celkem čtyři báze. Vytváříme triplety po třech bázích. Báze se mohou opakovat a záleží na jejich pořadí. Máme k dispozici celkem 43 = 64 kodonů. Tři jsou tzv. STOP kodony – ukončují translaci. Pro 20 aminokyselin zbývá 61 kodonů. Některé aminokyseliny jsou určovány více kodony.
 Tři nukleotidy (kodon) kódují jednu aminokyselinu. 2) V genetickém kódu se kodony nepřekrývají. 3) Triplety (kodony) nejsou od sebe odděleny tečkou. 4. Genetický kód je degenerovaný. Máme celkem čtyři báze. Vytváříme triplety po třech bázích. Báze se mohou opakovat a záleží na jejich pořadí. Máme k dispozici celkem 43 = 64 kodonů. Tři jsou tzv. STOP kodony – ukončují translaci. Pro 20 aminokyselin zbývá 61 kodonů. Některé aminokyseliny jsou určovány více kodony.")
54
V genetickém kódu se kodony nepřekrývají.

55
Pořadí bází je čteno sekvenčně od počátku (startu) bez dělících znamének.
 bez dělících znamének.")
56
Hlavní charakteristiky genetického kódu
Jedině Trp (UGG) a Met (AUG) jsou určeny jedním kodonem. Leu, Arg a Ser jsou determinovány šesti kodony. Počet kodonů koreluje s četností výskytu aminokyseliny v proteinech. Kodony, které určují stejnou aminokyselinu se nazývají synonyma. Většina synonym se liší pouze v bázi na třetím místě. Čte se od 5´ke 3´. Jaký je biologický smysl degenerovaného genetického kódu ? Pokud by nebyl degenerován, určovalo by dvacet kodonů dvacet aminokyselin a zbylých 44 by určovalo STOP. Pravděpodobnost mutace by častěji vedla k terminaci což má za následek inaktivní protein, zatímco záměna jedné aminokyseliny za jinou není tak nebezpečná.
 a Met (AUG) jsou určeny jedním kodonem. Leu, Arg a Ser jsou determinovány šesti kodony. Počet kodonů koreluje s četností výskytu aminokyseliny v proteinech. Kodony, které určují stejnou aminokyselinu se nazývají synonyma. Většina synonym se liší pouze v bázi na třetím místě. Čte se od 5´ke 3´. Jaký je biologický smysl degenerovaného genetického kódu Pokud by nebyl degenerován, určovalo by dvacet kodonů dvacet aminokyselin a zbylých 44 by určovalo STOP. Pravděpodobnost mutace by častěji vedla k terminaci což má za následek inaktivní protein, zatímco záměna jedné aminokyseliny za jinou není tak nebezpečná.")
58
mRNA obsahuje START a STOP kodony pro syntézu proteinů.
Syntéza proteinů se odehrává na ribosomech. STOP kodony jsou UAA, UAG a UGA. Tyto jsou čteny specifickými proteiny – uvolňovací faktory (RF). Po vazbě těchto faktorů se syntetizovaný protein oddělí. Signály START jsou kmplexnější. 1) U baktérií – START formylMet, specifická fMet-tRNA rozpozná kodon AUG nebo méně často GUG. Pozor – AUG je kodon pro Met uvnitř řetězce a GUG pro vnitřní Val.
. Po vazbě těchto faktorů se syntetizovaný protein oddělí. Signály START jsou kmplexnější. 1) U baktérií – START formylMet, specifická fMet-tRNA rozpozná kodon AUG nebo méně často GUG. Pozor – AUG je kodon pro Met uvnitř řetězce a GUG pro vnitřní Val.")
61
Eukaryota – AUG v blízkosti 5´konce mRNA je startovním signálem
Eukaryota – AUG v blízkosti 5´konce mRNA je startovním signálem. Této signál je přečten iniciační tRNA spojenou s Met.

62
Genetický kód je téměř univerzální
Genetický kód je téměř univerzální. Výjimkou je zvláště lidská mitochondrie, mírně kvasničná….

63
Proteosyntéza. Proteiny se syntetizují spojením aminoskupiny aktivované aminoacyl-tRNA s karboxylovým koncem peptidu.

64
Sekvence kvasničné Ala-tRNA
Sekvence kvasničné Ala-tRNA.Modifikované nukleosidy: methylinosin (mI), dihydrouridin (UH2), ribothymidin (T), methylguanosin (mG), dimethylguanosin (m2G) a inosin (I).
, dihydrouridin (UH2), ribothymidin (T), methylguanosin (mG), dimethylguanosin (m2G) a inosin (I).")
65
Aminoacyl-tRNA. Aminokyseliny jsou spojeny s tRNA přes esterovou vazbu v polohách 2´nebo 3´ (-hydroxyl).
..")
66
Ribosom prokaryot je ribonukleoproteinová částice (70S) tvořená malou (30S) a velkou (50S) podjdnotkami. A) 30S podjednotka, B) 50S podjednotka, C) 70 S ribosom.
 30S podjednotka, B) 50S podjednotka, C) 70 S ribosom..")
67
Prokaryota mají 70S ribosomy, složené z malé podjednotky 30S a velké 50S. Podjednotka 50S je složena z 5S RNA podjednotky obsahující 120 nukleotidů a 23S RNA podjednotky se nukleotidů a 34 proteinů. Podjednotka 30S je složena ze 1540 nukleotidových RNA a 21 proteinů. Eukaryota mají 80S ribosomy, každý složen z malé 40S a velké 60S podjednotky. Podjednotka 60S je složena z 5S RNA podjednotky se 120 nukleotidy a 28S RNA podjednotky se nukleotidy, a 5, 8S RNA podjednotky se 160 nukleotidy, a ~49 proteiny. Podjednotka 40S obsahuje 1900 nukleotidových RNA a ~33 proteinů. Sedimentační koeficient je vyjádřen v jednotkách S (Svedberg) = 10-13s.
 = 10-13s.")
68
Tvar ribosomu. Spojení velké podjednotky (1) s malou podjednotkou (2).
 s malou podjednotkou (2).")
69
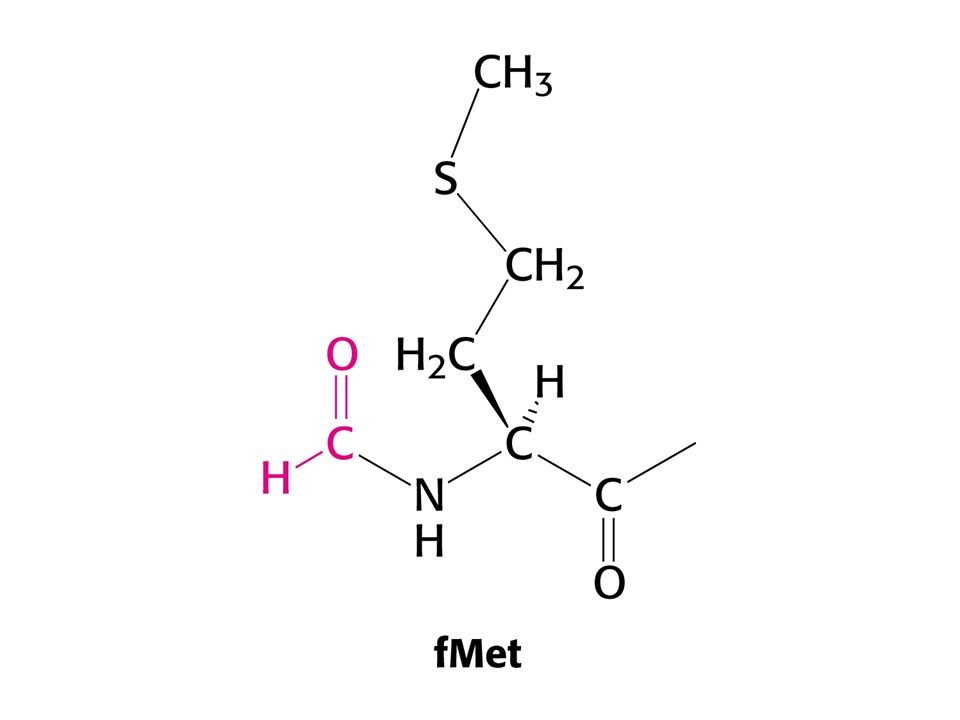
Formylace Met-tRNA.

70
Vazebná místa tRNA na 70S ribosomu
Vazebná místa tRNA na 70S ribosomu. A-aminoacyl, P- peptidyl a E uvolnění (exit). Každá molekula tRNA je v kontaktu s oběma podjednotkami. B) Molekuly tRNA v místech A a P se párují s mRNA.
. Každá molekula tRNA je v kontaktu s oběma podjednotkami. B) Molekuly tRNA v místech A a P se párují s mRNA.")
71
Mechanismus syntézy proteinů. Cyklus startuje s peptidyl-tRNA v místě P. Aminoacyl-tRNA se váže do místa A. Pokud jsou obě tato místa obsazena dojde k tvorbě peptidové vazby.

72
Tvorba peptidové vazby.

73
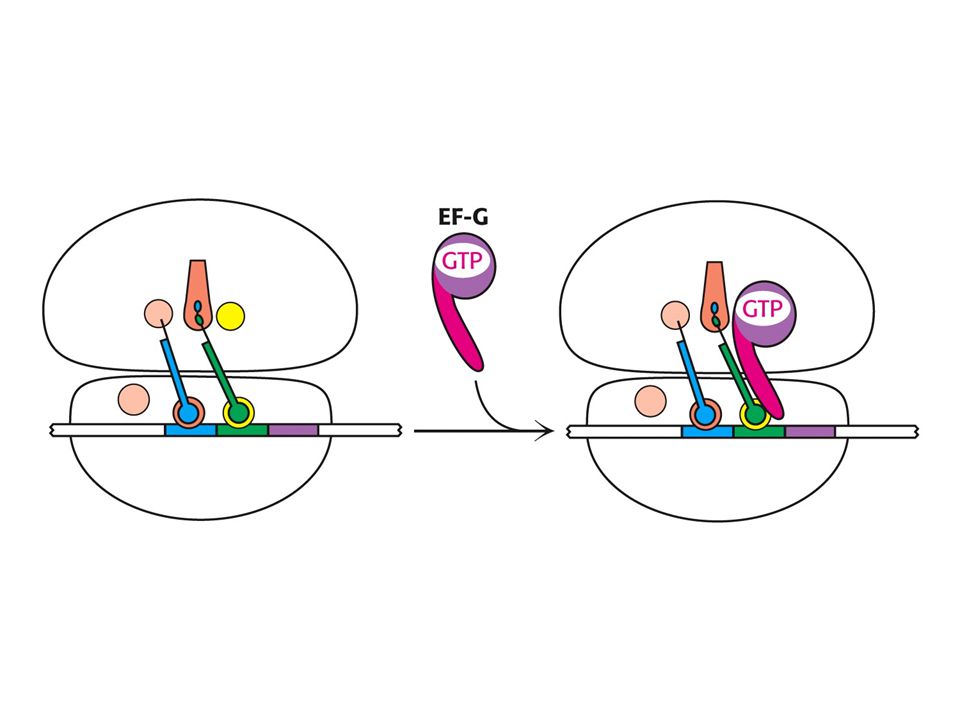
tRNA a mRNA jsou translokovány působením elongačního faktoru G, který posune deacylovanou tRNA do místa E.

74
Když se deacylovaná tRNA dostane do místa E, oddisociuje – cyklus je kompletní.

75
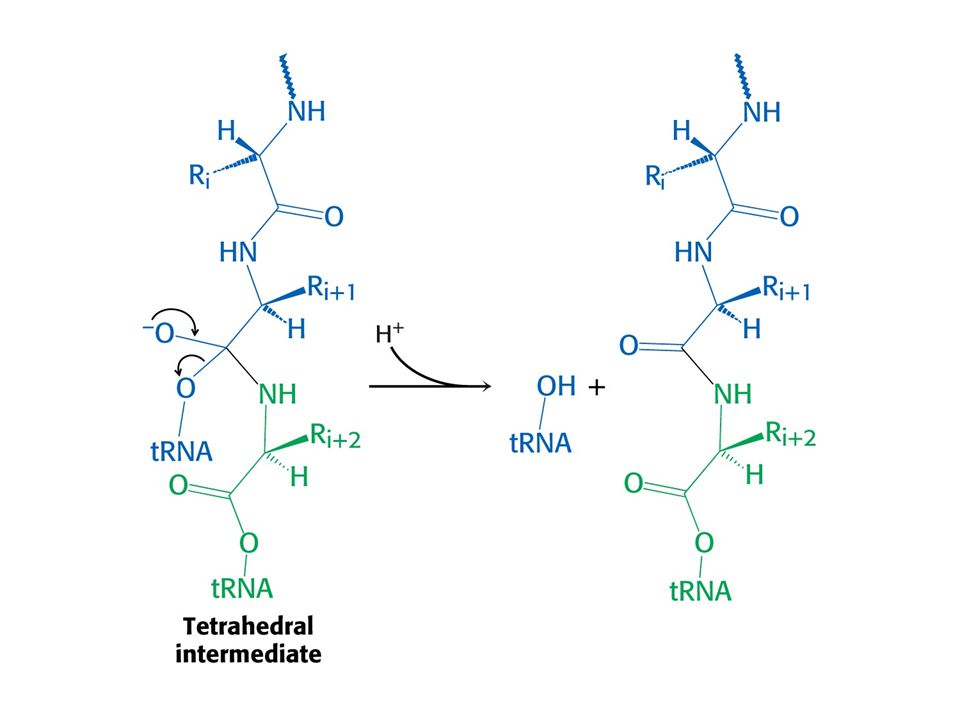
Tvorba peptidové vazby
Tvorba peptidové vazby. Aminoskupina aminoacyl-tRNA atakuje karbonylovou skupinu esterové vazby peptidyl-tRNA.

78
Role formylace. Pokud je koncová aminoskupina volná, může se dipeptidyl-tRNA cyklizovat a tak odštěpit z tRNA. Formylace této reakci zabraňuje.

79
Proteinové faktory hrají klíčové role při syntéze proteinů
Proteinové faktory hrají klíčové role při syntéze proteinů. FormylMet-tRNA a mRNA vstupují na ribosom za účasti tří iniciačních faktorů (IF1, IF2 a IF3).
.")
80
Elongační faktory přenášejí aminoacyl-tRNA na ribosom ((EF-Tu, G-protein).
Elongační faktor Ts indukuje disociaci GDP. Tvorba peptidové vazby následuje po GTP-poháněné translokaci tRNA a mRNA.

81
Mechanismus translokace
Mechanismus translokace. Ve formě GTP se váže na EF-Tu-vazebné místo na podjednotce 50S. To stimuluje hydrolýzu GTP a indukuje konformační změny v EF-G, které pohání vstup EF-G do místa A na 30S.

84
Syntéza proteinů je ukončena uvolňovacími faktory, které přečtou STOP kodón.
Aminoacyl-tRNA se normálně neváže do místa A na ribosomu, pokud se objeví kodóny UAA, UGA a UAG. Tyto kodóny čtou tři uvolňovací faktory (RFs), což jsou proteiny. Jejich funkcí je spuštění hydrolýzy namísto syntézy peptidové vazby.
, což jsou proteiny. Jejich funkcí je spuštění hydrolýzy namísto syntézy peptidové vazby.")
85
Mnohé geny u eukaryont jsou mozaikou intronů a exonů. Např
Mnohé geny u eukaryont jsou mozaikou intronů a exonů . Např. gen pro b-globin.

86
Ve kterém stádiu exprese genů se intervenující sekvence oddělují ?
Části oddělující se z primárního transkriptu se nazývají introny (intervening sequences). Části zůstávající v hotové RNA se nazývají exony (expressed regions)
. Části zůstávající v hotové RNA se nazývají exony (expressed regions)")
87
Kontinuální gen.

88
Dvě smyčky jednovláknové DNA (modrá) a a smyčka dvojvlákna (zelená a modrá) v případě, že gen obsahuje intervenční sekvence.
 a a smyčka dvojvlákna (zelená a modrá) v případě, že gen obsahuje intervenční sekvence.")
89
Gen je přepsán na primární transkript, který je modifikován přidáním čepičky a poly(A). Vnitřní sekvence jsou odstřiženy a vzniká mRNA.

90
Splicing (spojení) je komplexní proces odehrávající se na splicesomech
Splicing (spojení) je komplexní proces odehrávající se na splicesomech. Introny se poznají obvykle podle toho, že začínají GU a končí AG čemuž předchází na pyrimidiny bohatý úsek.
 je komplexní proces odehrávající se na splicesomech. Introny se poznají obvykle podle toho, že začínají GU a končí AG čemuž předchází na pyrimidiny bohatý úsek.")
Podobné prezentace










>")


>")




>")
>")




>")


