Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Ing. Zuzana Balounová, PhD.
Fyziologie rostlin Základní kurs 10 Růst a vývoj Ing. Zuzana Balounová, PhD.

2
složité fyzikální a biochemické reakce - o jejich pochopení a vysvětlení se snaží vědecké týmy fyziologů, biochemiků, biofyziků, genetiků, molekulárních biologů apod. mnoho nevyřešených otázek - nevíme jak rostliny měří čas (tzv. biologické hodiny) jaký je mechanismus působení světelného signálu jak vzniká z oplodněného vajíčka dospělá rostlina zda semeno za příznivých podmínek vůbec vyklíčí jak je zajištěno, aby se jednotlivé orgány vytvářely ve správné velikosti a ve vhodnou dobu jak pracují systémy vnitřní regulace růstu rostlin jaký je mechanismus aktivace a inaktivace určitého souboru genů…
 jaký je mechanismus působení světelného signálu. jak vzniká z oplodněného vajíčka dospělá rostlina. zda semeno za příznivých podmínek vůbec vyklíčí. jak je zajištěno, aby se jednotlivé orgány vytvářely ve. správné velikosti a ve vhodnou dobu. jak pracují systémy vnitřní regulace růstu rostlin. jaký je mechanismus aktivace a inaktivace určitého. souboru genů…")
3
Obecné problémy růstu a vývoje
Ontogeneze - růst a vývoj (dva vzájemně závislé procesy) - změny strukturální a funkční (organismus vzniká, vyvíjí se a zaniká) Růst - nevratné narůstání hmotnosti (zvětšování objemu, velikosti, délky a tvaru těla (orgánu) - v souladu s komplexní genetickou informací jedince - a ve vazbě na komplex faktorů vnitřních (v rámci celistvosti rostliny) ale i faktorů vnějšího prostředí tvorba nových buněk (závislá na dostatku stavebního materiálu z biochemických procesů) kvantitativní růst výměna informací tvarová a funkční specializace nových buněk diferenciace a morfogeneze - kvalitativní růst (postupné tvarové změny) Vývoj - časové úseky růstu a diferenciace (kvantitativní a kvalitativní změny) během ontogeneze
 - změny strukturální a funkční. (organismus vzniká, vyvíjí se a zaniká) Růst - nevratné narůstání hmotnosti. (zvětšování objemu, velikosti, délky a tvaru těla (orgánu) - v souladu s komplexní genetickou informací jedince. - a ve vazbě na komplex faktorů vnitřních (v rámci celistvosti rostliny) ale i faktorů vnějšího prostředí. tvorba nových buněk (závislá na dostatku stavebního materiálu z. biochemických procesů) kvantitativní růst. výměna informací tvarová a funkční specializace nových buněk. diferenciace a morfogeneze - kvalitativní růst (postupné tvarové změny) Vývoj - časové úseky růstu a diferenciace (kvantitativní a kvalitativní změny) během ontogeneze.")
4
Obecné problémy růstu a vývoje
V ontogenezi rostliny lze rozlišit: morfogenezi rostliny jako celku organogenezi formování jednotlivých orgánů histogenezi pletiv metabolické procesy růstu a vývoje genetické vazby na regulační mechanismy, na komplexní informaci jedince (v DNA) ekologickou adaptaci rostliny evoluční stránku ve vazbě na fylogenetický vývoj rostliny - druhu Růst je úzce spojen s buněčným metabolismem (dodává látky a energie nutné k výstavbě těla).
 ekologickou adaptaci rostliny. evoluční stránku ve vazbě na fylogenetický. vývoj rostliny - druhu. Růst je úzce spojen s buněčným metabolismem (dodává látky a energie nutné k výstavbě těla).")
5
Růstové procesy u většiny rostlin nejsou tak pevně určeny jako u živočichů.
axiální charakter růstu - zřetelná orientace nových struktur podle jisté osy modulární charakter růstu - opakování některých strukturních jednotek - modulů U rostlin má zvláštní význam polarita buněk i celých orgánů, projevující se nestejnými vlastnostmi protilehlých, zdánlivě stejných struktur. Je ovlivněna brzy po vzniku nové buňky faktory vnějšího prostředí.

6
Dělení a růst buněk zopakovat za domácí úkol
4 fáze buněčného cyklu: 1. presyntetická G1: narůstá jádro a cytoplazma 2. syntetická S: replikace DNA 3. postsyntetická G2: příprava na mitózu 4. mitóza M: člení se na čtyři úseky: profáze, metafáze, anafáze, telofáze

8
dva kritické okamžiky:
Po dokončení cytokineze vchází buňka do fáze G1v níž nastává více možností jejího dalšího vývoje. buňka může vkročit do etapy dělení nebo se může její vývoj zastavit (fáze G0) nebo se může degradovat a odumřít Po odeznění této kritické etapy vchází bunka do fáze S v níž se začíná syntéza zdvojením(replikací) DNA kritický bod regulace přchodu od fáze G1 do S = START nebo R (angl. Restriction point) Druhý kritický okamžik nastává na konci fáze G2, kdy se provádí zpětná kontrola dosavadního průběhu buněčného cyklu. Teprve jeho překročením je podmíněno buněčné dělení.
 nebo se může degradovat a odumřít. Po odeznění této kritické etapy vchází bunka do fáze S v níž se začíná syntéza zdvojením(replikací) DNA. kritický bod regulace přchodu od fáze G1 do S = START nebo R. (angl. Restriction point) Druhý kritický okamžik nastává na konci fáze G2, kdy se provádí zpětná kontrola dosavadního průběhu buněčného cyklu. Teprve jeho překročením je podmíněno buněčné dělení.")
9
Regulace buněčného cyklu
podléhá reakcím fosforylace (katalyzovaným různými typy kináz) defosforylace (prostřednictvím fosfatáz). Proces fosforylace bílkovinných substrátů odpovídá kinázám založeným na cyklinech (CDK – ang. cyclin dependet kinase). Cyklin B interaguje s proteinem který je nazýván p34cdc2. Právě tento protein je hlavním regulátorem buněčného cyklu. Svoji úlohu plní tím, že fosforyluje velké množství proteinu v jádře. Vazbacyklinu B na p34cdc2 vede k jeho aktivaci
 defosforylace (prostřednictvím fosfatáz). Proces fosforylace bílkovinných substrátů odpovídá kinázám založeným na cyklinech (CDK – ang. cyclin dependet kinase). Cyklin B interaguje s proteinem který je nazýván p34cdc2. Právě tento protein je hlavním regulátorem buněčného cyklu. Svoji úlohu plní tím, že fosforyluje velké množství proteinu v jádře. Vazbacyklinu B na p34cdc2 vede k jeho aktivaci.")
10
Způsoby oddělování dceřiných buněk
zaškrcování mateřská buňka se protáhne do tvaru piškotu a rozdělí na dvě buňky dceřiné. (baktérie, sinice, řasy, plastidy aj.) pučení na mateřské buňce se vytvoří hrbolek, jádro se protáhne a nakonec rozdělí. Původní hrbolek s doroste do velikosti mateřské buňky. Na mateřské buňce se 4asto zakládá hned další buňka (u kvasinek vznikají retízky buněk). přehrádečné dělení v telofázi se mezi mateřskou a dceřinou buňkou vytvoří prepážka: · současně v celém rozsahu u buněk bohatých na základní cytoplazmu, · postupně dostředivě (centripetálně) nebo · odstředivě (centrifugálně). volné novotvoření dělení původně jediného buněčného jádra se několikrát opakuje, takže vzniká větší počet jader, která se rozptýlí a mezi nimi se dodatečně vytvoří buněčné stěny (typické pro zásobní pletiva)
 pučení na mateřské buňce se vytvoří hrbolek, jádro se protáhne a nakonec rozdělí. Původní hrbolek s doroste do velikosti mateřské buňky. Na mateřské buňce se 4asto zakládá hned další buňka (u kvasinek vznikají retízky buněk). přehrádečné dělení v telofázi se mezi mateřskou a dceřinou buňkou vytvoří. prepážka: · současně v celém rozsahu u buněk bohatých na základní cytoplazmu, · postupně. dostředivě (centripetálně) nebo. · odstředivě (centrifugálně). volné novotvoření dělení původně jediného buněčného jádra se několikrát. opakuje, takže vzniká větší počet jader, která se rozptýlí a mezi nimi se dodatečně vytvoří buněčné stěny (typické pro zásobní pletiva)")
11
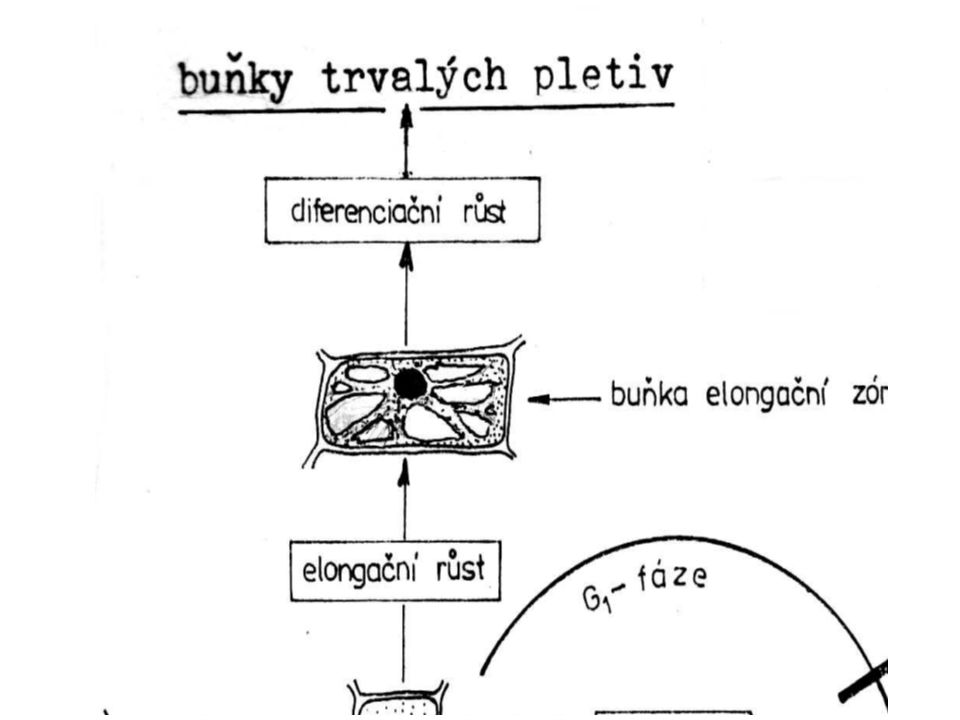
Pri růstu prodělává rostlinná buňka tři růstové fáze:
· zárodečnou (embryonální) · prodlužovací (prolongační) · rozlišovací (diferenciační).
 · prodlužovací (prolongační) · rozlišovací (diferenciační).")
12
– meristematické buňky malé vyplněny cytoplazmou
zárodečná fáze – meristematické buňky malé vyplněny cytoplazmou velké kulovité centrální jádro při růstu se zvětšuje množství základní cytoplazmy a zvětšuje i rozměr každé dělivé buňky jádro se rozdělí nakonec se oddělí dvě dceřiné buňky novou buněčnou stěnou přibývá množství živé hmoty (protoplastu) a narůstá počet buněk
 a narůstá počet buněk.")
13
- narůstá buněčná stěna do plochy vmezeřeným růstem (intususcepcí)
prodlužovací fáze - narůstá buněčná stěna do plochy vmezeřeným růstem (intususcepcí) objem buňky se zvětšuje (20 až 30krát) avšak množství základní cytoplazmy zůstává téměř stejné. - vznikají vakuoly – spojují se - centrální vakuola základní cytoplazma: nástěnný povlak Buňky mají nízký vodní potenciál, který podmiňuje nasávání vody do jejich vakuol i na úkor buněk sousedních pletiv - tento osmotický proces podmiňuje nevratné rozpínání buňky - nápadně se zvětšuje její objem
 objem buňky se zvětšuje (20 až 30krát) avšak množství základní cytoplazmy zůstává téměř stejné. - vznikají vakuoly – spojují se - centrální vakuola. základní cytoplazma: nástěnný povlak. Buňky mají nízký vodní potenciál, který podmiňuje nasávání vody do jejich vakuol i na úkor buněk sousedních pletiv. - tento osmotický proces podmiňuje nevratné rozpínání buňky - nápadně se zvětšuje její objem.")
14
diferenciační fáze - specializace dle funkce (na buňku krycí, vodivou aj.) - nabývání konečného tvaru - buněčná stěna tloustne - nabývá stavby a tvaru, který odpovídá funkci vznikajícího pletiva.

15
- strukturní a funkční rozlišení jednotlivých buněk, pletiv a orgánů
Diferenciace - strukturní a funkční rozlišení jednotlivých buněk, pletiv a orgánů její podstatou je diferenční genová aktivita (aktivita pouze určitých genů v určitých buňkách (pletivu) a v určitých fázích ontogeneze) u apikálních meristémů: dvě buňky dceřiné: se obě budou dále dělit nebo jedna zůstává dělivou, druhá již ne (především na okraji meristému) každá mateřská meristematická buňka je polarizována - - dělení není rovnocenné (vzhledem ke strukturám a rozložení látek v buňce podél osy polarity)
 a v určitých fázích ontogeneze) u apikálních meristémů: dvě buňky dceřiné: se obě budou dále dělit. nebo jedna zůstává dělivou, druhá již ne. (především na okraji meristému) každá mateřská meristematická buňka je polarizována - - dělení není rovnocenné. (vzhledem ke strukturám a rozložení látek v buňce podél osy polarity)")
16
velká perioda růstu: objem každé buňky (a proto i každého orgánu) - roste zpočátku zvolna - pak se rychlost růstu nápadně zvětšuje (fáze prodlužovací) - později klesá, až se zastavuje
 - později klesá, až se zastavuje.")
17
(opak = ekvální dělení)
Inekvální dělení - dělení buňky, při kterém jsou nově vzniklé dceřiné buňky morfologicky nebo funkčně nestejné (opak = ekvální dělení) - liší se regulačními mechanismy včetne genové aktivity (= jedna dceřinná b. zůstává meristematickou, druhá se diferencuje dle funkce)
 - liší se regulačními mechanismy včetne genové aktivity (= jedna dceřinná b. zůstává meristematickou, druhá se diferencuje dle funkce)")
18
Inekvální dělení je základem buněčné diferenciace
Probíhá i při diferenciaci průduchů kořenových vlásků pylových zrn (vznik vegetativní a generativní buňky) sítkovic (vznik původních buněk) … Rozhodující vliv na určení tohoto asymetrického dělení buňky má specifické uspořádání cytoskeletu
 sítkovic (vznik původních buněk) … Rozhodující vliv na určení tohoto asymetrického dělení buňky má specifické uspořádání cytoskeletu.")
19
Inekvální dělení při vzniku průduchu Dceřiná buňka, která je menší a bohatší cytoplazmou se dělí (=zvláštní typ diferenciace) a dá vznik svěracím buňkám průduchu
 a dá vznik svěracím buňkám průduchu")
20
Inekvální dělení při tvorbě kořenového vlásku (Phleum sp
Inekvální dělení při tvorbě kořenového vlásku (Phleum sp.) - kořenové vlásky se mohou diferencovat téměř z každé buňky rhizodermis - při dělení buněk rhizodermis se diferencují kratší buňky bohatší cytoplazmou, dále se již nedělící (trichoblasty) - z nich vyrůstají kořenové vlásky
 - kořenové vlásky se mohou diferencovat téměř z každé buňky rhizodermis - při dělení buněk rhizodermis se diferencují kratší buňky bohatší cytoplazmou, dále se již nedělící (trichoblasty) - z nich vyrůstají kořenové vlásky.")
21
terminální buňky - na vzrostných vrcholech řas, hub, mechů a u některých výtrusných rostlin
- oddělují další buňky - segmenty Iniciály (protomeristém = několik buněk) - vytvářejí soubor tzv. primárních meristémů (dermatogen, periblem, plerom, kalyptrogen), která oddělují již trvalá pletiva. U vyšších rostlin obnovují někdy i trvalá pletiva dělivou schopnost - meristematická pletiva sekundární (kambium, felogen)
 - vytvářejí soubor tzv. primárních meristémů (dermatogen, periblem, plerom, kalyptrogen), která oddělují již trvalá pletiva. U vyšších rostlin obnovují někdy i trvalá pletiva dělivou schopnost - meristematická pletiva sekundární (kambium, felogen)")
22
Diferenciace v širším smyslu - tvorba pletiv a vznik celých orgánů.
kořeny – jednoduchá a jednotná stavba apikální meristém se uvádí v činnost v klíčícím semeni rychle a stejně rychlý je i celý další růst kořene. kořenová čepička chrání dělivé buňky před poškozením (slizové látky) podílí se i na řízení růstu kořene při poškození se aktivuje klidové centrum (mezi čepičkou a vlastní dělivou zónou) - vyrovnává vzniklé ztráty. - diferenciace vodivých pletiv začíná na rozhraní mezi vlastním meristémem a zónou dlouživého růstu.
 podílí se i na řízení růstu kořene. při poškození se aktivuje klidové centrum (mezi čepičkou a vlastní dělivou zónou) - vyrovnává vzniklé ztráty. - diferenciace vodivých pletiv začíná na rozhraní mezi vlastním meristémem a zónou dlouživého růstu.")
23
Diferenciace v širším smyslu - tvorba pletiv a vznik celých orgánů.
prýt - základy již v embryu (apikální meristému): - zajišťuje růst a větvení stonku zakládání dalších meristémů pro růst listu a květu - buňky uspořádány specificky pro různé druhy - diferenciace je velmi dokonale řízena již v první fázi zakládání listu (kdy se rozhoduje např. o počtu listů, o způsobu a frekvenci větvení stonku, o časových intervalech mezi tvorbou listů aj). - listy se rozvíjejí v listových primordiích pomocí několika dalších specifických meristémů.
: - zajišťuje růst a větvení stonku. zakládání dalších meristémů pro růst listu a květu. - buňky uspořádány specificky pro různé druhy. - diferenciace je velmi dokonale řízena již v první fázi zakládání listu (kdy se rozhoduje např. o počtu listů, o způsobu a frekvenci větvení stonku, o časových intervalech mezi tvorbou listů aj). - listy se rozvíjejí v listových primordiích pomocí několika dalších specifických meristémů.")
24
Největší dělivá aktivita v prvních fázích růstu:
vrstva buněk na vnějším konci primordia později na okrajích někdy i v bazální části

25
dělivá (meristematická, meristémy)
Rostlinná pletiva: zopakujte si! trvalá dělivá (meristematická, meristémy) · vrcholová - apikální (vegetační vrchol kořene nebo stonku) · vmezeřená - interkalární (v koléncích trav), · boční - laterální (kambium, felogen) · okrajová - marginální (na okraji čepele listu), · bazální - na bázi listu v konečné fázi růstu rozptýlená - difuzní (endosperm)
 · vrcholová - apikální (vegetační vrchol kořene nebo stonku) · vmezeřená - interkalární (v koléncích trav), · boční - laterální (kambium, felogen) · okrajová - marginální (na okraji čepele listu), · bazální - na bázi listu v konečné fázi růstu. rozptýlená - difuzní (endosperm)")
26
Podle vzniku · pletiva pravá
nepravá (vznikají dodatečným spojením původně samostatných buněk, jako např. u rasy Hydrodyctyon reticulatum, u níž vlastne spory srostou v sítovitou strukturu, která opustí materskou bunku) nebo druhotným srůstem pletiv (plektenchym hub). Podle tvaru a stavby · parenchym · prosenchym · kolenchym · sklerenchym Podle funkce · krycí, ochranná · prvotní: pokožka (epidermis, rhizodermis) · druhotná: korek, borka · mechanická: kolenchym, sklerenchym · asimilační: palisádový parenchym · sací: tracheje, tracheidy, sítkovice · zásobní: parenchym, v němž se ukládají rezervní látky · provzdušňovací: houbový parenchym, aerenchym, · vyměšovací: žláznaté chlupy, medníky aj.
 nebo druhotným srůstem pletiv (plektenchym hub). Podle tvaru a stavby. · parenchym. · prosenchym. · kolenchym. · sklerenchym. Podle funkce. · krycí, ochranná. · prvotní: pokožka (epidermis, rhizodermis) · druhotná: korek, borka. · mechanická: kolenchym, sklerenchym. · asimilační: palisádový parenchym. · sací: tracheje, tracheidy, sítkovice. · zásobní: parenchym, v němž se ukládají rezervní látky. · provzdušňovací: houbový parenchym, aerenchym, · vyměšovací: žláznaté chlupy, medníky aj.")
27
Látky regulující růst rostlin
Koordinace růstových procesů v rostlinách je zajišťována dokonalým řídícím mechanismem rostlina však nemá centrální řídící systém -koordinace spočívá výhradně na korelacích řízení procesů uvnitř jedné buňky souhra v rámci pletiv přenos morfogenetických signálů se uskutečňuje převážně pomocí poměrně jednoduchých sloučenin

28
Fytohormony - vážou se na specifické vazebné bílkoviny, receptory (na membránách, v cytoplazmě nebo v jádře)
")
29
Fytohormony Každý hormon má několik funkcí a každá z nich je regulována dalším fytohormonem Fytohormony vzájemně ovlivňují svůj metabolismus a transport, což vyvolává velmi komplexní systémy regulace Citlivě reagují na podněty vnějšího prostředí podílí se na zapojování obranných mechanismů ve stresových podmínkách

30
Fytohormony jsou organické sloučeniny, syntetizované v určitých pletivech (částech) rostliny. Z místa vzniku jsou pak transportovány vodivými pletivy do jiných částí rostliny, kde ovlivňují různé fyziologické procesy. Látky, které růst povzbuzují, nazýváme stimulátory, brzdící růst - inhibitory. Všechny tyto látky působí ve velmi malých tzv. fyziologických koncentracích.

31
stimulátory inhibitory nativní syntetické nativní syntetické auxiny
gibereliny cytokininy auxinoidy kyselina abscisová etylén velké množství látek

32
Auxiny

33
Auxiny se v rostlinách se syntetizuje z tryptofanu
Nejznámější je kyselina b indolyloctová (IAA) a kyselina fenyloctová. Auxinoidy uměle syntetizované, s podobným účinkem Mezi nejznámější patří kyselina b indolylmáselná (IBA) a - naftyloctová (NAA) 2,4-dichlorfenoxyoctová (2,4-D)
 a kyselina fenyloctová. Auxinoidy uměle syntetizované, s podobným účinkem. Mezi nejznámější patří kyselina b indolylmáselná (IBA) a - naftyloctová (NAA) 2,4-dichlorfenoxyoctová (2,4-D)")
34
Auxiny se tvoří především v lodyžním vrcholu
a v nejmladších listech, odkud proudí do ostatních částí rostliny Transport auxinů má výrazně polární charakter V nadzemních částech rostliny se pohybuje vždy bazipetálně (od vzrostného vrcholu a z mladých listů směrem ke kořenům)
")
35
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
36
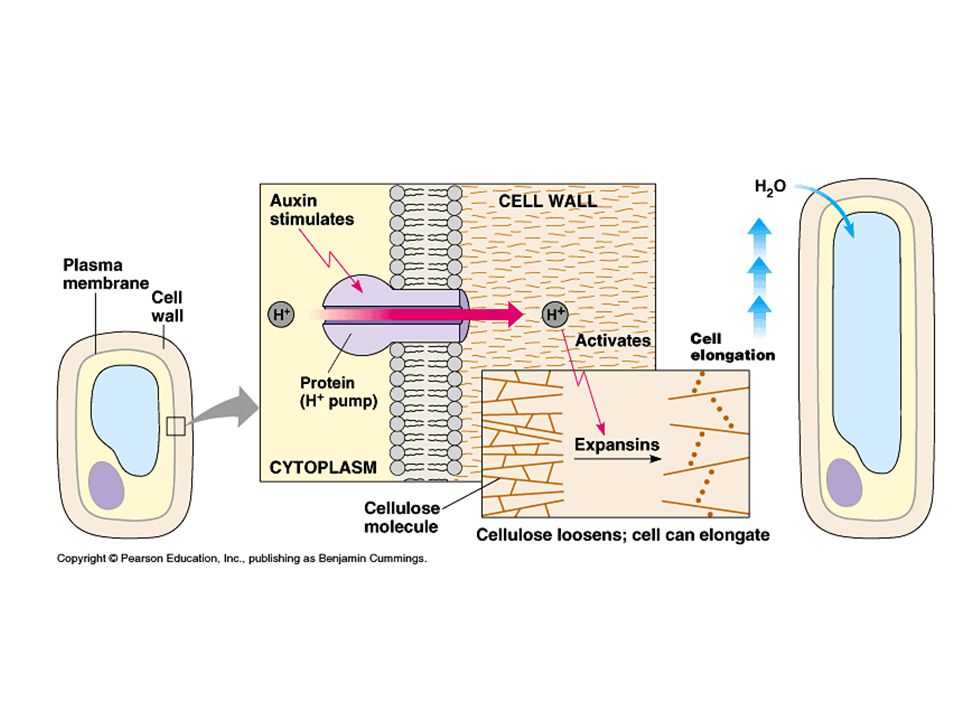
stimulace dlouživého růstu buněk
Působení auxinů v rostlinách je mnohostranné. Často působí v interakci s dalšími fytohormony stimulace dlouživého růstu buněk auxin stimuluje činnost H – pump okyselování buněčné stěny – zvýšení roztažitelnosti -zesilují mitotickou aktivitu pletiv (spolupůsobení cytokininů) -při poranění pletiva stimulují tvorbu ochranné vrstvy zkorkovatělých buněk zacelujících ránu.
 -při poranění pletiva stimulují tvorbu ochranné vrstvy zkorkovatělých buněk zacelujících ránu.")
37
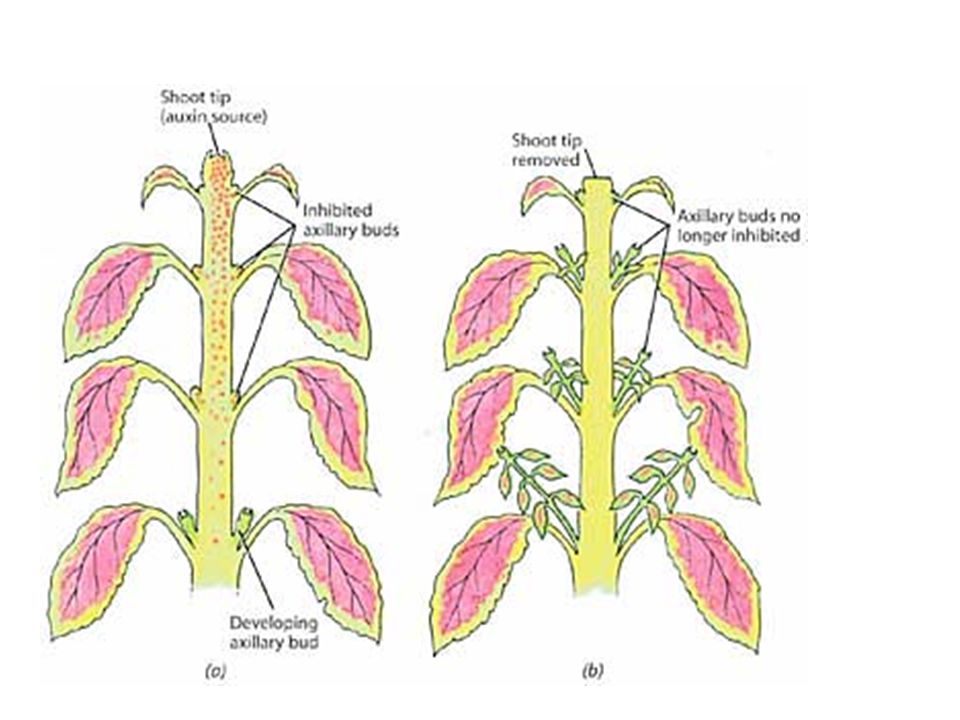
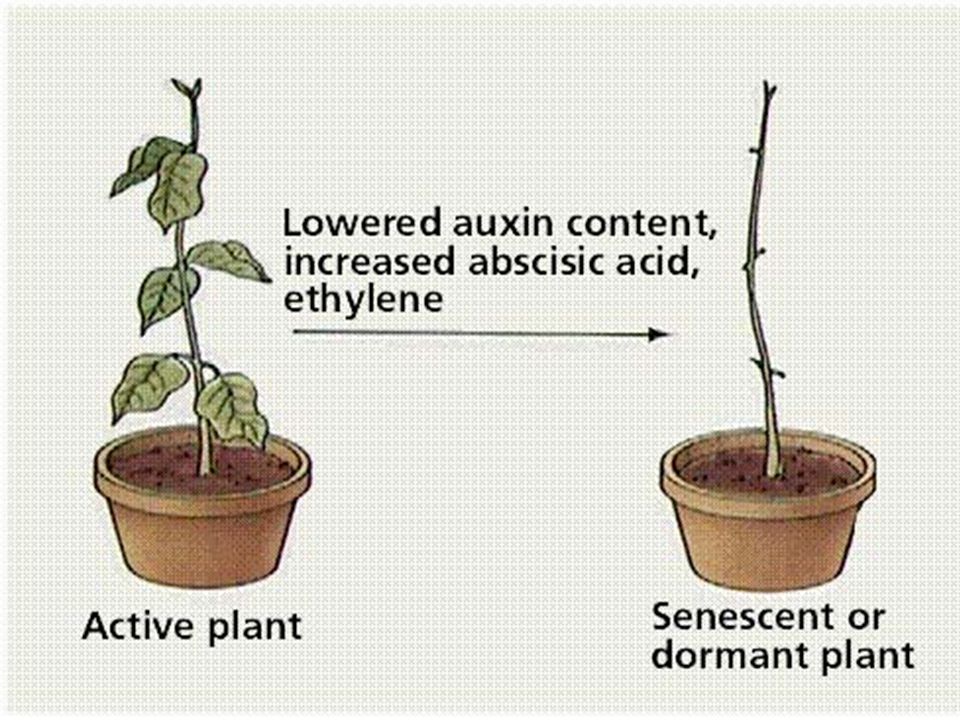
velmi významný je účinek auxinů na větvení kořenů
inhibiční vliv vegetačního vrcholu na níže položené pupeny (apikální dominance) velmi významný je účinek auxinů na větvení kořenů tvorbu adventivních kořenů oddalují stárnutí a opad listů, květů, plodů. Mají opačný účinek než cytokininy.
 velmi významný je účinek auxinů na. větvení kořenů. tvorbu adventivních kořenů. oddalují stárnutí a opad listů, květů, plodů. Mají opačný účinek než cytokininy.")
39
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
40
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
41
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
43
Auxinoidy Mezi auxinoidy patří např. kyselina b indolylmáselná (IBA), a - naftyloctová (NAA), 2,4-dichlorfenoxyoctová (2,4-D). Použití mají při zakořeňování řízků dřevin jako herbicidy při stimulaci klíčení osiva mohou zábranit předčasnému opadu plodů

44
auxinoidy

45
Gibereliny Jsou známy pouze jako nativní, vyrábějí se fermentací houby Gibberella fujikuroi Patří mezi terpenoidní sloučeniny Je jich více než 60 druhů U vyšších rostlin se jich běžně vyskytuje asi 20 Jsou označovány jako GA1, GA2-..atd.. Nejznámnější je kyselina giberelová (GA3).
.")
46
Gibereliny Syntéza giberelinů u vyšších rostlin probíhá zejména v nejmladších listech, mladých vyvíjejících se semenech a kořenech Pravděpodobně každá buňka má schopnost tvořit gibereliny Na rozdíl od auxinů jsou aktivní delší dobu, protože jejich degradace je velmi pomalá. Nepůsobí toxicky ani ve vyšších koncentracích.

47
gibereliny

50
Jejich transport je obousměrný - bazipetální i akropetální
Jejich transport je obousměrný - bazipetální i akropetální. Často se však pohybují v rostlině za zdrojem auxinů, a to bez ohledu na polaritu. Stimulují dělení buněk a hlavně prodlužovací růst zvýšením roztažitelnosti buněčné stěny - brání reakcím, které buněčnou stěnu ztužují. Stimulují dlouživý růst stonku. Vyšší koncentrace vyvolávají převahu prodlužovacího růstu nad tloustnutím.

51
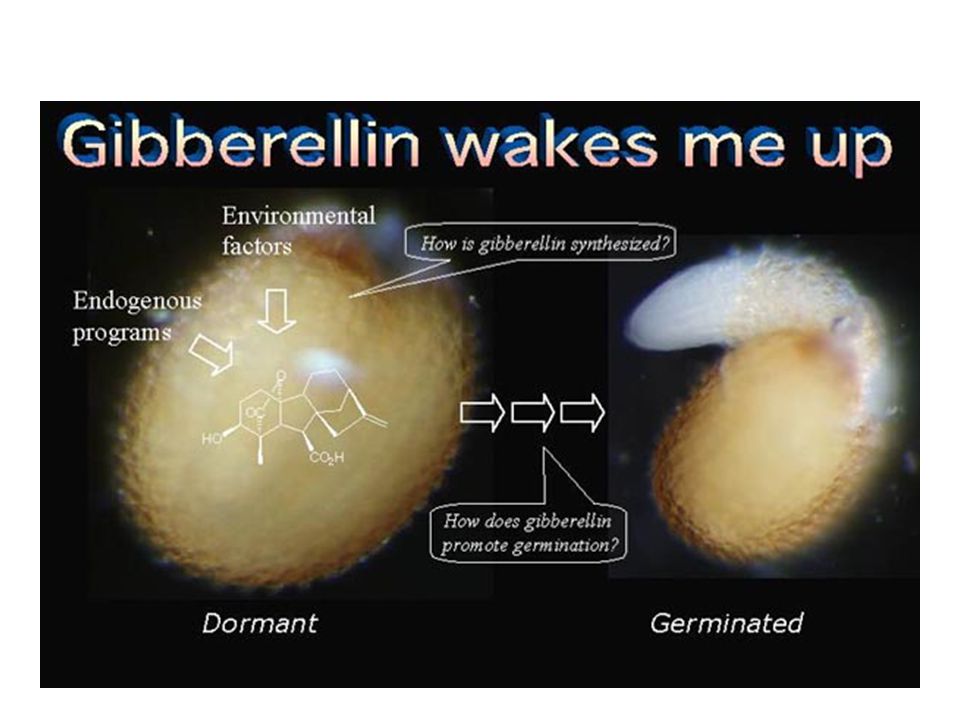
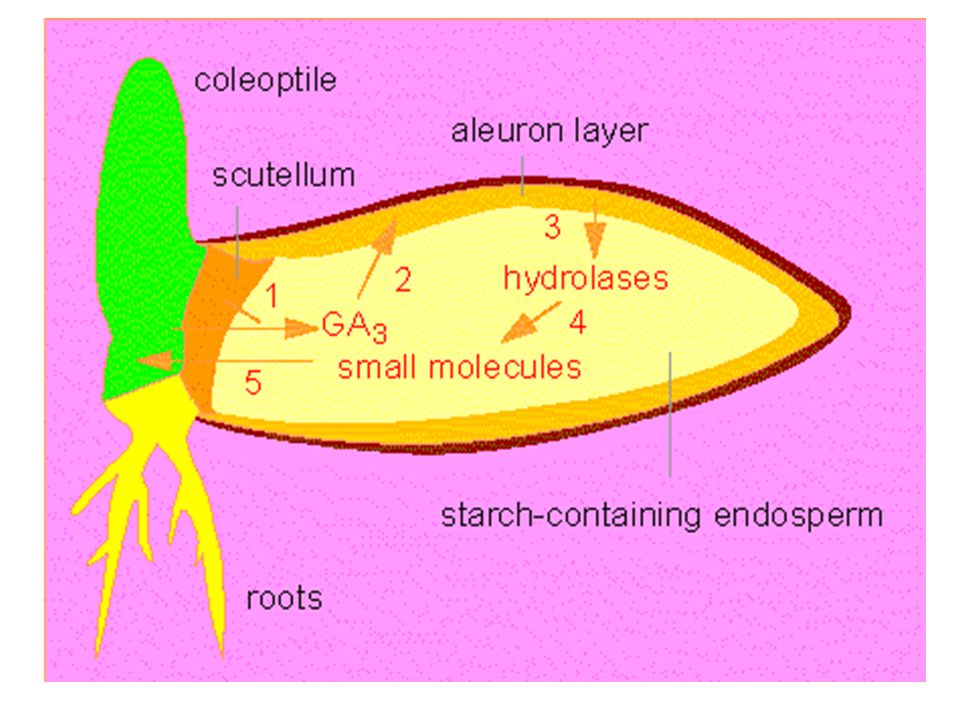
Mají rovněž důležitou úlohu při klíčení semen:
Stimulují tvorbu a vylučování hydrolytických enzymů a tím i rychlost uvolňování lehce rozložitelných cukrů (sacharózy) potřebných pro růst klíční rostlinky. Aplikací giberelinů lze přerušit dormanci semen i pupenů. Mohou tak do určité míry nahradit působení nízkých teplot. nahradit účinek dlouhého dne při indukci kvetení.
 potřebných pro růst klíční rostlinky. Aplikací giberelinů lze přerušit dormanci semen i pupenů. Mohou tak do určité míry. nahradit působení nízkých teplot. nahradit účinek dlouhého dne při indukci kvetení.")
52
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
54
Převzato z: http://www.whfreeman.com/biology (27.10. 2003)
")
57
Cytokininy

58
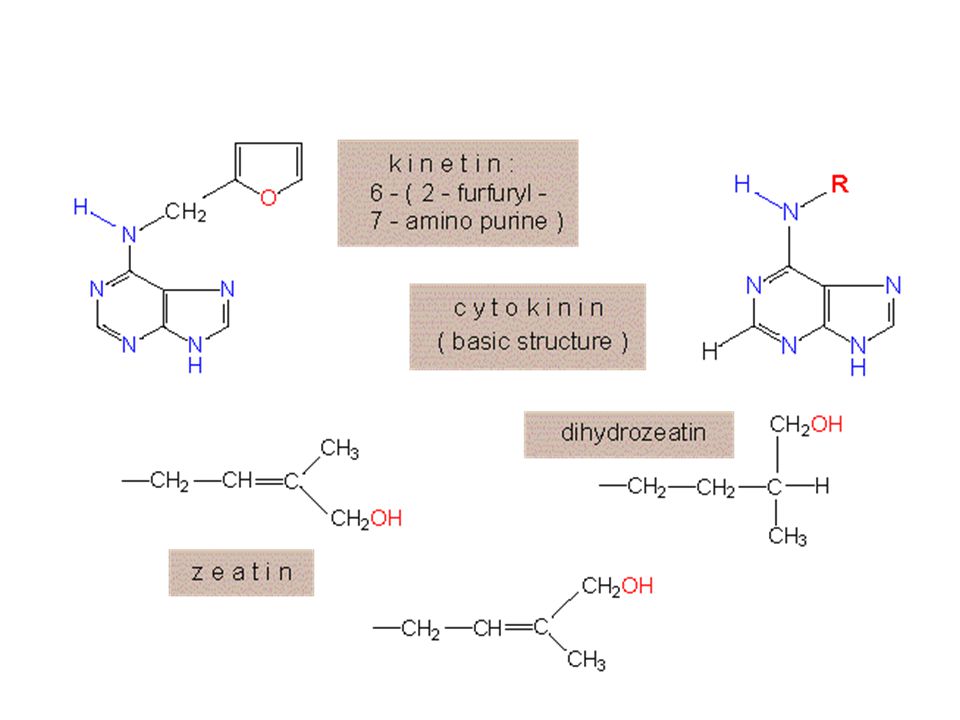
zrychlují buněčné dělení
cytokininy zrychlují buněčné dělení zřejmě stimulací tvorby proteinů. Účinek cytokininů na dělení buněk je podmíněn přítomností auxinů V současné době je známo asi 50 nativních cytokininů. Jako první byl objeven zeatin. Z chemického hlediska jsou všechny nativní cytokininy podobné, jejich základní kostru tvoří adenin.

59
cytokininy Někdy jsou mezi cytokininy řazeny i tzv. bios látky, které jsou důležité pro normální růst buněk rostlin: biotin (vitamín H) thiamin (vitamín B1) kyselina nikotinovou (vitamín PP) kyselina pantotenovou (vit. B5) Tyto látky jsou obsaženy v dělohách, pupenech, listech.
 thiamin (vitamín B1) kyselina nikotinovou (vitamín PP) kyselina pantotenovou (vit. B5) Tyto látky jsou obsaženy v dělohách, pupenech, listech.")
60
Z uměle připravovaných cytokininů jsou dnes nejpoužívanější kinetin (K)
benzylaminopurin (BAP). Cytokininy se v pletivech rostliny enzymaticky rozkládají až na močovinu. K intenzivní syntéze cytokininů dochází v kořenových špičkách, odkud jsou rozváděny xylémem do ostatních orgánů (akropetálně). Mohou vznikat i v pupenech, odkud proudí floémem bazipetálně. .
. Cytokininy se v pletivech rostliny enzymaticky rozkládají až na močovinu. K intenzivní syntéze cytokininů dochází v kořenových špičkách, odkud jsou rozváděny xylémem do ostatních orgánů (akropetálně). Mohou vznikat i v pupenech, odkud proudí floémem bazipetálně. .")
62
ABA

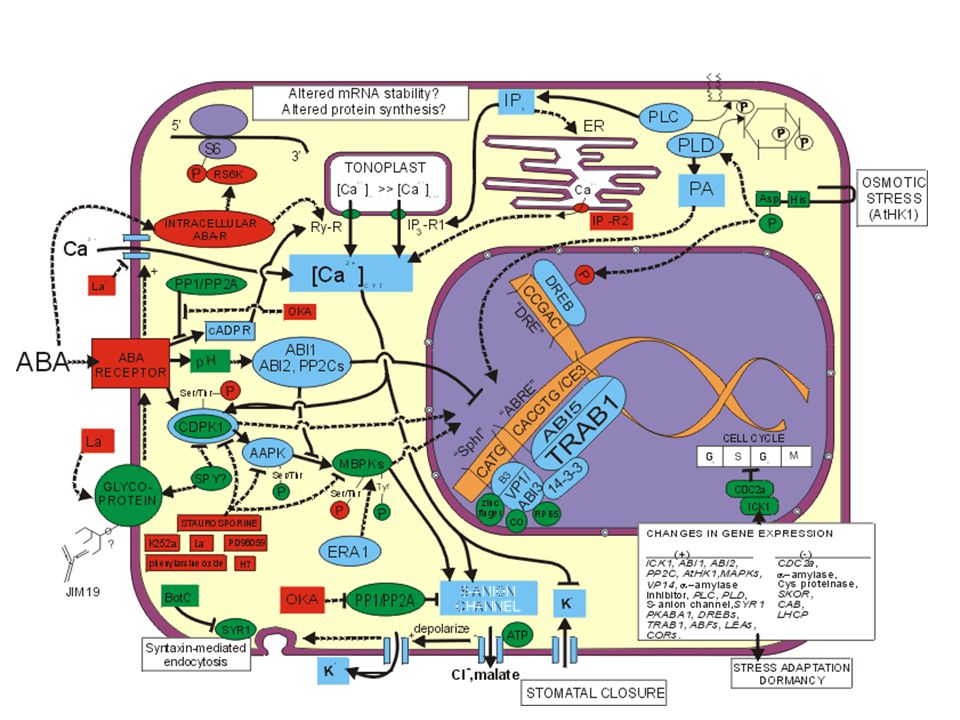
63
ABA Kyselina abscisová (Abscisic acid) je přirozeným inhibitorem rozšířeným zejména u krytosemenných rostlin Byla objevena až v roce 1963 při studiu opadu plodů bavlníku. Nejvíce je ABA syntetizována v dospělých listech v plodech a vysokou aktivitu má i v kořenovém vrcholu. ABA není toxická ani při vysokých koncentracích .

64
ABA Inhibiuje činnost protonových pump v plazmalemě, dochází tak k poklesu gradientu H, což je spojeno s pasivním výstupem K z buněk pokles jejich turgoru (následné zavírání průduchů) celková inhibice syntézy proteinů (i enzymů, např. hydroláz v semenech - zejména při navozování dormance. V tomto směru působí ABA antagonisticky s gibereliny. Nepřímo spolupůsobí na tvorbě oddělovací vrstvy mezi řapíkem a stonkem při opadu listů, květů, plodů.
 celková inhibice syntézy proteinů (i enzymů, např. hydroláz v semenech - zejména při navozování dormance. V tomto směru působí ABA antagonisticky s gibereliny. Nepřímo spolupůsobí na tvorbě oddělovací vrstvy mezi řapíkem a stonkem při opadu listů, květů, plodů.")
67
Etylén Fyziologické účinky etylénu jsou rozmanité a někdy i protichůdné zpomaluje dlouživý růst stonků i kořenů vyvolává tvorbu adventivních kořenů tvorbu odlučovací vrstvy buněk v řapících listů může stimulovat klíčení semen urychluje zrání plodů v důsledku zvýšení aktivity enzymů hydrolyzujících polysacharidy (celuláza, pektinázy) Působí na degradaci chlorofylu stimuluje biosyntézu karotenoidů. Z uměle připravených etylénotvorných látek je nejznámější 2-chlorethylfosfonová kyselina (CEPA), která dovede uvolňovat etylen do prostředí.
 Působí na degradaci chlorofylu. stimuluje biosyntézu karotenoidů. Z uměle připravených etylénotvorných látek je nejznámější 2-chlorethylfosfonová kyselina (CEPA), která dovede uvolňovat etylen do prostředí.")
68
Syntetické fyziologicky aktivní látky
CCC - (chlorcholin chlorid) se považuje za antigiberelin - omezuje růst lodyhy podporuje růst kořenů brzdí syntézu GA a auxinů (podporuje větvení, odnožování, zkracuje stéblo) TIBA - kyselina 2,3,5-trijódbenzoová antiauxin brzdí transport auxinů zeslabuje růst vrcholu podporuje větvení lodyhy urychluje tvorbu květů a hlíz
 se považuje za antigiberelin - omezuje růst lodyhy. podporuje růst kořenů. brzdí syntézu GA a auxinů (podporuje větvení, odnožování, zkracuje stéblo) TIBA - kyselina 2,3,5-trijódbenzoová antiauxin. brzdí transport auxinů. zeslabuje růst vrcholu. podporuje větvení lodyhy. urychluje tvorbu květů a hlíz.")
69
Syntetické fyziologicky aktivní látky
MH - maleinhydrazid brzdí růst Nelze použít u rostlin potravinářsky významných (retardace trávníků). ethrel - kyselina 2-chloreylfosfonová morforegulátor ovlivňuje dozrávání plodů, opadání listů a plodů, podporuje uvolňování etylenu… Nezanechává toxická rezidua.
. ethrel - kyselina 2-chloreylfosfonová. morforegulátor. ovlivňuje dozrávání plodů, opadání listů a plodů, podporuje uvolňování etylenu… Nezanechává toxická rezidua.")
Podobné prezentace