Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Organizace a kontrola eukaryotického genomu
2008 Organizace a kontrola eukaryotického genomu

2
Organizace genomu I prokaryotický genom není jen nahá DNA uložená v cytoplasmě, ale je spojena s řadou proteinů a je skládána do smyček Eukaryotický chromatin je však mnohem komplexnější DNA jednoho „průměrného“ chromosomu člověka měří 6 cm (!) DNA všech 46 lidských chromosomů měří 2 metry (!!)
 DNA všech 46 lidských chromosomů měří 2 metry (!!)")
3
Nukleosomy: „korálky na šňůře“
První úroveň sbalení DNA realizují histony. Histonů je v chromosomu přibližně stejné množství jako DNA! Histony jsou tvořeny převážně z + nabitých aminokyselin, jako je Arg a Lys, které se dobře vážou k negativně nabitým fosfátům DNA Existuje 5 typů histonů: H1, H2A, H2B, H3 a H4 Geny pro histony jsou prakticky totožné u všech eukaryot a dokonce i prokaryota mají podobné proteiny. Jedná se tedy jistě o velmi důležité a velmi konservativní geny

4
Eukaryotický genom

5
DNA eukaryotického chromosomu z vyvíjejícího se vajíčka mloka

6
Nukleosomy Nukleosom je tvořen oktamerem histonů: H2A,H2B, H3 a H4 jsou každý obsaženy dvakrát Histon H1 funguje jako svorka mezi jednotlivými nekleosomy Histony opouští DNA pouze při replikaci… …naopak při transkripci zůstávají připevněny k DNA – zdá se, že změnou tvaru a pozice nějak umožní RNA polymeráze čtení minus řetězce DNA

7
„Korálky na šňůře“ „Beads on a string“
N konec každého histonu směřuje ven z oktameru

8
DNA so obtáčí kolem histonu
prostorový model

9
Histony tradičně se myslelo, že přepisování DNA je řízeno transkripčními faktory sedající na 5´konec DNA to je ale jen část příběhu posttranslační úpravy histonů zřejmě odpovídají za regulaci genové exprese: na upravené histony nasedají proteiny, které určí, které části DNA budou přepsány RNA polymerázou (!) velmi častá je acetylace N konců (to jsou ty, které „trčí“ ven) – acetylací se zmenšuje kladný náboj a histony jsou tak náchylnější k transkripci, neboť DNA se již na ně tak pevně neváže schopnost acetylace je zřejmě klíčová pro život pokud pokusně mají kvasinky zaměněný lysin na arginin (který si zachovává kladný náboj a nemůže být acetylován), kvasinky umírají
 velmi častá je acetylace N konců (to jsou ty, které „trčí ven) – acetylací se zmenšuje kladný náboj a histony jsou tak náchylnější k transkripci, neboť DNA se již na ně tak pevně neváže. schopnost acetylace je zřejmě klíčová pro život. pokud pokusně mají kvasinky zaměněný lysin na arginin (který si zachovává kladný náboj a nemůže být acetylován), kvasinky umírají.")
10
Histony acetylace je dynamický proces
transkripční koaktivátory obsahují acetyltransferázy které histonové lysiny acetylují, korepresory mohou obsahovat deacetylázy, které acetyly odstraňují

11
Důležitost histonu H1 A: obvyklé sbalení DNA do 30nm vlákna E: bez H1 F: s H1

12
Vyšší úroveň organizace chromosomů
Nukleosomy tvoří vlákno 10nm Oproti nahé DNA nukleosomy 7x zkracují délku DNA jádro buňky člověka obsahuje 3,3x107 nukleosomů S pomocí H1 vzniká 30nm vlákno Oproti nahé DNA 30nm vlákno 40x zkracuje délku DNA Toto 30nm vlákno tvoří smyčky, které jsou připevněny k nehistonovým bílkovinám, které tvoří tzv. proteinové lešení 300nm silné Dalšími ohyby a skládáním těchto smyček vzniká chromosom – jedna chromatida je cca 700 nm silná

13
30 nm vlákno

14
300 nm vlákno

15
Chromosom

16
Chromosomy Ovšem i interfásový chromosom si udržuje jistou úroveň sbalení Předpokládá se, že v interfázi zůstává zachováno 30nm vlákno, které nehistonové proteiny váží k nukleární lamině a snad i k nukleární matrix Toto připevnění tak dává každému z interfázových chromosomů své přesné místo v jádře a zabraňuje tak zamotání jednotlivých chromosomů

17
Heterochromatin a euchromatin
I u interfázových chromosomů se některé jejich části nachází ve velmi kondensovaném stavu, tak že jsou barvitelné a pozorovatelné mikroskopem – jedná se o tzv. heterochromatin. Heterochromatinová DNA není přepisována Euchromatin = „pravý chromatin“ je přepisován a nachází se v mnohem více rozvolněném stavu

18
Odlišní exprese genů u odlišných buněk
všechny buňky našeho těla vznikly mitózou, první buňkou byla zygota (téměř) všechny buňky našeho těla obsahují stejný genom jak je tedy možné, že se naše tělo skládá z cca 200 typů buněk? (svalové, nervové…?) typická lidská buňka přepisuje v daném čase jen asi 20 % svých genů velmi diferencované buňky, jako jsou svalové buňky,přepisují dokonce ještě menší procento genů
 všechny buňky našeho těla obsahují stejný genom. jak je tedy možné, že se naše tělo skládá z cca 200 typů buněk (svalové, nervové… ) typická lidská buňka přepisuje v daném čase jen asi 20 % svých genů. velmi diferencované buňky, jako jsou svalové buňky,přepisují dokonce ještě menší procento genů.")
19
Odlišní exprese genů u odlišných buněk
jednotlivé buňky se tedy od sebe liší ani ne tak tím, že by obsahovaly odlišné geny, nýbrž tím, že odlišné geny jsou exprimovány otázkou tedy jest, jak může RNA polymeráza najít v nezměrném moři písmen začátek správného genu, který má v této buňce v tomto čase přepsat

20
Geny „housekeeping genes“ – geny, které se přepisují ve všech buňkách
patří sem např. geny pro histony, geny pro ribosomální proteiny atd.

21
v každé z těchto oblastí může proběhnout regulace genové exprese

22
Kontrola genové exprese

23
Kontrola genové exprese
Organizace chromatinu v jádře DNA metylace. Zejména cytosin. Asi 5% cytosinů lidské DNA je metylováno. Velmi je metylována DNA neaktivního X chromosomu (Barrova tělíska). Metylované geny se předávají v jednotlivých mitózách dál, protože metylační enzymy jsou schopny je rozeznat a metylují dceřinný řetězec při replikaci – tapisérie metylovaných genů se tak při růstu tkáně předává dál. Metylace rovněž hraje roli při genovém imprintingu.
. Metylované geny se předávají v jednotlivých mitózách dál, protože metylační enzymy jsou schopny je rozeznat a metylují dceřinný řetězec při replikaci – tapisérie metylovaných genů se tak při růstu tkáně předává dál. Metylace rovněž hraje roli při genovém imprintingu.")
24
Kontrola iniciace transkripce
transkripce začíná vytvořením transkripčně iniciačního komplexu = transkripční faktory + promotor + RNA polymeráza kontrolní elementy = segmenty nekódující DNA, na které se navazují transkripční faktory (striktně vzato je i promotor kontrolním elementem)
")
25
Kontrola iniciace transkripce

26
Role transkripčních faktorů
RNA polymeráza nemůže sama zahájit transkripci jen jeden transkripční faktor rozeznává TATA box, jiné rozeznávají proteiny - buď jeden druhý nebo RNA polymerázu takto ale probíhá transkripce jen pomalu a nedokonale klíčem k rychlé transkripci jsou kontrolní elementy

27
Posilovače transkripce - enhancery
enhancery mohou být až tisíce pb vzdáleny od promotory proti proudu nebo i po proudu mohou se dokonce nacházet i v intronech do kontaktu s promotorem se dostávají ohybem DNA aktivátor = transkripční faktor, který se váže na enhancer a popoří transkripci daného genu

28
Posilovače transkripce

29
Vazba transkripčních faktorů k DNA

30
Posttranskripční mechanismy kontrolují genovou expresi
mezi syntézou primárního transkriptu a činností aktivního proteinu v buňce se může nacházet řada kontrolních kroků může např. dojít k alternativnímu sestřihu

31
Regulace degradace mRNA
prokaryotické mRNA mají typicky velmi krátkou životnost a enzymy je degradují po několika minutách - tím je také způsobeno, že bakterie pružně a rychle reagují na měnící se životní podmínky u eukaryot mRNA přetrvává typicky hodiny, ale i dny a často i týdny pověstná dlouhým životem je např mRNA pro oba globinové řetězce

32
Regulace degradace mRNA
degradace mRNA začíná enzymatickým zkracováním poly(A) konce to rozběhne aktivitu enzymů, které odstraní čepičku po odstranění čepičky nukleázy rychle odbourají mRNA od 5´ konce
 konce. to rozběhne aktivitu enzymů, které odstraní čepičku. po odstranění čepičky nukleázy rychle odbourají mRNA od 5´ konce.")
33
Kontrola translace většinou je regulována iniciace translace (připojení mRNA na malou podjednotku, připojení tRNAMet a připojení velké podjednotky) např. v oocytu je velké množství předem připravených mRNA, které se po fertilizaci překládají ve správném čase a správném pořadí. některé faktory zastavují translaci všech mRNA. Pokud např. ve vznikajícím erytrocytu je málo hemů, rewgulační proteiny zastavují celou translaci - většina mRNA je ovšem mRNA pro globiny
 např. v oocytu je velké množství předem připravených mRNA, které se po fertilizaci překládají ve správném čase a správném pořadí. některé faktory zastavují translaci všech mRNA. Pokud např. ve vznikajícím erytrocytu je málo hemů, rewgulační proteiny zastavují celou translaci - většina mRNA je ovšem mRNA pro globiny.")
34
Kontrola translace v oocytech mnoha organismů je připraveno množství mRNA - teprve po fertilizaci dojde k rychlé aktivaci translačních iniciačních faktorů. Výsledkem je explozívíní přepis mnoha mRNA některé řasy a rostliny za noci zastavují translaci, kterou aktivuje až světlo

35
Posttranslační úpravy proteinů a jejich degradace
z insulinu musí být vyštěpena část řetězce, na jiné proteiny musí být navěšena sacharidová skupina, jiné proteiny mohou být aktivovány/deaktivovány reversibilním připojením fosfátové skupiny, jiné musí být dopraveny na správné místo v buňce - zde všude může probíhat regulace

36
Posttranslační úpravy proteinů a jejich degradace
např. cystická fibrosa je způsobena tím, že (díky mutaci v genu) protein, který má být normálně membránový a obsahuje v sobě kanál pro transport chloridových iontů, se nedostane na své místo do plasmatické membrány a v cytoplasmě je rychle degradován např. cykliny musí mít krátkou životnost a musí být rychle degradovány, aby pracovaly správně
 protein, který má být normálně membránový a obsahuje v sobě kanál pro transport chloridových iontů, se nedostane na své místo do plasmatické membrány a v cytoplasmě je rychle degradován. např. cykliny musí mít krátkou životnost a musí být rychle degradovány, aby pracovaly správně.")
37
Posttranslační úpravy proteinů a jejich degradace
degradace začíná připevněním malé molekuly zvané ubiquitin na protein obrovský proteinový komplex, zvaný proteasom, pak rozezná ubquitinovou „visačku“ a degraduje označený protein cykliny, které vzdorují degradaci mohou způsobit rakovinu

38
Proteasom degraduje proteiny

39
Organizace genomu na úrovni DNA
U prokaryot většina DNA kóduje proteiny a jednotlivé geny jsou odděleny jen krátkými úseky nekódující DNA U eukaryot naopak jen asi 1,5% (!) DNA kóduje proteiny další malé procento kóduje nějaký jiný typ RNA než mRNA Zbytek - nekódující DNA u eukaryot je tvořena částečně introny, ale především tzv. repetitivní DNA
 DNA kóduje proteiny. další malé procento kóduje nějaký jiný typ RNA než mRNA. Zbytek - nekódující DNA u eukaryot je tvořena částečně introny, ale především tzv. repetitivní DNA.")
40
Tandemové repetice Tandemové repetice jsou krátké sekvence nukleotidů, opakované mnohokrát za sebou, např. GTTACGTTACGTTACGTTACGTTACGTTAC Sekvence GTTAC se může až několik set tisíckrát opakovat – evoluční význam tohoto jevu (je-li jaký) je záhadou. Možná se jedná jen o „odpadní“ DNA Počet nukleotidů v sekvenci je obvykle
 je záhadou. Možná se jedná jen o „odpadní DNA. Počet nukleotidů v sekvenci je obvykle")
41
Tandemové repetice Díky tomu, že tandemové repetice se opakují tolikrát za sebou, má tata část chromosomu poněkud jiné složení CG a AT než ostatní části chromosomu Pokud je chromosom rozbit na malé kousky, tandemová DNA při centrifugaci vytvoří v centrifugační zkumavce pruh v poněkud jiné výšce, než zbytek chromosomu Proto se této DNA začalo říkat „satelitní DNA“ Nyní je tento termín užíván pro všechnu tandemovou DNA

43
Satelitní DNA Satelitní DNA se dělí na tři typy:
Satelitní DNA ( – 10 milionů opakování) Minisatelitní DNA (100 – opakování) Mikrosatelitní DNA (10 – 100 opakování)
 Minisatelitní DNA (100 – opakování) Mikrosatelitní DNA (10 – 100 opakování)")
44
Satelitní DNA Některé genetické choroby jsou způsobeny nesprávným počtem repetic Fragilní X chromosom = mentální retardace. Počet repetic CGG na nepřekládaném začátku genu je u zdravých osob přibližně 30. U nemocných se tento triplet vyskytuje ve stovkách nebo i tisících opakováních. Počet opakování se zvyšuje v průběhu generací Huntingtonova choroba: zde je triplet CAG překládán (do glutaminu). U zdravých osob je počet repeticí do 35, u nemocných 40 – několik set
. U zdravých osob je počet repeticí do 35, u nemocných 40 – několik set.")
45
Satelitní DNA Mnoho satelitní DNA se nachází v telomerách a v centromeře

46
Vmezeřená repetitivní DNA Intrespersed Repetitive DNA
Tvoří 25% - 40% DNA savců Repetice zde nenásledují těsně za sebou, ale jsou rozptýleny na různých místech genomu Repetice je obvykle dlouhá stovky až tisíce pb a její jednotlivé kopie se obvykle mírně odlišují Alu elementy: tvoří 5% genomu člověka. Jedná se o 300pb dlouhou repetici. Výjimka: Ale elementy se překládají do proteinu. Význam tohoto proteinu v buňce, je-li jaký, není znám

47
Genové rodiny U prokaryot i u eukaryot se většina genů vyskytuje jako unikátní sekvence DNA, v jediném provedení v haploidním genomu Některé geny se však vyskytují ve více než jednom provedení. Soubor stejných nebo velmi podobných genů se nazývá multigenová rodina Genovou rodinu můžeme chápat jako tandemovou repetici s velmi dlouhou (=jeden gen) repetiční jednotkou
 repetiční jednotkou.")
48
Genové rodiny Některé genové rodiny obsahují identické geny tandemově uspořádané - např. geny pro histony Většinou však genové rodiny neobsahují geny pro bílkoviny, ale pro RNA Za příklad mohou posloužit rodiny pro rRNA – zde jedna jednotka obshuje geny pro tři rRNA (18S, 5,8S a 28S rRNA) Tato jednotka se opakuje ve stovkách – tisících kopiích. Primární transkript je štěpen na tři rRNA. Takto má buňka materiál pro miliony ribosomů
 Tato jednotka se opakuje ve stovkách – tisících kopiích. Primární transkript je štěpen na tři rRNA. Takto má buňka materiál pro miliony ribosomů.")
49
Genové rodiny

50
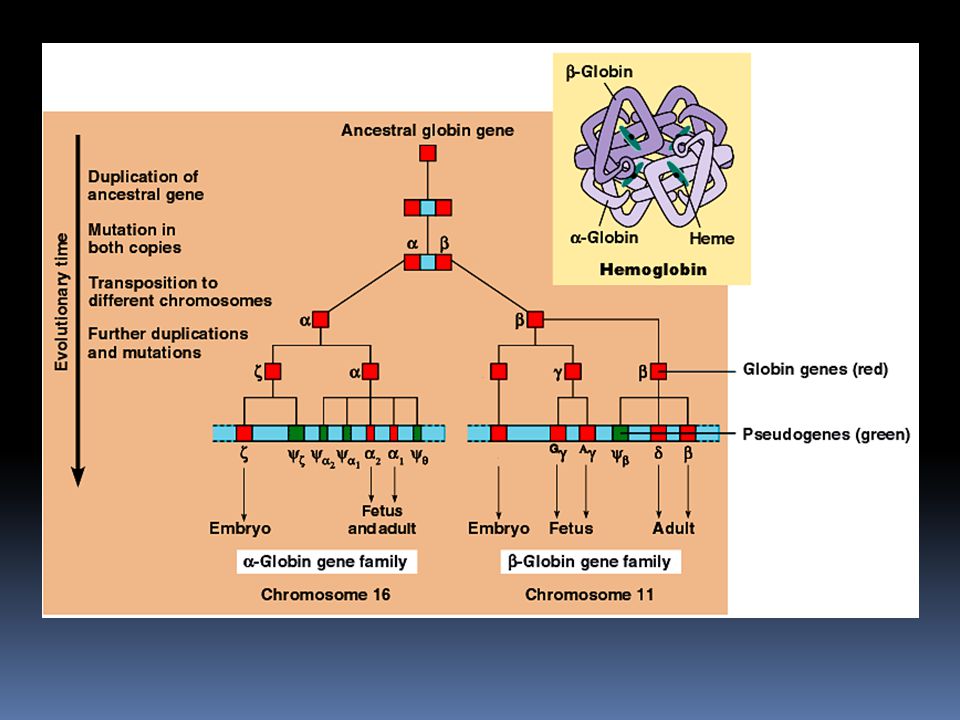
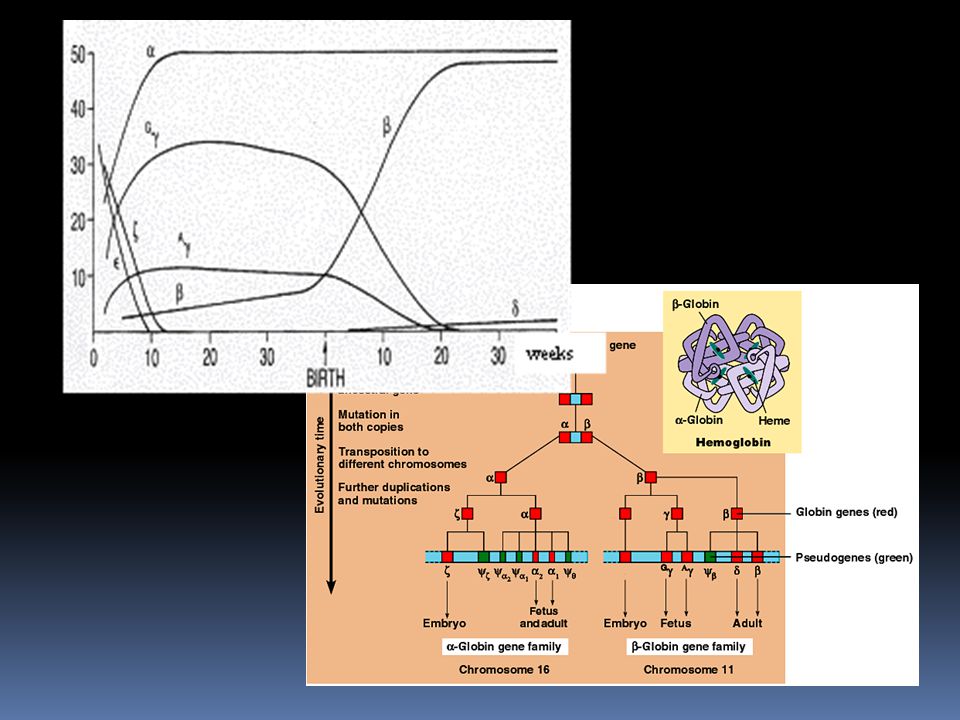
Multigenové rodiny Klasickým příkladem jsou geny pro globiny – proteiny, ze kterých se sestavuje hemoglobin Rodina na chromosomu 16 obsahuje geny kódující různé verze α-globinu Rodina na chromosomu 11 obsahuje geny kódující různé verze β-globinu Různé verze globinových genů jsou přepisovány v různých fázích embryonálního a fetálního vývoje

53
Globinová rodina Nejpravděpodobnější vznik je opakovaným zdvojením původně jednoho genu Rozdíly mezi jednotlivými geny pravděpodobně vznikly nahromaděním mutací v průběhu evoluce Pseudogeny: útvar velmi podobný genu, ale buď se nepřekládá, nebo kóduje nefunkční produkt. Globinová rodina obsahuje několik pseudogenů

54
Transpozony a retrotranspozony
Transpozon = sekvence DNA, která se může přesunout z jednoho umístění na genomu do jiného Pokud transpozon „skočí“ doprostřed jného genu, může jej vyřadit z činnnosti Trasnpozony tvoří 50% genomu kukuřice a cca 10% genomu člověka Většina transpozonů jsou retrotranspozony

55
Např. Alu elementy jsou retrotranspozony
Např. Alu elementy jsou retrotranspozony. U člověka aktivní transpozony nejsou známy.

56
Kontrola genové exprese
V průběhu buněčné diferenciace jsou zapínány a vypínány různé geny – výsledkem je, že se jedna buňka liší od druhé, např. neuron od svalové buňky Typická buňka přepisuje v daném čase jen asi 3-5% svých genů Enzymy přepisující DNA (RNA polymeráza) musí najít správné geny ve správném čase
 musí najít správné geny ve správném čase.")
57
Molekulární biologie rakoviny

58
Protoonkogeny rakovina vzniká poškozením genů kontrolující buněčný růst a dělení, a to buď fyzikálními nebo chemickými agens (mutageny, X-zářením atd.) protoonkogen = gen, který stimuluje normální buněčný růst a dělení onkogen = gen, který způsobuje rakovinu onco = řec. tumor, nádor
 protoonkogen = gen, který stimuluje normální buněčný růst a dělení. onkogen = gen, který způsobuje rakovinu. onco = řec. tumor, nádor.")
59
Změna protoonkogenu na onkogen
přesun genu na jiné místo v genomu amplifikace (zmnožení) protoonkogenu bodová mutace v protoonkogenu
 protoonkogenu. bodová mutace v protoonkogenu.")
60
Změna protoonkogenu na onkogen

61
Změna protoonkogenu na onkogen
Maligní buňky mají často chromosomové translokace, díky kterým se dostávají protoonkogeny do blízkosti silných promotorů a jsou přepisovány častěji. Jindy je to promotor, který se přesune do blízkosti protoonkogenu mutace může způsobit, že protein je více aktivní nebo více odolný vůči degradaci

62
Tumor supresorové geny
translační produkty těchto genů normálně pomáhají zastavit nekontrolovatelný růst translační produkty tumor supresorových genů mají normálně v buňce různé úkoly: opravují poškozenou DNA kontrolují adhezi buněk k sobě či k extracelulární matrix jiné přímo inhibují buněčný cyklus

63
ras protoonkogen a p53 tumor supresorový gen
30% rakovin člověka má poškozený ras gen 50% rakovin člověka má poškozený p53 gen Ras protein i p53 protein jsou součástmi drah, kterými je z cytoplasmy ovlivněna aktivita jádra

64
Ras protein a p53 protein

65
Ras protein jedná se o tzv G protein (aktivovaný GTP), který je součástí dráhy, předávající signál růstového faktory přes celou cytoplasmu až do jádra normálně je dráha spuštěna růstovým faktorem mutovaný ras protein spouští dráhu i v nepřítomnosti růstového faktoru
, který je součástí dráhy, předávající signál růstového faktory přes celou cytoplasmu až do jádra. normálně je dráha spuštěna růstovým faktorem. mutovaný ras protein spouští dráhu i v nepřítomnosti růstového faktoru.")
66
p53 protein „guardian angel of the genome“
je tumor supresorový protein o hmotnosti Daltonů p53 je trasnkripční faktor, který stimuluje syntézu proteinů inhibujících růst buňky: aktivuje gen p21, jehož proteinový produkt se váže k cyklin-dependentním kinázám, čímž je získán čas pro opravu DNA aktivuje geny, jejichž proteiny opravují DNA pokud je DNA neopravitelná, aktivuje „sebevražedné“ geny, které rozběhnou apoptózu

67
Rakovinu způsobí až mnohonásobná mutace
…což vysvětluje, že rakovin přibývá s věkem příklad: kolorektální karcinom nových případů v USA každý rok nejprve vzniká polyp, tvořený „normálními“ buňkami, které se jen rychleji dělí tumor roste a může se stát maligním - tak, že se v jeho buňkách hromadí mutace tvořící onkogeny a knock-outované tumor supresorové geny

68
Rakovinu způsobí až mnohonásobná mutace
tumor supresorové alely jsou většinou recesívní - musí tedy zmutovat obě naopak onkogeny jsou většinou dominantní - stačí, aby zmutovala jedna nakonec je aktivován gen pro telomerázu

69
Rakovina tlustého střeva

70
Další příčiny vzniku rakoviny
viry: asi 15% případů retroviry způsobují leukémie viry hepatitidy způsobují rakovinu jater způsobují rakovinu děložního hrdla buď přinesou s sebou onkogen, nebo se vloží do tumor supresorového genu, nebo aktivují protoonkogen

71
Dědičnost rakoviny rakovina není dědičná obvyklou mendelovskou dědičností dědičná může být např. jedna mutace tumor supresorového genu - potomstvo je tedy o jeden krok blíže k vypuknutí rakoviny 15% kolorektální rakoviny obsahuje již zděděné mutace, řada z nich zasahuje geny produkující proteiny opravující DNA

72
Rakovina prsu v USA 180 000 žen (!) každý rok
u 5%-10% silná dědičná disposice zejména geny BRCA1 a BRCA2 (BReast CAncer) - jejich mutace podporuje vznik rakoviny prsu i vaječníků. Oba jsou možná tumor supresorové a jsou recesívní. Snad se i podílejí na opravě DNA
 - jejich mutace podporuje vznik rakoviny prsu i vaječníků. Oba jsou možná tumor supresorové a jsou recesívní. Snad se i podílejí na opravě DNA.")
73
Rakovina prsu

74
S díky za pozornost Orko

Podobné prezentace


>")





>")
>")










