Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

2
Životní strategie – life histories
Všechny organismy se v čase nějakým způsobem vyvíjejí – rodí se, rostou, mění se, rozmnožují se. Otázkou je, zda-li v těchto životních procesech můžeme vysledovat nějaké obecné vzory, a zda-li mají tyto obecné (či společné) vzory z hlediska ekologie, respektive z hlediska působení přírodního výběru, nějaké vysvětlení. Při hledání odpovědí musíme mít na paměti, že přírodní výběr je z jedné strany omezen tím, co má k dispozici, že působí v určitých mantinelech daných předchozím vývojem, ale že z druhé strany, ač jsou možnosti toho kterého organismu omezeny možnostmi genotypu, mohou různí jedinci jednoho druhu reagovat za různých okolností různě, tj. že v rámci genotypu existuje plasticita. Pro přiblížení příklad: životní strategie letniček může být popsána charakteristickým počtem semen pro daný druh (genotyp) tím, že tento počet se v období sucha snižuje (plasticita) ale že míra poklesu produkce semen je opět kontrolována geneticky (např. některé druhy sníží počet semen, ale jiné počet zachovají, ale zmenší jejich velikost)
 vzory z hlediska ekologie, respektive z hlediska působení přírodního výběru, nějaké vysvětlení. Při hledání odpovědí musíme mít na paměti, že přírodní výběr je z jedné strany omezen tím, co má k dispozici, že působí v určitých mantinelech daných předchozím vývojem, ale že z druhé strany, ač jsou možnosti toho kterého organismu omezeny možnostmi genotypu, mohou různí jedinci jednoho druhu reagovat za různých okolností různě, tj. že v rámci genotypu existuje plasticita. Pro přiblížení příklad: životní strategie letniček může být popsána. charakteristickým počtem semen pro daný druh (genotyp) tím, že tento počet se v období sucha snižuje (plasticita) ale že míra poklesu produkce semen je opět kontrolována geneticky (např. některé druhy sníží počet semen, ale jiné počet zachovají, ale zmenší jejich velikost)")
3
Životní strategie - komponenty
(komponenty musí ovlivňovat fitness, tj. reprodukci a přežívání) Velikost je jedním z nejnápadnějších aspektů životních strategií. Liší se mezi taxony, mezi populacemi stejného druhu, mezi jedinci jedné populace, a mění se i během ontogenetického vývoje jedince samotného. Vnitrodruhová variabilita je patrná zejména u modulárních organismů. Velikost může například ovlivňovat kompetiční úspěšnost organismu, jeho menší zranitelnost vůči predátorům, a nebo naopak jeho efektivitu v roli predátora. Větší organismy mohou lépe udržovat konstantnost životních funkcí vzhledem k vnějšímu prostředí (mají vůči prostředí exponován relativně menší povrch). V rámci jednoho druhu větší organismy obyčejně produkují více mláďat. (click). Na druhou stranu větší velikost může přinášet i rizika: větší strom je zranitelnější ve vichřici nebo vůči bleskům, známe i mnoho případů, kdy predátor preferuje větší kořist oproti stejné, ale menší. Větší organismy také potřebují (v absolutních jednotkách) více energie pro růst a reprodukci – proto při omezených zdrojích mohou trpět více než menší. Někdy z toho všeho vychází, že nejlepší je být „někde uprostřed“. (click) Určité velikosti lze během života dosáhnout různými cestami: narodit se už velký, nebo rychle růst, nebo růst dlouho, a samozřejmě toto vše různě kombinovat. Rychlý vývoj může vést k brzkému rozmnožování, ke krátkým generační dobám a tedy k vysoké reprodukční rychlosti = zvýšení fitness. Na druhou stranu, potlačený vývoj (dormance, diapauza, apod.) může zvyšovat šance organismu na přežití za nepříznivých podmínek.
 Velikost je jedním z nejnápadnějších aspektů životních strategií. Liší se mezi taxony, mezi populacemi stejného druhu, mezi jedinci jedné populace, a mění se i během ontogenetického vývoje jedince samotného. Vnitrodruhová variabilita je patrná zejména u modulárních organismů. Velikost může například ovlivňovat kompetiční úspěšnost organismu, jeho menší zranitelnost vůči predátorům, a nebo naopak jeho efektivitu v roli predátora. Větší organismy mohou lépe udržovat konstantnost životních funkcí vzhledem k vnějšímu prostředí (mají vůči prostředí exponován relativně menší povrch). V rámci jednoho druhu větší organismy obyčejně produkují více mláďat. (click). Na druhou stranu větší velikost může přinášet i rizika: větší strom je zranitelnější ve vichřici nebo vůči bleskům, známe i mnoho případů, kdy predátor preferuje větší kořist oproti stejné, ale menší. Větší organismy také potřebují (v absolutních jednotkách) více energie pro růst a reprodukci – proto při omezených zdrojích mohou trpět více než menší. Někdy z toho všeho vychází, že nejlepší je být „někde uprostřed . (click) Určité velikosti lze během života dosáhnout různými cestami: narodit se už velký, nebo rychle růst, nebo růst dlouho, a samozřejmě toto vše různě kombinovat. Rychlý vývoj může vést k brzkému rozmnožování, ke krátkým generační dobám a tedy k vysoké reprodukční rychlosti = zvýšení fitness. Na druhou stranu, potlačený vývoj (dormance, diapauza, apod.) může zvyšovat šance organismu na přežití za nepříznivých podmínek.")
4
Životní strategie - komponenty
Rozmnožovaní (a velikost) Organismy se mohou lišit v délce pre-reprodukčního vývoje. Dále se liší ve četnosti rozmnožování (krátce a jednou – semelparie) nebo častěji, kdy se mezi jednotlivými reprodukčními epizodami udržují v takové kondici, aby příště mohli znovu (= iteroparie). V tomto případě se navíc mohou lišit snůšky – jejich počet i počet potomků v nich. Jednotliví potomci se mohou lišit ve velikosti – to záleží mj. na tom, kolik zdrojů (energie) do nich investovali rodiče. To může záviset třeba na tom, jestli má vajíčko velké žloutkové zásoby, nebo jestli je embryo vyživováno placentou (nebo nějak podobně). Jistě sem patří i množství zdrojů/energie investovaných do postnatální péče. Snaha toto vše zkombinovat vede k vyjádření relativní míry investic do reprodukce – reprodukční alokace nebo též reprodukční úsilí. Můžeme ji definovat jako ‚podíl dostupných zdrojů investovaný do rozmnožování během určité časové jednotky‘. Bohužel, je to mnohem snazší definovat než měřit. Ideální by bylo možné zdroje identifikovat a sledovat jejich rozdělování do tkání a přeměny ve fyziologických pochodech sledovaného organismu. V praxi se to ovšem omezuje většinou na biomasu či (lépe) energii – technika měření je mnohem snazší. Viz další slide.
 Organismy se mohou lišit v délce pre-reprodukčního vývoje. Dále se liší ve četnosti rozmnožování (krátce a jednou – semelparie) nebo častěji, kdy se mezi jednotlivými reprodukčními epizodami udržují v takové kondici, aby příště mohli znovu (= iteroparie). V tomto případě se navíc mohou lišit snůšky – jejich počet i počet potomků v nich. Jednotliví potomci se mohou lišit ve velikosti – to záleží mj. na tom, kolik zdrojů (energie) do nich investovali rodiče. To může záviset třeba na tom, jestli má vajíčko velké žloutkové zásoby, nebo jestli je embryo vyživováno placentou (nebo nějak podobně). Jistě sem patří i množství zdrojů/energie investovaných do postnatální péče. Snaha toto vše zkombinovat vede k vyjádření relativní míry investic do reprodukce – reprodukční alokace nebo též reprodukční úsilí. Můžeme ji definovat jako ‚podíl dostupných zdrojů investovaný do rozmnožování během určité časové jednotky‘. Bohužel, je to mnohem snazší definovat než měřit. Ideální by bylo možné zdroje identifikovat a sledovat jejich rozdělování do tkání a přeměny ve fyziologických pochodech sledovaného organismu. V praxi se to ovšem omezuje většinou na biomasu či (lépe) energii – technika měření je mnohem snazší. Viz další slide.")
5
reprodukční alokace

6
Životní strategie - komponenty
Pokud dojde ve studiích na měření reprodukční alokace, počítá se s tím, že se zdroje nějak rozdělují (aniž bychom věděli, jaké zdroje to jsou) – proto jsou častým vyjádřením reprodukční alokace poměry typu hmotnost gonád x hmotnost těla nebo hmotnost semen x biomasa rostliny nebo velikost snůšky x hmotnost těla. Ale je to jen z nouze ctnost. Není od věci počítat do komponent životních strategií též alokaci zdrojů/energie do zásob. Ty se vyplatí hlavně tam, kde organismus musí přečkávat období, kdy je zdroje nedostatek.
 – proto jsou častým vyjádřením reprodukční alokace poměry typu hmotnost gonád x hmotnost těla nebo hmotnost semen x biomasa rostliny nebo velikost snůšky x hmotnost těla. Ale je to jen z nouze ctnost. Není od věci počítat do komponent životních strategií též alokaci zdrojů/energie do zásob. Ty se vyplatí hlavně tam, kde organismus musí přečkávat období, kdy je zdroje nedostatek.")
7
REPRODUKČNÍ HODNOTA (RV)
Přírodní výběr favorizuje ty organismy, jejichž příspěvek do budoucnosti populace je relativně (vůči ostatním v populaci) nejvyšší. Z toho se vracíme k otázce – Jak vlastně měřit fitness? Nejlépe z life tables, ale to vždy nejde, a navíc je i tam více možností. Co třeba růstovými rychlostmi? Nebo jiní přísahají na reprodukční hodnotu. V zásadě jde o parametr stejného druhu jako ty dříve zmiňované; jejich podobnost je daleko větší než vzájemné rozdíly. Reprodukční hodnota vychází z parametrů známých z life tables, a pro jedince v daném stadiu (daného věku) ji lze stručně popsat jako součet již vyprodukovaných potomků s (pravděpodobným) budoucím reprodukčním příspěvkem, tzv. reziduální reprodukční hodnotou (RRV). RRV v sobě kombinuje očekávanou budoucí délku života a očekávanou budoucí fekunditu, a to tak, že bere do úvahy příspěvek jedince do budoucích generací relativně vůči příspěvkům ostatních jedinců. Přírodním výběr bude favorizovat takovou životní strategii, jež má součet již realizovaného a budoucího reprodukčního příspěvku nejvyšší.
 nejvyšší. Z toho se vracíme k otázce – Jak vlastně měřit fitness Nejlépe z life tables, ale to vždy nejde, a navíc je i tam více možností. Co třeba růstovými rychlostmi Nebo jiní přísahají na reprodukční hodnotu. V zásadě jde o parametr stejného druhu jako ty dříve zmiňované; jejich podobnost je daleko větší než vzájemné rozdíly. Reprodukční hodnota vychází z parametrů známých z life tables, a pro jedince v daném stadiu (daného věku) ji lze stručně popsat jako. součet již vyprodukovaných potomků s (pravděpodobným) budoucím reprodukčním příspěvkem, tzv. reziduální reprodukční hodnotou (RRV). RRV v sobě kombinuje očekávanou budoucí délku života a očekávanou budoucí fekunditu, a to tak, že bere do úvahy příspěvek jedince do budoucích generací relativně vůči příspěvkům ostatních jedinců. Přírodním výběr bude favorizovat takovou životní strategii, jež má součet již realizovaného a budoucího reprodukčního příspěvku nejvyšší.")
8
REPRODUKČNÍ HODNOTA (RV)
parametry rovnice známe z life tables. První člen rovnice (mx) označuje současný počet potomků, druhý člen rovnice pak RRV. Přitom ly/lx (. my) je pravděpodobný počet potomků ve věku y, je to tedy plodnost, které by jednotlivec dosáhl, kdyby se dožil daného věku, násobená pravděpodobností, že se mu to podaří. člen Rx-y je čistá reprodukční rychlost celé populace za jednotku času za časový interval změny z věku x na x+1, modifikovaná pro případ, že populace roste, tedy R>1, Rx-y<1, protože x < y, nebo klesá (opak). Pokud roste, tak protože jedinců je víc a víc směrem k času y, relativní příspěvek konkrétního jedince v čase od x k y klesá. Naopak, pokud klesá (tedy R < 1, Rx-y>1), jedinců v budoucnosti je méně a relativní příspěvek konkrétního jedince v čase od x k y roste.
 označuje současný počet potomků, druhý člen rovnice pak RRV. Přitom ly/lx (. my) je pravděpodobný počet potomků ve věku y, je to tedy plodnost, které by jednotlivec dosáhl, kdyby se dožil daného věku, násobená pravděpodobností, že se mu to podaří. člen Rx-y je čistá reprodukční rychlost celé populace za jednotku času za časový interval změny z věku x na x+1, modifikovaná pro případ, že populace roste, tedy R>1, Rx-y<1, protože x < y, nebo klesá (opak). Pokud roste, tak protože jedinců je víc a víc směrem k času y, relativní příspěvek konkrétního jedince v čase od x k y klesá. Naopak, pokud klesá (tedy R < 1, Rx-y>1), jedinců v budoucnosti je méně a relativní příspěvek konkrétního jedince v čase od x k y roste.")
9
NĚCO ZA NĚCO – TRADE-OFFS – vynucené kompromisy
Není těžké popsat ideální organismus – rozmnožuje se co nejdříve po narození, má hodně velkých potomků, o které pečuje, potomstvo plodí během svého dlouhého života co nejčastěji, zároveň je skvělým kompetitorem, uniká v pohodě predaci a sám chytí, na co si pomyslí. Popsat jej je snadné, představit si ho už jde špatně. Pokud nastrká co nejvíce zdrojů do rozmnožování, těžko je může investovat do přežívání, pokud se bude pořád zaobírat mláďaty, těžko si bude snadno hledat a lovit potravu. Je potřeba dělat kompromisy. Trade-off (směna) je vztahem mezi dvěma prvky životní strategie, při němž dochází ke zvyšování výhod plynoucích z jednoho na úkor snižování výhod plynoucích z druhého, protože dostupnost zdroje je limitovaná.
 je vztahem mezi dvěma prvky životní strategie, při němž dochází ke zvyšování výhod plynoucích z jednoho na úkor snižování výhod plynoucích z druhého, protože dostupnost zdroje je limitovaná.")
10
NĚCO ZA NĚCO – TRADE-OFFS

11
NĚCO ZA NĚCO – TRADE-OFFS
Ne všechny negativní korelace jsou nutně „směnou“. Pokud je málo zdroje a jedinec je zdatný kompetitor, tak bude dobrý ve všem, zatímco „losers“ budou pořád „losers“ . Náklady na rozmnožovaní (cost of reproduction, CR) Pokud jedinec investuje aktuálně do reprodukce, asi tím sníží rychlost svého růstu a délku přežívání a tím také potenciál pro rozmnožování v budoucnu. Je to vidět na předchozím obrázku (samci drosofil), ještě snadněji se to ukazu-je u rostlin. Když se kytka nenechá vykvést hned v prvním roce dospělosti (a pak by chcípla), ale zaštíp-ne se a nechá se do roku příštího, tak má daleko lepší květ (a víc semen). Absolutně i relativně vyšší alokace = chcípli Absolutně i relativně nižší alokace = přežili do dalšího roku
 Pokud jedinec investuje aktuálně do reprodukce, asi tím sníží rychlost svého růstu a délku přežívání a tím také potenciál pro rozmnožování v budoucnu. Je to vidět na předchozím obrázku (samci drosofil), ještě snadněji se to ukazu-je u rostlin. Když se kytka nenechá vykvést hned v prvním roce dospělosti (a pak by chcípla), ale zaštíp-ne se a nechá se do roku příštího, tak má daleko lepší květ (a víc semen). Absolutně i relativně vyšší alokace. = chcípli. Absolutně i relativně nižší alokace. = přežili do dalšího roku.")
12
obr 14.9 NĚCO ZA NĚCO – TRADE-OFFS Počet mláďat a jejich fitness
Zjednodušeně jde o trade-off mezi velikostí a počtem potomků. Dané množství zdroje může být rozděleno buď mezi hodně malých anebo méně velkých. Rodiče navíc mohou investovat „rodičovskou péči“ – pak se jedná o trade-off mezi počtem mláďat a množstvím potravy pro jejich krmení. obr 14.9 zlatobýl

13
OPTION SETS AND FITNESS COUNTOURS Podle čeho se stanovuje „kolik čeho kam“
Option set popisuje celou řadu možných kombinací mezi dvěma životními projevy, mezi kterými si může organismus vybírat – odráží fyziologické stavy organismu. Např. jedním projevem může být rozmnožování mx a druhým růst (který potencionálně ovlivňuje RRV). Hranice kombinací možností je křivka „trade-off“. Pokud je konvexní, implikuje, že jen o trošku menší než současná reprodukce umožňuje velké zvýšení růstu. Pokud je křivka konkávní, implikuje, že k podstatnému zvýšení růstu se musí reprodukce o hodně snížit. Fitness countours jsou přímky spojující body, kde kombinace (v našem příkladě) mx a růstu je konstantní. Přímky dále od počátku představují větší fitness, blíže k počátku menší fitness. Vyjadřují spíše vlastnosti prostředí (habitatu) než vlastnosti jedince.
. Hranice kombinací možností je křivka „trade-off . Pokud je konvexní, implikuje, že jen o trošku menší než současná reprodukce umožňuje velké zvýšení růstu. Pokud je křivka konkávní, implikuje, že k podstatnému zvýšení růstu se musí reprodukce o hodně snížit. Fitness countours jsou přímky spojující body, kde kombinace (v našem příkladě) mx a růstu je konstantní. Přímky dále od počátku představují větší fitness, blíže k počátku menší fitness. Vyjadřují spíše vlastnosti prostředí (habitatu) než vlastnosti jedince.")
14
OPTION SETS AND FITNESS COUNTOURS Podle čeho se stanovuje „kolik čeho kdy kam“
Text i k předchozímu obrázku

15
obr 14.11 Klasifikace stanovišť
Pokud fitness countours vyjadřují spíše vlastnosti stanoviště (habitatu) než vlastnosti jedince, znamená to, že odrážejí vliv stanoviště na organismus neboli reakci organismu na stanoviště. Pro usazené, etablované jedince (ne tedy pro nově narozené) lze na základě nákladů na reprodukci (CR) stanovit kontrastní typy stanovišť : 1. High CR (vysoké náklady), kde jakkoli snížený růst vinou aktuální reprodukce se podstatně negativně projeví na budoucí RRV, a tedy na fitness. Dosažení odpovídající fitness je proto možné kombinací vysoké reprodukce s pomalým růstem nebo nízkou reprodukcí s velkým růstem. 2. Low CR (nízké náklady na reprodukci), kdy je RRV jen málo ovlivněna aktuálním růstem. Fitness tak závisí jen na současné reprodukci – přímky jdou vertikálně. obr 14.11
 než vlastnosti jedince, znamená to, že odrážejí vliv stanoviště na organismus neboli reakci organismu na stanoviště. Pro usazené, etablované jedince (ne tedy pro nově narozené) lze na základě nákladů na reprodukci (CR) stanovit kontrastní typy stanovišť : 1. High CR (vysoké náklady), kde jakkoli snížený růst vinou aktuální reprodukce se podstatně negativně projeví na budoucí RRV, a tedy na fitness. Dosažení odpovídající fitness je proto možné kombinací vysoké reprodukce s pomalým růstem nebo nízkou reprodukcí s velkým růstem. 2. Low CR (nízké náklady na reprodukci), kdy je RRV jen málo ovlivněna aktuálním růstem. Fitness tak závisí jen na současné reprodukci – přímky jdou vertikálně. obr")
16
Klasifikace stanovišť
Tato klasifikace je komparativní – stanoviště se tak dají popisovat relativně vůči ostatním. Důvody pro zařazení stanoviště do příslušné skupiny mohou být různé: relativně High-CR tehdy, 1) je-li tam intenzivní kompetice a přežívají jen ti nejschopnější. Snížení aktuální reprodukce ve prospěch růstu vede k vysoké RRV (jeleni – jen ti nejlepší mají harém laní). 2) pokud jsou maličcí dospělci vystaveni silné predaci. Pak rychlý růst vede k úniku ze zranitelných velikostních tříd. Škeble únik krabům a kajkám. relativně Low-CR 1) mortalita je významná a neselektivní, takže jakákoliv investice do růstu je pravděpodobně na draka, když se stejně chcípne (vysychání periodických vod). 2) stanoviště je tak pohostinné, že všichni mají velkou šanci přežít a v budoucnosti se hodně množit, takže nějak se na počátku uskrovňovat nemá cenu (kolonisté). 3) na stanovišti působí mortalita zejména na velké jedince – čili růst znamená naopak mít menší RVV v budoucnu.
 je-li tam intenzivní kompetice a přežívají jen ti nejschopnější. Snížení aktuální reprodukce ve prospěch růstu vede k vysoké RRV (jeleni – jen ti nejlepší mají harém laní). 2) pokud jsou maličcí dospělci vystaveni silné predaci. Pak rychlý růst vede k úniku ze zranitelných velikostních tříd. Škeble únik krabům a kajkám. relativně Low-CR 1) mortalita je významná a neselektivní, takže jakákoliv investice do růstu je pravděpodobně na draka, když se stejně chcípne (vysychání periodických vod). 2) stanoviště je tak pohostinné, že všichni mají velkou šanci přežít a v budoucnosti se hodně množit, takže nějak se na počátku uskrovňovat nemá cenu (kolonisté). 3) na stanovišti působí mortalita zejména na velké jedince – čili růst znamená naopak mít menší RVV v budoucnu.")
17
Klasifikace stanovišť
Pro čerstvě narozené se dá také klasifikovat podle ↑velikost mláďat ↓ počet mláďat: Stanoviště citlivá na velikost mláďat tehdy, kdy RV jednotlivého mláděte s jeho velikostí podstatně roste (lepší v kompetici, nebo malí pod predací). Vzrůst znamená podstatný posun fitness contours. Stanoviště necitlivá na velikost mláďat tehdy, kdy se RV jednotlivého mláděte s jeho velikostí moc nemění (nespecifická mortalita, hodně zdrojů, predace orientována na velké). Vzrůst neovlivní podstatně posun fitness contours. Kombinace obou hledisek – 4 možné viz minulý obrázek
. Vzrůst znamená podstatný posun fitness contours. Stanoviště necitlivá na velikost mláďat tehdy, kdy se RV jednotlivého mláděte s jeho velikostí moc nemění (nespecifická mortalita, hodně zdrojů, predace orientována na velké). Vzrůst neovlivní podstatně posun fitness contours. Kombinace obou hledisek – 4 možné viz minulý obrázek.")
18
Klasifikace stanovišť
Kombinace obou hledisek – 4 možné

19
Low-CR High-CR High RA Low RA Reprodukční alokace
Pokud předpokládáme situaci s konvexním průběhem „options set“, tak by stanoviště s relativně nízkými náklady na reprodukci (low-CR) mělo povzbuzovat vyšší reprodukční alokaci, zatímco high-CR stanoviště naopak favorizovat nižší RA. Pokus se smetánkou lékařskou – pěstování různých biotypů (klonů A-D, A nejradši na low-CR, D nejradši na high-CR) na místech s různou CR, příklady ještěrek (běhající lovci nízká RA x číhající vyšší RA), živorodky ve sladké (nižší RA) a brakické vodě (vyšší RA), protože ve sladké vodě více energie na osmotickou rovnováhu.
 mělo povzbuzovat vyšší reprodukční alokaci, zatímco high-CR stanoviště naopak favorizovat nižší RA. Pokus se smetánkou lékařskou – pěstování různých biotypů (klonů A-D, A nejradši na low-CR, D nejradši na high-CR) na místech s různou CR, příklady ještěrek (běhající lovci nízká RA x číhající vyšší RA), živorodky ve sladké (nižší RA) a brakické vodě (vyšší RA), protože ve sladké vodě více energie na osmotickou rovnováhu.")
20
Kdy se začít množit – dříve anebo později?
Protože high-CR favorizují nízkou RA, dospělost jedince (bráno jako zahájení reprodukce) by na takových stanovištích měla být relativně opožděna – oddálena do pozdějšího věku, a dospělí jedinci by měli být větší (při oddálení reprodukce je nulová RA). Příklad Poa annua na dvou typech stanovišť – s nízkou hustotou a neselektivní mortalitou (= low-CR, oportunisté), a s vysokou hustotou a kompeticí (high CR, pastvina)
 by na takových stanovištích měla být relativně opožděna – oddálena do pozdějšího věku, a dospělí jedinci by měli být větší (při oddálení reprodukce je nulová RA). Příklad Poa annua na dvou typech stanovišť – s nízkou hustotou a neselektivní mortalitou (= low-CR, oportunisté), a s vysokou hustotou a kompeticí (high CR, pastvina)")
21
obr. 14.14 Kdy se začít množit – dříve anebo později?
Rozdíl mezi věkem dospělosti samců a samic ve vztahu k typu rozmnožování: U polygamních populací, kde se rozmnožování účastní jen omezený počet samců, tito by měli při opoždění dospělosti mít výhodu – budou později větší a snadněji se prosadí proti jiným. Naopak, při promiskuitním rozmnožování, a zejména tam, kde je vnější oplození, samci „nekontrolují“ přístup k samicím. Proto naopak samice (platí-li čím větší, tím více potomků) by měly mít při opoždění reprodukce relativně větší zisk vůči samcům. Pokud srovnáme na jedné straně savce a ptáky (mezi nimi převládají polygamní svazky) a na straně druhé ryby (běžně externí oplození), vychází to. obr
 by měly mít při opoždění reprodukce relativně větší zisk vůči samcům. Pokud srovnáme na jedné straně savce a ptáky (mezi nimi převládají polygamní svazky) a na straně druhé ryby (běžně externí oplození), vychází to. obr")
22
Kdy se začít množit – dříve anebo později?
Vztah mezi očekávanou délkou života samice a věkem, kdy dospívá: relativně konstantní uvnitř taxonomických skupin, mezi skupinami ale velké rozdíly. Možnosti životních strategií jsou dány evoluční historií jednotlivých skupin.

23
Semelparie - rozmnožování jen jedenkrát za život (x iteroparie)
Při srovnání low-CR a high-CR stano-višť je zřejmé, že semelparie bude typičtější spíše pro low-CR stanoviště. Synchronizace rozmnožování je u mnoha dlouhověkých druhů často asociována právě se semelparií (bambus, cikády). Evoluční příčinou je patrně „záchrana potomků před predací“, protože je-li potomstvo prokukováno masově synchronně v určitý rok, má daleko vyšší šanci přežít než kdyby se narodilo v roce, kdy se jich rodí málo. V „rozmnožovacím“ roce je tak fitness určena zejména aktuální úrovní reprodukce, a proto případná změna na iteroparii („teď se moc nerozmnožím, budu se šetřit ještě na příští rok“) nemá valného smyslu, protože by ta příští mláďata měla nízkou šanci přežít). Semelparie bude typická i pro organismy s konkávním option set, protože tam i malé reprodukční úsilí vede k významnému poklesu budoucího přežívání, ale změna z velkého na menší už tak dramatický efekt na přežití nemá. Pravděpodobně právě toto je důvodem, proč hodně druhů lososů vykazuje sebevražednou semelparii: nejvíce je stojí už samotný ‚akt‘ reprodukce (tah do řek na trdliště), a risk s ním spojený je víceméně nezávislý na velikosti RA.
. Evoluční příčinou je patrně „záchrana potomků před predací , protože je-li potomstvo prokukováno masově synchronně v určitý rok, má daleko vyšší šanci přežít než kdyby se narodilo v roce, kdy se jich rodí málo. V „rozmnožovacím roce je tak fitness určena zejména aktuální úrovní reprodukce, a proto případná změna na iteroparii („teď se moc nerozmnožím, budu se šetřit ještě na příští rok ) nemá valného smyslu, protože by ta příští mláďata měla nízkou šanci přežít). Semelparie bude typická i pro organismy s konkávním option set, protože tam i malé reprodukční úsilí vede k významnému poklesu budoucího přežívání, ale změna z velkého na menší už tak dramatický efekt na přežití nemá. Pravděpodobně právě toto je důvodem, proč hodně druhů lososů vykazuje sebevražednou semelparii: nejvíce je stojí už samotný ‚akt‘ reprodukce (tah do řek na trdliště), a risk s ním spojený je víceméně nezávislý na velikosti RA.")
24
Počet a velikost potomků
jak už víme z předchozího, rozdělení dané RA mezi méně velkých potomků je charakteristické pro stanoviště relativně citlivá na velikost mláďat. A naopak stanoviště relativně necitlivá na velikost mláďat, budou preferovat hodně malých. Kombinaci trade-off mezi počtem potomků a jejich velikostí řešil Lack (1947), a vycházelo, mu, že přírodní výběr by neměl favorizovat snůšky maximální, ale o trochu menší, optimalizující pravděpodobnost násled-ného přežití potomků. Tomuto se říká „Lackova velikost snůšky“ a mnoho dalších se to pokoušelo všemož-nými způsoby dokázat. Ve velké většině jim to ovšem nevycházelo. Až teď se ovšem ukazuje proč – obecně je totiž Lackův předpoklad správný, ale: Mnoho těchto studií mělo neadekvátní odhady fitness jednotlivých mláďat (nebrali do úvahy přežívání v dalším časovém úseku, a až vůbec ne reprodukční výsledky mláďat). Nynější pokusy s parazitoidem (Trichogramma) ukazují, že pokud deponoval do hostitele více vajec než normálně, tak i když měl i více potomků, jejich reprodukční schopnosti byly menší než u těch z normálních snůšek. Druhé selhání studií testujících „Lackovu velikost snůšky“ je právě v opomenutí kalkulace CR – nákladů na reprodukci. Přírodní výběr favorizuje takové strategie a životní historie, kde je maximální CELKOVÁ fitness. Proto velká a v daném okamžiku produktivní snůška může stát příliš mnoho z hlediska RRV. Ale stále pořád chybí detailní studie a data. … ale je to takhle: Lackova velikost snůšky
, a vycházelo, mu, že přírodní výběr by neměl favorizovat snůšky maximální, ale o trochu menší, optimalizující pravděpodobnost násled-ného přežití potomků. Tomuto se říká „Lackova velikost snůšky a mnoho dalších se to pokoušelo všemož-nými způsoby dokázat. Ve velké většině jim to ovšem nevycházelo. Až teď se ovšem ukazuje proč – obecně je totiž Lackův předpoklad správný, ale: Mnoho těchto studií mělo neadekvátní odhady fitness jednotlivých mláďat (nebrali do úvahy přežívání v dalším časovém úseku, a až vůbec ne reprodukční výsledky mláďat). Nynější pokusy s parazitoidem (Trichogramma) ukazují, že pokud deponoval do hostitele více vajec než normálně, tak i když měl i více potomků, jejich reprodukční schopnosti byly menší než u těch z normálních snůšek. Druhé selhání studií testujících „Lackovu velikost snůšky je právě v opomenutí kalkulace CR – nákladů na reprodukci. Přírodní výběr favorizuje takové strategie a životní historie, kde je maximální CELKOVÁ fitness. Proto velká a v daném okamžiku produktivní snůška může stát příliš mnoho z hlediska RRV. Ale stále pořád chybí detailní studie a data. … ale je to takhle: Lackova velikost snůšky.")
25
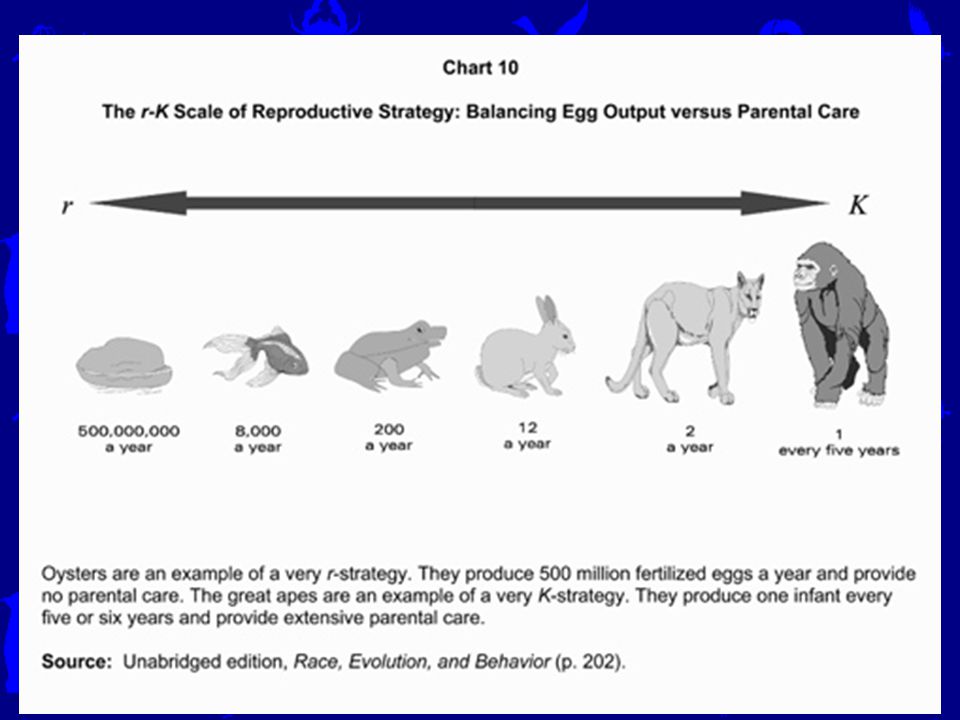
r a K selekce Základní koncepce životních strategií, pocházející od pánů McArthura a Wilsona (1967), rozvinutá Piankou (1970). Písmena v jejím názvu odrážejí parametry z logistické rovnice růstu populace: r strategové jsou organismy selektované pro výhody rychlého rozmnožování (vysoké r), zatímco K-stratégové jsou organismy selektované pro schopnost mít nejvyšší fitness (nejvíce reprodukčně přispívat do populace) při naplnění nosné kapacity K. Tato koncepce tak vychází z existence dvou kontrastních typů stanovišť: r-selektujících a K-selektujících. Je samozřejmě velké zjednodušení, ale pro ekologii velmi plodné. K selektující prostředí je prostředí, které jen velmi málo zažívá náhodné fluktuace. V důsledku toho populace dosáhne velmi rychle stále velikosti je silná kompetice mezi dospělými jedinci výsledek této kompetice určuje pravděpodobnost přežívání a fekunditu dospělců. Mladí také musí soutěžit, aby přežili, a mají jen málo příležitostí, aby dospěli v plodného jedince. Ve stručnosti – stanoviště je jak high-CR, tak i citlivé vůči velikosti mláďat. Pro K selektované jedince lze proto předpovědět, že budou větší, s oddálenou reprodukcí do staršího věku, iteroparičtí, s menším podílem RA a s většími (a proto o menším počtu) mláďaty.
, zatímco K-stratégové jsou organismy selektované pro schopnost mít nejvyšší fitness (nejvíce reprodukčně přispívat do populace) při naplnění nosné kapacity K. Tato koncepce tak vychází z existence dvou kontrastních typů stanovišť: r-selektujících a K-selektujících. Je samozřejmě velké zjednodušení, ale pro ekologii velmi plodné. K selektující prostředí je prostředí, které jen velmi málo zažívá náhodné fluktuace. V důsledku toho populace dosáhne velmi rychle stále velikosti je silná kompetice mezi dospělými jedinci výsledek této kompetice určuje pravděpodobnost přežívání a fekunditu dospělců. Mladí také musí soutěžit, aby přežili, a mají jen málo příležitostí, aby dospěli v plodného jedince. Ve stručnosti – stanoviště je jak high-CR, tak i citlivé vůči velikosti mláďat. Pro K selektované jedince lze proto předpovědět, že budou větší, s oddálenou reprodukcí do staršího věku, iteroparičtí, s menším podílem RA a s většími (a proto o menším počtu) mláďaty.")
26
r a K selekce (+ S) Naopak, r selektující prostředí je prostředí, jehož budoucnost je nepředpověditelná, nebo vůbec krátká. Populace zde žijící zažívá období rychlého růstu, bez kompetice (jsou zrovna báječné podmínky s hojností pro všechny, nebo se vůbec jedná o první kolonizaci). Ale pak následují období s nevyhnutelnou mortalitou (vše je sežráno, nebo se habitat prudce změní/zanikne). Rychlost vymírání jak dospělých, tak i mláďat je potom velmi variabilní a nepředpověditelná – a velmi často nezávislá na hustotě populace a na velikosti a stavu jedinců samotných. Ve stručnosti – stanoviště je jak low-CR, tak i NEcitlivé vůči velikosti mláďat. Pro r selektované jedince lze proto předpovědět, že budou menší velikosti, budou rychle dospívat, budou asi semelparní, budou mít velkou RA a menší (a proto početné) potomstvo. Jednotlivci investují jen málo do přežívání, ale toto je závislé hlavně na vnějších (nepředpověditelných) faktorech. ‚r/K koncepce‘ byla původně odvozená od průběhu kolonizace ostrovů, ale má daleko širší dosah. Jedná se vlastně o případ odpovídající nám známé klasifikaci stanovišť. Proto je třeba si připomenout, že jsou i jiné kombinace, a také že r/K schéma může resultovat z mnoha jiných příčin (predace malých dospělců x intenzivní kompetice mezi dospělci; mortalita velkých mláďat x fluktuace mezi dobou hojnosti a dobou neselektující mortality atd.) R-C-S strategie u rostlin (Grime 1974,1979): U rostlin (respektive asi sesilních organismů) často třetí možnost – Stres tolerance (S), na stanovištích trvale extrémních, kde je investice do velikosti (být „ K “) na nic (či spíše k horšímu), ale nemá cenu ani mohutně se občas množit (být „ r “).
. Ale pak následují období s nevyhnutelnou mortalitou (vše je sežráno, nebo se habitat prudce změní/zanikne). Rychlost vymírání jak dospělých, tak i mláďat je potom velmi variabilní a nepředpověditelná – a velmi často nezávislá na hustotě populace a na velikosti a stavu jedinců samotných. Ve stručnosti – stanoviště je jak low-CR, tak i NEcitlivé vůči velikosti mláďat. Pro r selektované jedince lze proto předpovědět, že budou menší velikosti, budou rychle dospívat, budou asi semelparní, budou mít velkou RA a menší (a proto početné) potomstvo. Jednotlivci investují jen málo do přežívání, ale toto je závislé hlavně na vnějších (nepředpověditelných) faktorech. ‚r/K koncepce‘ byla původně odvozená od průběhu kolonizace ostrovů, ale má daleko širší dosah. Jedná se vlastně o případ odpovídající nám známé klasifikaci stanovišť. Proto je třeba si připomenout, že jsou i jiné kombinace, a také že r/K schéma může resultovat z mnoha jiných příčin (predace malých dospělců x intenzivní kompetice mezi dospělci; mortalita velkých mláďat x fluktuace mezi dobou hojnosti a dobou neselektující mortality atd.) R-C-S strategie u rostlin (Grime 1974,1979): U rostlin (respektive asi sesilních organismů) často třetí možnost – Stres tolerance (S), na stanovištích trvale extrémních, kde je investice do velikosti (být „ K ) na nic (či spíše k horšímu), ale nemá cenu ani mohutně se občas množit (být „ r ).")
28
r a K selekce – důkazy Stromy v lese – K stratégové (malá RA, iteroparie, velcí jedinci, velká semena, dlouho žijí, pozdě se rozmnožují)
")
29
r a K selekce – důkazy Dva druhy orobince v Americe

30
R+C+S R-C-S strategie u rostlin (Grime 1974,1979):
U rostlin (respektive asi sesilních organismů) často třetí možnost – Stres tolerance (S), na stanovištích trvale extrémních, kde je investice do velikosti (být „ K “) na nic (či spíše k horšímu), ale nemá cenu ani mohutně se občas množit (být „ r “). S-stratégové: Různá životní formy nízká růstová rychlost, pozdní reprodukce nízké reprodukční úsilí nepravidelná reprodukce malá schopnost konkurence Dlouhověkost malá produkce biomasy dlouhověký asimilační aparát
 často třetí možnost – Stres tolerance (S), na stanovištích trvale extrémních, kde je investice do velikosti (být „ K ) na nic (či spíše k horšímu), ale nemá cenu ani mohutně se občas množit (být „ r ). S-stratégové: Různá životní formy. nízká růstová rychlost, pozdní reprodukce. nízké reprodukční úsilí. nepravidelná reprodukce. malá schopnost konkurence. Dlouhověkost. malá produkce biomasy. dlouhověký asimilační aparát.")
31
R+C+S R-C-S strategie u rostlin (Grime 1974,1979):
U rostlin (respektive asi sesilních organismů) často třetí možnost – Stres tolerance (S), na stanovištích trvale extrémních, kde je investice do velikosti (být „ K “) na nic (či spíše k horšímu), ale nemá cenu ani mohutně se občas množit (být „ r “).
 často třetí možnost – Stres tolerance (S), na stanovištích trvale extrémních, kde je investice do velikosti (být „ K ) na nic (či spíše k horšímu), ale nemá cenu ani mohutně se občas množit (být „ r ).")
32
R+C+S R-C-S strategie u rostlin (Grime 1974,1979):
U rostlin (respektive asi sesilních organismů) často třetí možnost – Stres tolerance (S), na stanovištích trvale extrémních, kde je investice do velikosti (být „ K “) na nic (či spíše k horšímu), ale nemá cenu ani mohutně se občas množit (být „ r “).
 často třetí možnost – Stres tolerance (S), na stanovištích trvale extrémních, kde je investice do velikosti (být „ K ) na nic (či spíše k horšímu), ale nemá cenu ani mohutně se občas množit (být „ r ).")
33
R+C+S R-C-S strategie u rostlin (Grime 1974,1979): S-stratégové:
Různá životní formy nízká růstová rychlost, pozdní reprodukce nízké reprodukční úsilí nepravidelná reprodukce malá schopnost konkurence Dlouhověkost malá produkce biomasy dlouhověký asimilační aparát

Podobné prezentace



















 6. předn.1 chování výběrového průměru nechť X 1, X 2,…,X n jsou nezávislé náhodné veličiny s libovolným rozdělením.>")
