Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Vývoj NS je formován procesy determinace, diferenciace a indukce
Determinace - dána procesem genové exprese Míra determinace různá u různých živočichů Vývoj determinovaný (všichni bezobratlí) Nederminovaný (chordata vč. obratlovců)
 Nederminovaný (chordata vč. obratlovců)")
2
Determinovaný vývoj Už ve fázi zygoty jsou jednotlivé bb embrya předurčeny ve svém vývoji Osud buněk je určován genetickými faktory Malá míra flexibility

3
Vývoj nedeterminovaný
Předurčenost buňky k určitému vývoji (daná genomem) je významná, ale Vývoj je ovlivňován vnějšími faktory (chemický gradient, chemické látky uvolňované sousedními bb, specifickými naváděcími bb, růstovými faktory, elektrickým gradientem embrya atd.)
 je významná, ale. Vývoj je ovlivňován vnějšími faktory. (chemický gradient, chemické látky uvolňované sousedními bb, specifickými naváděcími bb, růstovými faktory, elektrickým gradientem embrya atd.)")
4
Osud buňky je výsledkem indukce a ne automatickým spuštěním genetického programu, odstranění jedné nebo více bb nemá fatální vliv na osud embrya

5
Neurulace NS se vyvíjí z ektodermu Indukcí mezodermem
Začíná vytvořením Spermanova organizátoru – amphibia Hensenovy zóny – ptáci, savci Produkce růstových faktorů (difusibilní proteiny) indukují neurální diferenciaci a tvorbu nervové rýhy a nervové ploténky, neurální trubice
 indukují neurální diferenciaci a tvorbu. nervové rýhy a nervové ploténky, neurální trubice.")
6
Buňky z okrajů neurální rýhy se oddělují a tvoří neurální lištu
Buňky z rýhy migrují na periferii – periferní orgány dorzální ganglia a ganglia autonomních nervů Ventrálně vzniká chorda Anteriorně – diferenciace – základy mozku

7
Neurulace schema a foto

8
Produkce neuronů a glií
Zpočátku je N trubice jednovrstevná, později vícevrstevná Dělení probíhá v germinální zóně (vnitřní strana) Během bb dělení jádra složitě migrují DNA syntéza probíhá na vnějším okraji, Dělení na vnitřním Během migrace jsou jádra ovlivňována různými cytoplasmatickými faktory Po několika cyklech dceřinné buňky nejsou schopny dalšího dělení a opouštějí germinální zónu
 Během bb dělení jádra složitě migrují. DNA syntéza probíhá na vnějším okraji, Dělení na vnitřním. Během migrace jsou jádra ovlivňována různými cytoplasmatickými faktory. Po několika cyklech dceřinné buňky nejsou schopny dalšího dělení a opouštějí germinální zónu.")
9
Migrace Dceřinné bb migrují Neurony se dále nedělí – bb postmitotické
Glie se mohou dělit N trubice se stává 3 vrstevnou Ventrikulární zóna – proliferační Šedá hmota – vrstva neuronální prekursorů a migrujících neuronů Bílá hmota – vrstva neuronálních výběžků

10
Produkce neuronálních a gliových prekurzorů
Neuroektoderm S Centrální kanál M Migrující prekurzory amitotické neurony a glie Vnější strana ventrikulární střední okrajová zóna

11
Migrace neuronů v CNS Mícha Migrující neurony Centrální kanál Pleny
Střední Šedá hmota Marginální Bílá hmota Ventrikulární

12
Migrace v mozkové části
3 vrstevná stra přetrvává v oblasti míchy a prodloužené míchy V mozkové a mozečkové části neurony migrují periferně a tvoří kortikální ploténky Neurony migrují podél radiálních gliových buněk

13
Migrace neuronů v CNS Mozeček Radiální gliové buňky Migrující neurony
Komora Granul. bb Purkyň. bb VZ MZ Budoucí bílá hmota Korová ploténka

14
Migrace neuronů v CNS Mozek Migrující neurony Komora VZ Střední vrstva
MZ Korová ploténka Bílá hmota

15
Radiální gliové buňky Udržují kontakt s vnitřní a vnější stranou NT po celou dobu vývoje Na migraci neuronů se podílí neznámé transportní bílkoviny Po ukončení migrace neuronů zanikají

18
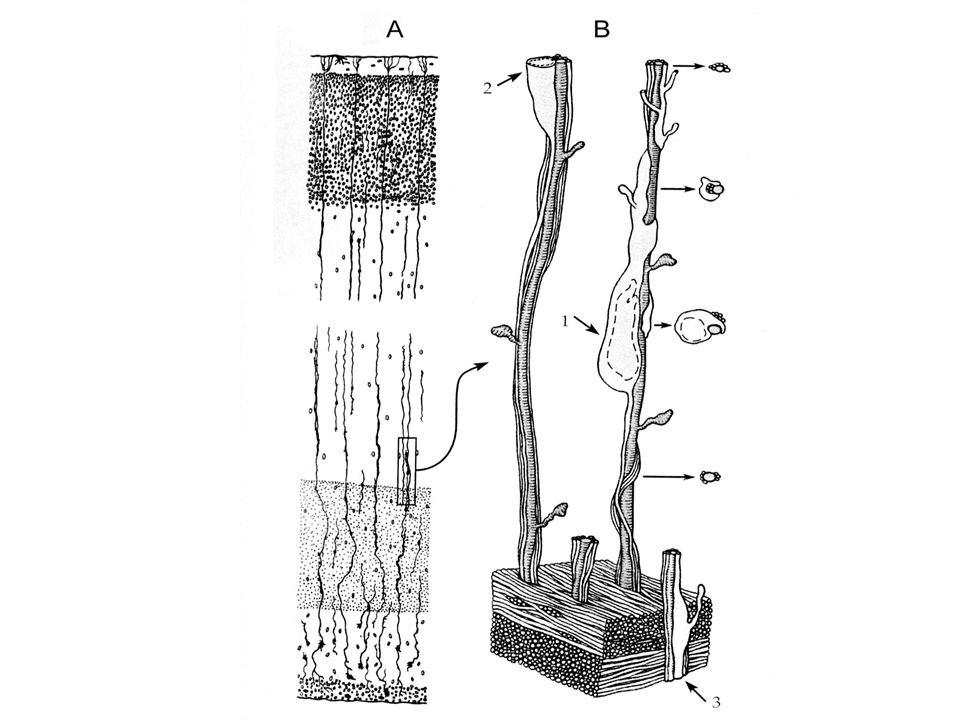
Migrace buněk neurální lišty
Bez pomoci glivých buněk Migrace řízena atraktivními a repulsivními interakcemi s buněčnými povrchy a komponentami extracelulárního prostoru (adhesivní faktory) V průběhu migrace se neurony dostávají do různých oblastí - indukce
 V průběhu migrace se neurony dostávají do různých oblastí - indukce.")
19
Migrace buněk neurální lišty
DR, autonomní ganglia, Schwann.bb, adrenergní chromafinní bb. Posteriorně Epidermis Neurální trubice Dermatomy Sklerotomy Chorda Anteriorně Anteriorní sklerotom Laminin, fibronektin Posteriorní Sklerotom F-spondin

20
Neurální lišta Naurální trubice Ektoderm Smyslová ganglia Somit Chorda
Sympatická ganglia Aorta Nadledvinky Tělní dutina Střevo

21
Neurony se po migraci dále nedělí
Výjimky tvoří – Interneurony bazálních ganglií Granulární bb mozkové kůry Primární aferentní neurony – čichové Postgangliové neurony autonomního Ns Bb nadledvinek, které tvoří A a NA

22
Mechanismus indukce – vyjádření genotypu
U dospělého jedince jsou různé oblasti NS fenotypově odlišné Fenotyp je determinován selektivní expresí genů Během vývoje dochází různými mechanismy indukce k Hierarchicky regulované expresi transkripčních faktorů

23
Forebrain Midbrain Hindbrain Enterická parasympatická ganglie Sympatická Ganglia a adrenergní dřeň

24
Některé geny kontrolují rozhodnutí o tvorbě tělních částí - rhombomer
V každém segmentu se neuronální bb diferencují ve stejném rytmu Proces diferenciace je specificky modifikován Hox geny – determinují osud neuronů v každém segmentu Vybírají k expresi ty geny, které jsou třeba v daném segmentu

25
Geny aktivované během ranného vývoje NS
Hox geny - osud segmentů Transkripční faktory (Kreisler, Krox-20) Receptor tyrosin kináza (Sek-1, Sek-4,transmembránový protein, ligand = Elf-2)
 Receptor tyrosin kináza (Sek-1, Sek-4,transmembránový protein, ligand = Elf-2)")
26
Hox – homeotické geny Nalezeny u Drosophily (mutace - tykadlo místo nohy) Geny zodpovědné za vznik určité struktury Kontrolují podřízené geny a transkripční faktory Exprese hox genů řízena kyselinou retinovou ze Spermanova organizátoru Systematicky organizovaná citlivost receptorů

27
Anterio-posteriorní gradient
Kyselina retinová

28
Hox geny – homeotické regulační geny – kyselina retinová
(reaguje s receptory v jádrech cílových buněk) Hierarchicky regulovaná exprese transkripčních faktorů
 Hierarchicky regulovaná exprese transkripčních faktorů.")
29
NS obratlovců také diferencován ve směru dorso-ventrálním
Indukční signály produkované chordou Produkt „sonic headgehog“ genu difusibilní protein kontroluje hox geny Později proužkem specializovaných gliových buněk bazální ploténky na ventrální straně míchy

30
Dorzo-ventrální gradient
Bazální ploténka Anterior Dorzálně Posterior Chorda Ventrálně Difuzibilní protein Produkovaný Headgehog genem

31
Anterioposteriorní a dorsomediální gradienty kontrolují transkripční faktoy a determinují regionální identitu CNS

32
Buněčné linie – vztah mezi narozením neuronů a jejich osudem
U obratlovců je obtížné sledovat buněčné linie Techniky vnesení markeru (fluorescenčně označený virus) do prekurzorové bb a sledování výskytu v hotovém NS Vnesení markeru do retiny nově narozené krysy – označené byly neurony i glie Do kůry – klony s virem byly jenom buď v gliích nebo neuronech – diferenciace probíhá dříve
 do prekurzorové bb a sledování výskytu v hotovém NS. Vnesení markeru. do retiny nově narozené krysy – označené byly neurony i glie. Do kůry – klony s virem byly jenom buď v gliích nebo neuronech – diferenciace probíhá dříve.")
33
Buněčné linie v savčím CNS
Retina Epitel Čočka 4-6 týdnů Injekce retroviru kódující b-galaktosidázu do prekurzorové buňky

34
Existuje závislost mezi vznikem neuronů a jejich uspořádáním v kůře
Injekce radioaktivního thymidinu do dělících se buněk kůry Nejhlouběji jsou ty, co se dřív narodily

35
Počet buněk v jednotlivých vrstvách kůry po injekci 3H tymidinu
1 2+3 4 5 6 Bílá hmota E30 E33 E39 E42 E48 E56

36
Zapojení neuronů v jednotlivých vrstvách kůry je ovlivňováno lokálními faktory
Transplantace neuronů vyvíjející se zrakové kůry do oblasti somatosensorické vedla k indukci neuronálních fenotypů typických pro somatosensorickou kůru i s propojením neuronů

37
Laminární determinace v kůře
1 Laminární determinace v kůře 2 3 E29 4 Korová ploténka 5 Vnitřní zóna 6 Ventrikulární zóna Bílá hmota VZ

38
Laminární determinace v kůře
1 2 P1 3 4 Korová ploténka 5 Vnitřní zóna 6 Ventrikulární zóna Bílá hmota VZ

39
V dospělém NS může být vývoj pod humorální kontrolou
Pěvci – kanáři Samci „high vocal center nucleus“ Zde probíhá turnover neuronů v závislosti na hladině testosteronu Zvýšení hladiny 2x ročně, jaro, podzim Po poklesu kontrolovaný zánik neuronů Samice toto centrum nemají, ale po injekci testosteronu je indukováno

40
Hormonální regulace vývoje NS
Nadřazené vokální centrum „Robust“ jádro v archistiatu Nové neurony v dospělém mozku asimilace do složité architektury substrát pro remodelování chování Hlasivky

41
Kmenové buňky High vocal centrum ptáků Hipokampus Čichový lalok savců
z kmenových buněk vnitřní vrstvy laterálních komor – rychle se dělí a dceřinné bb migrují do čichového epitelu a zabudovávají se do stávajících neuronálních okruhů Možnost diferenciace do Neuronů Oligodendrocytů Astrocytů

42
Kontrola fenotypu v periferním NS
Kuřecí a křepelčí zárodky Pozice v neurální liště předurčuje typ periferního NS a typ vznikajícího zapojení I na expresi genů periferního NS mají vliv regionální faktory

43
Fenotyp v periferním NS
Křepelčí embryo Kuřecí embryo Fenotyp v periferním NS

44
Křepelčí embryo Kuřecí embryo

45
Výběr mediátoru cílovou tkání
Některé buňky během vývoje syntetizují jiný mediátor než v dospělém zapojení Změna metabolismu je indukována látkami cílové tkáně Potní žlázy, sympatické neurony, norepinephrine – Ach

46
Hladká svalovina žlaz indukuje
Cholinergní diferenciaci sympatických neuronů LIF (leukemia inhibitory faktor) Ciliary neurotrophic factor Cardiotropin – 1 Aktivace Ach syntézy v sympatických neuronech
 Ciliary neurotrophic factor. Cardiotropin – 1. Aktivace Ach syntézy v sympatických neuronech.")
47
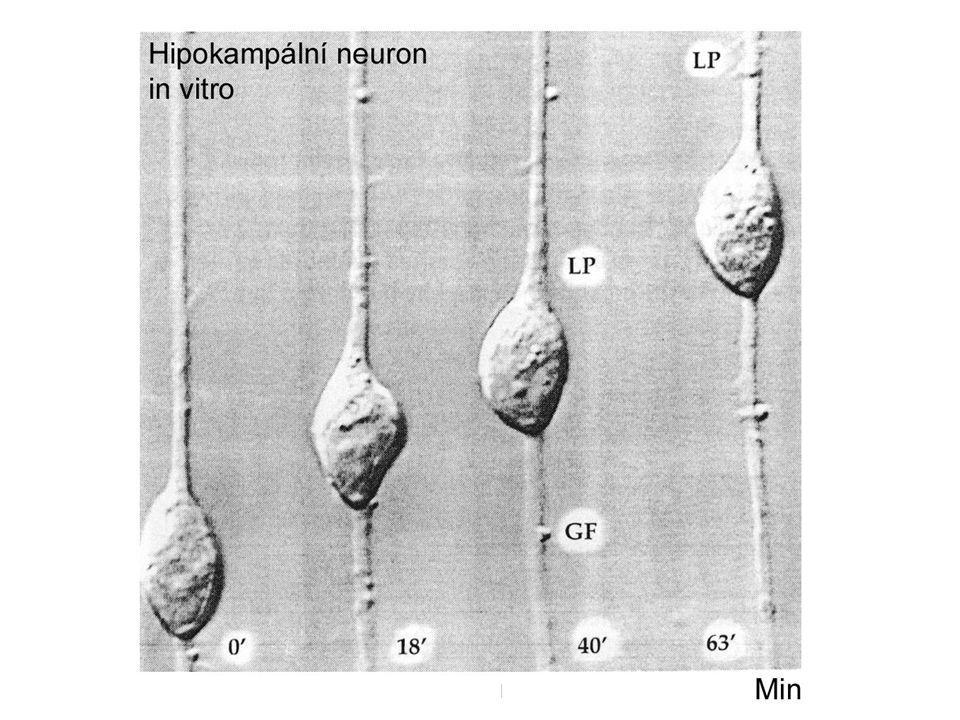
Růst neuronálních výběžků
Na růstovém vrcholu Amoeboidní pohyb – lamelipodia a filopodia Přichycují se k substrátu a orientují růstový vrchol Obsahují vlákna aktinu Filopodia „chutnají“ prostředí a odpovídají na chemické stimuly Short range stimuly na stěnách sousedních bb Long-range stimuly – v ect – přitahují či odpuzují

48
Růst axonů

49
Myosin zasouvá mikrotubuly Filopodia v klidu Aktinové monomery Myosin
Mikrotubulus Aktinová filamenta Aktinová polymerizace Vysouvání filopodií Myosin posouvá mikrotubuly Mikrotubulus Myosin Aktinová filamenta ATP, Actin-binding-protein Ca, proteinkináza Aktinová vlákna přichycená k podkladu

50
Naváděcí signály – extracelulární adhesní proteiny
Short range Na membrány vázané adhesní molekuly Imunoglobulinové molekuly Např. populární glykoprotein axonálního povrchu TAG1 (Transiently expressed Axonal surface Glykoprotein) Pomáhá řízenému prorůstání neuronů v míše a způsobuje u jistých axonů změnu směru růstu Některé působí prostřednictvím specifických receptorů, jiné spojují podobné struktury na sousedních bb
 Pomáhá řízenému prorůstání neuronů v míše a způsobuje u jistých axonů změnu směru růstu. Některé působí prostřednictvím specifických receptorů, jiné spojují podobné struktury na sousedních bb.")
51
Adheze často závisí na Ca++
Např. N cadherin – spojení buněk v ect tekutině, plasmě, krvi V tkáňových kulturách stimulují adhezi neuronů k podkladu, navzájem a stimulují růst výběžků A adhezi výběžků do primitivních nervů

52
Adhezní extracelulární glykoproteiny
Laminin, fibronektin, tenascin, trombospondin Spolupracují s imunoglobuliny Specifické kombinace zajistí, že ve vyvíjejícím se embryu určitá skupina vysílá výběžky do patřičné oblasti Adhezivní interakce působí na malou vzdálenost – 1 mm

53
Adhezivní molekuly Buněčné membrány
Transmembránové proteiny nebo asociované glykoproteiny, extracelulární část podobná imunoglobulinům N-CAM (neural cell adhesion molecule) CAM (Ng-CAM, glial cell adhesion molecule) TAG-1, MAG, DCC Homofilické vazby Heterofilické N-CADHERIN (Ca-dependentní buněčná adheze)
 CAM (Ng-CAM, glial cell adhesion molecule) TAG-1, MAG, DCC. Homofilické vazby. Heterofilické. N-CADHERIN (Ca-dependentní buněčná adheze)")
54
Adhezivní molekuly Extracelulární matrix LAMININ, FIBRONECTIN TENASCIN
CYTOTAXIN

55
Hmyzí NS má své dispečery
Fascikliny – vyvíjející se nervové pásce určují směr a dávají příkazy „shlukujte se“ „nerozcházejte se! Pionýrské neurony na větší vzdálenosti Některé smyslové neurony diferencují dřív hledají dráhu do příslušných oblastí a slouží jako orientační základna pro později narozené druhy

56
Navádění axonů na větší vzdálenosti
Naváděcí buňky Označují přechodné cíle Většinou nezralé neurony Přechodné synaptické kontakty Po splnění úkolu zanikají

57
Růst axonů

58
Chemoorientace Axon se může orientovat v závislosti na cílové tkáni i nezávisle Axony spinálních motoneuronů se orientují na periferii prostřednictvím interakce různých faktorů Adhezní proteiny – krátké vzdálenosti Delší vzdálenosti Netrin – chemický gradient (atrakce) Semaforin – chemický gradient (repulse) Příklad složitosti procesů navigace – navádění komisurálních interneuronů v dorsální části míchy
 Semaforin – chemický gradient (repulse) Příklad složitosti procesů navigace – navádění komisurálních interneuronů v dorsální části míchy.")
59
Chemo-atraktivní interace
Long-range Close-range Comisurální neuron Receptor DCC Exprimov V axonech TAG-1 adhezivní molekula Netrin-1 Bazální ploténka NrCAM Adhezivní molekula Jinde a v jiné souvislosti DCC funguje jako buněčná adhezivní molekula

60
Chemo-repulzní interakce
Short-range Long-range Motoneuron Periferie Receptor Robo Slit Netrin-1 Slit Difuzibilní protein

61
Rozpoznávání cíle Aby NS fungoval jak má a produkoval příslušné chování, musí být spoje specifické Tvorba spojů je geneticky podmíněna a závisí na environmentálních faktorech Neurony se chovají jakoby věděly kam růst Jak poznají cíl?

62
Zrakový systém obojživelníků a ptáků
Gangliové bb retiny prorůstají do tekta Z temporální oblasti retiny do anteriorní Tkáňové kultury Chemické naváděcí signály – EPHRINY EPH Receptory v tektu V retině - naso-temporální gradient receptorů V tektu - antero-posteriorní gradient ephrinů

63
Specifická inervace Levé tektum Interakce
receptor tyrosin kináz (Eph kinázy) a ephrinů - mediují vždy repelenci Levé tektum Ephriny exprimovány na cílových neuronech tekta Eph kinázy exprimovány na axonech ganliových buněk Pravé oko
 a ephrinů. - mediují vždy repelenci. Levé tektum. Ephriny exprimovány. na cílových neuronech tekta. Eph kinázy exprimovány. na axonech ganliových. buněk. Pravé oko.")
64
Retina Tektum Anteriorní (přední) Temporální Posteriorní (zadní) Nazální Gradient Eph receptorů Gradient Ephrinu

65
Axony gangliových buněk temporární sítnice
Posteriorní tektum Posteriorní tektum Eph-receptory Ephrin

66
Naváděcí signály a receptory
Short-range (kontaktní) TAG-1 Slit N-CAM ECM adhezní proteiny Ephriny Long-range (difusibilní) Netrin Semaphorin NrCAM Robo N-CAM Integriny Eph receptory DCC Neuropiliny A R Ca, proteinkinázy modulují
 TAG-1. Slit. N-CAM. ECM adhezní proteiny. Ephriny. Long-range (difusibilní) Netrin. Semaphorin. NrCAM. Robo. N-CAM. Integriny. Eph receptory. DCC. Neuropiliny. A. R. Ca, proteinkinázy modulují.")
67
Synaptogeneze na nervosvalové ploténce
Embryonální svalové vlákno má embryonální Ach receptory rovnoměrně na povrchu Při prorůstání axonu se uvolňuje spontánně Ach – depolarizační potenciál Po kontaktu se tvoří základ synapse Zvyšuje se uvolňování Ach, synapse se dotvoří během minut a receptory se zahušťují pod synapsí Objevuje se acetylcholinesteráza

68
Formování synapse nervosvalové spojení
Embryonické Ach abjd mRNA Dospělé Ach abde mRNA Agrin motoneuronů Heparan sulfát Proteoglykan Receptory MUSK Retrográdní signály - laminin

69
Formování synapsí v CNS
V CNS na jednom neuronu konverguje mnoho synapsí s různými mediátory I v CNS dochází ke shlukování příslušných receptorů pod příslušnými synapsemi Synaptogeneze a na periferii je shodná

70
Formování synapsí v CNS

71
Principy synaptogeneze
Neexistují předem daná místa pro tvorbu synapse Po kontaktu axon indukuje změny v postsynaptickém elementu Trofické změny působí anterográdně i retrográdně Významně působí elektrická aktivita Inervace je źpočátku polyneurální, Počet synapsí je následně upravován kompeticí neuronů o trofické faktory cílové tkáně

72
Kompetitivní interakce během vývoje

73
Kompetitivní interakce během vývoje
Po vzniku polyneurální inervace Některé spoje zanikají Dochází k reorganizaci Nadprodukce spojů a následná řízená smrt je charakteristická pro vývíjející se NS

74
velikosti inervované cílové tkáně a syntézu proteolytických enzymů
Mechanismus, který zajišťuje, aby neuronální vstup odpovídal velikosti inervované cílové tkáně zahrnuje aktivaci genu pro řízený rozpad DNA a syntézu proteolytických enzymů

75
Neuronální aktivita a eliminace synapsí
V procesu eliminace spojů má významnou úlohu neuronální aktivita Stimulace vede k eliminaci neaktivních vláken Stimulované spoje tvoří více větvení než nestimulované

76
Přirozená smrt neuronů během ontogeneze
Programovaná smrt Selekce programu vlivem mimobuněčných faktorů Součást přirozeného vývoje

Podobné prezentace