Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Mgr. Jiřina Procházková, Ph.D. jipro@sci.muni.cz
Změny aktivity transkripčního faktoru NF-kappaB ve vztahu k regulaci cytokinetiky Mgr. Jiřina Procházková, Ph.D.

2
Osnova Struktura NF-kappaB Charakterizace členů rodiny NF-kappaB
Klasická vs. alternativní dráha aktivace Transaktivace, ubiquitinace NF-kappaB a apoptóza NF-kappaB a imunitní systém NF-kappaB a proliferace NF-kappaB a stres NF-kappaB a karcinogeneze Metody detekce aktivního NF-kappaB

3
NF-kappaB Nuclear Factor – kappa B (NF-kB)
- Jaderný faktor B lymfocytů, který se váže na enhancer pro gen kódující lehký k řetězec imunoglobulinů Objeven v roce 1986 (Sen, Baltimore et al.) Pro-přežití, pro-zánětlivý transkripční faktor Aktivován min. 210 podněty a ovlivňuje transkripci min. 150 genů (Pahl 1999) Úzká vazba na procesy proliferace, karcinogeneze, aktivace imunitního systému
 Pro-přežití, pro-zánětlivý transkripční faktor. Aktivován min. 210 podněty a ovlivňuje transkripci min. 150 genů (Pahl 1999) Úzká vazba na procesy proliferace, karcinogeneze, aktivace imunitního systému.")
4
Struktura NF-kappaB NLS
dimerizace, jaderná lokalizace, vazba na DNA maskují NLS, interakce protein-protein obsahuje rozpoznávací sekvenci pro kinázy NLS TD PEST chrání protein před degradací v proteasomu, nutná pro správné štěpení proteinu neubiquitinylovaná chrání před proteasom. degradací PEST rich in prolineP, glutamic acidE, serineS, and threonineT residues ankyrin repeat is a 33-residue motif in proteins consisting of two alpha helices separated by loops, first discovered in signaling proteins in yeast Cdc10 and Drosophila Notch. Ankyrin repeats mediate protein-protein interactions and are among the most common structural motifs in known proteins. They appear in bacterial, archaeal, and eukaryotic proteins, but are far more common in eukaryotes. Most proteins that contain the motif have four to six repeats, although its namesake ankyrin contains 24 and the largest known number of repeats is 34, predicted in a protein expressed by Giardia lamblia.[1] ankyrins are a family of proteins that mediates the attachment of integral membrane proteins to the cytoskeleton. TD Transactivation Domain NLS Nuclear Localization Signal PEST rich in P, E, S, T aminoacids Podle Dejardin (2006), Biochemical Pharmacology 72:1161–1179
, Biochemical Pharmacology 72:1161–1179.")
5
Rodina NF-kappaB/IkappaB
PEST /p65 Transaktivační doména Members of the Rel/NF- B and I B families of proteins. The arrows indicate the endoproteolytic cleavage sites of p105 and p100 which give rise to p50 and p52, respectively. Black boxes indicate the PEST domains (obsahuje rozpoznávací sekvenci pro kinázy (prolin, glutamová kys., serineS, and threonineT residue), shaded boxes on Bcl-3 indicate transactivation domains, and gray boxes on RelB indicate leucine zipper domains. Abbreviations: RHD, Rel homology domain; ANK, ankyrin repeat; SS, signal-induced phosphorylation sites. Podle Caamano and Hunter (2002), Clinical Microbiology Reviews, 15 (3):414
, shaded boxes on Bcl-3 indicate transactivation domains, and gray boxes on RelB indicate leucine zipper domains. Abbreviations: RHD, Rel homology domain; ANK, ankyrin repeat; SS, signal-induced phosphorylation sites. Podle Caamano and Hunter (2002), Clinical Microbiology Reviews, 15 (3):414.")
6
Dimerizace členů rodiny NF-kappaB
Figure 1. Schematic of the canonical nuclear factor (NF)-kB pathway. The key factors involved in the regulation of the canonical NF-kB pathway include members of the IkB (inhibitor of kB) family and the IkB kinase (IKK) complex that consists of IKKa–IKKb heterodimers and the scaffold protein IKKg/NEMO. A variety of stimuli including the cytokines tumor necrosis factor (TNF)-a and interleukin (IL)-1 activate the IKK complex leading to the phosphorylation of the IkB proteins. This phosphorylation leads to IkB polyubiquitination and its subsequent degradation by the 26S proteasome. The NF-kB proteins are, thus, liberated from IkB and translocate to the nucleus where they bind to the promoter regions of NF-kB-responsive genes to result in increased gene expression. Figure 3. Nucleosomal role of IkB (inhibitor of kB) kinase-a (IKK-a). IKKa plays important part in regulating nuclear factor (NF)-kB transcriptional activation in the nucleus. In response to cytokines such as tumor necrosis factor (TNF)-a, IKKa is recruited to the promoter regions of NF-kB-responsive genes and forms a complex on the promoter with p65 and the coactivator cAMP-response-element-binding protein (CREB)-binding protein (CBP). The formation of this complex facilitates the phosphorylation (P) of histone H3 on Ser10 and leads to the subsequent acetylation (AC) by CBP to induce the expression of NF-kB-responsive genes. Yamamoto and Gaynor, (2004), TRENDS in Biochemical Sciences, 29 (2)
-kB pathway. The key factors involved in the regulation of the canonical NF-kB pathway include members of the IkB. (inhibitor of kB) family and the IkB kinase (IKK) complex that consists of IKKa–IKKb heterodimers and the scaffold protein IKKg/NEMO. A variety of stimuli including the. cytokines tumor necrosis factor (TNF)-a and interleukin (IL)-1 activate the IKK complex leading to the phosphorylation of the IkB proteins. This phosphorylation leads to IkB. polyubiquitination and its subsequent degradation by the 26S proteasome. The NF-kB proteins are, thus, liberated from IkB and translocate to the nucleus where they bind. to the promoter regions of NF-kB-responsive genes to result in increased gene expression. Figure 3. Nucleosomal role of IkB (inhibitor of kB) kinase-a (IKK-a). IKKa plays important part in regulating nuclear factor (NF)-kB transcriptional activation in the nucleus. In. response to cytokines such as tumor necrosis factor (TNF)-a, IKKa is recruited to the promoter regions of NF-kB-responsive genes and forms a complex on the promoter. with p65 and the coactivator cAMP-response-element-binding protein (CREB)-binding protein (CBP). The formation of this complex facilitates the phosphorylation (P) of. histone H3 on Ser10 and leads to the subsequent acetylation (AC) by CBP to induce the expression of NF-kB-responsive genes. Yamamoto and Gaynor, (2004), TRENDS in Biochemical Sciences, 29 (2)")
7
NF-kappaB knock-outs IKK mutace Ectodermal dysplasia and immune deficiency (oportunní infekce) Caamano and Hunter (2002), Clinical Microbiology Reviews, 15 (3):414
, Clinical Microbiology Reviews, 15 (3):414.")
8
Signálová dráha NF-kappaB
Caamano and Hunter (2002), Clinical Microbiology Reviews, 15 (3):414
, Clinical Microbiology Reviews, 15 (3):414.")
9
Transaktivace kB vazebné místo: 5'-GGGRNYYYCC-3' (R - purin, Y - pyrimidin, N jakákoli báze) JANSSEN-HEININGER et al. (2000), Free Radical Biology & Medicine,28(9):131713 HAT uvolnění DNA z histonů a tak umožní nasednutí iniciačního komplexu a transkripci Transkripční koaktivátory: P/CAF Histon acetyl transferázová (HAT) aktivita CBP/p300 cAMP response element (CREB) vázající protein, HAT aktivita HMG High mobility group proteins – citlivost DNA k nukleázám RNA pol 2 RNA polymeráza 2 TFIIB Nutný pro asociaci RNA pol. s iniciačním komplexem TBP TATA-box vázající protein TAF TBP asociovaný faktor
, Free Radical Biology & Medicine,28(9): HAT uvolnění DNA z histonů a tak umožní nasednutí iniciačního komplexu a transkripci. Transkripční koaktivátory: P/CAF Histon acetyl transferázová (HAT) aktivita. CBP/p300 cAMP response element (CREB) vázající protein, HAT aktivita. HMG High mobility group proteins – citlivost DNA k nukleázám. RNA pol 2 RNA polymeráza 2. TFIIB Nutný pro asociaci RNA pol. s iniciačním komplexem. TBP TATA-box vázající protein. TAF TBP asociovaný faktor.")
10
Ubiquitin proteasomový systém
Ubiquitin – 76 Akys E1- Ub. aktivující e. E2- Ub. přenášející e. E3- Ub. ligáza

11
Kanonická dráha aktivace NF-kappaB
Hlavní dimer p65/p50 Description: Survival of an organism is dependent on its ability to rapidly and effectively respond to adverse changes in its environment. Eukaryotic cells possess a number of distinct signal transduction pathways that couple environmental stimuli to specific changes of gene expression. One such pathway is the transcription factor NF-KappaB (Nuclear Factor-KappaB), which is implicated in the regulation of many genes that code for mediators of apoptosis, viral replication, tumorigenesis, various autoimmune diseases and inflammatory responses (Ref.1). NF-KappaB is composed of homo- and heterodimeric complexes of members of the Rel (NF-KappaB) family: p50, p65 (RelA), c-Rel, p52 and RelB. The most common and best-characterized form of NF-KappaB is the p65/p50 heterodimer. Each dimer combination exhibits differences in DNA binding affinity and transactivation potential. The activation of NF-KappaB is thought to be part of a stress response as it is activated by a variety of stimuli that include growth factors, cytokines, such as TNF (Tumor Necrosis Factor) and IL-1 (Interleukin-1) lymphokines, components of bacterial cell walls, such as LPS (Lipopolysaccharide), UV, pharmacological agents, and stress (Ref.2). Depending on the stimulus, the mechanism of activation involves overlapping and nonoverlapping steps. Among all the stimuli, perhaps the most is known about the mechanism by which TNF activates NF-KappaB. In its inactive form, NF-KappaB is sequestered in the cytoplasm, bound by members of the I-KappaB family of inhibitor proteins. The activity of NF-KappaB is regulated by inhibitory proteins, which include I-KappaB-Alpha, I-KappaB-Beta, I-KappaB-Gamma, I-KappaB-Epsilon, BCL3, p105, and p100. I-KappaB proteins are phosphorylated by I-KappaB Kinase complex consisting of at least three proteins, IKK-Alpha, IKK-Beta, and IKK-Gamma (Ref.3). NF-KappaB pathway involves the interaction of the ligand with its receptor at the cell surface (TNFR), which then recruits a protein called TRADD (TNF Receptor-Associated Death Domain). This protein binds to TRAF2 (TNF Receptor-Associated Factor-2), which activates RIP (Receptor-Interacting Protein). RIP interacts with MEKK (Mitogen-Activated Protein Kinase Kinase) and NIK (NF-KappaB-Inducing Kinase) to phosphorylate and activate the IKK (I-KappaB-Alpha kinase complex). The IKK complex phosphorylates I-KappaB-Alpha, which leads to ubiquitination and then leads to the degradation of I-KappaB-Alpha by the proteosome, resulting in the translocation of NF-KappaB to the nucleus. In the nucleus it binds to its consensus sequence (5'-GGGACTTTC-3') and positively regulates the transcription of genes involved in immune and inflammatory responses, cell growth control, and apoptosis. Genes encoding cytokines, cytokine receptors, cell adhesion molecules, chemoattractant proteins, and growth regulators are positively regulated by NF-KappaB (Ref.4). TRAF2 also interacts with A20, a zinc finger protein whose expression is induced by agents that activate NF-KappaB. A20 functions to block TRAF2-mediated NF-KappaB activation IL-1 has similar downstream affects through NF-KappaB, including immunoregulation, proinflammatory, and hematopoietic activities. IL-1 induced signaling is mediated through association of IL-1 receptors with adaptor proteins, such as IRAK (IL-1 Receptor-Associated Kinases) and MyD88 (Myeloid Differentiation Primary Response Gene-88). Activation of NF-KappaB by bacterial LPS promotes the upregulation of proinflammatory cytokines. Following recognition of LPS, the adapter protein MyD88 is recruited to the cytoplasmic domain of TLR4 (Toll Like Receptor-4). MyD88 contains a highly conserved DD (Death Domain) that facilitates its interaction with another DD-containing signaling molecule, IRAK. Following recruitment to MyD88, IRAK undergoes rapid autophosphorylation and dissociation from the signaling complex. Phosphorylated IRAK subsequently interacts with TRAF6, initiating the activation of a kinase cascade involving NIK and IKK. Activation of this cascade culminates in the phosphorylation and degradation of the I-KappaB, enabling NF-KappaB to translocate to the nucleus and promote new gene expression (Ref.5). NF-KappaB-dependent gene expression involves a growing family of proteins termed transcriptional coactivators that probably function by facilitating or bridging the sequence-specific activators to the basal transcriptional machinery and altering chromatin structure. The p65 component of NF-KappaB binds to the coactivator CBP (cyclic AMP response element binding protein [CREB]-binding protein) and its structural homolog p300. Phosphorylation of p65 by PKA (Protein Kinase-A) stimulates NF-KappaB-dependent gene expression by enhancing p65 association with CBP. Recent observations have shown that levels of the CBP homolog, p300, are limiting relative to those of p65 and that competition for CBP may regulate p65 transactivation. Deacetylation of p65 through specific interactions with HDAC3 (Histone Deacetylase-3) promotes effective binding to newly synthesized I-kappaB-Alpha, which subsequently leads to I-kappaB-Alpha dependent nuclear export. Overexpression of the p110 catalytic subunit of PI3K (Phosphoinositide-3 Kinase) also induces p65 -mediated transactivation and that the specific PI3K inhibitor LY represses this process. Additionally, the expression of a constitutively activated form of either p110 or the PI3K-activated protein kinase Akt also induces p65/RelA-mediated transactivation. NF-KappaB pathways provide many targets for developing specific drugs to treat inflammatory diseases. Inappropriate activation of NF-KappaB has been linked to inflammatory events associated with autoimmune arthritis, asthma, septic shock, lung fibrosis, glomerulonephritis,
, which is implicated in the regulation of many genes that code for. mediators of apoptosis, viral replication, tumorigenesis, various autoimmune diseases and inflammatory. responses (Ref.1). NF-KappaB is composed of homo- and heterodimeric complexes of members of the. Rel (NF-KappaB) family: p50, p65 (RelA), c-Rel, p52 and RelB. The most common and best-characterized. form of NF-KappaB is the p65/p50 heterodimer. Each dimer combination exhibits differences in DNA. binding affinity and transactivation potential. The activation of NF-KappaB is thought to be part of a stress. response as it is activated by a variety of stimuli that include growth factors, cytokines, such as TNF. (Tumor Necrosis Factor) and IL-1 (Interleukin-1) lymphokines, components of bacterial cell walls, such as. LPS (Lipopolysaccharide), UV, pharmacological agents, and stress (Ref.2). Depending on the stimulus, the mechanism of activation involves overlapping and nonoverlapping steps. Among all the stimuli, perhaps the most is known about the mechanism by which TNF activates. NF-KappaB. In its inactive form, NF-KappaB is sequestered in the cytoplasm, bound by members of the. I-KappaB family of inhibitor proteins. The activity of NF-KappaB is regulated by inhibitory proteins, which. include I-KappaB-Alpha, I-KappaB-Beta, I-KappaB-Gamma, I-KappaB-Epsilon, BCL3, p105, and p100. I-KappaB proteins are phosphorylated by I-KappaB Kinase complex consisting of at least three proteins, IKK-Alpha, IKK-Beta, and IKK-Gamma (Ref.3). NF-KappaB pathway involves the interaction of the ligand with its receptor at the cell surface (TNFR), which then recruits a protein called TRADD (TNF Receptor-Associated Death Domain). This protein binds. to TRAF2 (TNF Receptor-Associated Factor-2), which activates RIP (Receptor-Interacting Protein). RIP. interacts with MEKK (Mitogen-Activated Protein Kinase Kinase) and NIK (NF-KappaB-Inducing Kinase) to. phosphorylate and activate the IKK (I-KappaB-Alpha kinase complex). The IKK complex phosphorylates. I-KappaB-Alpha, which leads to ubiquitination and then leads to the degradation of I-KappaB-Alpha by the. proteosome, resulting in the translocation of NF-KappaB to the nucleus. In the nucleus it binds to its. consensus sequence (5 -GGGACTTTC-3 ) and positively regulates the transcription of genes involved in. immune and inflammatory responses, cell growth control, and apoptosis. Genes encoding cytokines, cytokine receptors, cell adhesion molecules, chemoattractant proteins, and growth regulators are positively. regulated by NF-KappaB (Ref.4). TRAF2 also interacts with A20, a zinc finger protein whose expression is. induced by agents that activate NF-KappaB. A20 functions to block TRAF2-mediated NF-KappaB. activation. IL-1 has similar downstream affects through NF-KappaB, including immunoregulation, proinflammatory, and hematopoietic activities. IL-1 induced signaling is mediated through association of IL-1 receptors with. adaptor proteins, such as IRAK (IL-1 Receptor-Associated Kinases) and MyD88 (Myeloid Differentiation. Primary Response Gene-88). Activation of NF-KappaB by bacterial LPS promotes the upregulation of. proinflammatory cytokines. Following recognition of LPS, the adapter protein MyD88 is recruited to the. cytoplasmic domain of TLR4 (Toll Like Receptor-4). MyD88 contains a highly conserved DD (Death. Domain) that facilitates its interaction with another DD-containing signaling molecule, IRAK. Following. recruitment to MyD88, IRAK undergoes rapid autophosphorylation and dissociation from the signaling. complex. Phosphorylated IRAK subsequently interacts with TRAF6, initiating the activation of a kinase. cascade involving NIK and IKK. Activation of this cascade culminates in the phosphorylation and. degradation of the I-KappaB, enabling NF-KappaB to translocate to the nucleus and promote new gene. expression (Ref.5). NF-KappaB-dependent gene expression involves a growing family of proteins termed. transcriptional coactivators that probably function by facilitating or bridging the sequence-specific. activators to the basal transcriptional machinery and altering chromatin structure. The p65 component of. NF-KappaB binds to the coactivator CBP (cyclic AMP response element binding protein [CREB]-binding. protein) and its structural homolog p300. Phosphorylation of p65 by PKA (Protein Kinase-A) stimulates. NF-KappaB-dependent gene expression by enhancing p65 association with CBP. Recent observations. have shown that levels of the CBP homolog, p300, are limiting relative to those of p65 and that competition. for CBP may regulate p65 transactivation. Deacetylation of p65 through specific interactions with HDAC3. (Histone Deacetylase-3) promotes effective binding to newly synthesized I-kappaB-Alpha, which. subsequently leads to I-kappaB-Alpha dependent nuclear export. Overexpression of the p110 catalytic. subunit of PI3K (Phosphoinositide-3 Kinase) also induces p65 -mediated transactivation and that the. specific PI3K inhibitor LY represses this process. Additionally, the expression of a constitutively. activated form of either p110 or the PI3K-activated protein kinase Akt also induces p65/RelA-mediated. transactivation. NF-KappaB pathways provide many targets for developing specific drugs to treat. inflammatory diseases. Inappropriate activation of NF-KappaB has been linked to inflammatory events. associated with autoimmune arthritis, asthma, septic shock, lung fibrosis, glomerulonephritis,")
12
Alternativní dráha aktivace NF-kappaB
(LTbR, BAFF-R,CD40, Fn14, RANK, CD27, CD30, LMP1) Aktivace - několik minut Aktivace - několik hodin Kinase cascade Fig. 2 – Model for the activation of the classical and the alternative NF-kB pathways by TNFR-related proteins (LTbR, BAFF-R, CD40, Fn14, RANK, CD27, CD30 and LMP1) and upon H. pylori infection. Upon receptor activation, TRAF proteins mediate the activation of the classical NF-kB pathway probably via the requirement of an upstreamkinase (still unknown),which activates the IKK complex. The latter phosphorylates IkBs (IkBa and IkBb), which become ubiquitinated and then fully degraded through the 26 S proteasome. Thus, NF-kB complexes p50/X and p52/X (X = p65 or c-Rel) are freed and translocate into the nucleus for modulating transiently the expression of their target genes. While the classical pathway is turned on within minutes, activation of the alternative pathway takes about a few hours. According to the receptor, TRAF proteins are either activated, or inhibited, in order to triggerNIK and IKKa kinase activities. NIK mediates p100 phosphorylation/ubiquitination in a IKKa-independent and IKKa-dependent manners. The phospho-p100 is recognized and polyubiquitinated by the E3 ligase SCFb-TrCP (not pictured here). Then, the phospho-ubiquitinated p100 interacts with the S9 subunit of the 19 S proteasome lid and is partially degraded through the proteasome for generating p52/RelB and p52/X dimers. Once generated, p52/RelB dimers are free tomove to the nucleuswhereas p52/X dimers are first captured by IkBs for their cytosolic retention and then activated through the classical pathway. Thus,while p52/RelB complexes are generated and activated through the alternative pathway, pre-bound p52/X dimers are generated via the alternative pathway but activated through the classical pathway. Therefore, these particular dimers are controlled by a ‘‘hybrid pathway’’. Dejardin, biochemical pharmacology 72 (2006) 1161–1179
 Aktivace - několik minut Aktivace - několik hodin. Kinase cascade. Fig. 2 – Model for the activation of the classical and the alternative NF-kB pathways by TNFR-related proteins (LTbR, BAFF-R, CD40, Fn14, RANK, CD27, CD30 and LMP1) and upon H. pylori infection. Upon receptor activation, TRAF proteins mediate the. activation of the classical NF-kB pathway probably via the requirement of an upstreamkinase (still unknown),which activates. the IKK complex. The latter phosphorylates IkBs (IkBa and IkBb), which become ubiquitinated and then fully degraded. through the 26 S proteasome. Thus, NF-kB complexes p50/X and p52/X (X = p65 or c-Rel) are freed and translocate into the. nucleus for modulating transiently the expression of their target genes. While the classical pathway is turned on within. minutes, activation of the alternative pathway takes about a few hours. According to the receptor, TRAF proteins are either. activated, or inhibited, in order to triggerNIK and IKKa kinase activities. NIK mediates p100 phosphorylation/ubiquitination in. a IKKa-independent and IKKa-dependent manners. The phospho-p100 is recognized and polyubiquitinated by the E3 ligase. SCFb-TrCP (not pictured here). Then, the phospho-ubiquitinated p100 interacts with the S9 subunit of the 19 S proteasome lid. and is partially degraded through the proteasome for generating p52/RelB and p52/X dimers. Once generated, p52/RelB dimers. are free tomove to the nucleuswhereas p52/X dimers are first captured by IkBs for their cytosolic retention and then activated. through the classical pathway. Thus,while p52/RelB complexes are generated and activated through the alternative pathway, pre-bound p52/X dimers are generated via the alternative pathway but activated through the classical pathway. Therefore, these particular dimers are controlled by a ‘‘hybrid pathway’’. Dejardin, biochemical pharmacology 72 (2006) 1161–1179.")
13
Dráhy aktivované TNF TNFa – prozánětlivý, proapoptotický cytokin
Nejsilnější induktor NF-kappaB aktivace NIK – NF-kappaB indukující kináza IKK – IkappaB kináza MEKKs – MAPK/ERK kináza kinázy MAPK - Mitogenem aktivovaná protein kináza (JNK, p38, ERK) ERK - Extracelulárním signálem regulovaná kináza JNK – cJun N-terminální kináza p38 – stresem aktivovaná kináza
 ERK - Extracelulárním signálem regulovaná kináza. JNK – cJun N-terminální kináza. p38 – stresem aktivovaná kináza.")
14
NF-kB a apoptóza DISC - – death inducing signaling complex Anti-
apoptotické cílové geny NFkB: c-Flip c-IAPs Bcl-XL A1 (Bcl-2 family) TRAF1/2 Pro-apoptotické cílové geny NF-kB: RIP, DR1-6, Fas, TNF-R
 TRAF1/2. Pro-apoptotické. cílové geny NF-kB: RIP, DR1-6, Fas, TNF-R.")
15
Podpora přirozené imunity
Produkce pro-zánětlivých působků Interleukiny, chemokiny, TNF; IFNb, RANTES Imunoreceptory (TNF-R, MHCI, imunoglobuliny) Buněčná adheze (ICAM1, ELAM1, VCAM1, selektiny, tenascin C) Proteiny akutní fáze zánětu (faktory komplementu, CRP, LPS R, angiotensinogen) Členové rodiny NF-kappaB Anti-apoptotická role je spojena s podporou zánětlivého procesu Apoptotická tělíska pohlcená makrofágy jsou neimunogenní Role Bcl-3 v T lymfocytech Vazba na homodimery p50 a p52 v jádře mění tyto represory na aktivátory transkripce Podpora dozrávání B lymfocytů IKKa podporuje aktivaci dimerů p52/RelB – produkce proteinů posilujících rezistenci B lymfocytů k apoptóze x PODPORA CHRONICKÉHO ZÁNĚTU!!! (revmatoidní artritida, kardiovaskulární nemoci, autoimunitní nemoci)
 Buněčná adheze (ICAM1, ELAM1, VCAM1, selektiny, tenascin C) Proteiny akutní fáze zánětu (faktory komplementu, CRP, LPS R, angiotensinogen) Členové rodiny NF-kappaB. Anti-apoptotická role je spojena s podporou zánětlivého procesu. Apoptotická tělíska pohlcená makrofágy jsou neimunogenní. Role Bcl-3 v T lymfocytech. Vazba na homodimery p50 a p52 v jádře mění tyto represory na aktivátory transkripce. Podpora dozrávání B lymfocytů. IKKa podporuje aktivaci dimerů p52/RelB – produkce proteinů posilujících. rezistenci B lymfocytů k apoptóze. x PODPORA CHRONICKÉHO ZÁNĚTU!!! (revmatoidní artritida, kardiovaskulární nemoci, autoimunitní nemoci)")
16
Podpora proliferace Cytokiny a jejich receptory produkované působením NF-kappaB působí jako růstové faktory - Aktivuje geny pro G-, M-, GM-CSF, VEGF Aktivuje expresi cyklinu D1 nutného pro přechod G1/S Aktivuje expresi c-Myc – transaktivuje geny pro cykliny a potlačuje expresi p21 v G1,G2 fázi – hl. role Akt - fosforyluje IKK + Chk1 je inaktivní v S fázi je to naopak + role RelA a p100

17
Regulátor stresové odpovědi
Stresory aktivující NF-kappaB: Infekce Ischemický/reperfúzní stres Hemorhagický šok Poškození jater Ozáření organismu (UV, g) Oxidační (ROS (?) a NOS metabolizmus) Hyperglykemický stres Xenobiotický (cigaretový kouř, těžké kovy, PCB, cytostatika)
 Oxidační (ROS ( ) a NOS metabolizmus) Hyperglykemický stres. Xenobiotický (cigaretový kouř, těžké kovy, PCB, cytostatika)")
18
NO metabolizmus aktivace nitrosylace
Fig. 2. Control of NF-kB activation by •NO. Low levels of •NO augment NF-kB activity potentially via regulation of the activity of IKKs. In contrast, high levels of •NO produced from iNOS or high concentrations of donors inhibit NF-kB in part via nitrosylation of cysteine 62 of p50 and provides a negative feedback loop to block prolonged activation. Free Radical Biology & Medicine, Vol. 28, No. 9, pp. 1317–1327, 2000 aktivace nitrosylace JANSSEN-HEININGER, et al.,Free Radical Biology & Medicine, 28, 9, 1317–1327, 2000

19
Podmínky vhodné pro karcinogenezi
Vliv na karcinogenezi NF-kappaB Potlačuje rozvoj apoptózy + Indukuje proliferaci, angiogenezi Zvyšuje metastatický potenciál Podporuje zánětlivé procesy = Podmínky vhodné pro karcinogenezi

20
Karcinogenetický potenciál NF-kappaB
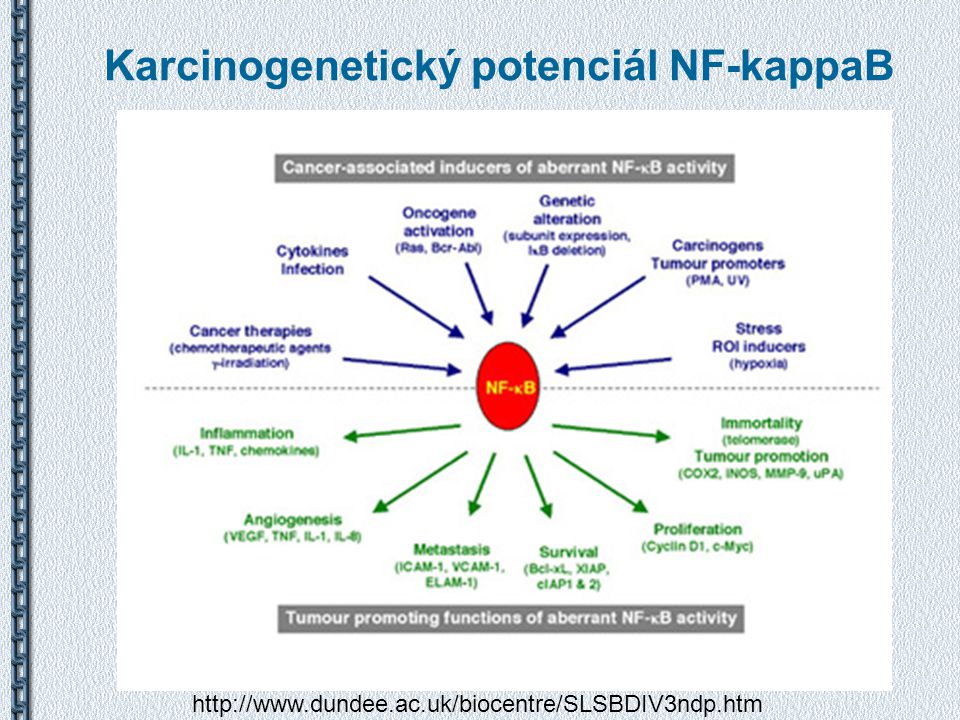
21
Modulace aktivity NF-kappaB
Tumor supresorové proteiny p53 a ARF indukují asociaci aktivního NF-kappaB s HDAC1 => Represe NF-kappaB aktivity Tumor supresory p53 a ARF indukují asociaci aktivovaných NFkB podjednotek s histon deacetylázami a tím potlačují expresi cílových genů NFkB Two roles for NF-kB in tumorigenesis. In this model, the broken arrows represent the progression of a normal cell into a transformed and malignant cancer cell. In normal cells, when an oncogenic signal, such as DNA damage, oncogene activation or infection by oncogenic viruses, is received, tumor suppressors, such as p53 and ARF, are activated. These tumor suppressors can modulate the activity of NF-kB complexes, which are either preexisting within the cell or are activated by the oncogenic stimuli. Consequently, the tumor-promoting characteristics of NF-kB are neutralized, ‘recruiting’ it to the tumor suppressor program (Figure 1). However, as the potential tumor cell accumulates additional mutations, tumor suppressor activity of NF-kB is lost, its oncogenic functions are now unleashed, and it can actively contribute to the growth and development of the cancer cell. Perkins (2004), Trends in Cell Biology, 14(2)
. However, as the potential. tumor cell accumulates additional mutations, tumor suppressor activity of NF-kB is lost, its oncogenic functions are now unleashed, and it can actively contribute to the. growth and development of the cancer cell. Perkins (2004), Trends in Cell Biology, 14(2)")
22
Detekce aktivního NF-kappaB
WB Degradace IkappaB - Nárůst p50 a p65 v nukleárním extraktu a úbytek v cytoplazmatickém extraktu EMSA - Electrophoretic Mobility Shift Assay NF-kappaB v nukleárním extraktu se váže na značené oligonukleotidy obsahující kB vazné místo Detekce komplexu nativní PAGE Reportérový plazmid – Transfekce buněk plazmidem nesoucím kB vazná místa v promotoru genu pro luciferázu/B-gal/CAT

23
Shrnutí Rodina transkripčních faktorů NF-kappaB
Klasická vs. alternativní dráha aktivace Nejsilnější aktivátor NF-kappaB je TNF-a Pro-zánětlivé a přežití podporující účinky Aktivován stresem Podílí se na rozvoji chronického zánětu a nemocí s ním spojených Podporuje rozvoj nádorových onemocnění

Podobné prezentace



















