Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
OXIDATIVNÍ FOSFORYLACE
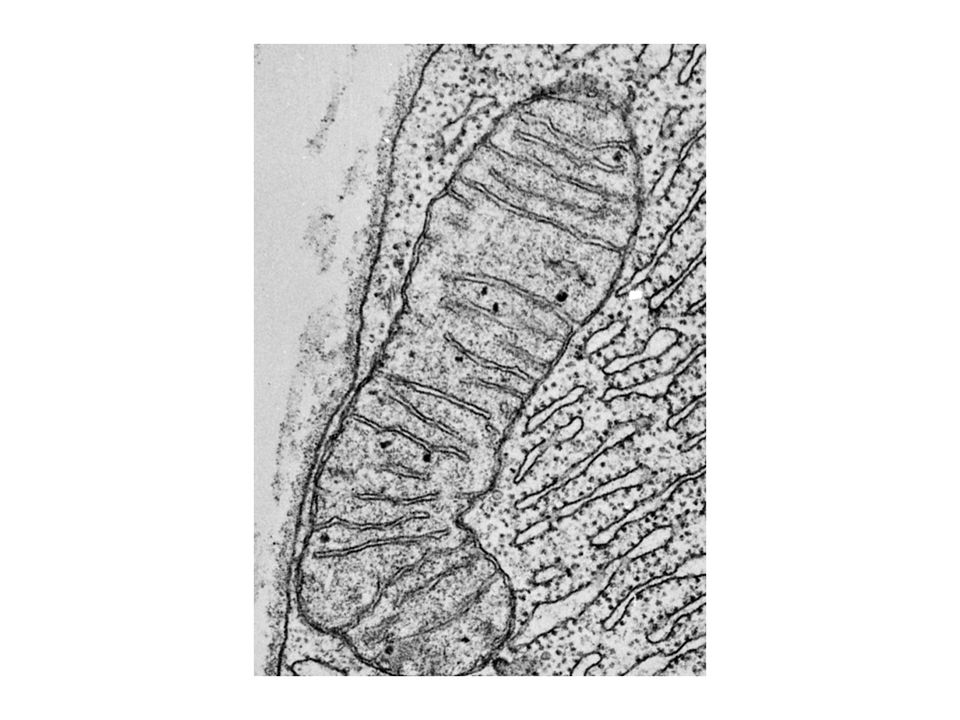
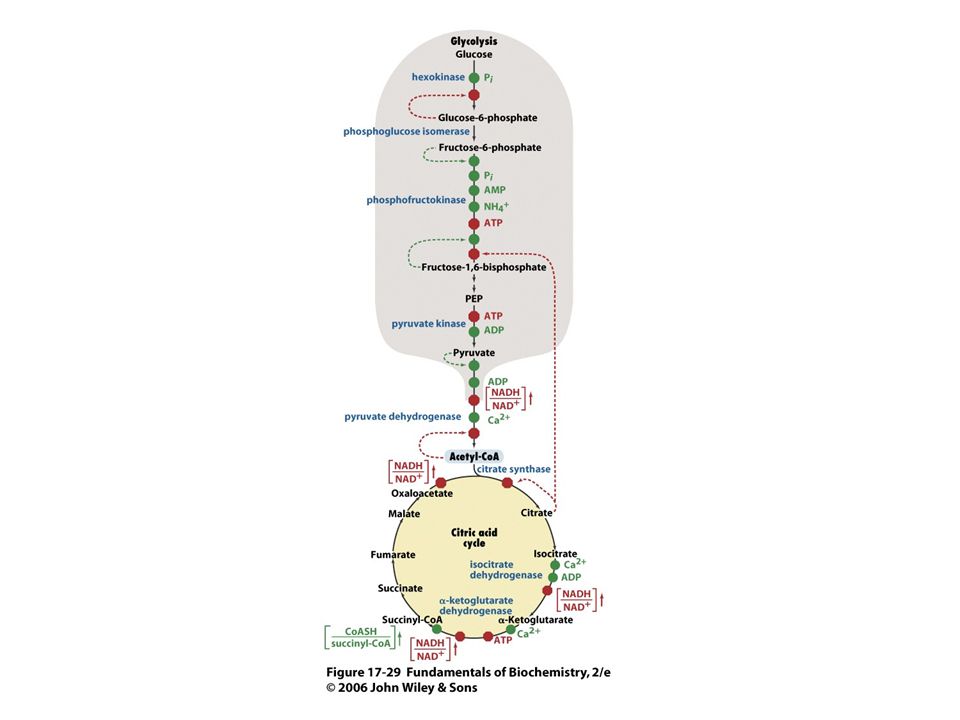
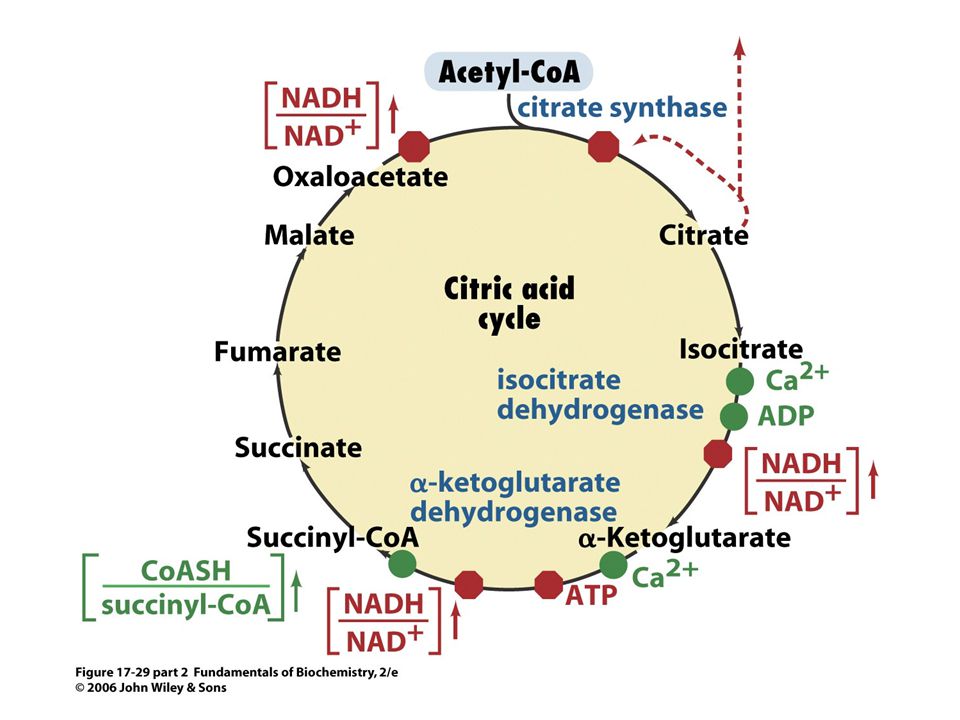
NADH a FADH2 vytvořené při glykolýze, oxidaci mastných kyselin a cyklu trikarboxylových kyselin jsou molekuly bohaté na energii. Při jejich oxidaci za tvorby vody se uvolňuje energie, která se ukládá ve formě ATP. Oxidativní fosforylace je proces tvorby ATP při přenosu elektronů z NADH a FADH2 na kyslík přes řadu přenašečů elektronů. Oxidativní fosforylace je hlavním zdrojem energie aerobních organismů.

4
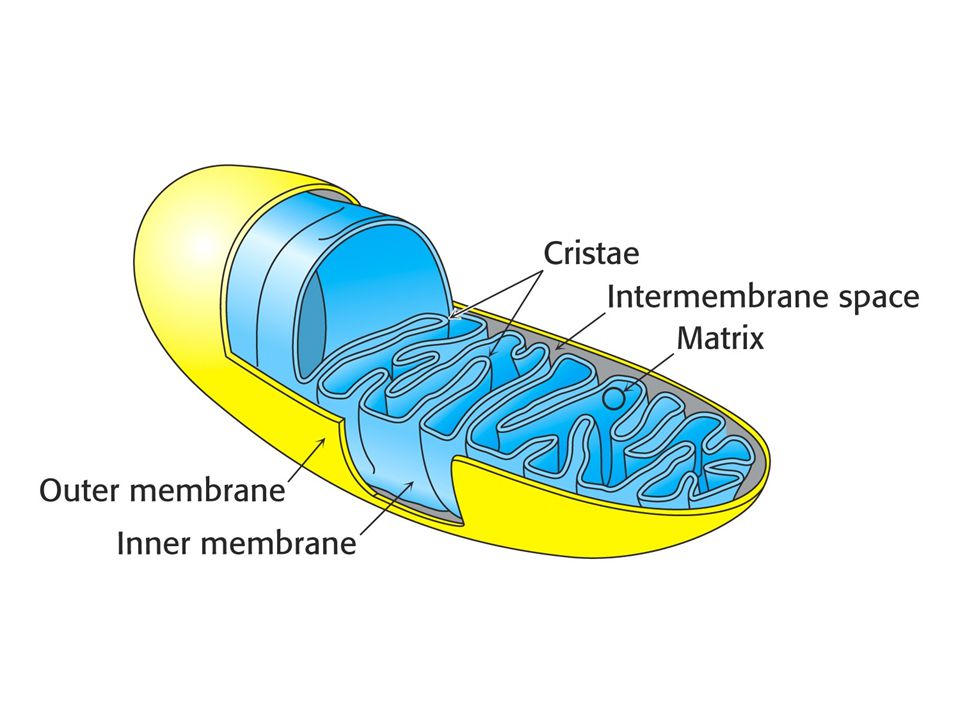
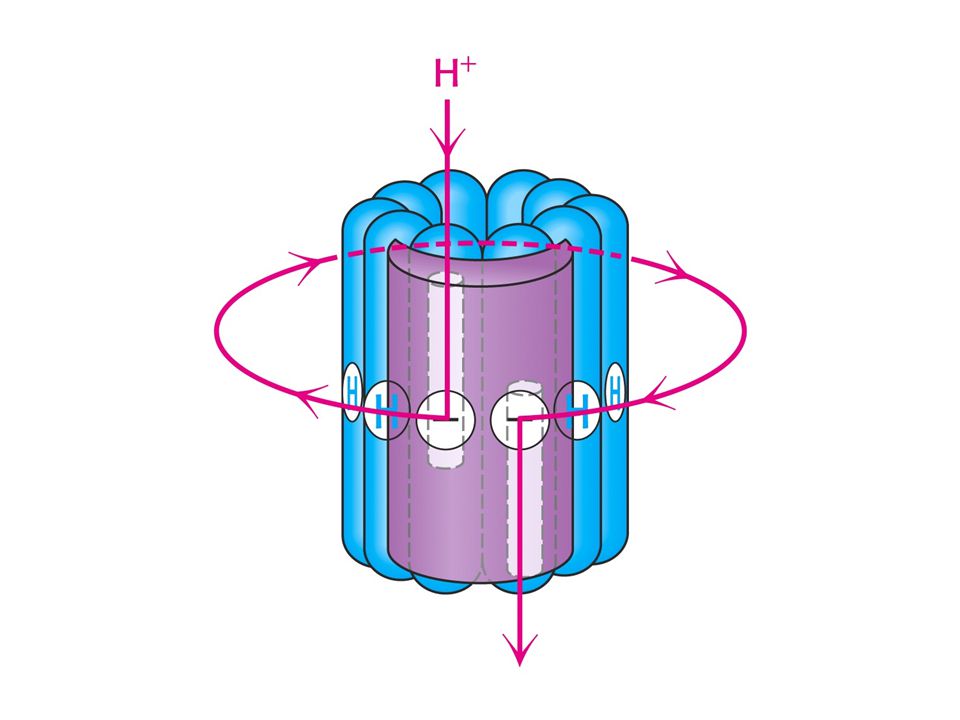
Oxidace a syntéza ATP jsou spojeny transmembránovým tokem protonů

5
Zařízení pro měření standardního redukčního (oxidačně-redukčního) potenciálu.
 potenciálu.")
6
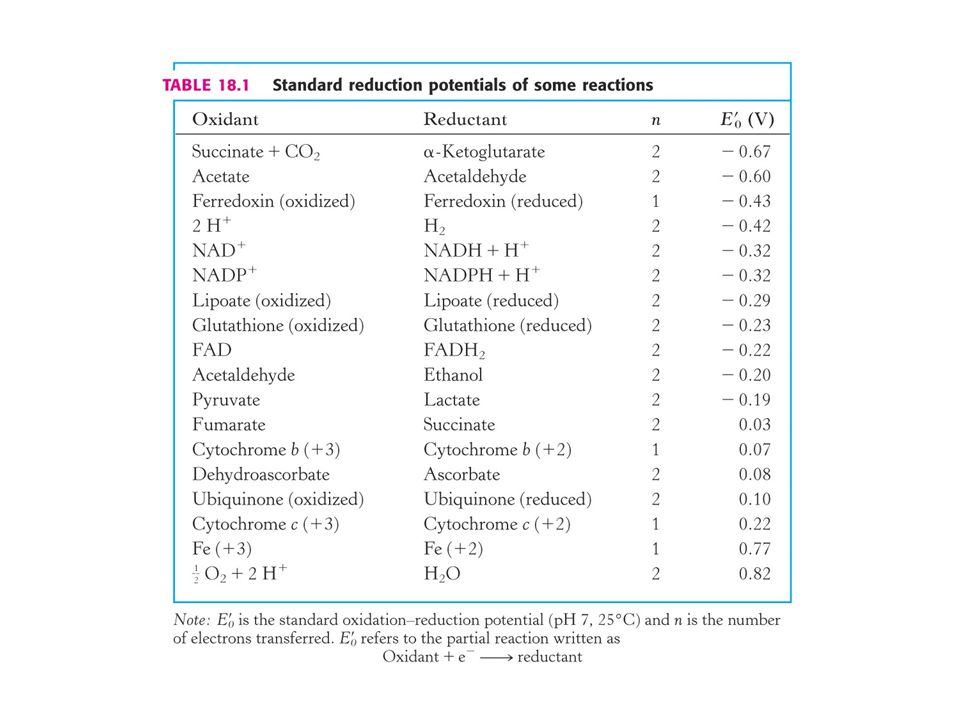
Redukční potenciál páru H+ : H2 je definován jako O voltů.
Např. NADH jako silné redukční činidlo předává elektrony a má tak negativní redukční potenciál. Silné oxidační činidlo jako je O2 přijímá elektrony a má tak pozitivní redukční potenciál. V BIOCHEMII: V biochemických textech je redukční potenciál uváděn při koncentraci H+ = 10-7M (pH 7), zatímco v chemických 1 M (pH O) Redukční potenciál v biochemii je tak označen jako Eo´ (obdobně jako D Go´) Vztah mezi změnou standardní volné (Gibbsovy) energie a změnou redukčního potenciálu: D Go´ = - nFDEo´ n = počet přenášených elektronů, F je Faradayův náboj 96, 48 kJ.mol-1.V-1) a DEo´ ve voltech.
, zatímco v chemických 1 M. (pH O) Redukční potenciál v biochemii je tak označen jako Eo´ (obdobně jako D Go´) Vztah mezi změnou standardní volné (Gibbsovy) energie. a změnou redukčního potenciálu: D Go´ = - nFDEo´ n = počet přenášených elektronů, F je Faradayův náboj. 96, 48 kJ.mol-1.V-1) a DEo´ ve voltech.")
8
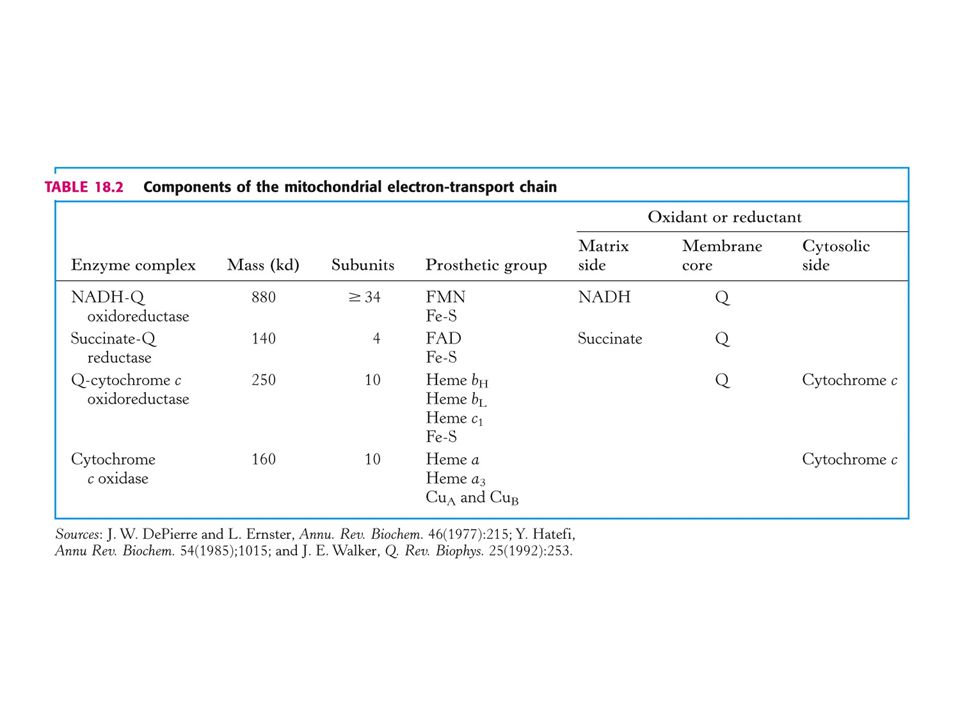
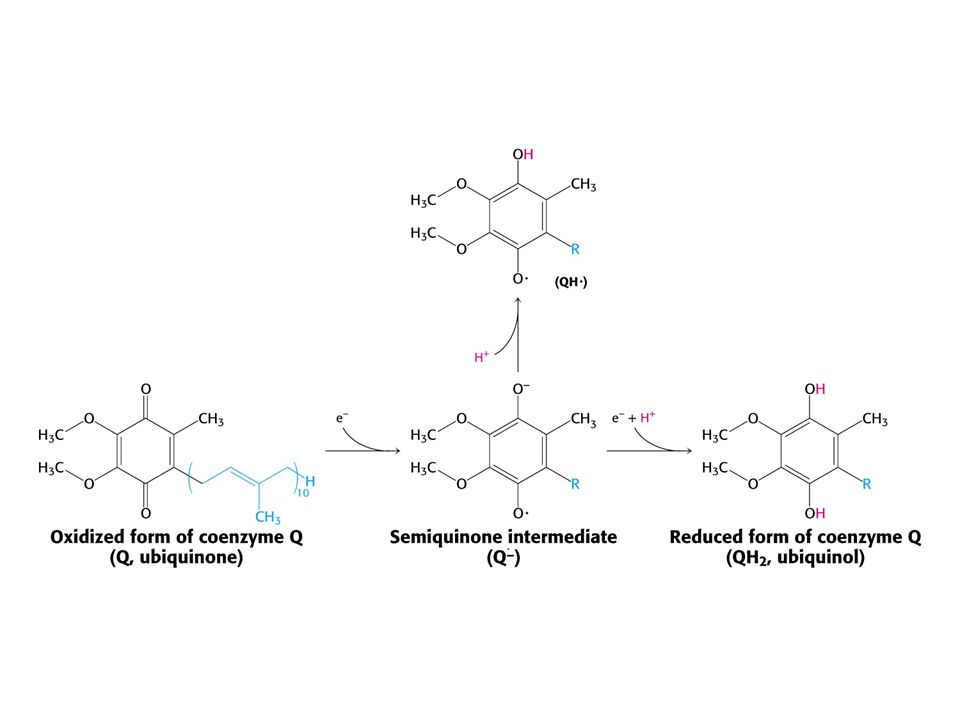
Sekvence elektronových přenašečů v dýchacím řetězci

10
Struktura NADH-Qoxidoreduktasy (Komplex I) Skládá se z části lokalizované v membráně a ramene v matrix. NADH se oxiduje v rameni a elektrony jsou přeneseny do membrány, kde redukují Q.

11
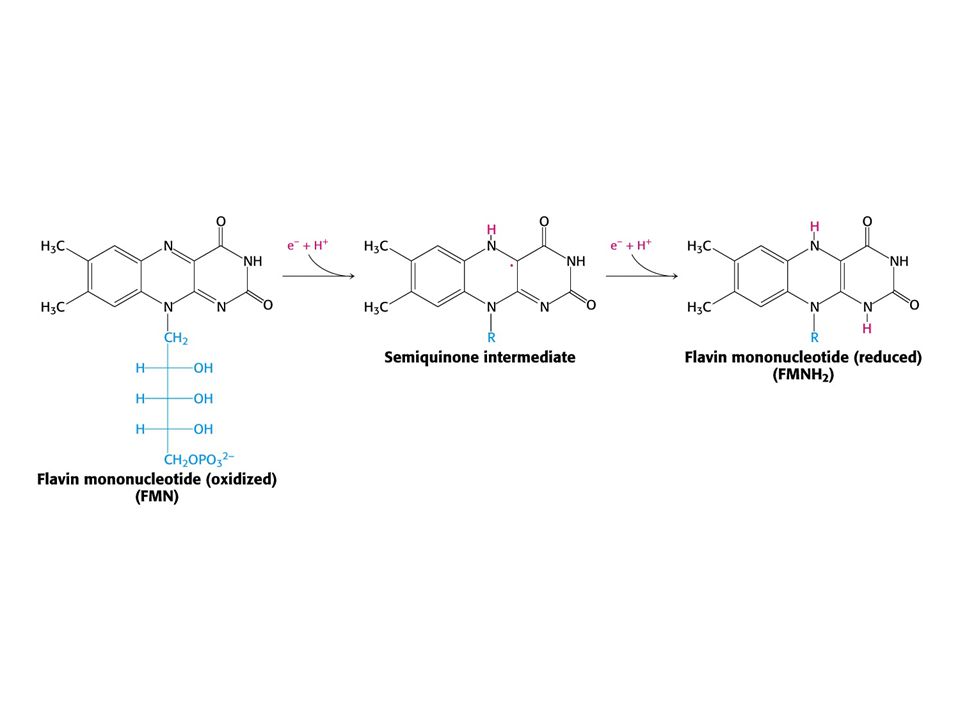
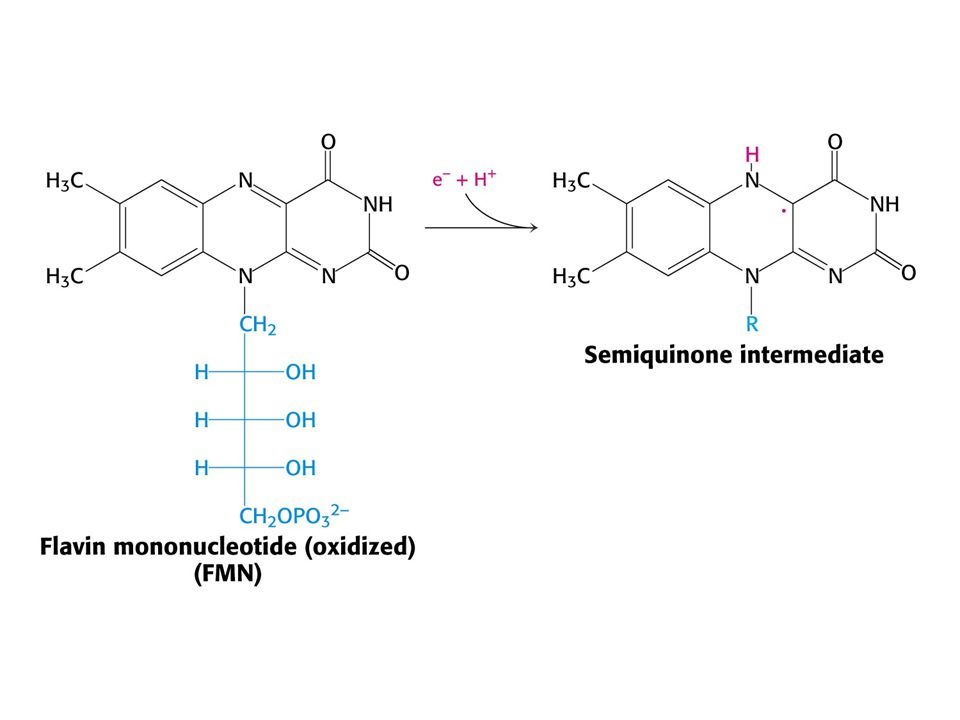
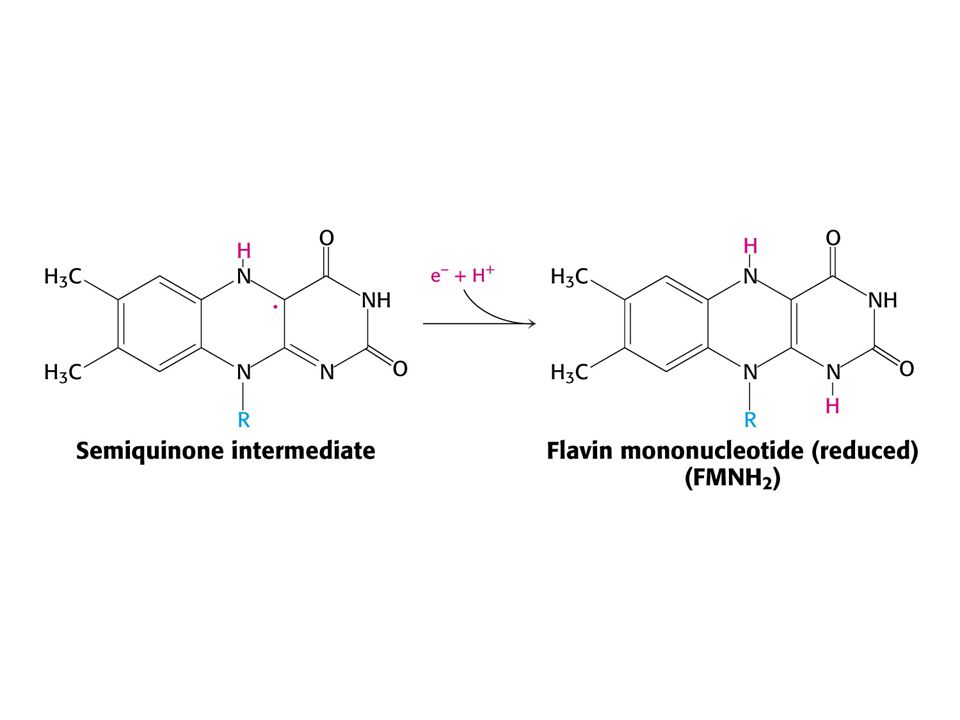
Reakce katalyzovaná NADH-Q oxidoreduktasou.
NADH + Q H+matrix NAD+ + QH2 + 4 H+cytoplasma První krok: vazba NADH a přenos dvou elektronů na FMN (FMNH2). Druhý krok: elektrony jsou přeneseny na soustavu sloučenin železo – síra. Třetí krok: Elektrony jsou přeneseny na vázaný Q. Tok dvou elektronů z NADH na vázaný Q přes komplex I pumpuje čtyři elektrony z matrix. Redukce Q na QH2 vyžaduje další dva protony z matrix. Čtvrtý krok: Pár elektronů z vázaného QH2 se přenáší na 4Fe – 4S a protony se uvolňují do cytoplasmy. Pátý krok: Elektrony z 4Fe – 4S se přenáší na mobilní Q v hydrofobní části membrány. To vede k odebrání dalších dvou protonů z matrix.
. Druhý krok: elektrony jsou přeneseny na soustavu sloučenin železo – síra. Třetí krok: Elektrony jsou přeneseny na vázaný Q. Tok dvou elektronů z NADH na vázaný Q přes komplex I pumpuje čtyři elektrony z matrix. Redukce Q na QH2 vyžaduje další dva protony z matrix. Čtvrtý krok: Pár elektronů z vázaného QH2 se přenáší na 4Fe – 4S a protony se uvolňují do cytoplasmy. Pátý krok: Elektrony z 4Fe – 4S se přenáší na mobilní Q v hydrofobní části membrány. To vede k odebrání dalších dvou protonů z matrix.")
16
Nehemové železnaté proteiny Iont železa vázaný čtyřmi cysteiny

17
2Fe – 2S „cluster“ seskupení s ionty železa vázanými sulfidovými ionty.

18
4Fe – 4S „cluster“ seskupení.

19
Spojení přenosu elektron – proton
Spojení přenosu elektron – proton. Redukce chinonu (Q) na QH2 vede k odčerpání dvou protonů z matrix
 na QH2 vede k odčerpání dvou protonů z matrix.")
20
Ubichinol - vstup elektronů z FADH2.
Sukcinátdehydrogenasa tvořící FADH2 v citrátovém cyklu je součástí sukcinát-Q reduktasového komplexu (Komplex II). Komplex II je integrální součástí vnitřní mitochondriální membrány. FADH2 komplex neopouští. Elektrony jsou přenášeny na Fe – S komplex a poté na ubichinon za tvorby ubichinolu. Další dva enzymy přenášející své elektrony také na FADH2 a poté na komplex II. a) Glycerolfosfátdehydrogenasa b) AcylCoAdehydrogenasa (mastné kyseliny) Sukcinát-Q-reduktasový komplex netransportuje protony!!!! Získá se méně ATP !!!!
. Komplex II je integrální součástí vnitřní mitochondriální membrány. FADH2 komplex neopouští. Elektrony jsou přenášeny na Fe – S komplex a poté na ubichinon za tvorby ubichinolu. Další dva enzymy přenášející své elektrony také na FADH2 a poté na komplex II. a) Glycerolfosfátdehydrogenasa. b) AcylCoAdehydrogenasa (mastné kyseliny) Sukcinát-Q-reduktasový komplex netransportuje protony!!!! Získá se méně ATP !!!!")
21
Q-cytochrom c oxidoreduktasa (Komplex III)
Druhá protonová pumpa. Funkcí komplexu III je katalyzovat přenos elektronů z QH2 na oxidovaný cytochrom c za současného pumpování protonů z matrix. QH2 + 2 Cyt cox + 2 H+matrix Q + 2 Cyt cred + 4 H+cytoplasma

22
Vazba Cyt c na protein. Vazba vzniká adicí SH skupiny Cys na vinylskupiny protoporfyrinu

23
Q cyklus. Přechod od dvouelektronového přenosu na jednoelektronový.
Dva elektrony z vázaného QH2 jsou přeneseny takto: Jeden na cyt c a druhý na vázaný Q za tvorby semichinonu. Nově vytvořený Q oddisociuje a je nahrazen druhým QH2, který obdobně poskytne své dva elektrony. Přenos druhého elektronu na semichinon vede k pumpování dvou protonů z matrix.

24
Stryer – Conc. Insights 18. ATPasa-19

25
Transmembránový protonový transport. Q cyklus.

26
Cytochrom c oxidasa (Komplex IV)
Oxidace redukovaného cyt c spojená s redukcí O2 na 2 H2O. Rovnice: 4 cyt cred + 8 H+matrix + O cyt cox + 2 H2O + 4 H+cytoplasma Všech osm protonů je z matrix !! Enzym neuvolňuje částečně oxidované kyslíkaté meziprodukty !! Brání tak vzniku reaktivních kyslíkatých radikálů.

27
Transport protonů z matrix cytochrom c oxidasou
Transport protonů z matrix cytochrom c oxidasou. Čtyři protony do cytoplasmy a čtyři na tvorbu vody.

28
Conceptual Insights

29
Protonový gradient pohání syntézu ATP. Chemiosmotická hypotéza.

30
Důkaz chemiosmotické hypotézy (Autor hypotézy Peter Mitchel, 1961).
.")
31
Procesy oxidace NADH a tvorby ATP
NADH + ½ O2 + H H2O + NAD+ D Go´ = - 220, 1 kJ/mol ADP + Pi + H ATP + H2O D Go´ = + 30, 5 kJ/mol

32
Struktura ATPsynthasy (mitochondriální ATPasa nebo F1FoATPasa)
Struktura ATPsynthasy (mitochondriální ATPasa nebo F1FoATPasa). Dvě části – protonvodivá (Fo) a katalytická (F1). Fo je v membráně a F1 v matrix.
. Dvě části – protonvodivá (Fo) a katalytická (F1). Fo je v membráně a F1 v matrix.")
34
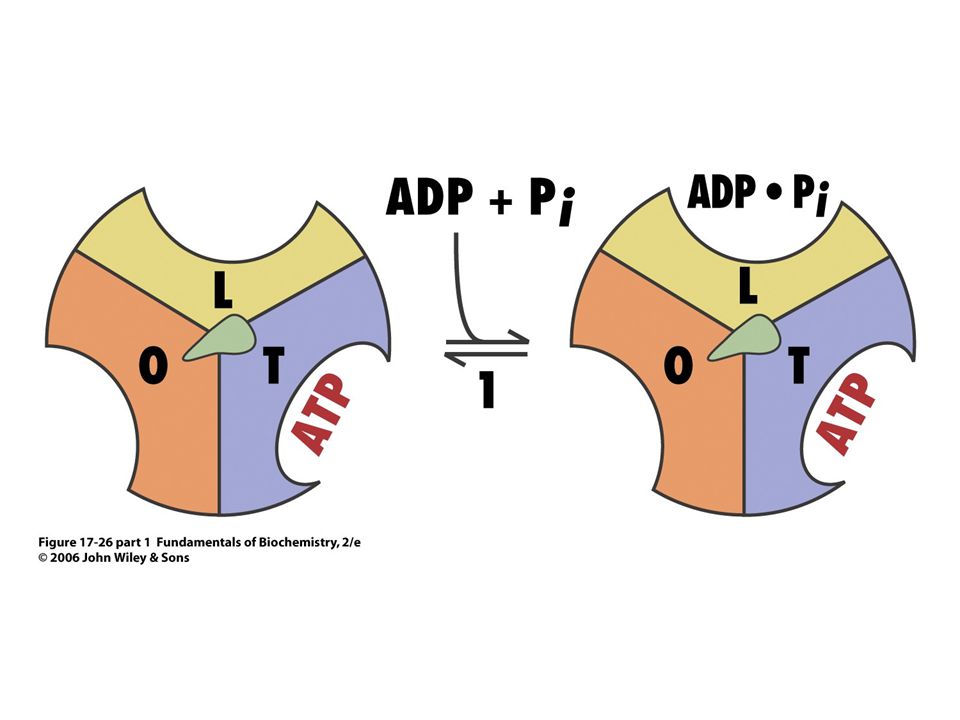
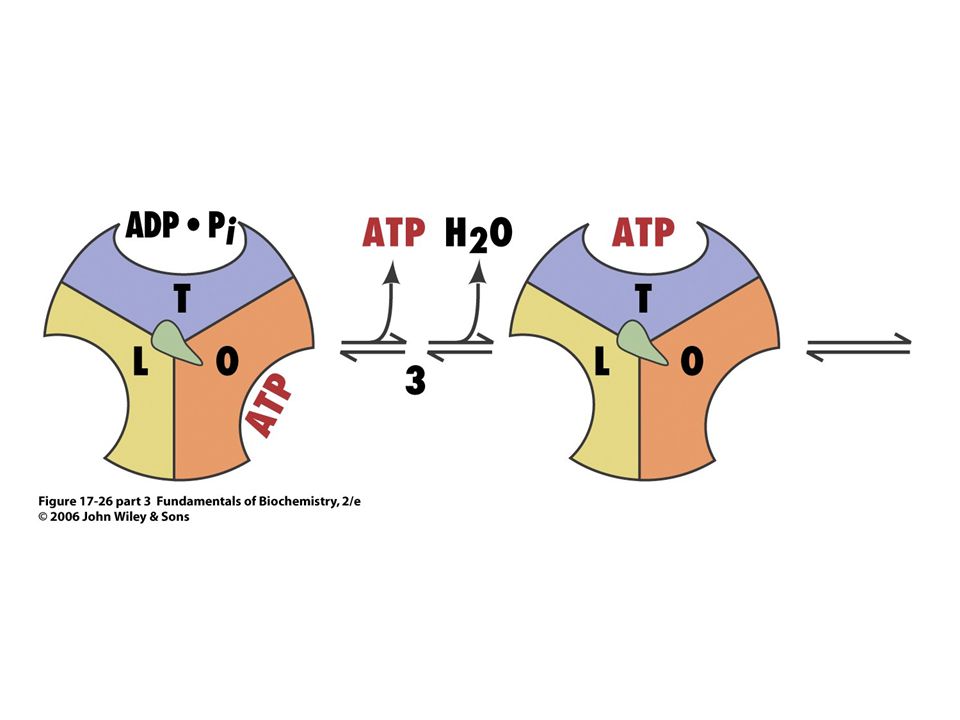
Mechanismus syntézy ATP
Role protonového gradientu nespočívá v syntéze ATP, ale v jeho uvolnění ze synthasy !!!

35
Tok protonů přes ATPsynthasu vede k uvolnění pevně vázaného ATP.
Rotace g podjednotky o 120o proti směru hodinových ručiček mění strukturu tří b podjednotek. Podjednotka s nově syntetizovaným ATP je ve formě T (tight) – nemůže uvolnit ATP. Pohybem podjednotky g se převede na O formu, uvolní ATP a váže nové ADP a Pi)
 – nemůže uvolnit ATP. Pohybem podjednotky g se převede na O formu, uvolní ATP a váže nové ADP a Pi)")
36
ATP se tvoří i bez protonmotivní síly, ale neuvolňuje se (hydrolyzuje)
")
41
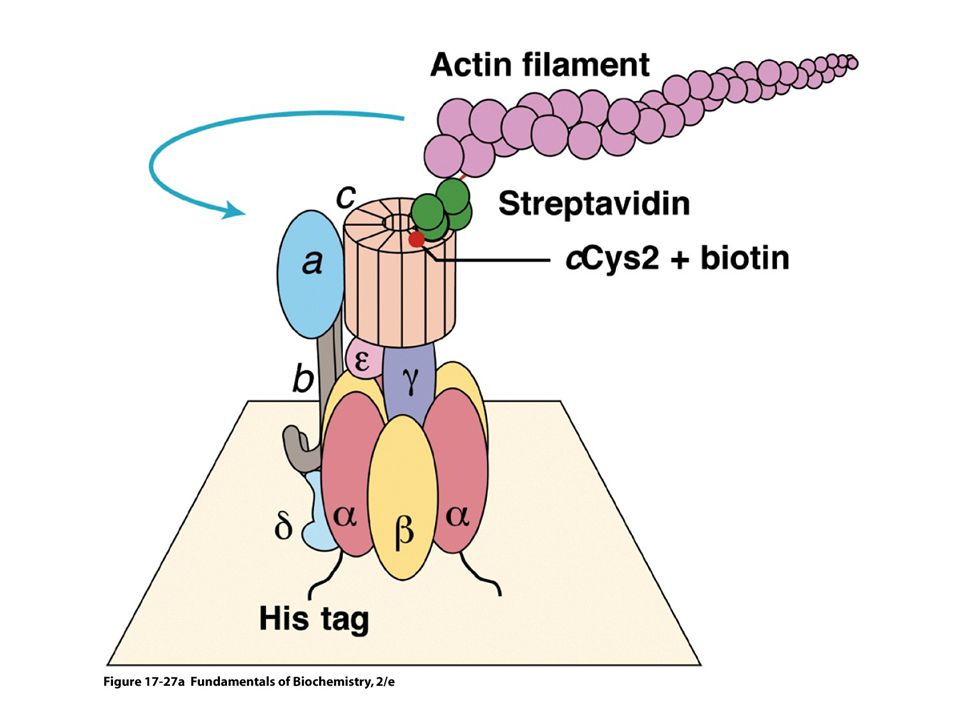
Přímé pozorování ATP poháněné rotace ATPsynthasy (nanomotor)
Přímé pozorování ATP poháněné rotace ATPsynthasy (nanomotor). Na g podjednotku je připevněno aktinové vlákno – pozorovatelné fluorescenčním mikroskopem.
. Na g podjednotku je připevněno aktinové vlákno – pozorovatelné fluorescenčním mikroskopem.")
43
Jak se dostává cytoplasmatické NADH do matrix mitochondrie ??

44
Glycerol-3-fosfátový člunek ve svalech(shuttle)
Glycerol-3-fosfátový člunek ve svalech(shuttle). Redukce dihydroxyacetonfosfátu na glycerol-3-fosfát a poté mitochondriální glycerol-3-fosfátdehydrogenasa. Tvoří se pouze 1, 5 ATP !!!
. Redukce dihydroxyacetonfosfátu na glycerol-3-fosfát a poté mitochondriální glycerol-3-fosfátdehydrogenasa. Tvoří se pouze 1, 5 ATP !!!")
45
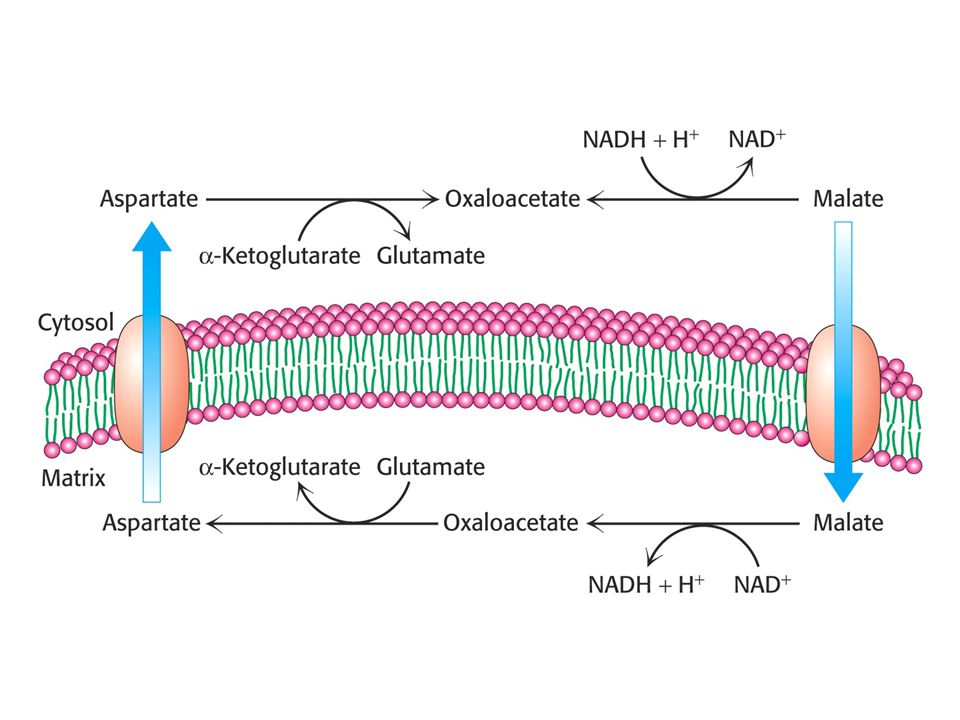
Malát – aspartátový člunek (srdce a játra)
Malát – aspartátový člunek (srdce a játra). Reversibilní – závisí na poměru NADH/NAD+ v cytoplasmě a matrix.
. Reversibilní – závisí na poměru NADH/NAD+ v cytoplasmě a matrix.")
47
Mechanismus mitochondriální ATP-ADPtranslokasy (14 % proteinů mitochondriální membrány) ANTIPORTER. ATP je transportován 30 x rychleji ven, než ADP dovnitř. Čtvrtina energie získaná ox. fosforylací jde na konto exportu ATP. Inhibice – atraktylosid (rostlinný glykosid) a bongkreková kyselina (antibiotikum z plísně).
 a bongkreková kyselina (antibiotikum z plísně)..")
48
Mitochondriální přenašeče

50
Respirační kontrola. Elektrony jsou trasportovány na O2 tehdy, když je současně ADP fosforylováno na ATP. Kontrolováno hladinou ADP.

51
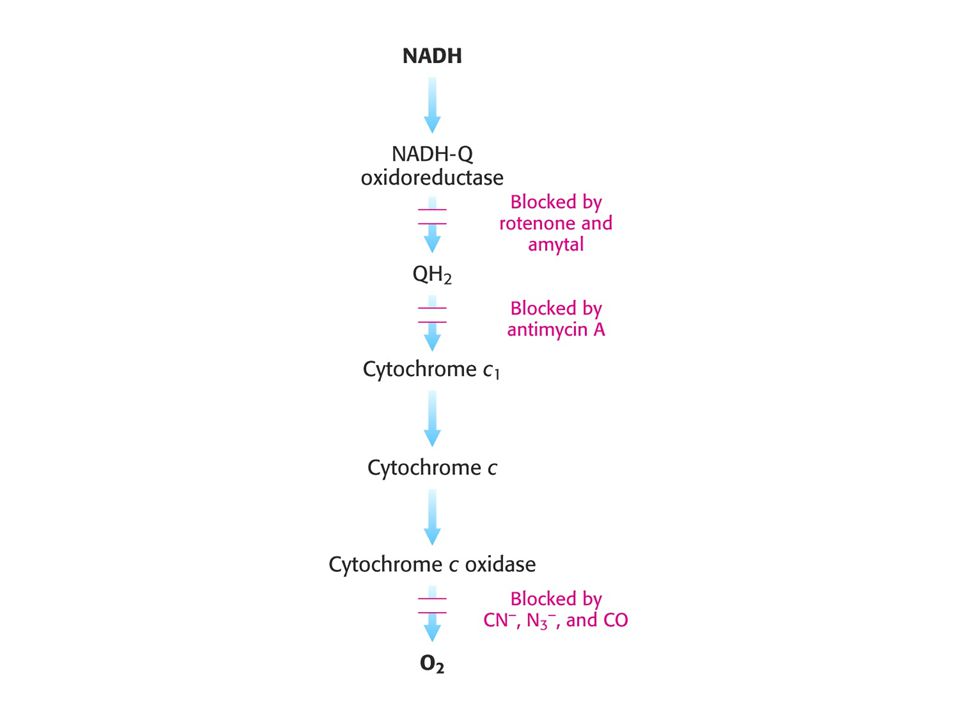
Inhibitory oxidativní fosforylace.

53
Rotenon – inhibitor mitochondriální oxidace na straně nehemového železa NADH dehydrogenasy. Insekticid

54
AMYTAL – barbiturát, inhibuje elektronový transportní řetězec mezi flavoproteiny a ubichinonem

55
Odpojovače(uncouplers) oxidatívní fosforylace
 oxidatívní fosforylace")
56
Funkce odpojovacího proteinu UCP-1 (thermogenin) – hnědá adiposní tkáň bohatá na mitochondrie (zazimující živočichové, novorozenci, zvířata adaptovaná na zimu)
 – hnědá adiposní tkáň bohatá na mitochondrie (zazimující živočichové, novorozenci, zvířata adaptovaná na zimu)")
Podobné prezentace











>")


>")








>")
