Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Úvod do obecné genetiky
2008

2
termín „genetika“ se poprvé objevuje v r. 1905
1909: pojem „gen“ 1911 termín „gen“ proniká do kontinentální Evropy

3
Johann Gregor Mendel 1822-1884 narozen 22. června 1822
v Hynčicích ve Slezsku 1843 vstup do augustiniánského kláštera na Starém Brně 1851 – 1853 studia ve Vídni 1856 počátek experimentů s hrachem, celkem okolo rostlin 1865 Versuche über Pflanzen-Hybride 1868 zvolen opatem 6. ledna 1884 umírá 1900 Mendelova práce znovuobjevena: Hugo de Vries (Nizozemí), Erich von Tschermak (Rakousko), Carl Correns (Německo)
, Erich von Tschermak (Rakousko), Carl Correns (Německo)")
4
Genetické křížení J.G. Mendel 1862 J.G. Mendel 1880

5
zatímco Mendelovi předchůdci se snažili zkoumat velký počet znaků u jedněch rodičů a zkoumali, jak se v potomstvu mísí… Mendel naopak zkoumal velmi malý počet znaků na velké populaci potomstva

6
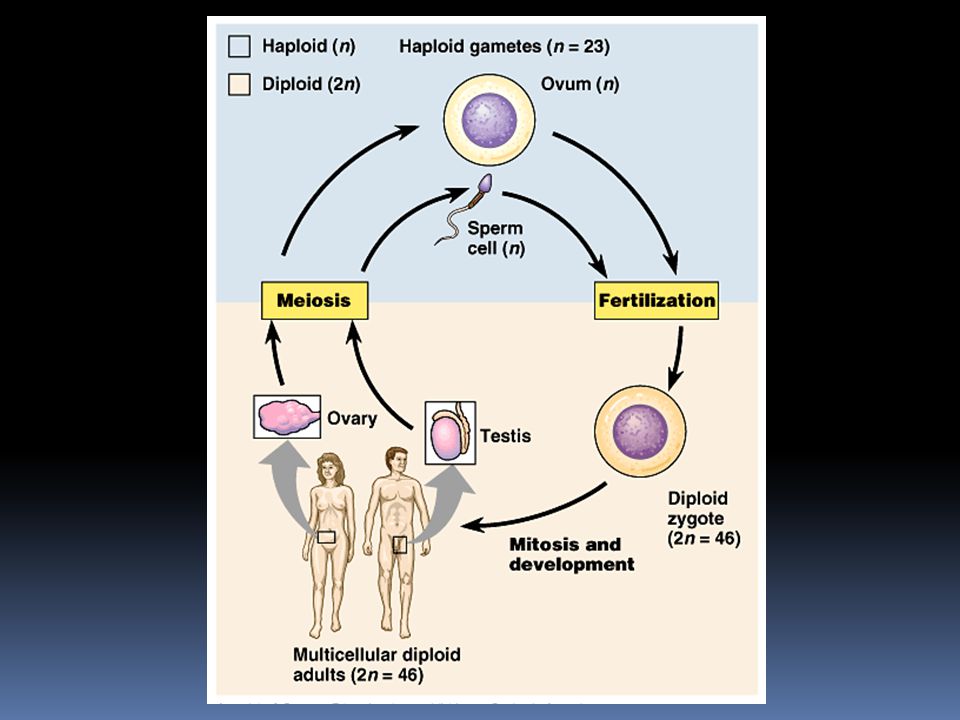
Důležité termíny každá „dědičná vlastnost“ je u diploidního organismu řízena dvěma geny; jeden jsme zdědili od otce, druhý od matky výjimkou jsou u muže geny na chromozómu X a na chromozómu Y. (Pozn. genů na chromozómu Y je velmi málo)
")
7
Důležité termíny Alely = různé varianty téhož genu. Pro jeden gen máme v našem těle dvě alely; jednu jsme zdědili od otce, druhou od matky. Hovoříme např. o genu pro barvu květů, a o alele způsobující fialovou barvu a o alele způsobující bílou barvu květu. To ale neznamená, že v populaci jsou vždy jen dvě alely. Známe např. tři alely pro dědičnost krevních skupin, IA, IB, i. V konkrétním člověku jsou vždy pouze dvě z nich. V populaci kolují všechny tři.

8
Alely lokus = konkrétní místo na chromozómu, na kterém se nachází určitý gen

9
Homologní chromosomy Žluté tečky označují jistý gen – a jeho lokus – na homologních chromosomech. Tyto chromosomy jsou již zreplikované po S fázi – což je poznat z toho, že na každém z obou homologních chromosomů jsou žluté tečky dvě.

10
Důležité termíny genom = kompletní genetický materiál daného organismu
genotyp = soubor alel, které má organismus k dispozici fenotyp = fyzické a fyziologické rysy daného organismu (=to, jak organismus aktuálně vypadá) čistá linie = rostliny, které při samoopylení vykazují opakovaně stejnou variantu sledovaného znaku (jsou homozygotní)
 čistá linie = rostliny, které při samoopylení vykazují opakovaně stejnou variantu sledovaného znaku (jsou homozygotní)")
11
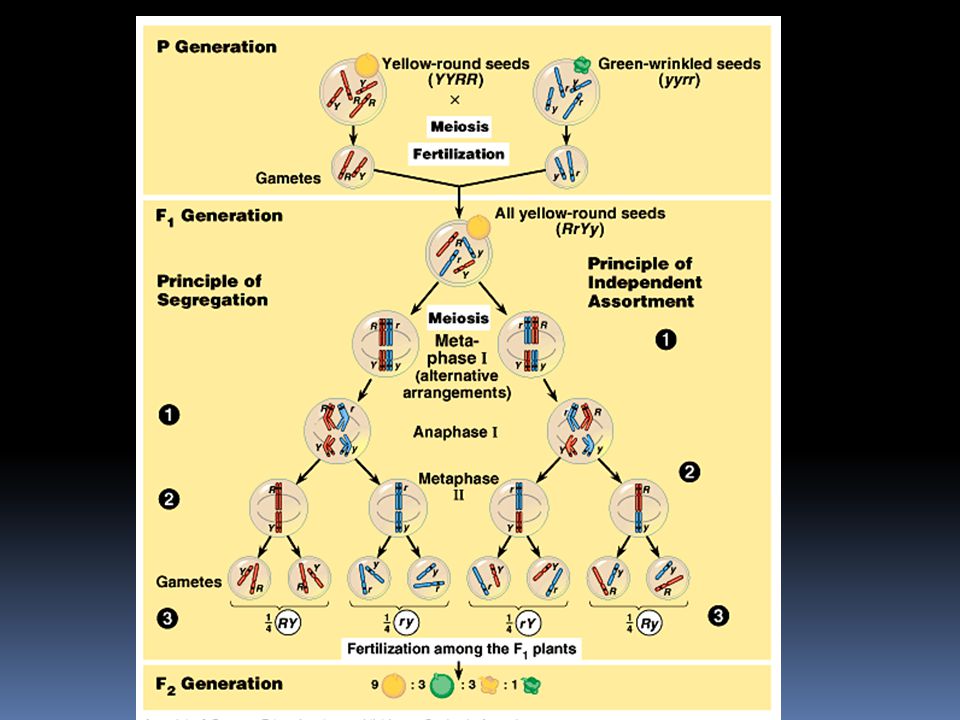
Základní Mendelův pokus
P generace = parentální, rodičovská F1 generace = první filiální generace (filius = syn) F2 generace = druhá filiální generace
 F2 generace = druhá filiální generace.")
12
Mendelovy úvahy dědičný faktor pro bílou barvu květů v F1 generaci nezmizel, jen ustoupil znaku pro fialovou barvu. fialová barva květu je tedy dominantní znak bílá barva květu je recesívní znak bílá barva květu se znovu objevuje u F2 generace. Znak pro bílou barvu tedy nebyl v F1 generaci ztracen, jen koexistoval spolu se znakem pro fialovou barvu

13
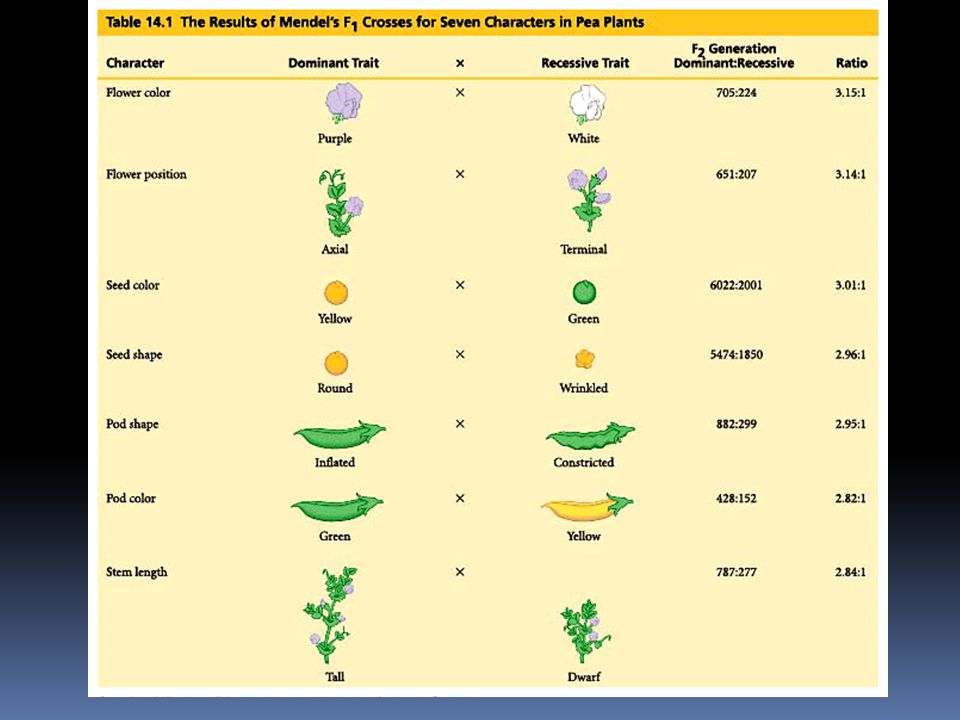
Mendel zkoumal u hrachu celkem 7 znaků
barva květů fialová x bílá 705:224 3,15:1 pozice květů axiální terminální 651:207 3,14:1 barva semen žlutá zelená 6022:2001 3,01:1 tvar semen kulatý svraštělý 5474:1850 2,96:1 tvar lusku nafouklý obtažený 882:299 2,95:1 barva lusků 428:152 2,82:1 výška stonku vysoký nízký 787:277 2,84:1

15
„Mendelovy zákony“ (Mendel žádné zákony nikdy neformuloval)
Alternativní verze genů (odlišné alely) odpovídají za rozdílnosti ve zděděných rysech gen pro barvu květů existuje ve dvou verzích, jeden odpovídá za fialovou barvu, druhý za bílou barvu. tyto alternativní verze genu se nazývají alely
 odpovídají za rozdílnosti ve zděděných rysech. gen pro barvu květů existuje ve dvou verzích, jeden odpovídá za fialovou barvu, druhý za bílou barvu. tyto alternativní verze genu se nazývají alely.")
16
„Mendelovy zákony“ (Mendel žádné zákony nikdy neformuloval)
pro každou „dědičnou vlastnost“ má organismus dvě alely, jednu získanou od otce, druhou od matky diploidní organismus má páry homologních chromozómů, jeden chromozóm z každého páru pochází od otce, druhý od matky. Genový lokus je tak v diploidní buňce přítomen dvakrát. Tyto homologní loci mohou obsahovat stejné alely, jako např. u čistých linií P generace hrachu z Mendelova pokusu, nebo se mohou lišit, jako v F1 generaci z tohoto pokusu

18
„Mendelovy zákony“ (Mendel žádné zákony nikdy neformuloval)
Pokud se obě alely liší, pak jedna, zvaná dominantní alela, se plně vyjádří ve vzhledu organismu, druhá, zvaná recesívní alela, nebude mít žádný pozorovatelný efekt na vzhledu organismu v Mendelově pokusu mají rostliny v F1 generaci fialové květy, přestože je v každé z nich krom alely pro fialovou barvu květu i alela pro bílou barvu květu

19
„Mendelovy zákony“ (Mendel žádné zákony nikdy neformuloval)
Dvě alely pro každou dědičnou vlastnost segregují (oddělují se od sebe) během vzniku gamet vajíčko i spermie obsahují pouze polovinu genetického materiálu, přítomného v somatické diploidní buňce. Homologní chromozómy se od sebe v průběhu meiózy oddělují. Pokud jsou obě alely na homologních chromozomech stejné, pak ve všech gametách se bude nacházet tatáž alela pro daný znak. Pokud se budou alely lišit, pak polovina gamet bude obsahovat dominantní alelu a polovina gamet recesívní alelu. Tento „zákon“ se nazývá „zákon o segregaci alel“
 během vzniku gamet. vajíčko i spermie obsahují pouze polovinu genetického materiálu, přítomného v somatické diploidní buňce. Homologní chromozómy se od sebe v průběhu meiózy oddělují. Pokud jsou obě alely na homologních chromozomech stejné, pak ve všech gametách se bude nacházet tatáž alela pro daný znak. Pokud se budou alely lišit, pak polovina gamet bude obsahovat dominantní alelu a polovina gamet recesívní alelu. Tento „zákon se nazývá „zákon o segregaci alel")
20
Zákon o segregaci alel Čisté linie obsahují totožné alely, PP nebo pp. Gameta obsahuje jen jednu alelu pro barvu květu, buď P nebo p. Spojením rodičovských gamet vznikne hybrid s kombinací Pp. Protože P alela je dominantní, budou mít všichni hybridi z F1 generace fialové květy. Nyní necháme hybridy z F1 zkřížit mezi sebou. V F2 generaci získáme ¾ potomků fialových a ¼ bílých.

21
Fenotypový a genotypový štěpný poměr
Fenotypový štěpný poměr pro F2 generaci je 3:1 (tři fialové : jeden bílý) Genotypový štěpný poměr pro F2 generaci je 1:2:1 (1PP:2Pp:1pp)
 Genotypový štěpný poměr pro F2 generaci je 1:2:1 (1PP:2Pp:1pp)")
22
Genotypový a fenotypový štěpný poměr v F2 generaci

23
Užitečné termíny homozygot = organismus, který má pro daný znak obě alely identické (např. PP nebo pp). Pokud má organismus obě alely dominantní pro sledovaný znak, užíváme termín dominantní homozygot (PP), pokud jsou obě alely recesívní, užíváme termín recesívní homozygot heterozygot = organismus, který má obě alely pro sledovaný znak odlišné (Pp)
. Pokud má organismus obě alely dominantní pro sledovaný znak, užíváme termín dominantní homozygot (PP), pokud jsou obě alely recesívní, užíváme termín recesívní homozygot. heterozygot = organismus, který má obě alely pro sledovaný znak odlišné (Pp)")
24
Analytické zpětné křížení (testcross)
pokud máme hrách s fialovými květy, nevíme, zda se jedná o homozygota (PP) nebo o heterozygota (Pp) pomůžeme si křížením této rostliny s bělokvětým hrachem jednalo-li se o heterozygota (Pp), potom polovina potomstva bude fialová a polovina bílá jednalo-li se o dominantního homozygota, pak bude potomstvo uniformě fialové křížení organismu s neznámým genotypem s recesívním homozygotem nazýváme analytické zpětné křížení. Je užitečné zejména u živočichů, kde nelze provést samoopylení
 nebo o heterozygota (Pp) pomůžeme si křížením této rostliny s bělokvětým hrachem. jednalo-li se o heterozygota (Pp), potom polovina potomstva bude fialová a polovina bílá. jednalo-li se o dominantního homozygota, pak bude potomstvo uniformě fialové. křížení organismu s neznámým genotypem s recesívním homozygotem nazýváme analytické zpětné křížení. Je užitečné zejména u živočichů, kde nelze provést samoopylení.")
25
Analytické zpětné křížení
= u živočichů = křížení s recesívním rodičem

26
Monohybridismus a dihybridismus
Doposud jsme sledovali pouze jediný znak - fialovou nebo bílou barvu květů. Hovoříme o monohybridismu co kdybychom si všímali zároveň dvou znaků? Mendel v jedné variantě pokusu sledoval zároveň barvu semen (žlutá x zelená) a tvar semen (kulatý x svraštělý). Hovoříme zde o dihybridismu.
 a tvar semen (kulatý x svraštělý). Hovoříme zde o dihybridismu.")
27
Dihybridismus s dominancí
Y - žlutá barva semen (yellow) y - zelená barva semen R - kulatá semena (round) r - svraštělá semena Fenotypový štěpný poměr: 9:3:3:1 Genotypový štěpný poměr: 1:2:1:2:4:2:1:2:1 tyto výsledky jsou známy jako „zákon o nezávislé kombinovatelnosti alel“ Pokud bychom zkoumali tyto znaky odděleně, dostali bychom obvyklý poměr 3:1 (resp.1:2:1)
 y - zelená barva semen. R - kulatá semena (round) r - svraštělá semena. Fenotypový štěpný poměr: 9:3:3:1. Genotypový štěpný poměr: 1:2:1:2:4:2:1:2:1. tyto výsledky jsou známy jako „zákon o nezávislé kombinovatelnosti alel Pokud bychom zkoumali tyto znaky odděleně, dostali bychom obvyklý poměr 3:1 (resp.1:2:1)")
28
Dihybridismus s dominancí

29
„Mendelovy zákony“ Zákon o uniformitě F1 (1. filiální = první generace potomků) generace. Zákon o náhodné segregaci genů do gamet. Při křížení 2 heterozygotů může být potomkovi předána každá ze dvou alel (dominantní i recesivní) se stejnou pravděpodobností. Zákon o nezávislé kombinovatelnosti alel. Při zkoumání 2 alel současně dochází k téže pravidelné segregaci. Máme-li 2 polyhybridy AaBb může každý tvořit 4 různé gamety (AB, Ab, aB, ab).
 se stejnou pravděpodobností. Zákon o nezávislé kombinovatelnosti alel. Při zkoumání 2 alel současně dochází k téže pravidelné segregaci. Máme-li 2 polyhybridy AaBb může každý tvořit 4 různé gamety (AB, Ab, aB, ab).")
30
Rozšíření Mendelových výzkumů
Mendel díky své genialitě (nebo štěstí či spíše dokonalé znalosti hrachu) si vybral 7 znaků, u nichž je kompletní dominance - fialová či bílá, nula nebo jednička každý z těchto sedmi znaků je kódován jen jedním genem, pro který existují jen dvě alely (s výjimkou postavení květu - tento znak kódují dva geny) vztah mezi genotypem a fenotypem je v přírodě málokdy tak jednoduchý...
 si vybral 7 znaků, u nichž je kompletní dominance - fialová či bílá, nula nebo jednička. každý z těchto sedmi znaků je kódován jen jedním genem, pro který existují jen dvě alely (s výjimkou postavení květu - tento znak kódují dva geny) vztah mezi genotypem a fenotypem je v přírodě málokdy tak jednoduchý...")
31
Nekompletní dominance
U tohoto typu dědičnosti vypadá hybrid z F1 jako průměr obou rodičů zde u hledíku má hybrid méně červeného pigmentu než dominantní rodič v F2 generaci je genotypový i fenotypový štěpný poměr totožný: 1:2:1

32
Neúplná dominance

33
Kodominance Zde ovlivní dvě alely heterozygota fenotyp
nejedná se ale o neúplnou dominanci, heterozygot má svou zvláštní kombinaci, kde se uplatní vedle sebe obě alely příkladem je dědičnost krevních skupin M,N a MN. Jedná se o určité molekuly na povrchu erytrocytů. Jedinci MM budou mít jeden typ molekuly, zatímco lidé NN budou mít druhý. Heterozygoti MN budou mít na povrchu erytrocytů oba typy těchto molekul MN není průměrem mezi M a N - vyjádří se jen obě alely nezávisle na sobě.

34
Problémy ve vztahu dominance-recesivita
Tay-Sachsova nemoc je autozomální dědičná choroba, postihující recesívní homozygoty. Mozkové buňky takovéhoto dítěte nejsou schopny metabolizovat jeden typ mozkových lipidů, zvaný gangliosidy, protože enzym, který tuto reakci katalyzuje nefunguje správně. Lipidy se tak hromadí v buňkách, znemožňují jim pracovat a výsledkem je smrt dítěte. Nemoc postihuje pouze děti, které zdědily obě alely pro Tay-Sachsovu nemoc.

35
Problémy ve vztahu dominance-recesivita
Na úrovni organismu se tedy alela chová jako recesívní a je možno hovořit o úplné dominanci na biochemické úrovni se však jedná a neúplnou dominanci - v buňkách heterozygotů je snížená hladina funkčního enzymu. I tato snížená hladina enzymu však „utáhne“ všechny potřebné reakce, proto jsou heterozygoti zdrávi na molekulární úrovni se ale jedná o kodominanci - v buňkách je syntetizován jak funkční, tak nefunkční varianta enzymu v poměru 1:1, vyjadřují se tak obě alely.

36
Problémy ve vztahu dominance-recesivita
Dominance rovněž neznamená, že by dominantní alela nějak přímo potlačovala recesívní alelu; rozdíly mezi oběma alelami jsou v nukleotidových sekvencích daného genu v příkladu semen hrachu kulatá x svraštělá je kulatost dominantní nad svraštělostí dominantní alela kóduje enzym, který je schopen přeměnit cukr na škrob; recesívní alela kóduje nefunkční formu tohoto enzymu

37
Problémy ve vztahu dominance-recesivita
U recesívního homozygota se tedy cukr nemůže přeměnit na škrob. Jak se semeno vyvíjí, vysoká koncentrace cukru způsobuje, že osmotickými silami do semena vniká voda. Když je semeno dospělé, vysouší se a svraskává je-li přítomna dominantní alela, cukr je proměněn na škrob a dospělé semeno zůstává kulaté. Jedna varianta zdravé alely stačí, aby se cukr přeměnil na škrob, heterozygoti jsou proto rovněž kulatí

38
Problémy ve vztahu dominance-recesivita
Vztah dominace - recesivita rovněž neznamená, že by dominantích alel bylo v populaci nutně více v USA se rodí jedno dítě z 400, které má navíc prst u nohy či ruky tato alela pro polydaktylii je dominantní. 399 z 400 dětí jsou proto recesívní homozygoti

39
Problémy ve vztahu dominance-recesivita shrnutí
Vztah dominance-recesivita: 1. jde od úplné dominance přes různé formy neúplné dominace až ke kodominanci 2. Vyjadřuje mechanismus, jakým jsou specifické alely vyjádřeny ve fenotypu, nikoli to, že by se alely potlačovaly na úrovni DNA 3. Nemá nic společného s frekvencí výskytu alel v populaci

40
Možné porušení Mendelových zákonů? Do popředí se dostává mRNA
Gen Kit způsobuje u černých myší skvrnité ocásky a bílé tlapky Dominantní homozygoti AA jsou celí černí Recesívní homozygoti aa umírají Heterozygoti Aa jsou skvrnití

41
Možné porušení Mendelových zákonů?
Pokud zkřížíme dva heterozygoty Aa a vznikne kombinace AA, mělo by teoreticky jít o zcela černou myš Myš je ale skvrnitá Jediné možné vysvětlení je, že by spolu s dominantní alelou šla do gamety i již připravená mRNA získaná přepsáním recesivní alely Možná jsme na stopě fundamentálnímu pravidlu, které je možná v živé přírodě včetně člověka značně rozšířeno

42
Vícealelové systémy Většina genů je v populaci zastoupena ve více než dvou alelách příkladem jsou alely pro dědičnost krevních skupin IA, IB a i alely IA a IB odpovídají za přítomnost dvou cukrů - A substance a B substance na povrchu erytrocytů člověk může mít na erytrocytech přítomnu jednu z nich (A nebo B), obě (AB), nebo žádnou (O)
, obě (AB), nebo žádnou (O)")
43
Dědičnost krevních skupin
Existují tedy 4 typy fenotypů, A,B,AB,O protože každá osoba má dvě alely, je možných 6 genotypů (IAIA, IB IB,IAIB ,IAi, IB i, ii) recesívní homozygoti mají skupinu 0, protože žádná substance není na povrchu erytrocytů přítomna IA a IB jsou k sobě kodominantní IA a IB jsou ve vztahu k alele i dominantní
 recesívní homozygoti mají skupinu 0, protože žádná substance není na povrchu erytrocytů přítomna. IA a IB jsou k sobě kodominantní. IA a IB jsou ve vztahu k alele i dominantní.")
44
Dědičnost krevních skupin

45
Dědičnost krevních skupin
Znalost krevních skupin je kritickým faktorem pro transfúzi krve. Imunitní systém příjemce reaguje na jakoukoli cizorodou látku pokud má krev donora substanci, která je příjemci cizí, nastane imunitní reakce vedoucí ke krevním sraženinám ústící ve smrt příjemce

46
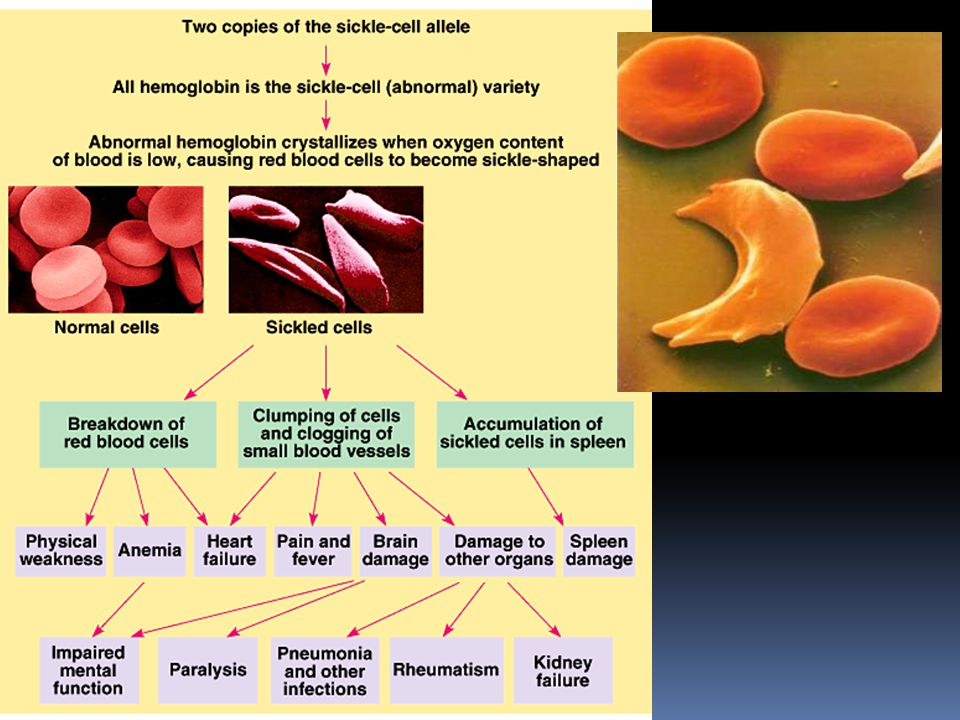
Pleiotropie Doposud jsme se zabývali případy, kdy jeden gen má jeden účinek na fenotyp organismu. Většina genů se ovšem projeví na fenotypu vícero způsoby jev, kdy jeden gen ovlivní organismus mnoha způsoby se nazývá pleiotropie typickým případem u člověka je gen způsobující srpkovitou anémii

47
Srpkovitá anémie – případ pleiotropie

48
Sickle cell disease The disease results from an altered hemoglobin, in which the aminoacid valin substitutes for glutamic acid at position 6 in the β- globin chain.

51
Epistáze = gen na jednom lokusu ovlivní expresi jiného genu na jiném lokusu (epistáze = řecky „zastavení“) Příklad: u myší je černá barva srsti dominantní nad hnědou barvou srsti B … alela pro černou barvu b …. alela pro hnědou barvu jiný gen, zvaný epistatický, rozhoduje o tom, zda barvivo bude „umístěno“ do srsti. Nazvěme tento gen písmenem C. bude-li přítomna alela C, pak bude srst podle situace hnědá či černá bude-li mít myš genotyp cc, pak se neuloží do srsti žádné barvivo a myš bude bílá

52
Epistáze Fenotypový štěpný poměr je 9:3:4

53
Komplementace 9:7 Fenotypový štěpný poměr je
Hrachor (Lathyrus odoratus). Křížením dvou bělokvětých rostlin vznikají červenokvěté i bělokvěté rostliny. Pro tvorbu červeného anthokyanu je nutná přítomnost jak alely C, tak alely R. Alela C řídí tvorbu enzymu, kterým vzniká bezbarvý prekurzor anthokyanu, alela R řídí tvorbu enzymu, který mění tento prekurzor na anthokyan.
. Křížením dvou bělokvětých rostlin vznikají červenokvěté i bělokvěté rostliny. Pro tvorbu červeného anthokyanu je nutná přítomnost jak alely C, tak alely R. Alela C řídí tvorbu enzymu, kterým vzniká bezbarvý prekurzor anthokyanu, alela R řídí tvorbu enzymu, který mění tento prekurzor na anthokyan.")
54
Polygenní dědičnost mnoho znaků, jako je např. tělesná výška člověka, odstín barvy kůže, inteligence apod. není v populaci diskrétních (fialové květy x bílé), ale plynule se mění hovoříme o dědičnosti kvantitativních znaků polygenní dědičnost je jev, kdy mnoho genů ovlivňuje jeden znak (jedná se o opak pleiotropie, kdy jeden gen ovlivňuje mnoho znaků)
, ale plynule se mění. hovoříme o dědičnosti kvantitativních znaků. polygenní dědičnost je jev, kdy mnoho genů ovlivňuje jeden znak (jedná se o opak pleiotropie, kdy jeden gen ovlivňuje mnoho znaků)")
55
Polygenní dědičnost odstín barvy kůže u člověka
řekněme, že u člověka je odstín barvy kůže kódován minimálně třemi geny (ve skutečnosti více) nazvěme tyto geny A,B,C každé velké písmeno tak přidává výslednému znaku jednu „jednotku“ osoba AABBCC bude mít velmi tmavou kůži, zatímco osoba aabbcc bude mít velmi světlou kůži protože se jedná o kumulativní efekt, osoby AaBbCc a AABbcc budou mít tutéž barvu kůže
 nazvěme tyto geny A,B,C. každé velké písmeno tak přidává výslednému znaku jednu „jednotku osoba AABBCC bude mít velmi tmavou kůži, zatímco osoba aabbcc bude mít velmi světlou kůži. protože se jedná o kumulativní efekt, osoby AaBbCc a AABbcc budou mít tutéž barvu kůže.")
56
Polygenní dědičnost odstín barvy kůže u člověka
Výsledná křivka je typická tzv. Gaussova křivka. Její konečný spojitý tvar je ovlivněn environmentálními faktory, jako je slunění apod.

57
Gaussova křivka

58
Gaussova křivka

59
Nature versus Nurture environmentální dopad na fenotyp
u člověka ovlivňuje výživa tělesnou výšku, opalování barvu kůže, posilování svalovou hmotu atd. i u jednovaječných dvojčat existují rozdíly díky odlišnému vlivu prostředí bonsaje, větší výnosy na pohnojených polích genotyp tedy není něco pevně daného, spíše se jedná o mantinely.

60
Nature versus Nurture environmentální dopad na fenotyp
inteligence = geny + trénink, studium barva kůže = geny + opalování tělesná výška = geny + výživa alkoholismus = geny + prostředí filosofie – u člověka je třeba ještě započítat vliv svobody

61
Nature versus Nurture environmentální dopad na fenotyp
norma reakce = mnoho různých fenotypů, které mohou vzniknout z jediného genotypu genotyp je klaviatura, na kterou může prostředí zahrát mnoho různých skladeb schizofrenie = geneticky podmíněná, ale zpravidla je pro vypuknutí zapotřebí environmentální stimul, stres (příprava na maturitu apod.)
")
62
Nature versus Nurture environmentální dopad na fenotyp
v některých případech prostředí nehraje žádnou roli a norma reakce je nulová: dědičnost krevních skupin, fialová/bílá barva květu hrachu, srpkovitá anémie atd. norma reakce bývá nejširší pro polygenní znaky, jak jsme viděli u barvy kůže. tyto znaky nazýváme multifaktoriální, neboť za ně odpovídá mnoho faktorů, a to genetických i faktory prostředí

63
Nature versus Nurture environmentální dopad na fenotyp
Norma reakce. Barva květu záleží u této rostliny na kyselosti půdy

64
Nature versus Nurture environmentální dopad na fenotyp
Plicník lékařský (Pulmonaria officinalis) má květy nejprve červené, později modré, při odkvétání až do fialova
 má květy nejprve červené, později modré, při odkvétání až do fialova.")
65
Mendelovská dědičnost u lidí
Nevýhoda člověka příliš dlouhá generační doba (stejná jako u výzkumníka) málo dětí bylo by neetické jej křížit provádí se analýza rodokmenů Příklad: alela W způsobuje „widow´s peak“, alela F volný ušní lalůček
 málo dětí. bylo by neetické jej křížit. provádí se analýza rodokmenů. Příklad: alela W způsobuje „widow´s peak , alela F volný ušní lalůček.")
66
Analýza rodokmenů

67
Recesívně děděné choroby
mnohem častější než dominantně děděné choroby heterozygoti jsou zdrávi, jsou zváni přenašeči postižený člověk se většinou narodí zdravým rodičům (Aa x Aa) u člověka od velmi lehkých (albinismus) po závažné (cystická fibrosa, fenylketonurie, Tay-Sachsova nemoc, srpkovitá anémie, různé thalassemie atd.) jsou v lidstvu nerovnoměrně rozloženy (Tay- Sachsova nemoc – aškenázští židé v USA, srpkovitá anémie – Afroameričané)
 u člověka od velmi lehkých (albinismus) po závažné (cystická fibrosa, fenylketonurie, Tay-Sachsova nemoc, srpkovitá anémie, různé thalassemie atd.) jsou v lidstvu nerovnoměrně rozloženy (Tay- Sachsova nemoc – aškenázští židé v USA, srpkovitá anémie – Afroameričané)")
68
Recesívně děděné choroby
každý z nás jsme heterozygotní pro asi 30 genetických chorob odtud zřejmě ve všech známých kulturách zákaz incestu a sňatků uvnitř rodiny tyto tabu se možná zrodily z empirických pozorování dnes známo asi genetických chorob

69
Cystická fibrosa nejčastější dědičná choroba v USA
1: 2500 bílých obyvatel každý 25 člověk je přenašeč (4%) zdravá alela kóduje membránový přenašeč pro chloridové ionty recesívní homozygoti mají tyto kanály nefunkční, výsledkem je mimořádně vysoká koncentrace extracelulárního Cl- výsledkem je hlen přítomen v pankreatu, plicích, trávicí soustavě a dalších orgánech.
 zdravá alela kóduje membránový přenašeč pro chloridové ionty. recesívní homozygoti mají tyto kanály nefunkční, výsledkem je mimořádně vysoká koncentrace extracelulárního Cl- výsledkem je hlen přítomen v pankreatu, plicích, trávicí soustavě a dalších orgánech.")
70
Cystická fibrosa baktérie i buňky imunitního systému jsou v tomto hlenu zachyceny, což dále zhoršuje stav pacienta neléčena způsobuje smrt před dosažením pátého roku věku při léčbě a denních dávkách antibiotik přežívá polovina pacientů 20 let.

71
Tay-Sachsova nemoc je způsobena nefunkčním enzymem, který není schopen rozložit jisté lipidy v mozku symptomy obvykle několik měsíců po narození; dítě má bolestivé křeče, slepne, nastává degenerace motorických i mentálních schopností smrt nastává do několika let po narození u aškenázů – židů – (cca Německo) je poměr 1:3600, což je 100x víc než u sefardských židů (cca Španělsko)
 je poměr 1:3600, což je 100x víc než u sefardských židů (cca Španělsko)")
72
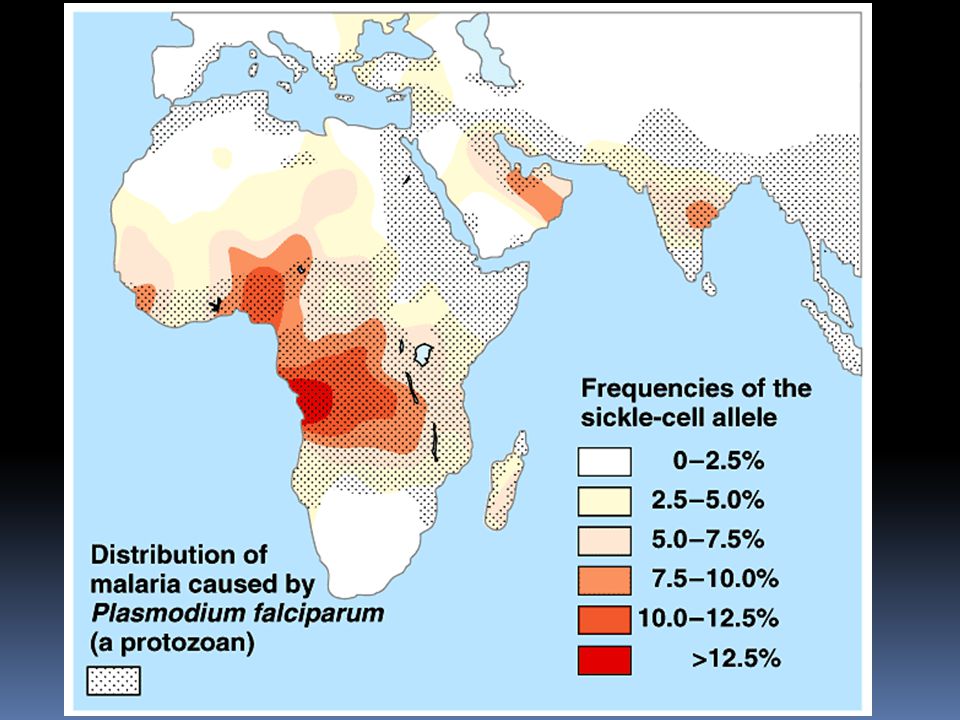
Srpkovitá anémie 1:400 u Afroameričanů
každý desátý Afroameričan je přenašeč u nemocných jsou třeba pravidelné transfúze, léčba zatím neexistuje heterozygoti jsou zdrávi, ale pokud se vyskytují delší dobu v místech s menším parciálním tlakem kyslíku, mohou mít problémy balancovaný polymorfismus: dominantní homozygot zemře na malárii recesívní homozygot zemře na anémii heterozygot přežije

73
Fenylketonurie (PKU) 1:10 000 až 1:15 000
neschopnost odbourat fenylalanin neléčena způsobuje u dětí mentální retardace (někdy je IQ kolem 50, jindy téměř 100) léčba neexistuje, jen dieta bez fenylalaninu
 léčba neexistuje, jen dieta bez fenylalaninu.")
74
Dominantně děděné choroby
mnohem vzácnější – neexistují přenašeči, kdo škodlivou alelu má, je nemocný achondroplasie, familiální hypercholesterolemie, Huntingtonova nemoc

75
Achondroplasie = trpasličí vzrůst výskyt 1:10000…
… z čehož vyplývá že 99,9% populace jsou recesívní homozygoti

76
Achondroplasie

77
Huntington´s disease auosomálně dominantní dědičnost

78
Huntingtonova choroba
= degenerativní neuropsychiatrická nemoc, zasahující basální ganglia, ovšem postihující rovněž široké oblasti celého nervového systému dědičnost je autosomálně dominantní: P: Aa x aa F1: Aa Aa aa aa 1 : 1 = každé dítě, které má jednoho postiženého rodiče má 50% risk, že je nositelem HD alely

79
Huntingtonova choroba
Symptomy se objevují obvykle mezi 35 a 50 rokem života... ...ačkoli nemoc může propuknout kdykoli mezi dětstvím a pokročilým věkem smrt nastává v průměru 15 let po objevení se prvních příznaků jedná se o tzv. late-onset disease, symptomy se objevují obvykle ve věku, kdy již postižená osoba má vlastní děti

80
Huntingtonova choroba
byl objeven gen způsobující Huntingtonovu chorobu. Nachází se na p raménku chromozómu 4. gen byl isolován gen je defektní kvůli abnormálně velkému počtu repeticí CAG jakou funkci v organismu má zdravý gen, není známo

81
Huntington´s Disease Objev HD genu v roce 1993 vyústil v dostupnost přímého genetického testování pro osoby v riziku choroby nebo pro osoby již nemocné pro potvrzení diagnózy. Z krevního vzorku se analyzuje DNA a zjistí se počet repetic CAG v HD lokusu terapie v současnosti neexistuje žádost o provedení testu je většinou založena na úmyslu založit rodinu a plánování rodičovství či na plánu pro další vlastní život.

82
Huntington´s Disease Zdraví jedinci mají obvykle 28 nebo méně repeticí CAG jedinci postižení Huntingtovou chorobou mají obvykle 40 nebo víc repeticí CAG malé procento osob má však počet repeticí mezi 30 až 40, tedy v zóně, kdy nelze přesně určit, zda nemoc propukne či ne. Nestabilní trinukleotidová repetice se může prodloužit při přenosu z rodiče na dítě

83
Huntingtonova choroba
Note: Dr. Michael McCormack cituje případ, kdy měla pacientka o jednu repetici více než její matka. Pacientka byla symptomatická již ve 40 letech, zatímco její matka byla v 70 roce věku ještě bez symptomů. Po nástupu nemoci se u někoho stav rapidně zhoršuje, zatímco jindy nemoc progreduje pomaleji.

84
Huntingtonova choroba klinické rysy
Emoční, kognitivní a motorické poruchy nejčastější emoční poruchy: deprese, iritabilita, apatie, výbuchy agrese, impulzivnost, sociální odtažitost kognitivní poruchy: ztráta rychlosti a pružnosti uvažování. Na pracovišti pacient není schopen udržet tempo práce.

85
Huntington´s Disease clinical features
Motorické poruchy: vědomé a nevědomé, involuntární pohyby, zejména chorea. Pacienti mají zprvu zhoršenou manuální zručnost, špatně artikulovanou řeč, potíže s polykáním, později se stávají rigidní a neschopní vykonat jakýkoli voluntární pohyb. Pacienti, kteří se dožijí tohoto stadia jsou upoutáni na lůžko a nejsou schopni se podílet na péči o sebe

86
Huntington´s Disease Genetický test odhalí přítomnost HD alely, ale nikoli nástup nemoci USA: v současnosti asi nemocných; dalších jsou v riziku onemocnění 1: je nemocných

87
Huntington´s Disease Varlam Shalamov ( )
")
88
Huntingtonova choroba etické problémy
Nancy Wexler

89
Huntington´s Disease ethical issues
Informational self – determination právo vědět právo nevědět v případě HD jen asi 10% osob chce vědět...

90
Testování heterozygotů
dnes možno u stále většího počtu chorob cystický fibrosa srpkovitá anémie Tay – Sachsova nemoc etické a sociální dopady: pojišťovny a zaměstnavatelé (přenašeči jsou naprosto zdrávi)
")
91
Fetální testování amniocentéza chorionické klky
14 – 16 týden těhotenství pro karyotyp ale buňky musíme nechat několik týdnů růst cca 1% riziko krvácení či potratu chorionické klky 8 – 10 týden těhotenství buňky rostou rychle dnes buňky plodu z krve matky - neinvazívní ultrazvuk – neinvazívní fetoskopie - neinvazívní

92
Amniocentéza a testování chorionických klků

93
Testování novorozenců
běžně na fenylketonurii (PKU)
")
94
Spojení mendelismu a chování chromosomů
se zlepšováním parametrů mikroskopů byla objevena mitóza 1875 meióza 1890 v roce 1900 v souvislosti se znovuobjevením Mendelových zákonů se začaly tyto zákony dávat do souvislosti s mitózou a meiózou kolem roku 1902 vzniká chromosomální teorie dědičnosti (Walter Sutton a Theodor Boveri)
")
95
Chromozomální teorie dědičnosti
= snaha spojit cytologii s Mendelovou dědičností

96
Thomas Hunt Morgan užívá Drosophila melanogaster
mnoho potomků (cca stovky) laciný chov krátká generační doba (14 dní) relativně snadné pozorování (lupa) jen 4 páry chromozómů (2n = 8) Morgan po roce experimentování získal drosophilu s bílýma očima
 laciný chov. krátká generační doba (14 dní) relativně snadné pozorování (lupa) jen 4 páry chromozómů (2n = 8) Morgan po roce experimentování získal drosophilu s bílýma očima.")
97
Drosophila melanogaster
Sameček Samička

98
Drosophila melanogaster

99
Drosophila melanogaster
3 páry autozomů a jeden pár gonozomů chromosomové určení pohlaví stejné jako člověk: sameček XY samička XX

100
Thomas Hunt Morgan (1866 – 1945) „To breed or not to breed. That´s the question.“
 „To breed or not to breed. That´s the question.")
101
Dědičnost vázaná na pohlaví klasický Morganův pokus
Drosophila má totéž chromosomální určení pohlaví jako člověk: XY … sameček XX … samička Na první pohled probíhá pokus přesně podle Mendela – v F2 máme poměr 3:1 Překvapivé ovšem je, že bílé oči mají vždy pouze samečci!

102
Morganovy závěry gen způsobující bílou barvu očí musí mít lokus na X chromosomu protože alela pro bílou barvu očí je recesívní, budou mít samičky bílé oči pouze v recesívně homozygotní konstelaci sameček má pouze jeden chromosom X, tedy stačí, aby právě tato jedna (a jediná) alela kódovala bílou barvu
 alela kódovala bílou barvu.")
103
Vazba genů genů je v organismu mnohem víc než chromosomů – na jednom chromosomu je tedy velké množství genů jsou-li lokusy dvou genů těsně vedle sebe na jednom chromosomu, je malá pravděpodobnost, že se crossing-over „trefí“ právě mezi ně výsledkem je fakt, že se tyto geny dostanou do gamety zpravidla spolu

104
Vazba genů v jedné variantě pokusu křížil Morgan mezi sebou drosophily s černým/normálním tělem a se zakrnělými/normálními křídly b+ šedé, normální tělo b černé tělo (b = black) vg+ normální křídla vg zakrnělá křídla (vg = vestigial)
 vg+ normální křídla. vg zakrnělá křídla (vg = vestigial)")
105
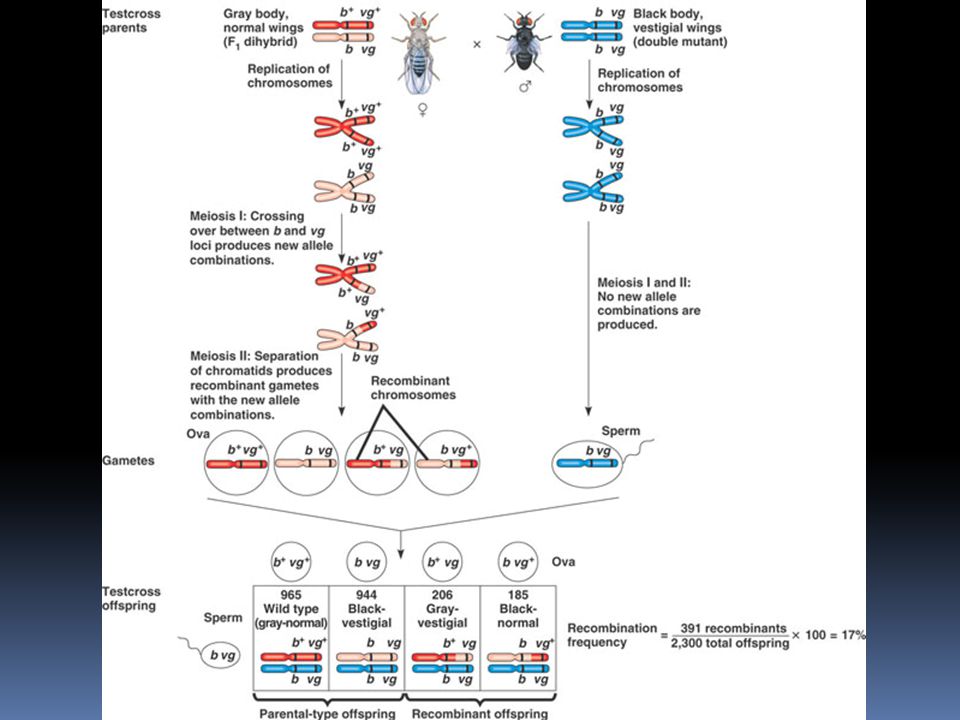
Vazba genů Morgan křížil divoké, normální drosophily b+ b+ vg+ vg+ s černými drosophilami se zakrnělýmí křídly bbvgvg b+ b+ vg+ vg+ x bbvgvg F1: b+ b vg+ vg - vzhledově normální, divoká drosophila (šedé tělo, normální křídla) nyní provedl Morgan klasické zpětné křížení: samičky z F1 zkřížil se samečky z P generace bbvgvg
 nyní provedl Morgan klasické zpětné křížení: samičky z F1 zkřížil se samečky z P generace bbvgvg.")
106
Vazba genů z tohoto křížení (b+ b vg+ vg x bbvgvg) by mělo podle Mendelových zákonů vzniknout potomstvo v poměru 1:1:1:1 tedy: ¼ šedých s normálními křídly ¼ černých se zakrnělými křídly ¼ šedých se zakrnělými křídly ¼ černých s normálními křídly avšak…

107
Vazba genů

108
Vazba genů křížení b+ b vg+ vg x bbvgvg
skutečné výsledky byly zřetelně odlišné: bylo velké množství divokých typů (šedé/normální křídla) a dvojitých mutantů (černí/zakrnělá křídla)
 a. dvojitých mutantů (černí/zakrnělá křídla)")
109
Vazba genů Morgan uzavřel, že gen pro barvu těla a gen pro zakrnělost křídel musí ležet na stejném chromosomu a při crossing-overu jdou většinou společně… …ovšem jen většinou - rekombinantní fenotypy přece jenom vznikly

110
Nezávislá segregace chromosomů a crossing-over produkují genetické rekombinanty
kdybychom křížili v podobném pokusu, jaký učinil Morgan žlutý a kulatý hrášek s genotypem YyRr se zeleným a svraštělým yyrr YyRr x yyrr vyšel by nám skutečný poměr ve shodě s předpovězeným 1:1:1:1 hrášky žluté a kulaté spolu se zelenými svraštělými bychom označili jako parentální typ a hrášky žluté a svraštělé spolu se zelenými kulatými bychom označili jako rekombinanty

111
Nezávislá segregace chromosomů a crossing-over produkují genetické rekombinanty
protože by v našem pokusu byla polovina (50%) parentálních typů a polovina (50%) rekombinantů, hovořili bychom o 50% frekvenci rekombinace celá situace je způsobena tím, že gen pro žlutá/zelená semena a gen pro kulatá/svraštělá semena leží na odlišných chromosomech což má za následek fakt, že při metafázi meiosy I se chromosomy s těmito geny dostávají zcela náhodně do dceřinných buněk
 parentálních typů a polovina (50%) rekombinantů, hovořili bychom o 50% frekvenci rekombinace. celá situace je způsobena tím, že gen pro žlutá/zelená semena a gen pro kulatá/svraštělá semena leží na odlišných chromosomech. což má za následek fakt, že při metafázi meiosy I se chromosomy s těmito geny dostávají zcela náhodně do dceřinných buněk.")
113
Vazba genů geny na jednom chromosomu ovšem nesegregují zcela náhodně – čím jsou blíže u sebe, tím je menší pravděpodobnost, že se mezi ně „trefí“ crossing-over

114
Vazba genů kdyby ležely geny v Morganově pokusu opravdu těsně vedle sebe, vznikla by kombinace 1:1:0:0 v Morganově pokusu bylo přibližně 17% rekombinantů – ačkoli zde vazba je, není zcela kompletní frekvence rekombinace = Rekombinanti/všichni x 100

116
Vazba genů

117
Z rekombinačních frekvencí lze zhotovit genetickou mapu
genetická mapa = lineární sekvence genů na chromosomu (Alfred Sturtevant) Sturtevant si uvědomil, že frekvence rekombinace – v našem případě 17% - vypovídá cosi o vzdálenosti mezi dvěma geny na chromosomu předpokládal, že crossing-overy se trefují se stejnou pravděpodobností do všech míst chromosomu… … čím jsou gny na chromosomu od sebe dál, tím je crossing-over mezi nimi pravděpodobnější
 Sturtevant si uvědomil, že frekvence rekombinace – v našem případě 17% - vypovídá cosi o vzdálenosti mezi dvěma geny na chromosomu. předpokládal, že crossing-overy se trefují se stejnou pravděpodobností do všech míst chromosomu… … čím jsou gny na chromosomu od sebe dál, tím je crossing-over mezi nimi pravděpodobnější.")
118
Z rekombinačních frekvencí lze zhotovit genetickou mapu
A.Sturtevant zkoumal tři geny: nám známý b, rovněž známý vg a gen cn (cinnabar = rumělka) drosophila s genem cn má světlejší oči než divoký typ Výsledky: cn a b = 9% cn a vg = 9,5% b a vg = 17%
 drosophila s genem cn má světlejší oči než divoký typ. Výsledky: cn a b = 9% cn a vg = 9,5% b a vg = 17%")
119
Z rekombinačních frekvencí lze zhotovit genetickou mapu
jediná možnost je umístit gen cn mezi geny b a vg 1% = 1 centimorgan

120
Genetická mapa některé geny jsou na jednom chromosomu od sebe natolik vzdáleny, že efekt vazby mezi nimi není pozorovatelný takové geny mají maximální hodnotu – 50% - frekvence rekombinace (=volná vazba) např. v Mendlových pokusech je barva semene a barva květu na stejném chromosomu 1, ale tak daleko od sebe, že se jedná o volnou vazbu pouze pro jednu dvojici ze sedmi Mendelových znaků objevili genetici vazbu: výška rostliny a tvar lusku: pro tuto dvojici Mendel dihybridní F2 nedělal (F1 ano)
 např. v Mendlových pokusech je barva semene a barva květu na stejném chromosomu 1, ale tak daleko od sebe, že se jedná o volnou vazbu. pouze pro jednu dvojici ze sedmi Mendelových znaků objevili genetici vazbu: výška rostliny a tvar lusku: pro tuto dvojici Mendel dihybridní F2 nedělal (F1 ano)")
121
Genetická mapa u Drosophily byly zjištěny celkem 4 skupiny genů tvořících vazbu, což odpovídá čtyřem chromosomům drosophily crossing overy se ale „netrefují“ do všech míst na chromosomu se stejnou frekvencí… …genetická mapa tak sice ukazuje seřazení chromosomů, ale není schopna stanovit abslotní vzdálenost mezi chromosomy

122
Genetická mapa i když vzdálenost mezi dvěma geny není natolik velká, aby je „rozpojila“ a vznikla volná vazba, přece může být natolik značná, že vznikne pravděpodobnost nikoli jen jednoho, ale dvou crossing-overů mezi oběma lokusy druhý crossing-over (d.c.o.) tak zruší účinek prvního mnohonásobný crossing-over odpovídá za matematické nepřesnosti na obrázku:
 tak zruší účinek prvního. mnohonásobný. crossing-over odpovídá. za matematické. nepřesnosti na obrázku:")
123
Genetická mapa A. Sturtevant objevil celkem čtyři vazbové skupiny genů u drosophily, což odpovídá počtu 4 chromosomů (2n=8) každý chromosom má lineární sekvenci lokusů pro konkrétní geny genetická mapa ovšem nedává absolutní velikost chromosomu: crossing-overy se netrefují ze stejnou pravděpodobností do všech míst chromosomu

124
Genetická mapa Zjednodušená genetuická mapa chromosomu II.
Barvu očí ovlivňuje mnoho genů

125
Cytogenetická mapa cytogenetická mapa se snaží najít geny v souvislosti s různě obarvenými místy na chromosomech cílem je určit přesnou sekvenci všech nukleotidů na chromosomu, délku genů jakož i vzdálenost mezi jednotlivými geny v párech bazí

126
Pohlavní chromosomy člověk: polovina spermií je 22X a polovina spermií 22Y; všechny oocyty jsou 22X anatomicky je pohlaví embrya odlišitelné ve stáří cca dvou měsíců – před tím jsou pohlavní orgány rudimentární – záleží na hormonech, zda vznikne chlapec nebo dívka lidské embryo se vyvíjí původně ženským směrem (VIZ syndrom testikulární feminizace – bude probráíno později)
")
127
Pohlavní chromosomy člověka

128
Pohlavní chromosomy zda se embryo bude vyvíjet mužským nebo ženským směrem záleží na přítomnosti či nepřítomnosti chromosomu Y v roce 1990 byl identifikován gen určující vývoj varlat – byl pojmenován SRY SRY = Sex determining Region of Y pokud gen SRY chybí, gonády se vyvíjejí ženským směrem gen SRY působí pouze jako spouštěč mnoha dalších procesů – SRY kóduje protein, který kóduje mnoho dalších genů

129
Pohlavní chromosomy v současnosti byly identifikovány další geny na chromosomu Y, nutné pro normální vývoj muže pokud jsou tyto geny poškozeny, narodí se chlapec XY, ale nebude schopen tvořit životaschopné spermie

130
Chromozomální určení pohlaví
alozom = nepárový pohlavní chromosom heterogametické pohlaví = to, které má alozom (např. muž) homogametické pohlaví = to, které má pár totožných pohlavních chromosomů
 homogametické pohlaví = to, které má pár totožných pohlavních chromosomů.")
131
Geny vázané na pohlaví geny na pohlavních chromosomech budou vykazovat tentýž vzorec dědičnosti jako běloocí samečkové Morganových drosophil muži budou předávat tyto geny (=na chromosomu X) na všechny své dcery avšak na žádného syna matky budou tyto geny předávat svým synům i dcerám
 na všechny své dcery avšak na žádného syna. matky budou tyto geny předávat svým synům i dcerám.")
132
Geny vázané na pohlaví

133
Geny vázané na pohlaví na chromosomu Y je obecně málo genů – a to jen ty, které určují pohlaví – většina genů vázaných na pohlaví bude na chromosomu X u žen budou recesívně děděné choroby velmi vzácné, protože by musely být recesívně homozygotní muži XY mají naopak jen jeden lokus na chromosomu X – termíny homozygotní či heterozygotní pro ně postrádají smyslu: někdy se říká že jsou hemizygotní

134
Genetické nemoci vázané na pohlaví
Duchenne muscular dystrofy: každý muž v USA; málokdy přežívají 20 rok věku; progresívní oslabení svalů a ztráta koordinace; nemoc je způsobena špatným proteinem zvaným dystrofin, který je klíčový pro správnou práci svalů; gen pro dystrofin leží na chromosomu X hemofilie – absence jednoho nebo více proteinů nutných pro srážlivost krve

135
Královna Viktorie ( ) spontánní mutace buď u otce či u matky královny Viktorie způsobila, že královna se stala přenašečkou prvním hemofilikem byl Leopold, syn královny Viktorie Leopold přežil a stal se otcem dcery- přenašečky, která alelu předala jednomu ze svých synů. Viktorie měla ovšem rovněž i dcery- přenašečky a skrze ně se alela dostala na královské dvory v Prusku, Rusku a Španělsku

136
Královna Viktorie

137
X inaktivace u savčích samiček
ačkoli mají všechny samičky savců dva X chromosomy, jeden z nich bývá inaktivován během embryonálního vývoje výsledkem je, že dospělé buňky muže i ženy mají pouze jeden aktivní X chromosom inaktivní X chromosom u žen kondensuje v kompaktní objekt v jádře, zvaný Barrovo tělísko (Barr body) většina genů na Barrově tělísku nejsou exprimovány (=nepřepisují se do mRNA a netvoří se z nich bílkoviny), ačkoli nějaké zůstávají aktivní (a jsou aktivní v buňkách, ze kterých vzniknou gamety)
 většina genů na Barrově tělísku nejsou exprimovány (=nepřepisují se do mRNA a netvoří se z nich bílkoviny), ačkoli nějaké zůstávají aktivní (a jsou aktivní v buňkách, ze kterých vzniknou gamety)")
138
Barrovo tělísko který z obou X chromosomů se v raném embryu stane Barrovým tělískem, je věcí náhody výsledkem je, že ženy se stávají mozaikou dvou typů somatických buněk: těch s aktivním X chromosomem od matky a s aktivním X chromosomem od otce ve chvíli, kdy je jeden z X chromosomů inaktivován, všechny buňky, které z této buňky vzniknou mitózou, budou mít inaktivován stejný X chromosom

139
Barrovo tělísko je-li tedy žena heterozygot pro gen ležící na X chromosomu, zhruba polovina jejích buněk bude exprimovat jednu alelu, zatímco druhá polovina jejích buněk bude exprimovat druhou alelu jev je krásně pozorovatelný na tříbarevných kočkách u žen je známa mutace, díky které nevznikají potní žlázy – žena pak má potní žlázy mozaikovitě jen na přibližně polovině těla

140
Barrovo tělísko

141
Barrovo tělísko Tři generace žen vykazující nemoc zvanou anhidrotická ektodermální dysplasie (absence potních žláz); všechny ženy jsou pro daný gen umístěný na X chromosomu heterozygotní. Oblasti bez potních žláz jsou vyznačeny zeleně. Rozsah a umístění oblastí bez potních žláz u jednotlivých žen je věcí náhody.
; všechny ženy jsou pro daný gen umístěný na X chromosomu heterozygotní. Oblasti bez potních žláz jsou vyznačeny zeleně. Rozsah a umístění oblastí bez potních žláz u jednotlivých žen je věcí náhody.")
142
Barrovo tělísko inaktivace chromosomu X spočívá v metylaci (=přidání –CH3 skupiny) cytosinů na jeho DNA ale co rozhoduje o tom, který z z obou X chromosomů se stane Barrovým tělískem? byl objeven gen, aktivní pouze na Barrově tělísku, který se nazývá XIST (X-inactive specific transcript). Jeho produktem je RNA, jejíž mnohé kopie pokrývají celý chromosom jaký je ale vztah mezi metylací a XIST genem? - a co rozhoduje o tom, který z obou X chromosomů bude mít aktivní XIST a stane se tak Barrovým tělískem? odpověď není známa
. Jeho produktem je RNA, jejíž mnohé kopie pokrývají celý chromosom. jaký je ale vztah mezi metylací a XIST genem - a co rozhoduje o tom, který z obou X chromosomů bude mít aktivní XIST a stane se tak Barrovým tělískem odpověď není známa.")
143
Změny v počtu chromosomů ústí v genetické choroby
nondisjunkce = jev, kdy se při meiose I. homologické chromosomy nerozdělí správně; nebo se nerozdělí správně sesterské chromatidy při meiose II. v těchto případech získá jedna gameta jednu kopii chromosomu navíc, zatímco druhá nemá žádnou kopii daného chromosomu

144
Změny v počtu chromosomů ústí v genetické choroby

145
Změny v počtu chromosomů ústí v genetické choroby
aneuploidie = stav, kdy se gameta s abnormálním počtem chromosomů spojí s normální gametou. Výsledkem je abnosrmální počet chromosomů v organismu trisomický organismus = buňky mají jeden chromosom navíc (2n+1) monosomický organismus = buňkám jeden chromosom chybí (2n-1) pokud organismus vývoj přežije, ve všech svých buňkách bude mít abnormální počet chromosomů
 monosomický organismus = buňkám jeden chromosom chybí (2n-1) pokud organismus vývoj přežije, ve všech svých buňkách bude mít abnormální počet chromosomů.")
146
Aneuploidie a polyploidie
někdy může nastat podobná chyba i při mitóze. Pokud k nondisjunkci došlo v rané embryonální fázi, řada buněk organismu bude zasažena, což může vést k závažným poruchám některé organismy mají více než dvě sady chromosomů. tento stav je zván polyploidie triploidní (3n) zygota může vzniknout např. tak, že v oocytu je abnormální diploidní stav chromosomů tetraploidie (4n) může vzniknout např. tak, že po první mitóze zygoty nenásleduje cytokineze.
 zygota může vzniknout např. tak, že v oocytu je abnormální diploidní stav chromosomů. tetraploidie (4n) může vzniknout např. tak, že po první mitóze zygoty nenásleduje cytokineze.")
147
Polyploidie polyploidie je relativně častá u rostlin; spontánní polyploidie je u rostlin zřejmě důležitou chvílí pro náhlý vznik nového druhu řada hospodářských druhů rostlin (pšenice, kukuřice) jsou polyploidní rostliny u živočichů je polyploidie velmi vzácná, i když je známa u některých ryb a obojživelníků
 jsou polyploidní rostliny. u živočichů je polyploidie velmi vzácná, i když je známa u některých ryb a obojživelníků.")
148
Polyploidie u savců v Chile byl identifikován zatím první savčí kandidát na polyploidii, hlodavec viskača Tympanoctomys barrerae, který je zřejmě tetraploidní

149
Změny ve struktuře chromosomů delece, duplikace, inverze, translokace
delece – z chromosomu je ztracena část neobsahující centromeru. Takovýto chromosom postrádá pochopitelně mnohé geny. Tento fragment ale nemusí být zcela ztracen, pokud se mu zdaří se připojit k sesterské chromatidě při meiose, vzniká duplikace inverze – stav, kdy se fragment sice připojí k sesterské chromatidě, ale v opačné orientaci

150
Změny ve struktuře chromosomů delece, duplikace, inverze, translokace
translokace – fragment je připojen k nehomologickému chromosomu delece a duplikace se objevují zejména během meiosy při crossing-overu, kdy se části nesesterských chromatid nesprávě připojují pokud má diploidní embryo značné delece či duplikace, zpravidla nepřežije. Rovněž při reciproční translokaci, kdy jsou segmenty nehomologických chromosomů vyměněné navzájem, je sice zachován stav 2n, ale rovněž zpravidla nepřežije. K tomu, aby geny se mohly řádně přepisovat, je totiž nutné, aby byly geny zařazeny ve správné konstelaci u správných promotorů

151
Změny ve struktuře chromosomů delece, duplikace, inverze, translokace

152
Chroroby u lidí způsobené chromosomálními alteracemi
většina aneuploidií je neslučitelná se životem, že buď se embryo ani neimplantuje do endometria dělohy, nebo je spontánně potraceno později některé aneuploidie ale jsou slučitelné se životem. Takoví jedinci mají sadu symptomů (sada symptomů = syndrom), který charakterizuje daný typ aneuploidie
, který charakterizuje daný typ aneuploidie.")
153
Downův syndrom trisomie 21. chromosomu
v USA 1:700 narozených dětí typické rysy tváře („mongolismus“) malá postava srdeční defekty poruchy sluchu náchylnost k respiračním onemocněním mentální retardace náchylní k leukémii a Alzheimerově nemoci smrt ve středním věku, i dříve většina sexuálně podvyvinutá či sterilní
 malá postava. srdeční defekty. poruchy sluchu. náchylnost k respiračním onemocněním. mentální retardace. náchylní k leukémii a Alzheimerově nemoci. smrt ve středním věku, i dříve. většina sexuálně podvyvinutá či sterilní.")
154
Downův syndrom trisomie 21. chromosomu
V některých případech mají lidé s Downovým syndromem obvyklých 46 chromosomů. Výzkum karyotypu však prozradí, že část nebo i celý třetí 21 chromosom je navíc translokován na jiném chromosomu

155
Downův syndrom trisomie 21. chromosomu
Downův syndrom je závislý na věku matky

156
Downův syndrom trisomie 21. chromosomu
těhotenství žen nad 35 let je považováno vůči Downovu syndromu za rizikové závislost věku matky na Downovu syndromu dítěte doposud nebyla vysvětlena většina případů je způsobená nondisjunkcí při meiose I. – monosomie je zřejmě letální. současné výzkumy předpokládají abnormality závislé na věku při napojování mikrotubulů na kinetochory při meiose I. trisomie ostatních chromosomů rovněž závisí na věku matky, i když plody s jinými trisomiemi nepřežívají dlouho

157
Nondisjunkce chromosomu X
nondisjunkce pohlavních chromosomů bývá slučitelná se životem, protože Y chromosom nese jen málo genů a protože všechny nadbytečné kopie X chromosomů jsou uloženy jako Barrova tělíska

158
Klinefelterův syndrom XXY muži
1:2 000 testes jsou malé a muž je sterilní často zvětšená prsa a další ženské znaky inteligence je o něco nižší než průměrná

159
Nondisjunkce pohlavních chromosomů
muži XYY mají tendenci mít vyšší postavu – náchylnost ke zločinům nebyla potvrzena ženy XXX se vyskytují v poměru 1:1000, jsou zdravé a nelze je rozlišit od normálních s výjimkou karyotypu, jejich IQ je však v průměru nižší než u XX žen a je kolem 85. Dva bratři. Vpravo XYY

160
Turnerův syndrom X0 ženy
1:5 000 – jediná známá monosomie u člověka slučitelná se životem sterilní, pohlavní orgány nevyvinuté; pokud je jim podáván estrogen, vyvinou normální sekundární pohlavní znaky většina má normální inteligenci

161
„Cri du chat“ syndrom delece v chromosomu 5 mentální retardace
malá hlava s neobvyklými rysy zvláštní typ pláče (cri du chat) zpravidla umírají v dětském věku
 zpravidla umírají v dětském věku.")
162
Genomický imprinting doposud jsme předpokládali, že každá alela bude mít na organismus stejný efekt, ať již původně pochází z otce nebo z matky… …což většinou můžeme předpokládat bezpečně když jsme například křížili hrášek s fialovými květy s hráškem s bílými květy, bylo jedno, zda fialová alela je ve vajíčku nebo v pylové buňce v současnosti však genetici objevili několik znaků u člověka, včetně některých chorob, u kterých překvapivě záleží na tom, zda je alela zděděna od otce či od matky

163
Genomický imprinting

164
Genomický imprinting Prader-Wiliho syndrom – mentální retardace, obezita, krátká postava, neobvykle malé ruce a nohy Angelmanův syndrom – spontánní, nekontrolovatelný smích, prudké pohyby, a další motorické a mentální symptomy obě choroby mají zřejmě tutéž příčinu: deleci části chromosomu 15

165
Genomický imprinting pokud zdědí dítě abnormální chromosom 15 od tatínka, výsledkem bude Prader-Wiliho syndrom pokud dítě zdědí abnormální chromosom 15 od maminky, výsledkem bude Angelmanův syndrom zdá se, že geny oblasti s delecí by se v normální situaci chovaly jinak pokud by byly zděděné od matky a jinak od otce

166
Genomický imprinting

167
Genomický imprinting Prader-Wiliho/Angelmanova hádanka může být vysvětlena jevem popsaným jako genomický imprinting zdá se, že alela na jednom chromosomu je nějak umlčena, zatímco druhá na homologickém chromosomu se může přepisovat alely stejného genu mají tedy na potomka různý vliv, podle toho, zda se do zygoty dostaly z vajíčka nebo ze spermie

168
Genomický imprinting v potomkovi se v buňkách, které tvoří gamety genomický imprint „vymaže“ a utvoří se znovu podle toho, zda se jedná o muže nebo o ženu imprinting zřejmě spočívá v metylaci (přidání skupiny –CH3) cytosinů na imprintované alele, která se nepřepisuje a je inaktivní někdy ale metylace naopak aktivuje gen! dnes známo asi 20 imprintovaných genů, odhaduje se, že jich budou stovky
 cytosinů na imprintované alele, která se nepřepisuje a je inaktivní. někdy ale metylace naopak aktivuje gen! dnes známo asi 20 imprintovaných genů, odhaduje se, že jich budou stovky.")
169
Genomický imprinting většina imprintovaných genů jsou mimořádně důležité při embryonálním vývoji experimentálně vytvořené myši, které měly dva homologické chromosomy od otce či dva homologické chromosomy od matky vždy umíraly před narozením androgenot = embryo s 2n, obě sady jsou od otce gynogenot = embryo s 2n, obě sady jsou od matky normální vývoj tedy zřejmě předpokládá, aby byla v buňce přítomna přesně jedna kopie aktivního genu: ne dvě a ne žádná

170
Syndrom fragilního X chromosomu
konec X chromosomu visí na tenkém vláknu DNA chlapci 1:500 dívky 1:2 500 mentální retardace velký počet repetic CGG (normální 6 – 54, nemocní nad 200) syndrom je častější, pokud je abnormální chromosom zděděný od matky… …to by odpovídalo pozorování, že syndrom je častější u chlapců: pokud muž zdědí X chromosom, musí to být od matky
 syndrom je častější, pokud je abnormální chromosom zděděný od matky… …to by odpovídalo pozorování, že syndrom je častější u chlapců: pokud muž zdědí X chromosom, musí to být od matky.")
171
Geny mimo jádro malé kruhové DNA se nacházejí v mitochondriích a plastidech tyto cytoplasmatické geny nevykazují mendelovskou dědičnost u rostlin přijímá zygota téměř vždy všechny plastidy z vajíčka a žádný z pylové buňky savci dědí mitochondrie po mateřské linii – ze spermie se zpravidla nedostane žádná mitochondrie do jádra

172
Panašování u rostlin Euonymus fortunei. Dvoubarevná mosaika na listech je způsobena přítomností dvou typů chloroplastové DNA, jedné normální a jedné způsobující bílé zbarvení. Výsledkem jsou různě zbarvené listy, nebo méně často, celé rostliny

173
Geny mimo jádro mitochondriální DNA může způsobit řadu nemocí u lidí – kóduje např. části enzymu ATP-syntázy chyby v mitDNA tedy vyústí v menší množství vyrobeného ATP choroby se tedy projeví nejčastěji v orgánech, které jsou závislé na přísunu energie: svaly a nervová soustava mitochondriální myopathie se např. projevuje slabostí, neschopností snášet fyzickou zátěž a oslabováním svalů

174
Geny mimo jádro chyby v mitDNA přispívají alespoň částečně k některým případům cukrovky, srdečního infarktu a Alzheimerově nemoci

175
Krásný advent přeje Orko

Podobné prezentace