Stáhnout prezentaci
1
Biologie a genetika I.

2
Genetický aparát buňky
DNA = nositelka genetické informace RNA: mRNA = messenger RNA – přenos genetické informace z jádra do cytoplazmy tRNA = transferová – přísun AK k místu syntézy bílkoviny tvar trojlístku, na 1 smyčce antikodon =trojice bazí paralelní ke kodonu na mRNA rRNA = ribozomální – funkce při syntéze bílkovin – součást ribozomů

3
Struktura DNA Watson, Crick 1953: dvoušroubovice = 2 řetězce nukleotidů stočené do spirály Nukleotid: cukr = deoxyriboza (5C) fosfát N baze – purinová : adenin (A), quanin( G) - pyrimidinová : cytozin (C), tymin (T), event. uracil (U) v RNA Komplemetarita bazí, párování vodíkovými můstky A T, C G
 fosfát. N baze – purinová : adenin (A), quanin( G) - pyrimidinová : cytozin (C), tymin (T), event. uracil (U) v RNA. Komplemetarita bazí, párování vodíkovými můstky. A T, C G.")
4
Struktura DNA řetězce antiparalelní Struktura RNA jednořetězcová

5
Struktura DNA

6
Replikace DNA = zdvojení DNA předchází buněčné dělení
Semikonzervativní = ke starému vláknu syntéza nového Replikace začíná na mnoha místech současně (u Eukaryot) Od počátku replikace postup oběma směry – replikační bublina ale vlastní připojování nukleotidů pouze v jednom směru (5´→ 3´)
 Od počátku replikace postup oběma směry – replikační bublina. ale vlastní připojování nukleotidů pouze v. jednom směru (5´→ 3´)")
7
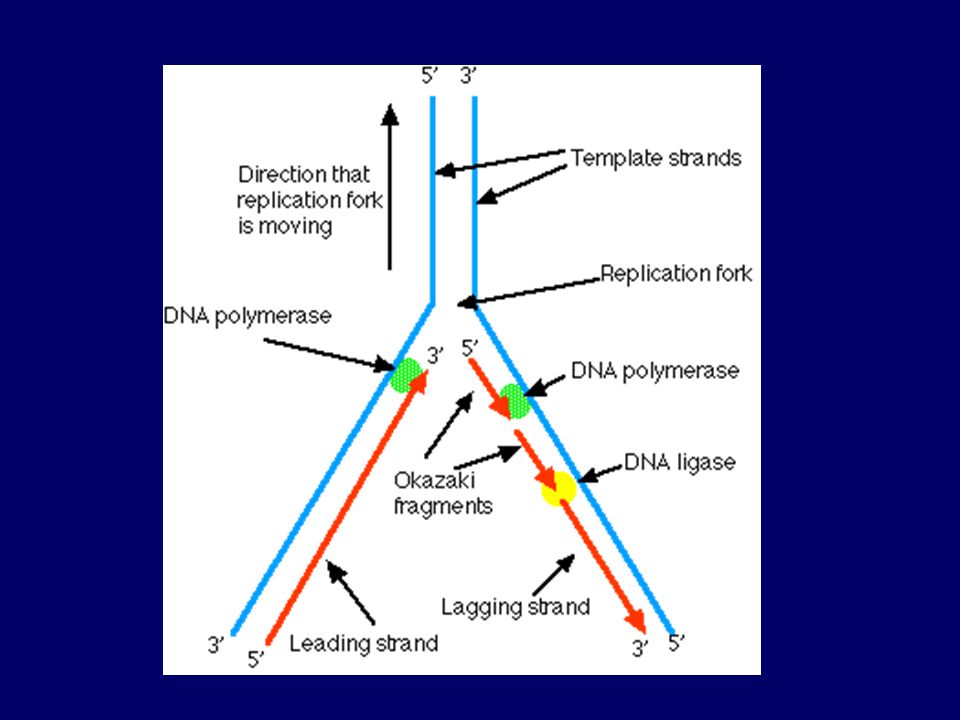
Replikace = enzymatický proces
DNA polymeráza – polymerace + rozpoznání chyb a jejich oprava Další enzymy = rozvíjejí dvoušroubovici a struktury vyšších řádů, spojují fragmenty (ligázy) RNA primer = očko= krátký řetězec ribonukleotidů, k němu připojovány nukleotity dle matrice starého řetězce (DNA polymeráza neumí spojit dva volné nukleotidy, pouze připojovat k již existujícímu řetězci Syntéza vlákna kontinuální ve směru 5´→ 3´, opačné vlákno - syntéza ve formě Okazakiho fragmentů
 RNA primer = očko= krátký řetězec ribonukleotidů, k němu připojovány nukleotity dle matrice starého. řetězce (DNA polymeráza neumí spojit dva volné. nukleotidy, pouze připojovat k již existujícímu řetězci. Syntéza vlákna kontinuální ve směru 5´→ 3´, opačné. vlákno - syntéza ve formě Okazakiho fragmentů.")
9
Centrální dogma molekulární biologie

10
Exprese genetické informace
Transkripce = přepis DNA do mRNA - enzym RNA polymeráza počátek= promotor Stavba eukaryontního genu : exony = kodující části introny = nekodující části Posttranskripční úpravy = vyštěpení intronů z mRNA = splicing a transport upravené mRNA do cytoplazmy na ribozom

11
Translace = překlad = syntéza polypeptidu dle matrice mRNA
probíhá na ribozomech – zde se mRNA napojí ribozom = proteiny + rRNA, ze 2 podjednotek tRNA přináší jednotlivé aminokyseliny k místu syntézy polypeptidu má antikodon = specifický triplet pro určitou AK a komplementární ke kodonu (trojici bazí) na mRNA kam se napojí při napojení dvou tRNA na specifická místa ribozomu – vznik peptidové vazby mezi dvěma aminokyselinami nebo AK a polypeptidem
 na. mRNA kam se napojí. při napojení dvou tRNA na specifická místa ribozomu – vznik peptidové vazby mezi dvěma aminokyselinami nebo AK a polypeptidem.")
12
Vlastnosti genetického kódu
Kodon = triplet(trojice bazí) na mRNA kodující 1 aminokyselinu (AK) Genetický kód: tripletový = trojice nukleotidů (bazí) určuje 1 AK degenerovaný pro 1 AK existuje více tripletů čtení je nepřekrývající univerzální iniciační a terminační kodony
 na mRNA kodující 1 aminokyselinu (AK) Genetický kód: tripletový = trojice nukleotidů (bazí) určuje 1 AK. degenerovaný pro 1 AK existuje více tripletů. čtení je nepřekrývající. univerzální. iniciační a terminační kodony.")
13
Definice genu Gen = takový úsek DNA molekuly, který svojí primární strukturou (pořadím nukleotidů, resp.tripletů) určuje primární strukturu jiné makromolekuly (polypeptidu, tRNA, rRNA)
 určuje primární strukturu jiné makromolekuly (polypeptidu, tRNA, rRNA)")
14
Ultrastruktura chromozomů:
Složení chromatinu: DNA histony = bazické bílkoviny: H1,H2A, H2B,H3,H4 nehistonové bílkoviny = kyselé

15
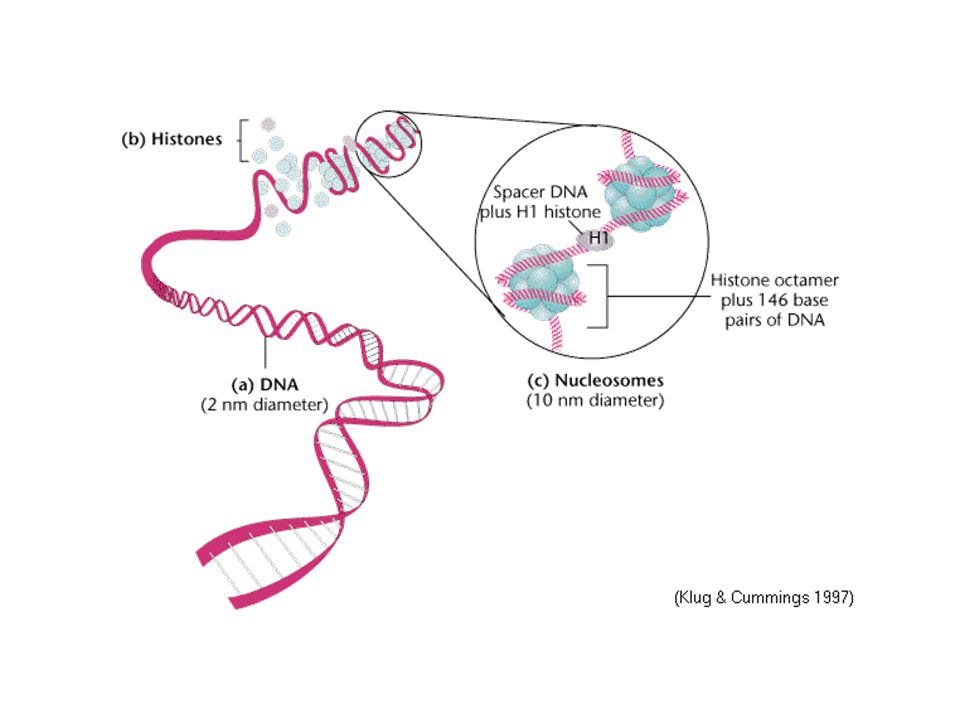
Organizace interfázního chromatinu
Nukleozom = jádro z oktameru histonů H2A,H2B,H3,H4 obtočené dvoušroubovicí DNA spojka mezi nukleozomy = vlákno DNA, volné nebo asociované s H1 histony (utažení vlákna) řetězec nukleozomů stočen do solenoidu
 řetězec nukleozomů stočen do solenoidu.")
16
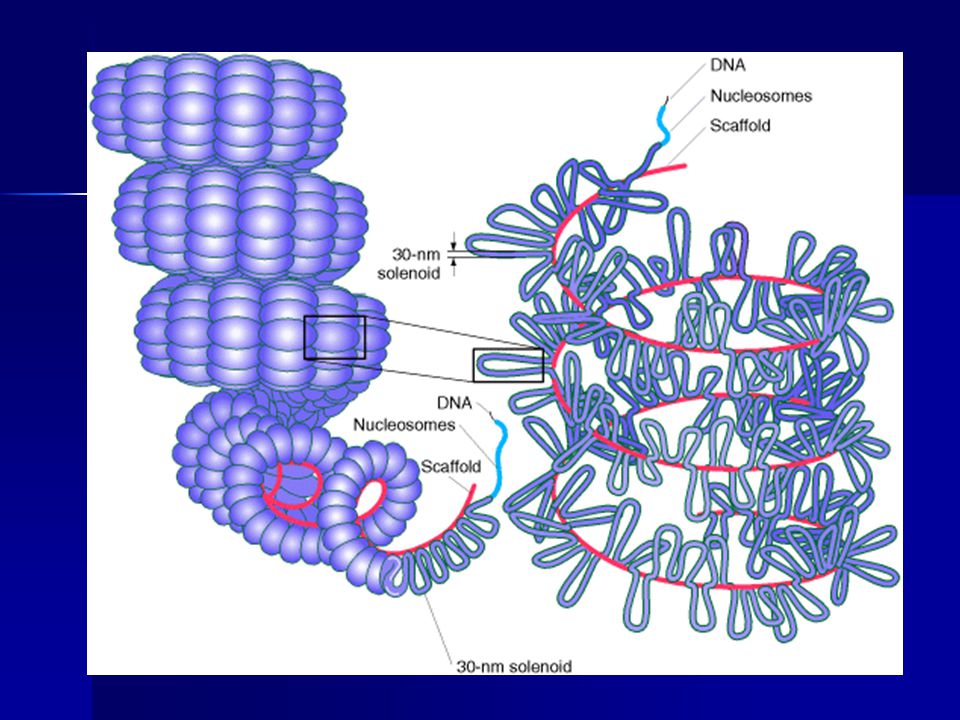
Další stupně spiralizace = kondenzace chromatinu do chromozomů
Solenoidy poskládány do smyček obtáčejících osu z nehistonových bílkovin Další skládání a utažení smyček při přechodu chromozomů z profáze do metafáze= mnohonásobné zkrácení vlákna DNA

17
Zkrácení v profázi na 1/3000 délky
v metafázi na 1/10000 délky celková délka DNA asi 2 m lidské chromozomy obsahují cca strukturních genů

21
metacentrický submetacentrický akrocentrický
p telomera centromera satelit sat. stopka (NOR) q chromatidy Chromozom metacentrický submetacentrický akrocentrický p = krátké rameno q = dlouhé rameno NOR = oblast organizátoru jadérka (rRNA geny)
 q. chromatidy. Chromozom. metacentrický submetacentrický akrocentrický. p = krátké rameno. q = dlouhé rameno. NOR = oblast organizátoru jadérka (rRNA geny)")
22
Chromatin Euchromatin Heterochromatin
despiralizován v interfázi spiralizován v mitoze obsahuje strukturní geny Heterochromatin repetitivní sekvence,neobsahuje strukturní geny v interfázi poněkud spiralizován – barvitelný

23
Heterochromatin konstitutivní – stálý
-v centromerických oblastech všech chromozomů - heterochromatinové bloky na 1q, 9q, 16q, Yq variabilita heterochromatinových částí fakultativní = strukturně euchromatin, chová se jako heterochromatin = neaktivní Inaktivní X = sex chromatin=Barrovo tělísko =X chromatin jeden ze dvou X chromozomů u samic savců je geneticky inaktivní (= není transkribován) heterochromatin-pozdější replikace v S fázi (inaktivní X na konci S) žena=mozaika buněk s inaktivním otcovským a mateřským X
 heterochromatin-pozdější replikace v S fázi. (inaktivní X na konci S) žena=mozaika buněk s inaktivním otcovským a mateřským X.")
24
X inaktivace XM XP XM XP XM XP XM XP XM XP XM XP XM XP XM XP XM XP XM XP XM XP XM XP XM XP XM XP XM XP

25
Chromozomy pozorovatelné jen v dělící se buňce
U člověka 46 chromozomů = diploidní počet 22 párů autozomů + 1 pár gonozomů XX u ženy XY u muže Homologní chromozomy – původ otcovský a mateřský (původní chromozom spermie a vajíčka) Gamety = haploidní počet
 Gamety = haploidní počet.")
26
Genetická determinace pohlaví
23,X 23,X 23,Y

27
Karyotyp ženy 46,XX – G pruhy

28
Karyotyp muže - 46,XY – G pruhy

29
Buněčný cyklus interfáze : G1, S, G2 mitoza
G1 fáze - nejdelší, variabilní část cyklu syntéza RNA, proteinů, doplnění organel (ribozomů, mitochondrií, ER apod.) syntéza nukleotidů, příprava na replikaci kontrolní bod cyklu pro vstup do S fáze
 syntéza nukleotidů, příprava na replikaci kontrolní bod cyklu pro vstup do S fáze.")
30
S fáze - replikace jaderné DNA
mimojaderná DNA (=mitochondriální) se replikuje i mimo S fázi G2 fáze - růst buňky, syntéza proteinů, RNA, tvorba buněčných struktur syntéza rychlejší, transkripce pravděpodobně z obou chromatid kontrolní bod cyklu pro vstup do mitozy Mitoza - spiralizace chromozomů, rozpad jaderné membrány, vytvoření mitotického aparátu- rozdělení chromatid, rozdělení jádra Cytokineze – rozdělení buňky
 se replikuje i mimo S fázi. G2 fáze - růst buňky, syntéza proteinů, RNA, tvorba buněčných struktur. syntéza rychlejší, transkripce pravděpodobně z obou chromatid. kontrolní bod cyklu pro vstup do mitozy. Mitoza - spiralizace chromozomů, rozpad jaderné membrány, vytvoření mitotického aparátu- rozdělení chromatid, rozdělení jádra. Cytokineze – rozdělení buňky.")
31
M G1 G2 S

32
Regulace buněč.cyklu: Signál k dělení přijat receptory na buněč.membráně nebo v cytoplasmě signální molekuly = cytokiny (růstové faktory), hormony Signál předán prostřednictvím kaskády regulačních molekul do nitra buňky až k jaderným genům, které svými produkty přímo aktivují buněčný cyklus

33
Geny regulující buněčné dělení:
Protoonkogeny produkty stimulují buněčné dělení (př. myc, fos, jun aj.) kódují růstové faktory, receptory růstových faktorů, regulační proteiny apod. mutované formy = onkogeny = stálá nebo nadměrná mitotická aktivita
 kódují růstové faktory, receptory růstových faktorů, regulační proteiny apod. mutované formy = onkogeny = stálá nebo nadměrná mitotická aktivita.")
34
Nádorové supresorové geny
produkty inhibují mitotické dělení př. Rb1 gen – mutace u retinoblastomu i jiných tumorů p53 – mutace genu v mnoha nádorech nadřazen dalším genům funkce: zástava cyklu v G1 při poškození DNA a navození reparace nebo apoptózy - programované buněčné smrti

35
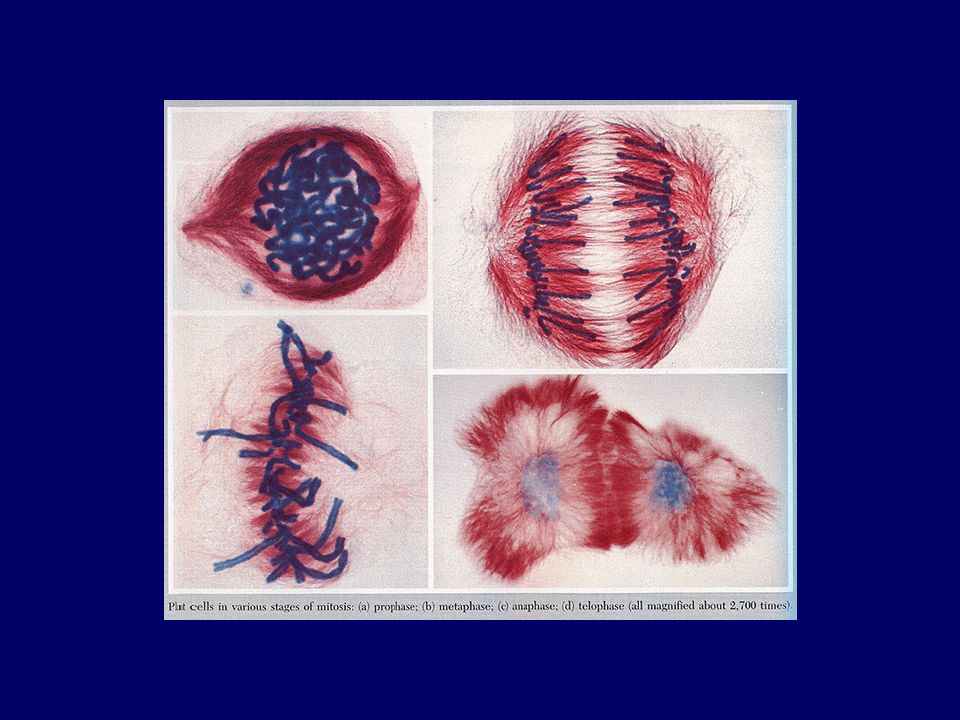
Mitoza = dělení somatických buněk
z diploidní buňky mateřské – 2 diploidní buňky dceřinné

36
profáze interfáze prometafáze Mitoza 2n 2n metafáze telofáze anafáze

37
Meioza – dělení pohlavních buněk
I.Meiotické dělení = heterotypické = vlastní redukční dělení – homologní chromozomy se jednotlivě rozcházejí do dceřinných buněk Z diploidní buňky mateřské – 2 haploidní buňky dceřinné II.Meiotické dělení = homeotypické = ekvační = mitóza – štěpení centromer a rozchod chromatid do dceřinných buněk

38
n M I = redukční 2n = heterotypické Profáze : leptotene - počátek spiralizace zygotene - párování (synapse homologů) = bivalenty synaptonemální komplex zajištuje párování homologů párování X a Y pouze konci - sexuální váček

39
pachytene - patrny sesterské chromatidy = tetrády
crossing over mezi nesesterskými chromatidami homologních chromozomů = rekombinace otcovského a mateřského chrom.materiálu diplotene - separace bivalentů - spojeny v místě crossing overu = chiasmata diakineze - maximální zkrácení chromozomů - terminalizace chiazmat

40
Metafáze: orientace párů chromozomů v ekvatoriální rovině, centromery se neštěpí
Anafáze: rozchod homologů - náhodný – náhodné kombinace otcovských a mateřských chromozomů Telofáze: haploidní sady na opačných polech Interkineze: bez replikace

41
M II = homeotypické = n ekvační n = mitotické
v metafázi štěpení centromer v anafázi separace chromatid

42
I. Meiotické dělení profáze leptotene zygotene pachytene diplotene
crossing over diakineze anafáze telofáze

43
II.Meiotické dělení anafáze

44
anafáze M I telofáze M I Rozchod chromozomů - náhodný z hlediska rodičovského původu

45
II.Meiotické dělení anafáze telofáze

48
Spermiogeneza – v době sexuální zralosti
spermatogonie Mitotické dělení růst primární spermatocyt M I sekundární spermatocyt meioza M II spermatidy zrání, diferenciace spermie Spermiogeneza – v době sexuální zralosti

49
Oogeneza – začátek v prenatálním období
oogonie Mitotické dělení růst 3.měs.fetál.života dictyotene MI v době porodu primární oocyt M I meioza 1.pol.tělísko Metafáze MII ovulace M II sekundární oocyt 2.pol.tělísko Anafáze,telofáze po oplození Oplození – pronukleus vajíčko dokončí MII- pronukleus zygota Oogeneza – začátek v prenatálním období pokračování v době sexuální dospělosti

50
Výsledek oogenese z 1 diploidní buňky = 1 vajíčko(většina cytoplasmy)
+ 2 až 3 pólová tělíska úbytek mnoha buněk: v 5. měsíci fetálního života x 106 buněk v době porodu x 106 buněk v pubertě buněk ovuluje buněk dlouhý interval mezi započetím a dokončením meiotického dělení = faktor nondisjunkce , chyby v rozdělování chromozomů u starších žen

51
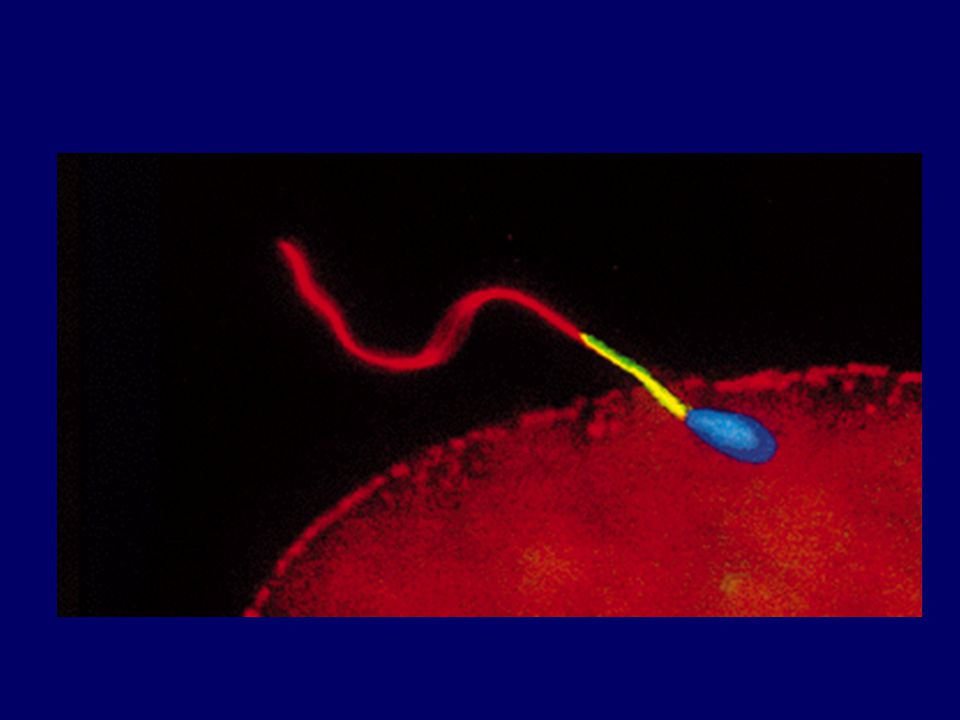
Oplození vajíčka v metafázi MII
spermie menší, nejsou zásobní látky akrozomální váček - enzymy vaječná b. - zásobní látky pod cytoplazmat.membránou sekreční váčky (kortikální granula) vně zona pellucida
 vně zona pellucida.")
52
spermie = samčí pronukleus
(22 autozomů + 1 gonozom-X nebo Y) vajíčko dokončí MII = samičí pronukleus (22 autozomů+1 gonozom X) fuze haploidních jader = zygota - replikace - mitotické dělení dává vznik populaci buněk budoucího zárodku
 vajíčko dokončí MII = samičí pronukleus. (22 autozomů+1 gonozom X) fuze haploidních jader. = zygota - replikace - mitotické dělení dává vznik populaci buněk budoucího zárodku.")
53
Spermiogeneza M I M II 2n n n spermatogonie spermatida
primární sekundární spermatocyt spermie

54
Oogeneza a fertilizace
I. meiotické dělení polové tělísko oogonie primární sekundární oocyt n 2n fertilizace a II.meiotické dělení

57
Genetické důsledky meiozy
redukce diploidního počtu chromozomů na haploidní segregace alel v MI , MII náhodný rozchod chromozomů – náhodné kombinace chromozomů v gametách (dle rodičovského původu) zvýšení genetické variability crossing overem (segregující chromozom složen z částí mateřského a otcovského původu)
 zvýšení genetické variability crossing overem (segregující chromozom složen z částí mateřského a otcovského původu)")
58
Chyby v meioze nebo mitoze
Nondisjunkce- chybný rozchod chromozomů nebo chromatid –jdou spolu do 1 dceřinné buňky Vzniklé gamety mají navíc chromozom - po oplození je výsledkem trizomie, např Downův syndrom 47,XX nebo XY,+21 nebo gameta nemá žádný chromozom, po oplození je monozomie Opoždění chromozomu v anafázi a nezačlenění do dceřinné buňky – zde pak chybí chromozom – po oplození místo chromozomálního páru je pouze 1 chromozom, např Turnerův sy – monozomie X

59
Chyby v meioze 46 46 M I 23 24 22 23 M II 23 23 23 23 24 24 22 22 nondisjunkce v M I důsledek: trizomie/monozomie po oplození normální průběh meiozy

60
46 46 M I 23 22 23 23 (X chrom.) M II 24 22 23 23 22 22 23 22 Opoždění chromozomu v anafázi M I nebo M II Důsledek: monozomie po oplození Nondisjunkce v M II Důsledek: trizomie/monozomie po oplození

61
46 46 M I 46 23 23 M II 46 46 23 23 46 Chyby v meiose – nondisjunkce celé sady chromozomů (M I nebo M II) Důsledek: neredukovaná gameta, triploidie po oplození

62
Poruchy oplození oplození 46 69 XX XXY
Dispermie- oplození vajíčka 2 spermiemi









>")






>")
 = M.>")


>")



