Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Remodelace srdce a cév za patologických stavů

3
Srdeční buňky Kardiomyocyty- 75% normálního tkáňového objemu myokardu, ale pouze 30–40% počtu buněk. Většina jsou ne-kardiomyocyty, zejména fibroblasty, dále endoteliální buňky, buňky hladké svaloviny cév aj. Fibroblasty jsou v kontaktu s myocyty; každý kardiomyocyt je v normální srdeční tkáni přiřazen ke „svému“ fibroblastu. Patologické stavy srdce jsou spojeny s myokardiální remodelací, často s fibrózou (ICHS, srdce po revmatické horečce, zánětu při hypertrofii, infarktu. Růst fibrózní tkáně je dán proliferativním potenciálem fibroblastů, který ovšem chybí cardiomyocytům.

4
Charakteristiky srdečních fibroblastů
Významná buněčná populace v srdci Spoluzodpovídá za structurální, biochemické, mechanické elektrické vlastnosti myokardu

5
Fibroblasty Buňky mezenchymálního původu, které produkují intersticilání kolagen, kolagen typu I, III a VI (myocyty tvoří kolegan typu IV, který je součástí jejich bazální mebrány). Jsou v principu motilní - obsahují aktin (zejména α-aktin hladké svaloviny) a myosin. Jsou pleiomorfní - jejich obsah aktinu a myosinu může být ovlivněn prostředím, především mechanickými silami.
. Jsou v principu motilní - obsahují aktin (zejména α-aktin hladké svaloviny) a myosin. Jsou pleiomorfní - jejich obsah aktinu a myosinu může být ovlivněn prostředím, především mechanickými silami.")
6
Původ srdečních fibroblastů
1. Proepikardiální orgán 2. Epiteliálně mezenchymální transformace během vývoje chlopní Málo pojivové tkáně v časně embryonálním srdci Podstatnější tvorba během tvorby epikardu a chlopní. 3D kolagenová síť se vytváří v pozdně fetálním období. Dokončení až po narození s rychlou proliferací fibroblastů a tvorbou kolagenu Po dokončení novorozeneckého vývoje už jen velmi nízká úroveň proliferace fibroblastů, reagující na fyziologickou i patofyziologickou stimulaci.

7
Původ srdečních fibroblastů
Obsah fibroblastů dále v průběhu života a stárnutí roste (až dvě třetiny všech buněk). Jsou nazývány “sentinelovými“ buňkami, které fungují jako lokální imunomodulátory a modulátory srdeční elektrofyziologie.
. Jsou nazývány sentinelovými buňkami, které fungují jako lokální imunomodulátory a modulátory srdeční elektrofyziologie.")
8
Kolagen v srdci Depozice kolagenu v zdravém dospělém srdci je velmi malá. Během stárnutí se zvyšuje podíl „cross-linked“ kolagenu, což vede k tuhnutí tkáně. Dramaticky navýšený podíl kolagenu u patologických stavů, jako jsou hypertrofie, srdeční selhání, IM. Tato asociovaná proliferace fibrózní tkáně se dělí dle Webera et al. Do dvou kategorií 1) reparační fibróza (disperzní), a (2) reaktivní fibróza, která se objevuje na začátku kolem kapilár, teprve později se rozšiřuje po myokardu. Tyto fibroblasty reagují na cirkulujícíc hormony, které jsou schopny ovlivňovat jejich odpovědi na patologické stimuly. Zdá se, že tyto perivaskulární, proliferující fibroblasty v srdci pocházejí z cirkulujících progenitorů, odvozených z kostní dřeně.
 reparační fibróza (disperzní), a. (2) reaktivní fibróza, která se objevuje na začátku kolem kapilár, teprve později se rozšiřuje po myokardu. Tyto fibroblasty reagují na cirkulujícíc hormony, které jsou schopny ovlivňovat jejich odpovědi na patologické stimuly. Zdá se, že tyto perivaskulární, proliferující fibroblasty v srdci pocházejí z cirkulujících progenitorů, odvozených z kostní dřeně.")
9
Vlastnosti srdečních fibroblastů
Udržují strukturu srdeční tkáně včetně homeostázy extracelulární matrix (ECM)a produkce faktorů udržujících rovnováhu mezi syntézou a degradací složek pojivové tkáně, jako jsou cytokiny, růstové faktory, matrix metaloproteinázy (MMP) aj. Hrají zásadní roku v srdeční remodelaci (hypertrofie kardiomyocytů, migrac a proliferace fibroblastů, změny rozsahu a složení srdeční ECM. Excesivní proliferace fibroblastů a nárůst obsahu proteinů ECM (fibróza) indukuje ztuhnutí myokardu jako vlastnost přispívající k dysfunci myokardu. Fibróza je spojena se zvýšenou produkcí MMP a dalších faktorů jako TGF-β, angiotenzin II, endotelin-1 and tumour necrosis factor-α (TNF-α).
a produkce faktorů udržujících rovnováhu mezi syntézou a degradací složek pojivové tkáně, jako jsou cytokiny, růstové faktory, matrix metaloproteinázy (MMP) aj. Hrají zásadní roku v srdeční remodelaci (hypertrofie kardiomyocytů, migrac a proliferace fibroblastů, změny rozsahu a složení srdeční ECM. Excesivní proliferace fibroblastů a nárůst obsahu proteinů ECM (fibróza) indukuje ztuhnutí myokardu jako vlastnost přispívající k dysfunci myokardu. Fibróza je spojena se zvýšenou produkcí MMP a dalších faktorů jako TGF-β, angiotenzin II, endotelin-1 and tumour necrosis factor-α (TNF-α).")
10
Vlastnosti srdečních fibroblastů
MMP jsou produkovány také kardiomyocyty, endoteliálními buňkami a prozánětlivými buňkami. Účastní se regulace buněčného růstu, migrace, buněčného přežití a angiogenezy. Angiotensin II, TGF-β a tumour necrosis factor-α se účastní autokrinní a parakrinní regulace hypertrofie myocytů, proliferace fibroblastů a obratu proteinů ECM. Angiotensin II dále stimuluje expresi genů pro kolagen a pro syntézu kolagenu a redukuje degradaci kolagenu (snížením aktivity MMP v srdečních fibroblastech), zatímco endotelin-1 indukuje hypertrofii myocytů a stimulaci syntézy kolagenu. Remodelace myokardu může být stimulována také chronickou adrenergní stimulací (chronické srdeční selhání). Statiny přímo inhibují proliferaci fibroblastů (prevence srdeční remodelace?)
, zatímco endotelin-1 indukuje hypertrofii myocytů a stimulaci syntézy kolagenu. Remodelace myokardu může být stimulována také chronickou adrenergní stimulací (chronické srdeční selhání). Statiny přímo inhibují proliferaci fibroblastů (prevence srdeční remodelace )")
11
Elektrická signalizace
Fibroblasty ji mohou ovlivnit pasivně (např. separací skupin svalových buněk, což může vést ke zpoždění šíření vzruchu). Intersticiáoní fibróza a akumulace kolagenu může být zdrojem lokální anizotropieu ischemie a hypertofie-predipsozice k arytmogenezi. Aktivně- jako mechanoelektričtí transduceři. Mají aktivní potenciál (napěťově (stretch)-aktivované iontové kanály, propustné pro Na+, K+ a Ca2+).
. Intersticiáoní fibróza a akumulace kolagenu může být zdrojem lokální anizotropieu ischemie a hypertofie-predipsozice k arytmogenezi. Aktivně- jako mechanoelektričtí transduceři. Mají aktivní potenciál (napěťově (stretch)-aktivované iontové kanály, propustné pro Na+, K+ a Ca2+).")
12
Elektrická signalizace
Přímé „gap-junction coupling“ fibroblastů a myocytů? Fibroblasty se za patologických stavů stávají senzitivnějšími na mechanický stres, což může při spojení s myocyty vést až k ovlivnění aktivity pacemakeru v síních.

13
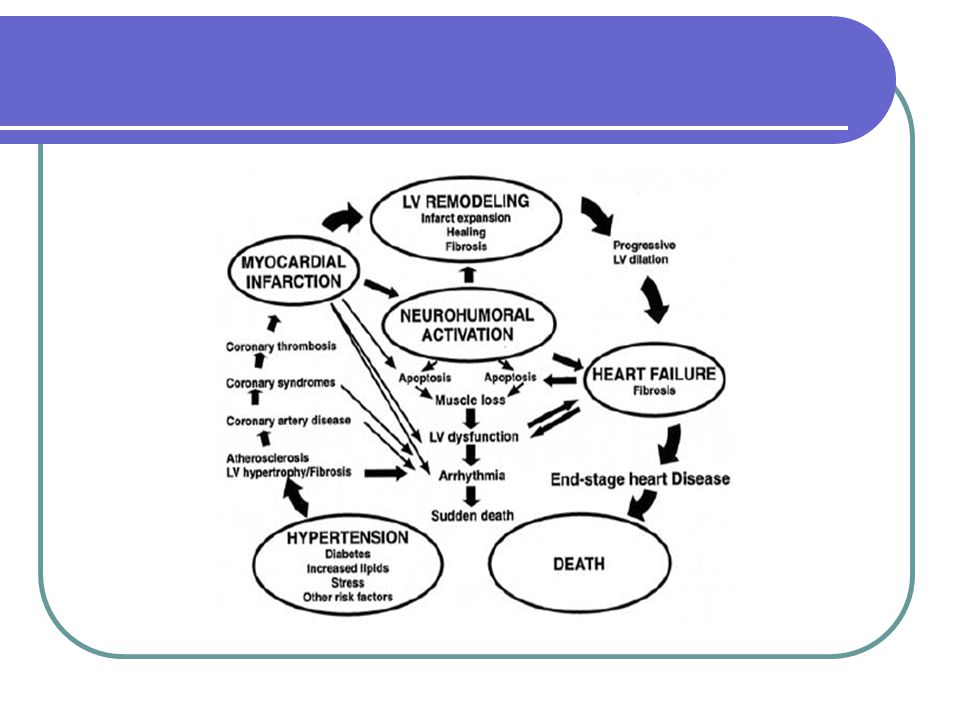
Remodelace levé komory
Up-regulace matricelulárních proteinů v důsledku poškození myokardu?

14
Matricellulární proteiny
Proteiny ECM, které modulují interakce buňka matrix. O jejich strukturální funkci je pochybnost. Rodina zahrnuje tenascin-C (TN-C), tenascin-X (TN-X), osteonektin, osteopontin, thrombospondin-1 (TSP1) a thrombospondin-2 (TSP2).
, tenascin-X (TN-X), osteonektin, osteopontin, thrombospondin-1 (TSP1) a. thrombospondin-2 (TSP2).")
15
Matricelulární proteiny
Jejich exprese je vysoká během embryogeneze, ale téměř chybí postnatálně. Znovu se objevuje jako reakce na poškození.

16
Chronický stres Chronický stres v srdci aktivuje expresi příslušných genů. Signální transdukce vyvolaná ANP a růstovými faktory iniciuje remodelaci (hypertrofii), která přispívá k elektrické nestabilitě zvýšenou senzitivitou mechanosenzitivních kanálů.
, která přispívá k elektrické nestabilitě zvýšenou senzitivitou mechanosenzitivních kanálů.")
17
Změny indukované akutním a chronickým napětím (stretch), což může vést
k arytmiím

18
Kolagen I Rovnováha mezi produkcí a degradací kolagenu typu I je rozhodující pro udržení tkáňové i orgánové integrity včetně procesu tkáňové reparace v reakci na poškození. Je regulována na transkripční úrovni různými cytokiny (transforming growth factor (TGF-). Tumor necrosis factor (TNF- ) redukuje expresi tohoto kolagenu.
. Tumor necrosis factor (TNF- ) redukuje expresi tohoto kolagenu.")
19
Signální transdukce prostřednictvímTGF-

20
Signální transdukce prostřednictvímTNF

21
Apoptóza v srdci Morfogeneze srdce se během vývoje účastní apoptóza.
Během embryogeneze dochází k apoptóze kardiomyocytů, zatímco po narození apoptóza moduluje vývoj převodního systému.

22
Ionty během adaptace Srdce se adaptuje na mechanické přetížení změnou velikosti buněk a změnou exprese iontových kanálů a iontových transportérů (remodelace buněk a buněčných kanálů). Tento stav je po nějakou dobu kompenzovatelný. Pokud dojde k trvalému nebo dalšímu přetížení, dojde k rozvoji systolické a diastolické dysfunkce. Hypertrofie vždy vede ke zvýšenému riziku rozvoje arytmií a náhlé srdeční smrti.
. Tento stav je po nějakou dobu kompenzovatelný. Pokud dojde k trvalému nebo dalšímu přetížení, dojde k rozvoji systolické a diastolické dysfunkce. Hypertrofie vždy vede ke zvýšenému riziku rozvoje arytmií a náhlé srdeční smrti.")
23
Strukturální remodelace síňového myokardu během fibrilace síní.
Nárůst objemu kardiomyocytů, zánik sarkomer, akumulace glykogenu, abniormality mitochondrií. Dediferenciace a degenerace kardiomyocytů. Rozvíjí se adaptační reakce, odlišná od apoptózy i nekrózy: programované buněčné přežití. Jde o adaptaci buněk na ischémii a zvýšené mechanické napětí (stretch).
.")
24
Srdeční hypertrofie Nárůst velikosti kardiomyocytů
Zvýšená syntéza proteinů Vyšší stupeň organizace sarkomer, kterou předchází a doprovází reaktivatce několika fetálních genů.

25
Srdeční hypertrofie Strukturální remodelace slouží jako adaptační reakce k udržení normálního napětí v srdeční stěně při normálním srdečním výdeji. U sportovců (atletů) je tato hypertrofická odpověď indukovaná fyzickou zátěží považovaná za kompenzační. Prolongovaná hypertrofie jako odpověď na patologické stavy nevyhnutelně vede l maladaptačním změnám, které zvyšují riziko arytmií a podporují rozvoj srdečního selhání.
 je tato hypertrofická odpověď indukovaná fyzickou zátěží považovaná za kompenzační. Prolongovaná hypertrofie jako odpověď na patologické stavy nevyhnutelně vede l maladaptačním změnám, které zvyšují riziko arytmií a podporují rozvoj srdečního selhání.")
26
Srdeční hypertrofie Mezi kompenzační a maladaptační hyoetrofií jsou rozdíly: U hypertrofie indukované fyzickou zátěží se tloušťka srdeční stěny zvětšuje jen málo a není přítomna srdeční fibróza. Není jasné, zda kompenzační hypertrofie vždy předchází maladaptivní, zda v ni přechází, nebo zda je část kompenzačních hypertrofií maladaptačních od začátku. Pomohou nám rozdíly v expresních profilech .

27
Hypertrofická remodelace
U maladaptačních hypertrofií zvýšené riziko fatálních komorových arytmií Alterace v délce akčního potenciálu Poruchy metabolismu Ca++ Re-entry fenomén Poruchy vedení

28
Hypertrofická remodelace
Zjištěné poruchy ve vedení elektrických impulsů od buňky k buňce může být dáno změnami v expresním profilu a ve složení gap junctions. Gap junctions jsou lokalizovány především v interkalátních discích kardiomyocytů a skládají se z mnohočetnách kanálů v gap junction. Kanály v gap junction jsou tvořeny jejich proteiny (konexiny); 6 konexinů vytváří konexon (hemikanál) na povrchu jedné buňky, který se spojuje s polokanálem sousední buňky a tvoří dohormoday intercelulární kanál.
; 6 konexinů vytváří konexon (hemikanál) na povrchu jedné buňky, který se spojuje s polokanálem sousední buňky a tvoří dohormoday intercelulární kanál.")
29
Hypertrofická remodelace
V kardiomycytech jsou exprimovány produkty tří konexinových genů (Cx43, Cx40, Cx45). V komorách převažuje exprese Cx43 jak v kardiomyocytech, tak v buňkách převodního systému. Cx40 a Cx45 jsou exprimovány v buňkách převodního systému. Cx45 exprimován také lokálně ve stěně komor. Porucha v elektrickém coupling a v expresi konexinů vede k poruchám v srdeční elektrofyziologii a k arytmogenezi. Poruchy může způsobit srdeční fibróza rozrušením spojů mezi buňkami.
. V komorách převažuje exprese Cx43 jak v kardiomyocytech, tak v buňkách převodního systému. Cx40 a Cx45 jsou exprimovány v buňkách převodního systému. Cx45 exprimován také lokálně ve stěně komor. Porucha v elektrickém coupling a v expresi konexinů vede k poruchám v srdeční elektrofyziologii a k arytmogenezi. Poruchy může způsobit srdeční fibróza rozrušením spojů mezi buňkami.")
30
Konexiny

31
Hypertrofická remodelace
Srdeční onemocnění jsou často spojena a alteracemi v množství a distribuci kanálů v gap junction. V iniciální fázi hypertrofie jsou konexiny zvýšeně exprimovány; později dochází k jejich down regulaci, což je projevem počínajícího srdečního selhání.

32
Hypertrofická remodelace
Redukce proteinů v sarkomerách patrně ovlivňuje kontraktilní vlastnosti srdečních síní. Změněné distribuce a exprese kanálových proteinů patrně interferují s udržováním hladin iontů a se spžažením excitace-kontrakce. Dochází ke změně vedení vzruchu v síních.

33
Ionty během adaptace U srdečního selhání dochází k up regulaci Na/Ca výměníku, což se považuje za mechanismus kompenzace nižšího vychytávání vápníku v cytoplasmě v důsledku horší funkce pumpy SERCA2. To ovlivňuje hladiny obou iontů.

35
Remodelace cévní stěny u patologických stavů

36
Cévní remodelace Fyziologický i patofyziologický proces
Kompenzační ztenčení aterosklerotických cév. Teprve po překročení kapacitních možností ztenčení dochází k jejich zúžení. Konstriktivní remodelace vede k omezení průtoku.

37
Cévní remodelace Dosud není jasné, zda remodelace je primárně odpovědí medie, adventicie nebo obou. Tyto stavy vedou k ztluštění nebo ztenčení medie, což je negativní nebo pozitivní remodelace.

38
Cévní remodelace Postupný proces
Konstriktivní remodelace (negativní) s malým podílem tvorby neointimy a velkým podílem stenózy Expanzivní (pozitivní) remodelace-velký podíl tvorby neointimy na dilatované cévě.
 s malým podílem tvorby neointimy a velkým podílem stenózy. Expanzivní (pozitivní) remodelace-velký podíl tvorby neointimy na dilatované cévě.")
39
Cévní remodelace Časná ateroskleróza-kompenzační vazodilatace, pozdní ateroskleróza-stenóza. Smysl angioplastiky je zabránit cévě v hojení a v nastolení nevhodného kalibru po odeznělé vazodilataci.

40
Cévní remodelace Adventicie má největší rychlost buněčné proliferace ve srovnání s ostatními vrstvami cévní stěny. Fibroblasty jsou zde schopny diferencovat se v myofibromyoblasty. Ty jsou schopny syntetizovat kolagen. Vytváří se restriktivní adventiciální jizva.

41
Cévní remodelace Účast mezibuněčné matrix.
Angioplastika způsobuje akutní změny v syntéze a degradaci extracelulární matrix. To vede ke zvýšení syntézy kolagenu a snížení aktivity matrix metaloproteináz (MMP) a tedy degradace extracelulární matrix. Denzita neointimálního kolagenu je vyšší v arteriích s negativní remodelací ve zvířecích modelech, což pdoporuje hypotézu, že zvýšená syntéza kolagenu podporuje vznik restenózy konstriktivní remodelací cévní stěny po provedené angioplastice.
 a tedy degradace extracelulární matrix. Denzita neointimálního kolagenu je vyšší v arteriích s negativní remodelací ve zvířecích modelech, což pdoporuje hypotézu, že zvýšená syntéza kolagenu podporuje vznik restenózy konstriktivní remodelací cévní stěny po provedené angioplastice.")
42
Cévní remodelace Zvýšený obsah kolagenu a snížená aktivita MMP identifikována v místech restenózy na rozdíl od primární plaky nebo normální cévy. Velký význam remodelace extracelulární matrix.

43
Cévní apoptóza Apoptóza hladkých svalových buněk je řízena:
Interakcemi mezi buňkami a produkcí cytokinů v cévní stěně Expresí pro a antiapoptotických proteinů v buňce (death receptory, protoonkogeny, supresorové geny).
.")
44
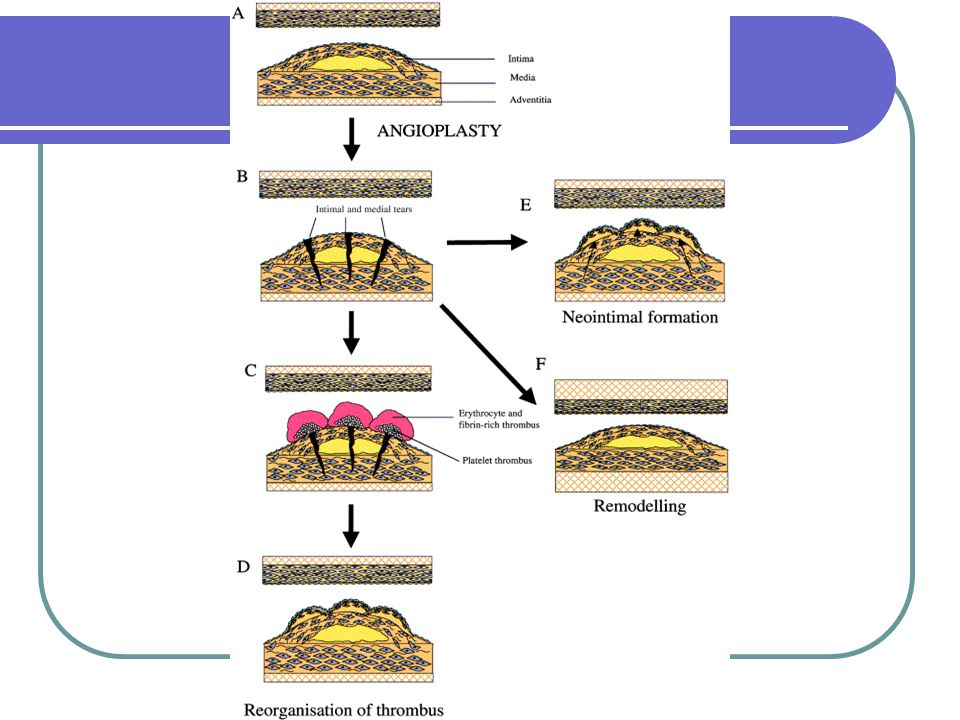
Mechanismy restenóz ve stentu
Distenze poškozené cévní stěny angioplastikoun nebo stentem způsobuje poškození endotelu, lamina elastica interna a media, často i adventicie. Rozšíření lumen je dáno kombinací redukce plaku (kompresí/embolizací), axiální redistribucí plaku v lezi, protlačením plaku a expanzí cévy.
, axiální redistribucí plaku v lezi, protlačením plaku a expanzí cévy.")
45
Mechanismy restenózy Restenóza je konečným produktem kombinacce biologických procesů, které všechny přispívají ke konečnému zúžení lumen postižené cévy. Mechanismy uplatněné při angioplastice a aplikaci stentu jsou podobné, proporce účinku jednotlivých faktorů je různá.

47
Mechanismy restenózy po angioplastice
Arteriální poškození po koronární intervenci vede ke změnám v cévní stěně. Angioplastika indukuje: Mechanické napětí cévy, které vede k ruptuře lamina elastica interna a k disekci médie Poškození až odstranění endotelu, což vede k expozici subendotelu cirkulujícím mitogenům (AT II, plasmin ...) Uvolnění mitogenů a cytokinů z destiček, endoteliálních buněk, hladkých svalových buněk cévní stěny a prozánětlivých buněk.
 Uvolnění mitogenů a cytokinů z destiček, endoteliálních buněk, hladkých svalových buněk cévní stěny a prozánětlivých buněk.")
49
Mechanismy restenózy po aplikaci stentu
V popředí je tvorba neointimy Proliferace hladkých asvalových buněk je dána chronickou přítomností stentu, která stimuluje proliferaci těchto buněk.

50
Děkuji za pozornost

Podobné prezentace





















 vylučované kůrou nadledvinek (aldosteron, kortisol); 2) vylučované pohlavními žlázami (progesteron, testosteron, estradiol)>")