Metabolismus sacharidů
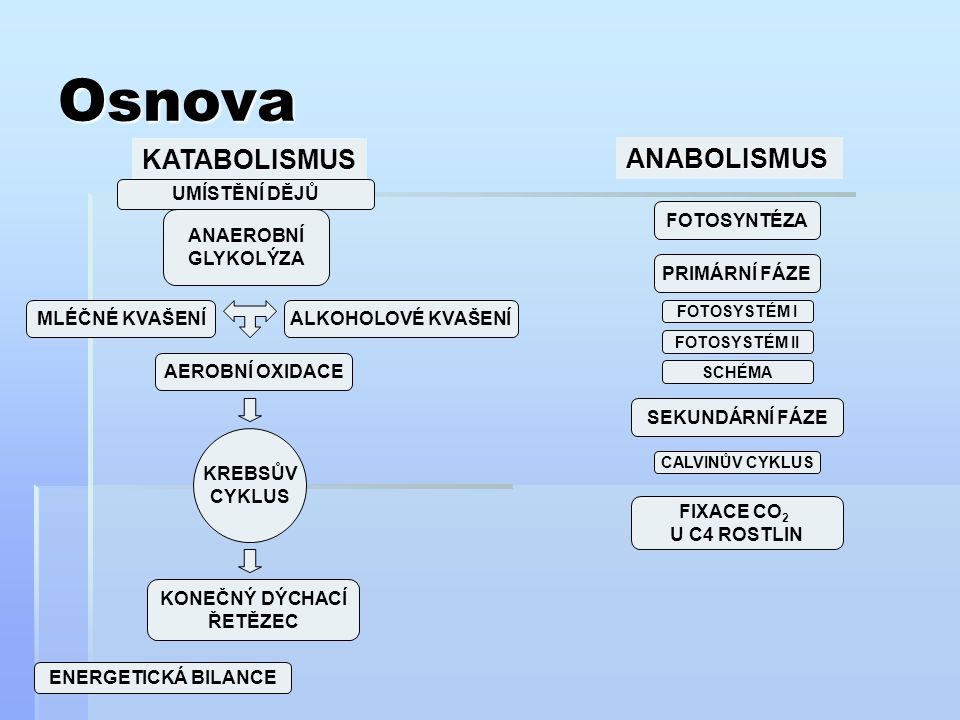
Osnova KATABOLISMUS ANABOLISMUS UMÍSTĚNÍ DĚJŮ FOTOSYNTÉZA ANAEROBNÍ GLYKOLÝZA PRIMÁRNÍ FÁZE MLÉČNÉ KVAŠENÍ ALKOHOLOVÉ KVAŠENÍ FOTOSYSTÉM I FOTOSYSTÉM II AEROBNÍ OXIDACE SCHÉMA SEKUNDÁRNÍ FÁZE KREBSŮV CYKLUS CALVINŮV CYKLUS FIXACE CO2 U C4 ROSTLIN KONEČNÝ DÝCHACÍ ŘETĚZEC ENERGETICKÁ BILANCE
Osnova KATABOLISMUS ANABOLISMUS UMÍSTĚNÍ DĚJŮ FOTOSYNTÉZA ANAEROBNÍ GLYKOLÝZA PRIMÁRNÍ FÁZE MLÉČNÉ KVAŠENÍ ALKOHOLOVÉ KVAŠENÍ FOTOSYSTÉM I FOTOSYSTÉM II AEROBNÍ OXIDACE SCHÉMA SEKUNDÁRNÍ FÁZE KREBSŮV CYKLUS CALVINŮV CYKLUS FIXACE CO2 U C4 ROSTLIN KONEČNÝ DÝCHACÍ ŘETĚZEC ENERGETICKÁ BILANCE
Katabolismus glukosy rozklad glukosy probíhá v několika na sebe navazujících dějích začíná anaerobní glykolýzou, jejímž výsledkem je pyruvát, který může být zpracován třemi způsoby: a) aerobně v Krebsově (citrátovém) cyklu pyruvát se před vstupem do Krebsova cyklu mění na Ac-CoA b) anaerobně mléčným kvašením c) anaerobně alkoholovým kvašením výsledkem těchto dějů je energie, která se ukládá do ATP a do redukovaných koenzymů (NADPH+H+ a FADH2) redukované koenzymy vstupují do dýchacího řetězce, kde z nich organismus získává další energii
Katabolismus glukosy (schéma) glukosa 2 ADP 2 Pi 2 NAD+ ANAEROBNÍ GLYKOLÝZA 2 NADH 2 ATP 2 pyruvát 2 NADH 2 NADH MLÉČNÉ KVAŠENÍ ALKOHOLOVÉ KVAŠENÍ 2 NAD+ 2 NAD+ AEROBNÍ OXIDACE 2 NAD+ 2 HS-CoA 2 laktát 2 ethanol 2 CO2 2 NADH 2 CO2 2 acetyl-CoA 6 NAD+ 2 FAD 2 GDP 2 Pi 2x citrátový cyklus KDE SE VZALO…? 10 NADH 2 FADH2 2 GTP 2 HS-CoA 4 CO2 6 NADH 2 FADH2 10 NADH 2 FADH2 34 ADP 34 Pi O2 KONEČNÝ DÝCHACÍ ŘETĚZEC H2O 10 NAD+ 2 FAD 34 ATP
Umístění dějů KATABOLICKÉ DĚJE SE ODEHRÁVAJÍ V CYTOPLAZMĚ BUŇKY A V MITOCHONDRIÍCH CYTOPLAZMA MITOCHONDRIE
Popis mitochondrie VNĚJŠÍ MEMBRÁNA VNĚJŠÍ KOMPARTMENT KRISTA VNITŘNÍ MEMBRÁNA MATRIX (VNITŘNÍ KOMPARTMENT)
Umístění dějů II CYTOPLAZMA MITOCHONDRIE GLUKOSA NADH+H+ PYRUVÁT AEROBNÍ OXIDACE MITOCHONDRIE PYRUVÁT KONEČNÝ DÝCHACÍ ŘETĚZEC H2O ANAEROBNÍ GLYKOLÝZA AcCoA ATP ATP ATP ATP NADH+H+ KREBSŮV CYKLUS NADH+H+ FADH2 GTP CO2
Anaerobní glykolýza O2 probíhá bez přístupu vzduchu evolučně se jedná o archaický děj, není cyklický!!! je málo energeticky výnosný probíhá u všech organismů probíhá v cytoplazmě, nejčastěji pak v buňkách příčně pruhovaného svalstva a v srdečním svalstvu děj řízený celou řadou enzymů vzniká málo ATP na úrovni substrátu (substrátová fosforylace) ATP ATP
Anaerobní glykolýza (souhrnná rovnice) glukosa + 2 NAD+ + 2ADP + 2 Pi vzniká rozpadem glukosy !!! ENERGIE!!! 2 NADH + 2 pyruvát + 2 ATP + 2 H2O + 4H+ redukovaný koenzym
Anaerobní glykolýza (popis) glukosa je fosforylována na glukosa-6-fosfát k tomu dochází proto, aby se glukose nepodařilo uniknout z buňky (stává se z ní iontová sloučenina) glukosa-6-fosfát se izomeruje na fruktosa-6-fosfát molekula fruktosa-6-fosfátu je symetričtější fruktosa-6-fosfát se fosforyluje na fruktosa-1,6-bisfosfát fruktosa-1,6-bisfosfát se štěpí na dva tříuhlíkaté štěpy dihydroxyacetonfosfát nepostupuje dále, dokud se nepřesmýkne na molekulu glyceraldehyd-3-fosfátu, tento jev slouží jako regulace anaerobní glykolýzy glyceraldehyd-3-fosfát je fosforylován na 1,3-bisfosfoglycerát z 1,3-bisfosfoglycerátu se uvolní fosfát za vzniku ATP a 3-fosfoglycerátu 3-fosfogylcerát se izomeruje na 2-fosfoglycerát, který je následně dehydratován na fosfoenolpyruát fosfoenolpyruvát ztrácí svůj fosfát za vzniku ATP a pyruvátu
Anaerobní glykolýza (schéma) OH O HO CH2O P OH O HO CH2OH GLUKOSA-6-FOSFÁT SE IZOMERUJE NA SYMETRIČTĚJŠÍ MOLEKULU FRUKTOSA-6-FOSFÁT ATP ADP IZOMERACE FOSFORYLACE GLUKOSAFOSFÁT- ISOMERASA GLUKOKINASA GLUKOSA GLUKOSA-6-FOSFÁT CHO HC–OH CH2O P CH2OH O – CH2 P OH HO O GLYCERALDEHYD-3-FOSFÁT ŠTĚPENÍ NA DVA TŘÍUHLÍKATÉ CUKRY TRIOSAFOSFÁTISOMERASA IZOMERACE FRUKTOSABISFOSFÁTALDOLASA FRUKTOSA-6-FOSFÁT CH2OH C=O CH2O P CH2 – O O – CH2 P OH HO O P ATP FOSFORYLACE 6-FOSFOFRUKTO- KINASA ADP DIHYDROXYACETONFOSFÁT FRUKTOSA-1,6-BIFOSFÁT
GLYCERALDEHYDFOSFÁT- Anaerobní glykolýza II (schéma) 2 NADH+H+ GLYCERALDEHYDFOSFÁT- DEHYDROGENASA COO HC–OH CH2O P 2 2 ATP P 2 ADP COO- HC–OH CH2O P 2 FOSFORYLACE FOSFOGLYCERÁTKINASA 2 NAD+ 1,3-BISFOSFOGLYCERÁT P 2 3-FOSFOGLYCERÁT CHO HC–OH CH2O P IZOMERACE FOSFOGLYCERÁTMUTASA COO- HC–O CH2OH P 2 GLYCERALDEHYD-3-FOSFÁT ENOLASA 2 ATP 2 ADP COO- C = O CH3 2 COO- C–O CH2 P 2 DEHYDRATACE 2-FOSFOGLYCERÁT PYRUVÁTKINASA 2 H2O PYRUVÁT FOSFOENOLPYRUVÁT
Anaerobní glykolýza (bilanční schéma) PYRUVÁT 2 GLUKOSA ANAEROBNÍ GLYKOLÝZA ATP ATP 2 NADH+H+
(anion kyseliny pyrohroznové) (anion kyseliny mléčné) Osudy pyruvátu I za anaerobních podmínek se pyruvát přeměňuje na laktát – MLÉČNÉ KVAŠENÍ NADH+H+ NAD+ COO- C = O CH3 COO- H – C – OH CH3 RED. LAKTÁTDEHYDROGENASA pyruvát (anion kyseliny pyrohroznové) laktát (anion kyseliny mléčné) Laktát vzniká při velké/nadměrné fyzické námaze. Protože se jedná o kyselinu, dochází k okyselování cytoplazmy svalových buněk, což se projevuje bolestí svalů. Při relaxaci (svalů) dochází k vyplavování laktátu do krve, kterou se dostává do jater, kde je resyntetizován na glukosu. Tento cyklus laktátu nazýváme Coriho cyklus.
Osudy pyruvátu II za anaerobních podmínek může dojít k alkoholovému kvašení COO- C = O CH3 C CH3 H O NADH+H+ NAD+ - CO2 CH2OH CH3 PYRUVÁTDEKARBOXYLASA ALKOHOLDEHYDROGENASA ETHANOL PYRUVÁT ACETALDEHYD Alkoholové kvašení se využívá při výrobě alkoholických nápojů (pivo, víno). Kvasinky jsou schopné žít v maximálně 15% alkoholu (pak se pro ně stává jedem) Silnější alkohol je nutné připravovat destilací.
Metabolismus alkoholu alkohol (ethanol) je postupně oxidován na acetaldehyd a následně na kyselinu octovou kyselina octová se buď spontánně štěpí na CO2 a H2O, nebo vstupuje v podobě acetylkoenzymuA do Krebsova cyklu z 1 molekuly ethanolu získáme energii 15 ATP C CH3 H O C CH3 OH O CH2OH CH3 NAD+ NADH+H+ ALKOHOLDEHYDROGENASA ACETALDEHYDOXIDASA ETHANOL ACETALDEHYD KYSELINA OCTOVÁ
Osudy pyruvátu III Aerobní odbourávání pyruvátu Dochází k tzv. oxidativní dekarboxylaci MAKROERGICKÁ VAZBA NAD+ NADH+H+ COO- C = O CH3 CH3 – C SCoA O + CO2 PYRUVÁTDEKARBOXYLASA AKTIVOVANÁ KYSELINA OCTOVÁ PYRUVÁT Tato reakce probíhá v cytoplazmě buňky. Aktivovaná kyselina octová neboli acetylkoenzymA se přenáší přes mitochondriální membránu a v mitochondriích vstupuje do Krebsova cyklu Ac-CoA obsahuje makroergní vazbu, je to tedy makroergická sloučenina
Krebsův cyklus (úvod) Krebsův cyklus má mnoho názvů: citrátový cyklus cyklus trikarboxylových kyselin cyklus kyseliny citrónové TCA cyklus Probíhá v matrix mitochondrie. Popis: jedná se o cyklický děj (může být kdykoliv přerušen, podle toho, jaké látky tělo zrovna potřebuje) dochází ke dvěma dekarboxylačním reakcím – oxidační reakce (produkce redukovaných koenzymů 1xFADH2 a 3xNADH+H+, které dále vstupují do dýchacího řetězce)
Krebsův cyklus (schéma) NAD+ CoASH CH3COSCoA NADH+H+ CITRÁT ISOCITRÁT CO2 H2O OXALACETÁT 2-OXOGLUTARÁT NADH+H+ NAD+ CoASH NADH+H+ NAD+ CO2 L-MALÁT SUKCINYLKOENZYM A FADH2 FAD GDP + P GTP H2O FUMARÁT SUKCINÁT CoASH
Krebsův cyklus (bilanční schéma) H2O CH3COSCoA KOENZYMY (oxidované) KREBSŮV CYKLUS GTP CO2 REDUKOVANÉ KOENZYMY
Konečný dýchací řetězec jedná se o „zakončení“ katabolické dráhy glukosy (i jiných sloučenin) jedná se o aerobní děj vstupuje do něj kyslík a redukované koenzymy, probíhá na vnitřní mitochondriální membráně elektronový transportního řetězec Enzym ATP-synthasa O2 NADH+H+ ELEKTRONOVÝ TRANSPORTNÍ SYSTÉM H2O FADH2 ATP- synthasa ATP ATP ATP
Konečný dýchací řetězec (průběh) redukované koenzymy se před vstupem do konečného dýchacího řetězce rozpadají dle rovnic: NADH+H+ → NAD+ + 2 H+ + 2e- FADH2 → FAD + 2H+ + 2e- elektrony, mající vysoký energetický potenciál, jsou této energie postupně zbavovány přechodem přes elektronový transportní řetězec (cytochromy…) ten je energie zbavuje postupně (kdyby elektrony vydaly všechnu svou energii zároveň, znamenalo by to smrt buňky) vodíkové kationty se dostávají ven přes mitochondriální membránu a při svém návratu dodávají energii enzymu ATP-syntasa, který ji váže do vazeb ATP
Konečný dýchací řetězec (schéma) NADH+H+ a FADH2 se dostanou k mitochondriální membráně a dojde k jejich rozkladu. ELEKTRONY procházejí elektronovým transportním řetězcem a nakonec se spojí s 2H+ a O2 za vzniku vody. VODÍKOVÉ KATIONTY se dostávají na vnější stranu membrány a při návratu roztáčí ATP-synthasu. 4H+ 6H+ 8H+ 12H+ cyt.a cyt.a3 „Cu2+“ IV cyt. b cyt. c1 Fe2S2 III I FMN FMNH2 Fe4S4 cyt. c CoQ i CoQH2 FAD FADH2 II MITOCHONDRIÁLNÍ MEMBRÁNA 2e- ATP-synthasa 2e- 2H+ 2H+ 2H+ 1/2O2 4H+ 2H+ ATP 2e- 2H+ NADH+H+ NAD+ H2O ADP+P ATP FADH2 FAD
Proč se H+ vrací? kationty H+ v konečném dýchacím řetězci procházejí vnitřní mitochondriální membránou a hromadí se ve vnějším kompartmentu VNĚJŠÍ KOMPARTMENT VNITŘNÍ MIT. MEMBRÁNA S ATP-SYNTHASOU MATRIX nahromadění H+ ve vnějším kompartmentu v něm způsobuje zvýšení koncentrace H+ (a tím i elektrického gradientu) H+ se vrací zpět do matrix proto, aby vyrovnaly rozdíl koncentrací (a tím i elektrického gradientu) Nezapomeňme, že při tomto ději vzniká ATP!!!
Energetická bilance energetická bilance nám ukáže energetický výtěžek všech výše popsaných cyklů při jejím sestavení vycházíme z toho, že: 1x NADH+H+ … … … 3 ATP 1x FADH2 … … … 2 ATP 1x GTP … … … 1 ATP Bilance na jednu molekulu glukosy: anaerobní glykolýza: spotřeba: 2 ATP zisk: 4 ATP + 2 NADH+H+ = 10 ATP celkem: 8 ATP oxidativní dekarboxylace pyruvátu (2x): 2 NADH+H+ = 6 ATP Krebsův cyklus (2x) = 6 NADH+H+ + 2 FADH2 + 2 GTP = 24 ATP Celková bilance na molekulu glukosy: 38 ATP
38, ale… v učebnicích biochemie a chemie se setkáváme s různými hodnotami zisku ATP na molekululu glukosy: 32 36 38 … 38 ATP je největší možný teoretický výtěžek 36 ATP započítává spotřebu 2 ATP, při přenosu dvou molekul NADH+H+ z cytoplazmy do mitochondrie přes membránový člunek (o kterém mnoho lidí neví) ve skutečnosti (v živé buňce) vzniká něco okolo 30 ATP
Anabolismus sacharidů Podle zdroje uhlíku pro tvorbu sacharidů rozlišujeme: Autotrofní organismy (litotrofní) schopnost tvorby sacharidů z jednoduchých anorganických látek (CO2)→ fotosyntéza zelené rostliny, sinice, některé řasy… Heterotrofní organismy (organotrofní) využívají pro tvorbu sacharidů 3-4 uhlíkové organické sloučeniny vzniklé v průběhu katabolismu: laktát glycerol meziprodukty Krebsova cyklu (pyruvát…) většina organismů Podrobnější rozdělení
Rozdělení organismů (podle zdroje E) ve vztahu ke zdroji energie rozlišujeme organismy: Organismy fototrofní využívají energii slunečního záření FOTOAUTOTROFNÍ (=FOTOLITOTROFNÍ) ENERGIE ZE SLUNCE, UHLÍK Z ANORGANICKÝCH LÁTEK FOTOHETEROTROFNÍ (=FOTOORGANOTROFNÍ) ENERGIE ZE SLUNCE, UHLÍK Z ORGANICKÝCH LÁTEK Organismy chemotrofní Využívají energii chemických vazeb CHEMOAUTOTROFNÍ (=CHEMOLITOTROFNÍ) ENERGIE Z CHEM.VAZEB, UHLÍK Z ANORGANICKÝCH LÁTEK CHEMOHETEROTROFNÍ (=CHEMOORGANOTROFNÍ) ENERGIE Z CHEM.VAZEB, UHLÍK Z ORGANICKÝCH LÁTEK
Fotosyntéza Kdo? vyšší rostliny, zelené a hnědé řasy, sinice, purpurové a zelené bakterie Co ji umožňuje? přítomnost fotoreceptorů (=pigmentů absorbujících záření) většinou se využívají chlorofyly a,b jejichž činnost doplňují karotenoidy Co to je? z hlediska syntézy glukosy anabolický děj více viz. fotosynéza II
Chlorofyl a,b
Karotenoidy
Fotosyntéza II Fotosyntéza je soubor chemických reakcí, v jejichž průběhu dochází k pohlcování energie slunečního záření, která je využita k přeměně jednoduchých anorganických sloučenin na látky organické. Fyzikální hledisko: přeměna energie slunečního záření na energii chemickou Chemické hledisko: převedení uhlíku z oxidačního čísla IV (nízký obsah energie) na redukovaný materiál s vysokým obsahem energie (sacharidy) ATP ATP +TEPLO FOTOSYNTÉZA ATP E CO2 REDUKCE sacharidy E
Souhrnná rovnice fotosyntézy ZÁKLADNÍ ROVNICE: ☼ 6 CO2 + 12 H2O C6H12O6 + 6 O2 + 6 H2O ZJEDNODUŠENĚ: 6 CO2 + 6 H2O C6H12O6 ☼ ΔG0 = 2826 kJ.mol-1 ΔG0 – Gibbsova energie
Fotosyntéza bez vody nižší fotosyntetyzující organismy užívají jako redukční činidlo místo vody sulfan, vodík, nebo organické kyseliny O H H+ e– H+ e– ½O OH O R C S H H SULFAN VODÍK KARBOXYLOVÁ KYSELINA neprodukují tedy kyslík
Průběh fotosyntézy Fotosyntéza probíhá ve dvou oddělených, ale na sebe navazujících fázích fáze primární (světelná) přítomnost světla vzniká při ní ATP a NADPH+H+ ATP slouží v další fázi jako zdroj energie NADPH+H+ slouží jako zdroj vodíkových protonů uvolňuje se kyslík fáze sekundární (temnostní) dochází během ní k syntéze glukózy z CO2 ATP ATP H+ NADP O2 O2 O2 glukosa
Primární (světelná) fáze všechny reakce jsou podmíněny adsorpcí slunečního záření chlorofylovými molekulami CHLOROPLAST molekula chlorofylu umožňuje přeměnu energie fotonů slunečního záření na energii excitovaných elektronů tato energie je využita při syntéze ATP a NADH+H+ primární fáze probíhá za účasti dvou fotosystémů (pouze u vyšších rostlin)
Fotosystém I fotosystém I obsahuje dlouhovlnější formy chlorofylu a s adsorpcí světla do 700 nm (proto označení P700) po adsorpci světelného kvanta dojde k redukci oxidačněredukčního potenciálu a k uvolnění elektronů, které se přesouvají na dosud neznámý akceptor Z a odtud: se přesouvají na NADP+ a redukují jej na NADPH+H+ (zdroj H+ je fotolýza vody) se můžou vrátit zpět na P700, přičemž část jejich energie se využije na tvorbu ATP (=cyklická fosforylace) Z NADP+ NADPH+H+ ADP 2h.ν 2H+ ATP P700 H2O FOTOLÝZA VODY 2 e- ½ O2
Fotosystém II fotosystém II obsahuje krátkovlnější formy chlorofylu a, maximální délka přijatého záření je 680 nm (odtud P680) po adsorpci světelného kvanta dojde k uvolnění elektronů, které: nejprve redukují systém Q pak přes plastochinon (nebo plastokyanin, či cytochrom f) přechází na fotosystém I a nahrazují elektrony, které byly použity v předchozích reakcích (=necyklická fosforylace); vzniká ATP fotosystém II se sám navrátí do původního stavu po přijetí elektronů vzniklých při fotolýze vody PLASTOCHINON Q P700 ADP 2H+ ATP 2h.ν P680 H2O FOTOLÝZA VODY 2 e- ½ O2
Primární fáze (schéma) FOTOSYSTÉM I NEBO ELEKTRONY „PODSTOUPÍ“ CYKLICKOU FOSFORYLACI NADP+ Z ELEKTRONY SE UPLATNÍ PŘI TVORBĚ NADPH+H+ NADPH+H+ 2 e- Q VZNIKÁ ATP 2h.ν ADP PO PŘIJETÍ ENERGIE SE UVOLŇUJÍ ELEKTRONY 2H+ P700 2 e- ATP ELEKTRONY UMOŽNÍ NÁVRAT FOTOSYSTÉMU I DO PŮVODNÍHO STAVU 2h.ν H2O PRŮBĚH PRIMÁRNÍ FÁZE DOPLŇUJE FOTOLÝZA VODY P680 2 e- ½ O2 PO PŘIJETÍ ENERGIE SE UVOLŇUJÍ ELEKTRONY FOTOLÝZA VODY FOTOSYSTÉM II
Primární fáze (poznámky) Cyklická x necyklická fosforylace při obou dějích vzniká ATP cyklická = elektrony poskytuje P700 a vrací se do P700 (cyklus) necyklická = elektrony poskytuje P680 a putují do P700 Fotolýza vody rovnicí ji lze vyjádřit: H2O → 2 H+ + 2e- + ½ O2 Vznik kyslíku a NADPH+H+ při cyklické fosforylaci se neuplatňuje voda → nevzniká kyslík, nevzniká NADPH+H+ při necyklické fosforylaci se uplatňuje voda → vzniká kyslík, vzniká NADPH+H+
Sekundární fáze nevyžaduje světelnou energii dochází k biosyntéze sacharidů z CO2 za využití: ATP (zdroj energie) NADPH+H+ (redukční činidlo) biosyntéza sacharidů se odehrává v kapalné části chloroplastu (stroma) a v cytosolu realizuje ji několik metabolických cest – nejznámější je Calvinův cyklus CO2 Calvinův cyklus produkce NADPH+H+ fotolýza vody necyklická a cyklická fosforylace H2O ATP ½ O2 HEXOSY PRIMÁRNÍ FÁZE SEKUNDÁRNÍ FÁZE
Calvinův cyklus (popis) Lze rozlišit tři fáze fixace CO2 v organické formě redukce aktivovaného CO2 za vzniku hexosy regenerace akceptoru CO2 CO2 se navazuje na molekulu ribulosa-1,5-bifosfátu za vzniku nestabilního meziproduktu (6 C), který se ihned rozpadá na 3-fosfogylcerát 3-fosfoglycerát je fosforylován pomocí ATP – vzniká 1,3-bisfosfoglycerát a ten je následně redukován pomocí NADPH+H+ na glyceraldehyd-3-fosfát: část glyceraldehyd-3-fosfátu se kondenzuje za vzniku fruktosa-1,6-bifosfátu (ten se mění na glukosa-6-fosfát) druhá část se mění na ribulosa-1,5-bifosfát, který umožňuje fixaci dalšího CO2
Calvinův cyklus (schéma) VZNIKÁ 6 molekul CO2 NA RIBULOSA-1,5-BIFOSFÁT SE NAVÁŽE CO2 ZE VZDUCHU nestabilní meziprodukt obsahující 6 atomů uhlíku 6 P ribulosa-1,5-bifosfát TEN SE ROZPADÁ NA: P 12 3-fosfoglycerát ČÁST OBNOVUJE RIBULOSA-1,5-BIFOSFÁT 6 ADP 6 ATP 12 NADPH+H+ 12 ATP PROBĚHNE FOSFORYLACE POMOCÍ ATP A REDUKCE POMOCÍ NADPH+H+ 4 Pi 12 NADP+ 12 ADP+Pi P 10 glyceraldehyd-3-fosfát VZNIKÁ: P 12 glyceraldehyd-3-fosfát P glukosa-6-fosfát P 2 glyceraldehyd-3-fosfát ČÁST KONDEZUJE ZA VZNIKU GLUKOSA-6-FOSFÁTU
C3 rostliny C3 rostliny jsou takové, které využívají Calvinův cyklus jedná se o většinu rostlin a řas název „C3 rostliny“ byl zvolen proto, že první produkt asimilace oxidu uhličitého je tříuhlíkatá sloučenina 3-fosfoglycerát COO- HC–OH CH2O P 3-FOSFOGLYCERÁT
C4 rostliny př. kukuřice, cuktrová třtina, plevele… primárním akceptorem je fosfoenolpyruvát a meziprodukt je oxalacetát (4 uhlíkatá sloučenina = C4) oxalacetát se redukuje na malát (kyselina jablečná), který se rozkládá na CO2 a pyruvát tímto způsobem se v místě Calvinova cyklu vytváří vysoká koncentrace CO2 a to umožňuje velkou rychlost a účinnost fotosyntézy
Fixace CO2 u C4 rostlin Calvinův cyklus CO2 ze vzduchu oxalacetát OXALACETÁT SE REDUKUJE NA: VZNIKÁ: MALÁT SE ROZPADÁ NA: CO2 ze vzduchu CO2 VSTUPUJE DO CALVINOVA CYKLU oxalacetát malát Calvinův cyklus FOSFOENOLPYRUVÁT NA SEBE VÁŽE CO2 CO2 fosfoenolpyruvát pyruvát ZMĚNA PYRUVÁTU VZNIKÁ: glukosa-6-fosfát