Mechorosty pro Botaniku vyšších rostlin M
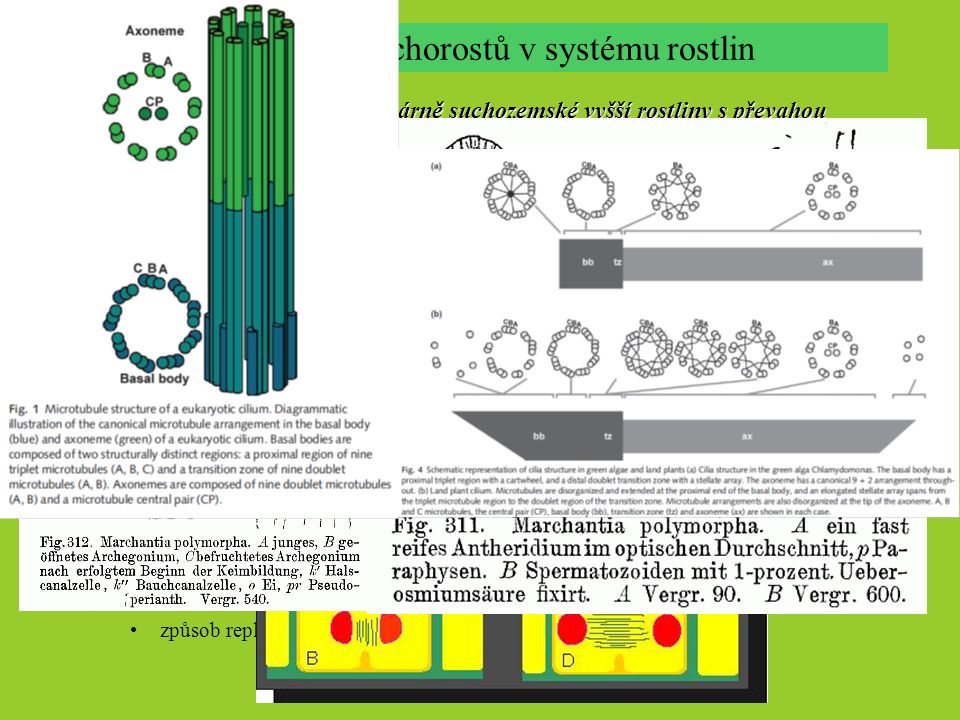
Postavení mechorostů v systému rostlin mechorosty – zelené, primárně suchozemské vyšší rostliny s převahou autotrofního gametofytu v životním cyklu, tvořící nevětvená sporangia Mechorosty součástí zelených rostlin („Viridiplantae“, tj. zelené řasy + vyšší r.): kombinace fotosyntetických barviv (chlorofyl a+b, α- a β-karoten, ztráta fykobilinů) stavba chloroplastu škrob jako zásobní látka ultrastruktura bazálního aparátu bičíků u pohyblivých buněk Streptophyta (Charophyceae s.l., Zygnematophycae, Embryophyta): fragmoplast – mikrotubuly vřeténka kolmo ke tvořící se buněčné stěně Mechorosty součástí vyšších rostlin (suchozemských, Embryophyta) tvorba embrya mnohobuněčná gametangia s vícevrstevnými sterilními obaly tvorba kutikuly, tlustostěnných spor s tetrádovým švem apikální meristematická buňka s >2 oddělujícími hranami prodlužování sporofytu po sporogenezi sporopolenin ve stěně spor způsob replikace centrozómů, ultrastruktura baz. tělísek a MLS
Společné znaky mechorostů Goremykin & Hellwig (2005; chloroplast. genom) Groth-Malonek & al. (2005; mitochondr. introny), Qiu & al. (2007,8; 7 cp+mt+nr genů) Společné znaky mechorostů Společné znaky mechorostů, odlišení ostatních vyš. rostlin (Polysporangiophyta) charakteristický životní cyklus; převaha gametofytu a jeho morfologická a anatomická diverzifikace krátkověkost sporofytu a jeho trofická závislost na gametofytu sporofyt nevětvený s jediným terminálním sporangiem absence ligninu gametofyt často tvořen fází protonematu a gametoforu gametofor přirůstá činností tzv. apikální buňky, ne meristematického pletiva patrně parafyletická skupina organismů, tři nejspíše monofyletické skupiny s dosud nejasnými vzájemnými vztahy: játrovky (Marchantiophyta), mechy (Bryophyta), hlevíky (Anthocerotophyta) patří mezi nejstarší suchozemské rostliny, první ± jasné fosilie (játrovek, Metzgeriothallus sharonae, Hepaticites devonicus) ze střed. a svrchního devonu, ca. 360-390 mil. let staré, ale ve skutečnosti jistě starší, možné spory játrovek už z ordoviku (475 mya), předchůdci pravděpodobně Charales) Mechorosty dnes: 15-20 tisíc druhů celosvětově, většina tropická, rozšířeny však téměř ve všech ekosystémech s výjimkou moře (Evropa ca. 1700 druhů) vzhledem k fyziologickým limitacím ve většině ekosystémů minoritní složkou s výjimkou rašelinišť; často však mezi prvními kolonizátory nově vzniklých substrátů
Játrovky (Marchantiophyta) monofyletická, velmi diverzifikovaná skupina mechorostů s asi 5-9 tis. druhy v asi 300-400 rodech. Nejstarší známé fosílie (střední devon, ale asi ještě starší). Olistěné i lupenité gametofory někdy komplexní stavby, sporofyty efemerní, jednoduše stavěné Hlavní společné znaky játrovek protonema redukované na 2-3 buňky v jednořadém klíč. vláknu; na protonematu diferencována jediná iniciála jediný gametofor gametofor lupenitý nebo listnatý, listy ve 2-3 řadách, obvykle jednovrstevné, bez diferenciace žebra jako svazku výztužných a vodivých pletiv, často laločnaté buňky gf. často isodiametrické a s rohovými ztluštěninami, obsahují (ca. 90%) siličná tělíska (membránové organely, terpenoidy, vznik z ER) sporofyty (noha, štět, výtrusnice) dozrávají uvnitř tkáně gametofytu, sporogenní tkáň z endothecia štět hyalinní, prodlužování štětu rychlé, pouze elongací jeho buněk, až po diferenciaci a dozrání sf., čepička protržena a zůstává na bázi sf. kolumela chybí, avšak obvykle přítomny sterilní buňky – elatery s helik. ztluštěninami; stěna sporangia bez průduchů, s nápadnými ztluštěninami, tmavá pigmentace; dozrání spor synchronní, otvírání sporofytu obv. 4 chlopněmi, někdy 1 chlopní, nepravid. rozpadem, víčkem Systém: bazální malé skupiny (Haplomitriopsida; <20 druhů), dvě velké třídy – Marchantiopsida (~450), Jungermanniopsida (~ 5000)
Játrovky: třída Marchantiopsida převážně lupenité játrovky s tendencí k tvorbě komplexní lupenité stélky (diferenciace pletiv); apikální buňka s obvykle 2 oddělujícími plochami (někdy 4); sporocyty před meiózou nelaločnaté, tendence k redukci sporofytu (jednovrstevná stěna, krátký až chybějící štět …); často xeromorfické adaptace Největší skupina třídy – Marchantiales vždy lupenité játrovky se zpravidla komplexní stavbou stélky stélka zpravidla diferencována ve svrchní fotosyntetickou část s dýchacími póry, ústícími do komůrek a spodní část s parenchymatickým zásobním pletivem rhizoidy často dvojího druhu, část s charakter. zluštěninami ve stěnách buňky gametoforu částečně silnostěnné, některé (bez chloroplastů) obsahují po 1 velkém siličném tělísku, ve stěnách anthokyanová barviva gametangia (zejména ♀) často vyrůstají na specializované větvi stélky – receptakulu (archegoniofory karpocephalum, antheridiofory), na karpocephalech tvorba zpravidla většího počtu sporofytů štět krátký až zcela redukovaný stěna sporangia jednovrstevná Kosmopolitní rozšíření, hodně subtropických druhů, obecně často extrémní podmínky, suchomilné. Marchantia, Riccia, ca. 400 druhů v ± 28 rodech a 12-15 čeledích
třída Jungermanniopsida: obecná char., lupenité typy listnaté i lupenité játrovky s jednoduchou stavbou stélky; apikální buňka často se 3 oddělujícími plochami; rhizoidy pouze hladké; buňky s velkým počtem siličných tělísek; sporocyty před meiózou laločnaté, sporofyty dobře vyvinuté (vícevrstevná stěna sporangia, dlouhý štět …); rostliny vlhkomilné, obvykle nesnášející vyschnutí Nejpočetnější skupina játrovek, ca. 5000 druhů ve > 300 rodech Převážně lupenité typy – podtřídy Peliidae a Metzgeriidae apikální buňka obvykle čočkovitá, 2 oddělující plochy lupenitá, zploštělá stélka, obvykle nepravidelně pseudodichotomicky se větvící stavba stélky obvykle homogenní, ale někdy diferencována jednoduchá vodivá pletiva (hydroidy; čeledi Pallaviciniaceae, Hymenophytaceae) archegonia nejsou terminální sporofyty vyrůstají pod vrcholem (anakrogynní) parafyletická skupina poměrně izolovaných linií, většina zástupců tropických, 500 druhů. Evropa: nejvýznamnější Pelliaceae (Pellia), Metzgeriaceae (Metzgeria). Přechod k listnatým typům – např. Fossombronia
Jungermanniopsida: listnaté typy – Jungermanniidae Převážně listnaté typy – podtřída Jungermanniidae apikální buňka obvykle tetraedrická, 3 oddělující plochy listnatá stélka s hlavní osou a dvěma postranními + 1 ventrální řadou listů listy obvykle jednovrstevné, laločnaté (typicky 2 laloky), buňky isodiametrické, s rohovými ztluštěninami, chloroplasty a siličná tělíska ve všech buňkách stavba stélky homogenní, vodivá pletiva chybí archegonia jsou terminální sporofyty vyrůstají na vrcholu (akrogynní) kolem archegonií a vyvíjejícího se sporofytu periant – válcovitý útvar list. původu sporofyt dobře vyvinutý, na obvykle dlouhém štětu, otvírání sporangia 4 chlopněmi zřejmě monofyletická skupina, několik řádů, většina zástupců tropických, téměř všechny druhy třídy patří sem Evropa: největší čeledi Lophoziaceae, Scapaniaceae Tropické oblasti: největší čeledi Plagiochilaceae, Lejeuneaceae, Jubulaceae
Mechy – oddělení Bryophyta největší skupina mechorostů s 10 tis. druhy v asi 700 rodech. Nejstarší známé fosílie z permu. Olistěné gametofory obvykle komplexní stavby, sporofyty s výztužnými (+ někdy vodivými) pletivy Hlavní společné znaky mechů protonema dobře vyvinuto, větvené; tvorba většího množství gametoforů rhizoidy mnohobuněčné gametofor listnatý, listy nelaločnaté, často mnohovrstevné v oblasti středního žebra (vodivá+výztužná pletiva) buňky listů obvykle rovnoměrně ztlustlé, často pigmentované stěny; siličná tělíska chybí štět sporangia se prodlužuje před dozrátím sporangia, kryt čepičkou; není efemerní uvnitř sporangia střední sloupek (kolumela), jiné sterilní buňky (elatery, pseudoelatery …) chybí; stěna mnohovrstevná, s průduchy spory dozrávají najednou, ale vypadávají ze sporangia postupně díky existenci obústí (peristomu) – věnec hygroskopických zubů, důležitý syst. znak Systém: na bázi asi 7 izolovaných skupin (dnes rozeznávaných jako třídy), většina mechů v nejodvozenější třídě Bryopsida
Třída Sphagnopsida – rašeliníky malá skupina mechorostů s asi 150 druhy ve 2 rodech, rozšířeny celosvětově Hlavní znaky protonema dvojího druhu – vláknité a lupenité gametofor – vzpřímená, spirálně olistěná lodyžka se svazečky větví, na vrcholu větve nahloučené v kapitulum; rhizoidy u dosp. gametoforů chybí listy dimorfické (lodyžní × větevní), jednovrstevné, bez středního žebra anatomická stavba listů: hyalocyty + chlorocyty (1:2), hyalocyty velké, přepážkované, s póry stavba lodyžek a větví: střední válec parenchymatického pletiva – sklerodermis – hyalodermis sporofyt vyrůstá na zvláštní větvi lodyžky připomínající štět – pseudopodiu; tvořen pouze nohou a vlastním sporangiem (štět chybí) uvnitř sporangia mohutná kolumela překlenutá sporogenní tkání; ve stěně nejsou průduchy otvírání sporangia víčkem, explozivně; peristom chybí ekologicky významná skupina, základ biomasy rašelinišť; absorpční kapacita; antibiotické účinky; rašelina
Třídy Takakiopsida, Andreaeopsida – štěrbovky malé skupiny mechorostů (Takakiopsida – 2 druhy, Andreaeopsida – asi 100 druhů ve 2 rodech, zejména v horských oblastech, více na již. polokouli; v Evropě pouze Andreaea (ca. 10 druhů) Hlavní znaky třídy Andreaeopsida gametofor – vzpřímená, drobná, spirálně olistěná lodyžka; často výrazná hnědočervená až černá pigmentace lodyžky bez vodivých pletiv, ± homogenní stavby listy se středním žebrem nebo bez něj; čepel ze silnostěnných, isodiametrických buněk, na bázi někdy prodloužených sporofyt vyrůstá jako u rašeliníků na pseudopodiu, tvořen pouze nohou a vlastním sporangiem (štět chybí) stavba sporangia velmi podobná rašeliníkům otvírání sporangia štěrbinami, na vrcholu jsou chlopně obvykle spojené; peristom chybí striktně epilitické horské mechy kyselých substrátů
Třída Polytrichopsida – ploníky menší skupina mechorostů v jediné čeledi s asi 23 rody a 200 druhy; mechy s nejrozvinutějším gametofytem vůbec Hlavní znaky protonema dobře vyvinuté, vláknité gametofor – vzpřímená, i > 50 cm vysoká, spirálně olistěná lodyžka nesoucí terminální sporofyt lodyžky s obvykle výrazně vyvinutými vodivými pletivy; svazek protostélické stavby – uprostřed s elementy odpovídajícími dřevní části (hydroidy + výztužná pletiva), obkroužený „lýkovou částí“ – leptomem (leptoidy, vedoucí asimiláty + průvodní parenchym. buňky) listy dlouze kopinaté se širokým středním žebrem, v žebru podpůrné + vodivé buňky, rudimentární list. stopy; výrazná epidermis, ventrálně fotosyntetické lišty, voskovitá kutikula. U některých rodů sekundární redukce stavba sporofytu odpovídá většině ostatních mechů (třídy Bryopsida) – tvořen nohou, štětem a vlastním sporangiem (tobolkou), ale i ve štětu leptoidy uvnitř sporangia kolumela, sporogenní tkáň a vzdušné komůrky; ve stěně funkční průduchy otvírání sporangia víčkem; peristom vyvinutý, nematodontního typu (zuby z celých buněk horní části sporangia; nehygroskopické, přirostlé k rozšířené části kolumely – epifragmě) kosmopolitně rozšířená skupina, obvykle nápadné, velké mechy vlhkých i vysychavých substrátů. Na sev. polokouli nejvýraznější ploníky (Polytrichum, Polytrichastrum…), v paleotropické oblasti Dawsonia
Třída Bryopsida – pravé mechy největší a nejdiverzifikovanější skupina mechorostů; ca. 20 řádů, 100 čeledí, > 600 rodů a kolem 10 tis. druhů; kosmopolitní rozšíření, ve většině suchozemských biotopů Hlavní znaky protonema dobře vyvinuté, vláknité gametofor – vzpřímená nebo poléhavá, někdy až zpeřeně větvená olistěná lodyžka, nesoucí terminální nebo laterálně vyrůstající sporofyt(y); uchycení v substrátu rhizoidy (někdy tvořícími gemy) lodyžky někdy s vyvinutými vodivými pletivy; ve svazku pouze hydroidy listy různých tvarů, často se středním žebrem, v žebru podpůrné + vodivé buňky; epidermis, někdy voskovitá kutikula. Buňky čepele často prodloužené sporofyt tvořen nohou, štětem a vlastním sporangiem; umístění terminální (akrokarpní mechy) nebo laterální (klado- a pleurokarpní mechy) uvnitř sporangia kolumela a sporogenní tkáň; ve stěně často průduchy otvírání sporangia víčkem; peristom často vyvinutý (někdy sek. redukce), arthrodontního typu (zuby ze zbytků bun. přepážek horní části sporangia; hygroskopické, uzavírání sporangia po odpadu víčka) spory opouštějí sporangium postupně
Bryopsida: přehled systému asi 8 podtříd, z nichž nejdůležitější: Podtřída Funariidae vzpřímeně rostoucí, akrokarpní mechy s dvěma kruhy obústí (exostom, endostom), zuby obou kruhů v zákrytu několik morfologicky odlišných řádů, největší Funariales: často efemerní mechy s jednoduše stavěným gametoforem (buňky listů téměř bez diferenciace, slabě protažené, tenkostěnné, vodivá pletiva ± nevyvinuta) a mohutným sporofytem. Kolonizátoři nových substrátů, často disturbované biotopy. Funaria hygrometrica. Podtřída Dicranidae vzpřímeně rostoucí, obvykle vrcholoplodé mechy s jediným kruhem obústních zubů (endostom) gametofory značně redukované, listy obvykle velmi úzké tendence k redukci obústí velká skupina mechů (>3000 druhů), řády Dicranales, Pottiales, Grimmiales ad. Spíše v mimotropických oblastech, často extrémní biotopy (aridní, polární, vysokohorské, striktní epiliti …).
Bryopsida: přehled systému Podtřída Bryidae obv. vzpřímeně rostoucí, vrcholoplodé mechy s typicky dvěma dobře vyvinutými kruhy obústí; zuby exo- a endostomu alternují gametofory velmi různé, svazek vodivých pletiv (hydroidů) často dobře vyvinutý; 6 řádů řád Splachnales – koprofilní mechy s gametofyty podobnými Funariales, sporofyty přizpůsobené entomofilii řád Bryales – obv. drobnější akrokarpní mechy s dobře vyvinutými vod. pletivy, buňky listů často rhombické, tobolky často převisle hruškovité Podtřída Hypnidae obv. poléhavé, větvené (i několikrát zpeřeně), pleurokarpní mechy sporofyt velmi podobný Bryidae gametofory velmi různé, svazek vodivých pletiv (hydroidů) často přítomen, listy často s redukovaným středním žebrem; 3 řády řád Hypnales – tropické i mimotropické mechy; typicky velmi prodloužené buňky listů, diferenciace křídelních buněk, tvorba parafylií na lodyžkách
Hlevíky (Anthocerotophyta) izolovaná evoluční linie s nejasným vztahem k ostatním mechorostům a zbytku vyšších rostlin; celosvětově asi 100 – 150 (–300?) druhů v 6 – 12 rodech Hlavní společné znaky stélka lupenitá, dorziventálně zploštělá, růžicovitá nebo pentlicovitá stélka z tenkostěnných buněk a schizogenních slizových dutin; stěny jsou neschopné vytvářet pigmenty anthokyanového typu; buňky neobsahují siličná tělíska, často pouze jeden obří chloroplast v buňce, ten často obsahuje pyrenoid antheridia tvořena endogenně v antheridiálních dutinách; spermatozoidy jsou bilaterálně symetrické první dělení zygoty je podélné bilaterální symetrie embrya a sporofytu sporofyt = válcovité (rohovité) zašpičatělé sporangium bez štětu, noha zanořena do gametoforu uvnitř sporofytu střední sloupek (kolumela); mezi výtrusy diferencovány sterilní buňky – pseudoelatery stěna sporangia mnohovrstevná, často s průduchy růst sporofytu obvykle neukončený, pomocí interkalárního meristému dozrávání postupné, od vrcholu; otvírání sporangia 1-2 podélnými liniemi, odshora chybí flavonoidy
Hlevíky (Anthocerotophyta) Zástupci, rozšíření Systém: bazální linie = Leiosporocerotopsida: jediný druh – Střední Amerika, Karibik, Ekvádor. Ostatní hlevíky – Anthocerotopsida: kosmopolitní, široce rozšířené rody se zástupci i v mírných oblastech: Anthoceros , Phaeoceros – růžicovitá stélka, 1 obří chloroplast s pyrenoidem, pseudoelatery dlouhé bez ztluštěnin. U nás na polích na kyselých podkladech. tropické rody – zejména Asie, Stř. a Již. Amerika: Dendroceros, Megaceros – pentlicovitá stélka se „středním žebrem“, jednotlivá antheridia, pseudoelatery se spir. ztluštěninami, absence průduchů. Často epifyti.
Mechorosty: hospodaření s vodou, fyziologické limitace mechorosty: alternativní způsob vyrovnání se s nepravidelnou dostupností vody – poikilohydrie (× kořenový systém + vodivá pletiva cévnatých) – za cenu nižší účinnosti metabolismu tolerance k vyschnutí u jednotlivých druhů velmi rozdílná příjem vody: ± celým povrchem stélky, pokud vyvinuta vosk. kutikula, na některých místech slabší usměrnění příjmu do určitých oblastí vedení vody: endohydrie – podstatná část vody vedena vnitřkem stélky (vod. pletiva – hydroidy; apoplast, vnitř. kapilární prostory), vnější povrch vodoodpudivý ektohydrie – voda rozváděna převážně po snadno smáčivém povrchu; využití externích kapilárních prostor – přiléhavé báze listů, papily mixohydrie – přechodný způsob mezi výše uvedenými Nedostatečná funkce vodivých pletiv neumožňuje velký vzrůst, ale naopak účinnější využití i malých a horizontálních srážek Fotosyntetické schopnosti – větší LAI (5-20), schopnost fotosyntézy při nižším světelném požitku, kladný výtěžek při nízkých teplotách Využití živin – ze substrátu velmi omezené, výhoda kořenů cévnatých; × lepší využití živin z atmosférické depozice
mechorosty: reakce na toxické látky, bioindikace vzhledem k relativně velkému povrchu stélky a malým bariérám snadná a poměrně rozsáhlá komunikace s okolím velká citlivost mechorostů na substrát i kvalitu okolního ovzduší nebo složení atmosf. srážek Vyhraněné substrátové nároky: bazifilnost, acidofilnost, ale i kuprofilní mechy, „spáleništní“ Funaria hygrometrica Citlivost ke znečištění ovzduší: závislost i na dalších faktorech (vzdušná vlhkost, charakter substrátu, životní forma…). obecně nejcitlivější epifytické, ektohydrické druhy (Orthotrichaceae, Jubulaceae, Lejeuneaceae…) relativně nejméně citlivé terestrické druhy, obzvlášť na bazickém substrátu (pufrační kapacita…) Biomonitoring těžkých kovů a radioaktivních izotopů využití velké absorpční schopnosti (viz výše), pomalého růstu a dlouhého živ. rozpětí některých druhů, až 100násobné koncentrace oproti cévnatým. Klasické používané druhy – velké lesní bokoplodé mechy Hylocomium splendens, Pleurozium schreberi
Bryologie: zdroje informací Učebnice, kompendia Goffinet B., Shaw A.J., ed. (2008): Bryophyte Biology, 2nd ed. Cambridge Univ. Press. Schofield W. B. (1985): Introduction to bryology. Macmillan, New York. Frahm J.-P. (2001): Biologie der Moose. Spektrum Akad. Verl., Heidelberg & Berlin. Kalina T. & Váňa J. (2005): Sinice, řasy, houby, mechorosty … – Karolinum, Praha. Plášek V. (2005): Základy bryologie (Systém, fylogeneze a ekologie mechorostů). – Skriptum, Ostravská univerzita v Ostravě. Váňa J. (2006-7): Obecná bryologie; Speciální bryologie I., II/1,2. Učební texty UK v Praze, Karolinum, Praha. Bryologie na internetu Bryoweb na katedře http://bryoweb.prf.jcu.cz/, dosažitelné i z http://botanika.prf.jcu.cz – prezentace k přednáškám a další Missouri Botanical Garden, Bryology: (Index of Mosses, Bibliography, Glossary, checklisty, flóry…) http://www.mobot.org/MOBOT/tropicos/most/welcome.shtml British Bryological Society, Portal: http://www.britishbryologicalsociety.org.uk/ Bryophyte Ecology – online publikace, J.M. Glime: http://www.bryoecol.mtu.edu/
Obrázky
Životní cyklus mechorostů Gametofyt lístek lodyžka noha Gametofor
Hlevíky: stavba stélky Anthoceros punctatus. Light micrograph showing dorsal schizogenous mucilage cavities. Photo by K. S. Renzaglia. Phaeoceros carolinianus. Light micrograph showing unicellular ventral rhizoids. Note lack of internal differentiation of the thallus. Photo by K. S. Renzaglia.. Duff, Nickrent & Renzaglia, hornwort pages (portal)
Hlevíky: chloroplasty Fig. 3. Transmission electron micrographs of chloroplasts in hornworts. A. Leiosporoceros dussii (Steph.) Hässel. Chloroplast in the assimilative layer of the sporophytes showing peripheral starch and centralized grana. B. Folioceros fuciformis Baradw. Central pyrenoid with lens-shaped subunits separated by narrow grana and surrounded by starch grains. Bar = 0.5 µm. Shaw & Renzaglia 2004 Phaeoceros laevis; A.M.F. Tomascu, Ohio Univ.
Hlevíky: průduchy Fig. 6. Scanning electron micrographs of pores in hornworts. A. Stoma in the sporophyte epidermis of Phaeoceros carolinianus (Michx.) Prosk. B. Ventral mucilage cleft in the gametophyte epidermis of Dendroceros crispatus (Hook.) Nees. Bars = 20 µm. Shaw & Renzaglia 2004 Dendroceros tubercularis. SEM of ventral side of thallus showing mucilage clefts (pores) along midrib. These are the sites of entry of the endosymbiont Nostoc bacteria. Bar = x µm. Specimen from Japan provided by Dr. J. Hasagawa. Photo by K. S. Renzaglia. Shaw & Renzaglia 2004
Hlevíky: spermatozoid, báze sporofytu Duff, Nickrent & Renzaglia, hornwort pages
Hlevíky: průřez sporangiem Fig. 4. Cross sections of hornwort sporophytes. A. Light micrograph of Leiosporoceros dussii (Steph.) Hässel. Tissue is differentiated from outside to inside as follows: single-layered epidermis, 9–10 layers of assimilative cells, abundant sporogenous tissue with several layers of tetrads intermixed with elaters, and an indistinct columella. The suture is clearly defined as a longitudinal groove that extends nearly to the sporogenous tissue. Bar = 100 µm. B. Scanning electron micrograph of Phaeoceros carolinianus (Michx.) Prosk. In contrast to Fig. 4A, this sporophyte contains an assimilative zone of four cell layers, sporogenous tissue with one layer of large tetrads intermixed with small elaters, and a columella of 16 cells. Bar = 1 mm. Shaw & Renzaglia 2004
Hlevíky: spory tetráda spor pseudoelatery Phaeoceros carolinianus, proximální a distální povrch spor mírka = 10 µm.; tetrády s pseudoelatery; http://www.science.siu.edu/landplants/anthocerophyta.html
Anthoceros Anthoceros agrestis, © M. Lüth
Dendroceros Dendroceros tubercularis. Color photo of an epiphytic hornwort with a swollen midrib and single-celled thallus wings. Note dark, spherical structures at the base of the thallus that house the endosymbiont Nostoc, a nitrogen-fixing cyanobacterium. Specimen from Japan provided by Dr. J. Hasagawa. Photo by Dan Nickrent. Dendroceros tubercularis, SEM, bar = 0,25 mm. Photo by K. S. Renzaglia.
Leiosporoceros American Journal of Botany 93(5): 693–705. 2006.
Játrovky: siličná tělíska Calypogeia mulleriana. Surface view of leaf cells showing oil bodies. Photo by Karen Renzaglia. http://www.science.siu.edu/lpo/hepatophyta.html Photograph of oil bodies in cells of leaves of the Calypogeia fissa complex: Bar = 50 µm. K. Buczkowska in Pl. Syst. Evol. 247: 187-201 (2004)
Játrovky: lupenitá stélka, sporofyt Pellia epiphylla, © M. Lüth
Játrovky: listnatá stélka Lophocolea bidentata, © M. Lüth
Játrovky: sporofyt, spory, elatery
Marchantiopsida: sporofyt Marchantia polymorpha, © Gilbert J. Muth, Pacific Union College
Marchantiopsida: stavba stélky zdroj:
Marchantiopsida: nosiče gametangií ♂ ♂ http://www.biology.lsa.umich.edu/courses/bio255/march3.jpg ♀ ♀ http://www.biology.iastate.edu/Courses/201L/Bryoph/marchantia.jpg
Marchantiales: Marchantia polymorpha © M. Lüth © M. Lüth
Marchantiales: Riccia Riccia cf. glauca, © P. Degroote Riccia ciliata. © M. Lüth
Jungermanniopsida: lupenité typy Metzgeria hamata Fossombronia alaskana
Jungermanniopsida: stavba stélky Aneura sharpii Pallavicinia sp., svazek hydroid
Jungermanniopsida: listnaté typy Lophocolea itoana
Jungermanniidae: zástupci – Evropa Diplophyllum albicans
Jungermanniidae: zástupci trop. skupin Frullania pycnantha, © Matt von Konrat, 2004
Mechy: stavba stélky, protonema
Mechy: tvar buněk listů
Mechy: gametofor, tvar a stavba listů
Mechy: stavba sporofytu čepička víčko peristom kolumela sporogenní tkáň apofýza štět průřez sporangiem
Mechy: obústí (peristom)
mechy: třída Sphagnopsida; gametofyt chlorocyty hyalocyty pór septum
mechy: třída Sphagnopsida; sporofyt pseudopodium víčko kolumela sporogenní tkáň noha
mechy: Takakia
mechy: třída Andreaeopsida – Andreaea
Polytrichopsida: stavba gametofytu
Polytrichopsida: vodivá pletiva
Polytrichopsida: nematodontní typ obústí
Polytrichopsida: zástupci Polytrichum commune Dawsonia longifolia Atrichum undulatum walter.obermayer@kfunigraz.ac.at
Bryopsida: gametofor
Bryopsida: Funariales
Bryopsida: Dicranidae Leucobryum glaucum
Bryopsida: Dicranidae Grimmia pulvinata Dicranum polysetum
Bryopsida: Dicranidae, sporofyty Grimmiaceae: Schistidium sp. Tortula muralis,© Bill Malcolm Grimmiaceae: Coscinodon australis
Bryopsida: Bryidae Rhodobryum ontariense Pohlia filum © M. Lüth
Bryidae: sporofyt Bryum sp., sporophyte © Z. Iwatsuki
Bryidae: Splachnaceae Splachnum ampullacum, Foto: Michael Lüth Splachnum luteum, Foto: Lena Eriksson
Hypnidae, znaky gametoforu Hylocomium splendens Hypnum plumaeforme, zpeřeně větvené lodyžky
Hypnidae: znaky gametoforu Thuidium sp., parafylie Brachythecium rutabulum, prosenchymatické buňky listu
Hypnidae: znaky sporofytu Hypnum tristoviride, dvojitý peristom, alternující zuby Daltonia cf. longifolia, bočně vyrůstající sporofyty © M. Lüth Brachythecium rutabulum, dvojitý peristom
papily
bioindikace - monitoring depozice těžkých kovů