Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Genetika mikroorganismů
Z latinského genaó = tvořím Přenos genetické informace je základní atribut živé hmoty Základní genetické pochody jsou konzervativní = podobné u všech organismů (společný původ?) Genetická informace je uložená v nukleových kyselinách
 Genetická informace je uložená v nukleových kyselinách.")
2
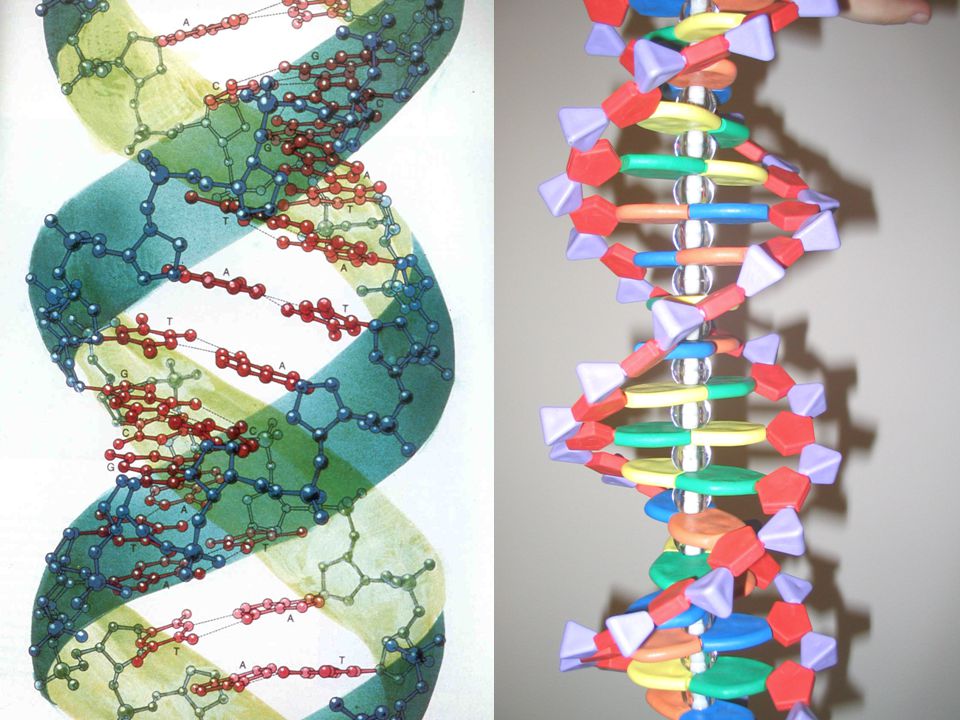
Nukleové kyseliny Základní nositelky dědičné informace
DNA = deoxirubonukleová kyselina (deoxyribonucleic acid) RNA = ribonukleová kyselina (ribonucleic acid) Fosfát – pentózová kostra s postranně navázanými bázemi Pořadí bází nese genetickou informaci
 RNA = ribonukleová kyselina (ribonucleic acid) Fosfát – pentózová kostra s postranně navázanými bázemi. Pořadí bází nese genetickou informaci.")
3
Nukleové kyseliny P P P P P P P P A T G C G A C C A G G G T C C T P P

4
Nukleové kyseliny DNA Deoxyribóza Báze Adenin, Guanin, Cytozin, Thymin
Stabilnější Obvykle dvouvláknová (double-stranded = ds) RNA Ribóza Báze Adenin, Guanin, Cytozin, Uracil Méně stabilní Obvykle jednovláknová (single-stranded ss)
 RNA. Ribóza. Báze Adenin, Guanin, Cytozin, Uracil. Méně stabilní. Obvykle jednovláknová (single-stranded ss)")
5
Báze nukleových kyselin
Purinové báze Adenin Guanin Pyrimidinové báze Cytosin Thymin Uracil

6
Dvouřetězcové NK NK bývají obvykle dvouvláknové = dva samostatné řetězce NK jsou spojené vodíkovými vazbami mezi bázemi Báze se tzv. párují Oba řetězce jsou tzv. komplementární = nesou opačnou genetickou informaci („negativ a pozitiv“)
")
7
Párování bazí Základ všech genetických pochodů
Párování umožňuje předávání a expresi genetické informace Purin (A, G) s pyridinem (T, U, C) 2 nebo 3 vodíkové můstky A = T A = U C ≡ G
 s pyridinem (T, U, C) 2 nebo 3 vodíkové můstky. A = T. A = U. C ≡ G.")
8
Šroubovicová struktura
Oba řetězce NK se vzájemně obtáčejí = dvojšroubovice Řetězce leží tzv. antiparalelně = konec jednoho leží u začátku druhého („hlava-pata“) podle popisu struktury pentózy jsou konce vlákna označovány jako 3’- a 5’-
 podle popisu struktury pentózy jsou konce vlákna označovány jako 3’- a 5’-")
10
Párování bazí DNA - DNA DNA – RNA RNA – RNA
5’- A T T C G G C T T A G G C G - 3’ 3’- T A A G C C G A A T C C G C - 5’ DNA – RNA 3’- U A A G C C G A A U C C G C - 5’ RNA – RNA 5’- A U U C G G C U U A G G C G - 3’

11
Genetická informace Pořadí bází určuje genetickou informaci
Gen = úsek NK kódující nějakou funkci Strukturní gen – kóduje strukturu bílkoviny Gen pro RNA – kóduje strukturu RNA Soubor všech genů = genom Kromě genů jsou v NK i další úseky Regulační (řídící) Nekódující – bez funkce nebo s neznámou funkcí – evolučně pokročilejší organismy mají více nekódujících sekvencí
 Nekódující – bez funkce nebo s neznámou funkcí – evolučně pokročilejší organismy mají více nekódujících sekvencí.")
12
Genetické pojmy Intron = sekvence DNA nekódující bílkovinu vmezeřená do strukturního genu Sestřih = proces odstraňování intronů ze strukturního genu Promotor = sekvence DNA uvozující gen nebo operon Operon = sekvence několika genů se společnou regulací Kodon = trojice bází kódující jednu aminokyselinu v peptidovém řetězci Antikodon = sekvence tří bazí komplementární ke kodonu

13
Typy RNA mRNA (messanger, mediátorová) = přenos exprimovaných genů z jádra na ribozómy rRNA (ribozomální) = stavební funkce v ribozómech, uplatňují se při translaci tRNA (transferová) = čtení genetického kódu, přenos aminokyselin při syntéze proteinů
 = stavební funkce v ribozómech, uplatňují se při translaci. tRNA (transferová) = čtení genetického kódu, přenos aminokyselin při syntéze proteinů.")
14
Základní genetické pochody
Replikace (zdvojení) = kopírování genetické informace do nové molekuly NK Transkripce (přepis) = kopírování malé části genetické informace z DNA do RNA Translace (překlad) = syntéza primární struktury bílkoviny podle informace v RNA
 = kopírování genetické informace do nové molekuly NK. Transkripce (přepis) = kopírování malé části genetické informace z DNA do RNA. Translace (překlad) = syntéza primární struktury bílkoviny podle informace v RNA.")
15
Základní genetické pochody
DNA replikace transkripce reverzní transkripce replikace RNA translace „Základní dogma molekulární biologie“ Bílkovina

16
Replikace Nutná pro předání genetické informace další generaci
Enzym DNA polymeráza Rozpletení dsDNA Ke každému vláknu je dosyntetizováno druhé komplementární Semikonzervativní (polovina nové molekuly DNA pochází od rodiče, polovina je nová)
")
17
Semikonzervativní replikace

18
P G C T A P P P A A T G A T T C P P P P P G C T A

19
Exprese genů Funkční protein
Soubor pochodů vedoucích od genu po funkční bílkovinu Možná regulace na všech úrovních transkripce translace gen mRNA polypeptid posttranslační modifikace transport Funkční protein

20
Transkripce Přepis jednoho nebo několika genů z DNA do mRNA
Enzym RNA polymeráza

21
Reverzní transkripce Pouze u tzv. retrovirů (např. HIV)
Přepis z RNA do DNA Enzym reverzní transkriptáza – nekontroluje chyby mutace Vzniklá DNA je integrována (začleněna) do genomu může vzniknout rakovina
 do genomu může vzniknout rakovina.")
22
Translace Syntéza bílkovin podle genetické informace
Probíhá na ribozómech Ribozóm se posouvá po mRNA a syntetizuje peptid Čtení genetické informace podle genetického kódu mRNA

23
Genetický kód Soubor kódů pro všechny aminokyseliny
20 kódovaných aminokyselin Jedna aminokyselina je kódována třemi bázemi (tzv. triplet nebo kodon) Triplet = 64 kombinací genetický kód je degenerovaný (více kódů pro jednu aminokyselinu) Genetický kód je univerzální (= až na výjimky stejný ve všech organismech)
 Triplet = 64 kombinací genetický kód je degenerovaný (více kódů pro jednu aminokyselinu) Genetický kód je univerzální (= až na výjimky stejný ve všech organismech)")
24
Genetický kód

25
Čtení genetického kódu
5’- A U U C G G C U U A G G - 3’ N Ile Arg Leu Arg C Nukleové kyseliny jsou čteny i syntetizovány od 5’ konce Bílkoviny jsou čteny i syntetizovány od N konce

26
Čtecí rámec 1 aminokyselina je kódována 3 bázemi = záleží na tom, kde se začne číst Čtecí rámec, jsou 3 možné začátky čtení, ale jen jeden je správný 5’- A U U C G G C U U A G G - 3’ N Ile Arg Leu Arg C

27
Čtecí rámec 5’- A U U C G G C U U A G G - 3’ N Ile Arg Leu Arg C

28
Čtecí rámec 5’- A U U C G G C U U A G G - 3’ N- Ile Arg Leu Arg -C
N Phe Gly Leu C

29
Čtecí rámec 5’- A U U C G G C U U A G G - 3’ N- Ile Arg Leu Arg -C
N Phe Gly Leu C N Ser Ala STOP C

30
Genomika Genetická informace uložena v DNA U některých virů i v RNA
Genomika = věda zabývající se genomy a genetickou informací Velikost genetické informace se udává v párech bazí (base pairs = bp)
")
31
Velikost genomů Organismus Bp Bakterioág MS2 3569 Escherichia coli
4.106 Mycoplasma genitalium Nanoarchaeum equitans Saccharomyces cerevisiae 2.107 Populus trichocarpa (topol) 4,8.108 Homo sapiens sapiens (člověk) 3,2.109 Fritillaria assyriaca 1,3.1011 Amoeba dubia (měňavka) 6,7.1011
 4, Homo sapiens sapiens (člověk) 3, Fritillaria assyriaca. 1, Amoeba dubia (měňavka) 6,")
32
Genetická informace Jaderná Mimojaderná
Chromózómy = hlavní „velké“ DNA Mimojaderná Plasmidy = malé kruhové DNA u bakterií DNA v organelách (mitochondrie, plastidy)
")
33
Bakteriální genóm Obvykle kruhová DNA Obvykle jeden chromozóm
Volně v cytoplazmě DNA obalena alkalickými aminy (spermin a spermidin) Časté plasmidy, i několik desítek Bez intronů Geny uspořádány do operonů
 Časté plasmidy, i několik desítek. Bez intronů. Geny uspořádány do operonů.")
34
Eukaryotický genom Lineární DNA Obvykle více chromozómů (člověk 46)
DNA je uložena v jádře obaleném dvojitou membránou Zahuštěna do kompaktní struktury pomocí alkalických bílkovin (histonů) Plasmidy výjimečné Geny mají introny Operony nenacházíme
 Plasmidy výjimečné. Geny mají introny. Operony nenacházíme.")
35
Archeální genóm Mezistupeň mezi bakteriemi a eukaryoty
Obvykle lineární Volně v cytoplasmě DNA obalena bílkovinami podobnými eukaryotickým histonům Primitivní introny Plasmidy (méně než bakterie) Geny organizované do operonů
 Geny organizované do operonů.")
36
Plasmidy Krátké cyklické úseky DNA Výskyt zejména u bakterií
V buňce může být i několik stovek plasmidů, stejných i různých Některé plasmidy vzájemně nekompatibilní (nemohou být v jedné buňce) Nezávisle se replikují Občas vymizení
 Nezávisle se replikují. Občas vymizení.")
37
Plasmidy Postaru nazývané faktory
Kódují vlastnosti, které bakterie k životu nutně nepotřebuje rezistenci vůči antibiotikům (tzv. RTF – Resistence Transfer Factor) nové metabolické dráhy (např. odbourávání uhlovodíků apod.) produkce toxinů
 nové metabolické dráhy (např. odbourávání uhlovodíků apod.) produkce toxinů.")
38
Plasmidy Význam v genetickém inženýrství
umělé plasmidy s požadovanými vlastnostmi

39
Replikace u bakterií Dvojsměrná 3 fáze Iniciace (zahájení)
v tzv. místě ori (origin = počátek) Elongace (prodlužování) Terminace (zakončení)
 Elongace (prodlužování) Terminace (zakončení)")
40
Replikační enzymy u bakterií
DNA polymerázy 3 druhy (DNA-pol I. II. a III.) dNTP + DNAn PP + DNAn+1 dNTP = dATP, dGTP, dCTP, dTTP napojování nukleotidů na 3’-konec neumí začátek řetězce, jen napojovat, potřebuje tzv. primer (očko) = krátká sekvence DNA nebo RNA DNA primáza syntetizuje krátký fragment RNA - primer
 dNTP + DNAn PP + DNAn+1. dNTP = dATP, dGTP, dCTP, dTTP. napojování nukleotidů na 3’-konec. neumí začátek řetězce, jen napojovat, potřebuje tzv. primer (očko) = krátká sekvence DNA nebo RNA. DNA primáza. syntetizuje krátký fragment RNA - primer.")
41
N H 2 O P -

42
Replikační enzymy u bakterií
DNA ligáza spojuje delší řetězce DNA DNA helikázy rozplétají dvojšroubovici na jednotlivá vlákna

43
Iniciace replikace 1. Rozpoznání ori místa 2. Navázání helikáz
DnaA proteiny – najdou ori místo a oddělí v něm oba řetězce DNA ori místo je bohaté na AT páry = snadno se oddělí vznik tzv. replikační vidlice 2. Navázání helikáz na oba konce vidlice se naváže helikáza a začne rozplétat DNA 3. Navázání DNA polymeráz a dalších replikačních enzymů

44
Iniciace replikace A A A A

45
Iniciace replikace A A A A

46
Iniciace replikace A A A A H H

47
Iniciace replikace A H H A A A

48
Iniciace replikace A H H A A A

49
Iniciace replikace Pol A H Pol H A A A

50
Iniciace replikace Pol A H Pol H A A A

51
Elongace DNA Syntézu obou řetězců katalyzuje enzymový komplex DNA-polymerázy III. Semidiskontinuální = jeden řetězec se syntetizuje nepřetržitě (vedoucí řetězec) a jeden po kratších úsecích (opožďující se řetězec) Okazakiho fragmenty = krátké úseky DNA ( nukleotidů) DNA polymeráza syntetizuje řetězec ve směru 5’-3’, ale řetězce mají opačnou orientaci cca 500 nukletidů / s
 a jeden po kratších úsecích (opožďující se řetězec) Okazakiho fragmenty = krátké úseky DNA ( nukleotidů) DNA polymeráza syntetizuje řetězec ve směru 5’-3’, ale řetězce mají opačnou orientaci. cca 500 nukletidů / s.")
52
Semidiskontinuální elongace
3’ 5’ vedoucí řetězec Pol 5’ 3’ Okazakiho fragmenty opožďující se řetězec 3’ 5’

53
Elongace DNA Syntéza opožďujícího se řetězce:
Syntéza RNA primeru (primáza) Elongace fragmentu (DNA polymeráza I.) Odstranění primeru (DNA polymeráza I.) Zaplnění mezery (DNA polymeráza I.) Spojení řetězců (DNA ligáza)
 Elongace fragmentu (DNA polymeráza I.) Odstranění primeru (DNA polymeráza I.) Zaplnění mezery (DNA polymeráza I.) Spojení řetězců (DNA ligáza)")
54
Semidiskontinuální elongace
3’ 5’ vedoucí řetězec Pol 5’ 3’ primery opožďující se řetězec 3’ 5’

55
Terminace replikace Na tzv. ter místě Tus protein – inhibice helikázy

56
Replikace plasmidů Analogická repliklaci chromozómů
Časově oddělená replikace vedoucího a opožďující se řetězce – tzv. replikace valivou kružnicí nejprve je syntetizován vedoucí řetězec, který vytlačí původní – 1. kopie plasmidu k vytlačenému původnímu řetězci se pomocí Okazakiho fragmentů dosyntetizuje nový řetězec – 2. kopie plasmidu

57
Replikace plasmidů

58
Replikace u eukaryot V principu stejná jako u bakterií Odlišnosti
jiné DNA polymerázy (ale s podobnou funkcí) replikace probíhá na mnoha místech naráz eukaryotické chromozómy jsou lineární na 5’ konci nových řetězců chybí úsek, který není na co navázat – speciální enzym telomeráza 3’ 5’ 3’ 5’ 5’ 3’ 5’ 3’
 replikace probíhá na mnoha místech naráz. eukaryotické chromozómy jsou lineární na 5’ konci nových řetězců chybí úsek, který není na co navázat – speciální enzym telomeráza. 3’ 5’ 3’ 5’ 5’ 3’ 5’ 3’")
59
Replikace u archeí Velice podobná bakteriální replikaci
DNA polymerázy strukturně podobné eukaryotickým, ale s podobnou funkcí jako u bakterií

60
Transkripce u bakterií
3 fáze Iniciace (zahájení) Elongace (prodlužování) Terminace (zakončení) Enzym RNA-polymeráza 5 podjednotek – a2bb’s Transkripce je zahajována na tzv. promotoru – uvozující úsek DNA různé promotory u různých genů Rychlost cca 40 nukleotidů / s
 Elongace (prodlužování) Terminace (zakončení) Enzym RNA-polymeráza. 5 podjednotek – a2bb’s. Transkripce je zahajována na tzv. promotoru – uvozující úsek DNA. různé promotory u různých genů. Rychlost cca 40 nukleotidů / s.")
61
Sigma faktor Podjednotka s (sigma faktor) má za úkol rozpoznat promotor bakterie mají více sigma faktorů pro geny různého typu Síla promotoru = pravděpodobnost zahájení transkripce geny se silnějším promotorem jsou více exprimovány závisí na sekvenci promotoru i sigma faktoru

62
Pozitivní a negativní řetězec
Transkripcí dsDNA vzniká ssRNA Přepisuje se jen jedno vlákno ze dvou = negativní vlákno Nepřepisované vlákno = pozitivní vlákno Analogie s fotografií – negativ a pozitiv

63
Iniciace transkripce 1. Spojení sigma-faktoru s RNA polymerázou s
a2bb’ promotor přepisované geny

64
Iniciace transkripce 2. Sigma podjednotka rozpozná promotor a naváže RNA polymerázu na DNA s a2bb’ promotor přepisované geny

65
Iniciace transkripce 3. Rozpletení DNA

66
Iniciace transkripce 4. Zahájení syntézy RNA

67
Elongace RNA 5. Syntéza RNA – sigma-faktor se oddělí

68
Terminace transkripce
Pomocí vlásenky terminátor = koncový úsek přepisované RNA palindromatická sekvence (symetrická) = páruje se sama se sebou a vzniká vlásenka vlásenka „vykolejí“ RNA polymerázu Pomocí r-faktoru bílkovina, rozpoznávající terminátor interakce s RNA polymerázou a ukončení transkripce
 = páruje se sama se sebou a vzniká vlásenka. vlásenka „vykolejí RNA polymerázu. Pomocí r-faktoru. bílkovina, rozpoznávající terminátor. interakce s RNA polymerázou a ukončení transkripce.")
69
Transkripce u eukaryot
V principu stejná jako u bakterií Odlišnosti více RNA polymeráz odlišných od bakteriální odlišné promotory k zahájení transkripce jsou potřeba iniciační faktory (bílkoviny); obvykle několik složitější iniciace složitější terminace, nejčastěji pomocí tzv. polyadenylačního signálu = sekvence AATAAA Posttranskripční úpravy mRNA
; obvykle několik. složitější iniciace. složitější terminace, nejčastěji pomocí tzv. polyadenylačního signálu = sekvence AATAAA. Posttranskripční úpravy mRNA.")
70
Posttranskripční úpravy mRNA
U eukaryot je přepsaná mRNA ještě podrobena tzv. posttranskripčním úpravám vytvoření tzv. čepičky na 5’-konci komplex se specifickými proteiny sestřih = odstranění intronů polyadenylace 3’-konce

71
Transkripce u archeí Podobnosti i odlišnosti proti bakteriím a eukaryím Podobnosti s eukaryotickou transkripcí podobné RNA-polymerázy podobné iniciační faktory Podobnosti s bakteriální transkripcí žádné posttranskripční úpravy mRNA podobné promotory přepis operonů do jedné mRNA

72
Bakteriální translace

73
tRNA Prostředník při překladu z genetického kódu do „řeči“ aminokyselin Jednu aminokyselinu může přenášet více tRNA = izoakceptorové Ve struktuře tRNA jsou zařazeny i nestandardní nukleotidy (pseudouridin, 1-methylguanozin…) – vznik modifikací standardních nukleotidů 74-95 nukleotidů sekundární struktura připomíná jetelový lístek
 – vznik modifikací standardních nukleotidů nukleotidů. sekundární struktura připomíná jetelový lístek.")
74
tRNA Vazebné místo pro aminokyselinu Smyčky Antikodon

75
Antikodon Sekvence 3 nukleotidů komplementární s kodonem pro danou aminokyselinu Nestandardní nukleotidy umožňují rozšířené párování

76
Aktivace aminokyselin
Proces napojování aminokyselin na tRNA Pro každý pár aminokyselina-tRNA existuje jeden enzym aminoacyl-tRNA-syntetáza Rozpoznává správné aminokyseliny i tRNA Syntetizuje jejich vazbu (energie ze štěpení ATP) aa + ATP aa-AMP + PP aa-AMP + tRNA aa-tRNA + AMP
 aa + ATP aa-AMP + PP. aa-AMP + tRNA aa-tRNA + AMP.")
77
Bakteriální ribozómy Kuličky složné z bílkovin a rRNA
Označení komponent podle sedimentačního koeficientu celý bakteriální ribozóm má sedimentační koeficient 70S 16S-rRNA, 23S-rRNA, 5S-rRNA Dvě podjednotky malá 30S velká 50S

78
Vazebná místa na ribozómu
Vazebné místo pro mRNA Aminoacylové místo (A-místo) – vazba nepřipojené aa-tRNA Peptidylové místo (P-místo) – vazba už hotového peptidového řetězce Výstupní místo pro tRNA (E-místo) – odchod deacylované tRNA
 – vazba nepřipojené aa-tRNA. Peptidylové místo (P-místo) – vazba už hotového peptidového řetězce. Výstupní místo pro tRNA (E-místo) – odchod deacylované tRNA.")
79
Průběh translace Iniciace Elongace – prodlužování řetězce Terminace
rozpoznání čtecího rámce zařazení první aminokyseliny (formylmethionin) iniciační faktory (IF) - bílkoviny Elongace – prodlužování řetězce elongační faktory (EF) Terminace terminační (nesmyslný) kodon = nekóduje žádnou aminokyselinu, ale konec řetězce účast terminačních faktorů (RF)
 iniciační faktory (IF) - bílkoviny. Elongace – prodlužování řetězce. elongační faktory (EF) Terminace. terminační (nesmyslný) kodon = nekóduje žádnou aminokyselinu, ale konec řetězce. účast terminačních faktorů (RF)")
80
Iniciace translace 1.Rozpad ribozómu na podjednotky E P A

81
Iniciace translace 2.Navázání fMet-tRNA fMet E P A

82
Iniciace translace 3.Navázání mRNA
správné umístění pomocí tzv. Shine-Dalgarnovy sekvence E P A fMet

83
Shine-Dalgarnova sekvence
Sekvence AGGA na mRNA, která se páruje s UCCU na 16S-rRNA Zajišťuje správné umístění mRNA na ribozóm ribozóm 3’ 5’ 16S-rRNA UCCU AGGA AUG iniciační kodon 5’ 3’ mRNA

84
Iniciace translace 4.Znovuspojení ribozomálních podjednotek E P A fMet

85
Elongace polypeptidového řetězce
6. Navázání další aminokyseliny do A místa Trp E P A fMet

86
Elongace polypeptidového řetězce
7. Vznik peptidové vazby fMet E P A Trp

87
Elongace polypeptidového řetězce
8. Posun ribozómu E A fMet P Trp

88
Elongace polypeptidového řetězce
9. Navázání další aminokyseliny E A fMet P Trp Lys

89
Elongace polypeptidového řetězce
10. Vznik peptidové vaby fMet Trp E A P Lys

90
Elongace polypeptidového řetězce
11. Posun ribozómu fMet Trp E A P Lys

91
Elongace peptidového řetězce
mRNA

92
Terminace translace Terminační faktory RF1 a RF2 rozpoznají terminační kodón a za spolupráce s RF3 způsobí uvolnění tRNA, peptidu a rozpad ribozómu na podjednotky

93
Rychlost proteosyntézy
10-20 aminokyselin / s chybovost cca 1 nesprávná aminokyselina na 2000 správných

94
Translace polygenních RNA
U bakteriích je mnoho mRNA polygenních = nesou více genů (např. operony) U každého genu nová iniciace translace
 U každého genu nová iniciace translace.")
95
Návaznost transkripce a translace
U bakterií dochází k rychlému navazování transkripce a translace na nehotovou mRNA už nasedají ribozómy a překládají polypeptid na jedné RNA může být současně ribozómů posunujících se „za sebou“

96
RNA pol DNA RNA

97
peptid ribozóm ribozóm ribozóm ribozóm ribozóm ribozóm RNA pol DNA RNA

98
Eukaryotická transalce
V principu podobná bakteriální ale s odlišnostmi ribozómy jsou větší a odlišné (80S) volné vázané na endoplasmatické retikulum – syntéza membránových bílkovin první aminokyselinou je methionin a ne formylmethionin eukarya nepoužívají Shine-Dalgarnovu sekvenci, správný začátek transalce je rozpoznáván pomocí čepičky více translačních faktorů
 volné. vázané na endoplasmatické retikulum – syntéza membránových bílkovin. první aminokyselinou je methionin a ne formylmethionin. eukarya nepoužívají Shine-Dalgarnovu sekvenci, správný začátek transalce je rozpoznáván pomocí čepičky. více translačních faktorů.")
99
Eukaryotická transalce
Prostorové oddělení eukaryální transkripce a translace transkripce probíhá v jádře translace probíhá mimo jádro – nutný transport Časové oddělení translace probíhá teprve po dokončení všech posttranskripčních úprav

100
Archeální transalce Podobnosti s bakteriální transalcí
podobné ribozómy používání Shine-Dalgarnovy sekvence Podobnosti s eukaryální translací podobné translační faktory

101
Mutace Změna dědičné (genetické) informace
Obvykle předávaná dalším generacím

102
Mutace Dělení podle příčiny
samovolné (spontánní) – chyby při replikaci DNA vyvolané (indukované) – způsobené faktory vnějšího prostředí (mutageny)
 – chyby při replikaci DNA. vyvolané (indukované) – způsobené faktory vnějšího prostředí (mutageny)")
103
Mutace Dělení podle rozsahu bodové – změna jednoho páru bazí
chromozómové – změna delšího úseku DNA genomové – změna počtu chromozómů (jen u eukaryot)
")
104
Mutace Dělení podle následků
neutrální – žádný vliv na vlastnosti a životaschopnost téměř neutrální – přibližně nulový vliv na vlastnosti a životaschopnost negativní – zhoršení vlastností či životaschopnosti pozitivní – zlepšení vlastností či životaschopnosti

105
Mutace Dělení podle následků pokles pravděpodobnosti
neutrální – žádný vliv na vlastnosti a životaschopnost téměř neutrální – přibližně nulový vliv na vlastnosti a životaschopnost negativní – zhoršení vlastností či životaschopnosti pozitivní – zlepšení vlastností či životaschopnosti pokles pravděpodobnosti

106
Bodové mutace Obvykle změna jedné báze, ke které se při další replikaci připáruje správná ATTCTTGCGGTCGAATGCCGCTCAATCGG TAAGAACGCCAGCCTACGGCGAGTTAGCC

107
Bodové mutace Obvykle změna jedné báze, ke které se při další replikaci připáruje správná ATTCTTGCGGTCGAATGCCGCTCAATCGG TAAGAACGCCAGCCTACGGCGAGTTAGCC

108
Bodové mutace Obvykle změna jedné báze, ke které se při další replikaci připáruje správná ATTCTTGCGGTCGAATGCCGCTCAATCGG TAAGAACGCCAGCTTACGGCGAGTTAGCC ATTCTTGCGGTCGGATGCCGCTCAATCGG TAAGAACGCCAGCCTACGGCGAGTTAGCC

109
Bodové mutace Typy bodových mutací Inzerce a delece mění čtecí rámec
tranzice – výměna purinu za purin resp. pyrimidinu za pyrimidin (A↔G C↔T) transverze – výmna purinu za pyrimidin a opačně (A,G↔C,T) inzerce – vložení nového páru bazí delece – odstranění jednoho páru bazí Inzerce a delece mění čtecí rámec
 transverze – výmna purinu za pyrimidin a opačně (A,G↔C,T) inzerce – vložení nového páru bazí. delece – odstranění jednoho páru bazí. Inzerce a delece mění čtecí rámec.")
110
Bodové mutace Mutace s chybným smyslem (missense) = mutací vznikne kodon pro jinou aminokyselinu kódovaná bílkovina si obvykle uchová aktivitu Nesmyslné mutace (nonsense) = mutací vznikne stop-kodon vzniká neúplná bílkovina obvykle bez funkce
 = mutací vznikne stop-kodon. vzniká neúplná bílkovina obvykle bez funkce.")
111
Chromozómové mutace Vznikají zlomem DNA a chybným znovuspojením
koncová deficience – chybí koncová část DNA interkalární delece – chybí vnitřní část DNA inverze – část DNA je vložena obráceně translokace – přeskupení DNA duplikace – zdvojení DNA

112
Chromozómové mutace Původní DNA A B C D E F G Koncová deficience
Interkalární delece A B C F G Inverze A D C B E F G Translokace A B D E F G C Duplikace A B C D C D C D E F G

113
Genomové mutace Změna počtu chromozómů Zvýšení a snížení
Polyploidie = znásobení genetické informace obvykle max. hexaploidie, víceplodiní jádra se při mitóze rozpadají na dvě Aneuploidie = neúplná genetická informace

114
Spontání mutace Způsobené bez viditelného vlivu mutagenu
Nesprávné párování bazí Deaminace bazí C U (páruje se s A) A hypoxantin (páruje se s C) G xantin (nepáruje se, zastavení translace) Oxidativní poškození kyslíkové radikály, hlavně OH·, vznik z H2O2 (vedlejší produkt dýchacího řetězce) různé produkty se změněným párováním nebo bez párování
 A hypoxantin (páruje se s C) G xantin (nepáruje se, zastavení translace) Oxidativní poškození. kyslíkové radikály, hlavně OH·, vznik z H2O2 (vedlejší produkt dýchacího řetězce) různé produkty se změněným párováním nebo bez párování.")
115
Mutageny mutagen = fyzikální faktor nebo chemická látka způsobující mutace Analoga bazí – strukturní podobnost, bazím, ale odlišné párování bodové mutace př. 5-bromuracil (AT↔GC) Kyselina dusitá – deaminace Alkylační látky – křížové vazby mezi řetězci Interkalární látky – planární molekuly, vmezeřují se mezi báze DNA a narušují operace s DNA polyaromatické uhlovodíky
 Kyselina dusitá – deaminace. Alkylační látky – křížové vazby mezi řetězci. Interkalární látky – planární molekuly, vmezeřují se mezi báze DNA a narušují operace s DNA. polyaromatické uhlovodíky.")
116
Mutageny Ionizující záření (rentgen, radioaktivní…)
excitace elektronů a vznik náhodných vazeb vliv přímo na DNA nebo nepřímo přes jiné molekuly, zejména vodu zlomy v DNA změny bazí UV záření – NK absorbují mezi nm vznik thyminových dimerů

117
Reparační procesy Buňky mají schopnost opravy (reparace) poškozené DNA
menší poškození je možno odstranit bez ztráty genetické informace Úplná oprava – chemická reakce odstraňující poškození odstranění thyminových dimerů Excizní oprava – vyštěpení jednoho řetězce v chybném místě a jeho správné dotvoření Tolerantní oprava – úprava poškozené DNA neodstraňující mutaci, ale umožňující funkci DNA (replikace, transkripce)
")
118
Konjugace u bakterií Proces výměny genetického materiálu mezi některými rody G- bakterií Je možný i mezi různými rody Zdroj variability a odolnosti bakterií Výměna probíhá pomocí kanálků tvořených fimbriemi Geny pro fimbrie jsou kódovány na F plazmidu (F+ / F-) F plazmid je epizóm = může se integrovat do genomu a vyštěpovat zpět jako plazmid
 F plazmid je epizóm = může se integrovat do genomu a vyštěpovat zpět jako plazmid.")
119
F+ F- konjugace Proces je jednosměrný z F+ buňky do F- buňky
F+ - mají F plazmid, jsou schopné tvorby fimbrií a přenosu genetické informace F- - nemají F plazmid, mohou být akceptory Konjugace neprobíhá ve směru F+ F+ F- buňka se po přijetí stává F+ buňkou F+ buňka zůstává F+ buňkou

120
F+ F- konjugace 1. Spojení buněk kanálkem F+ F-

121
F+ F- konjugace 1. Spojení buněk kanálkem F+ F-

122
F+ F- konjugace 1. Spojení buněk kanálkem F+ F-

123
F+ F- konjugace 2. replikace F-plazmidu valivou kružnicí F+ F-

124
F+ F- konjugace 3. Dokončení druhého vlákna plazmidu F+ F-

125
F+ F- konjugace 4. Oddělení buněk F+ F-

126
Hfr konjugace Hfr = High frequency of recombination (vysoká frekvence rekombinace) Nastává, pokud se F-plazmid integruje do chromozómu S F-plazmidem se přenáší i kus chromozómu přenos mnoha genů začátek v místě integrace F-plazmidu Velmi pomalý proces (u E.coli cca 100 minut) – často předčasné přerušení Obvykle se nepřenese celý F-plazmid – recipientní buňka zůstane F-
 – často předčasné přerušení. Obvykle se nepřenese celý F-plazmid – recipientní buňka zůstane F-")
127
Hfr konjugace 1. Spojení buněk kanálkem Hfr F-

128
Hfr konjugace 1. Spojení buněk kanálkem Hfr F-

129
Hfr konjugace 1. Spojení buněk kanálkem Hfr F-

130
Hfr konjugace 2. přenos části DNA do druhé buňky Hfr F-

131
Hfr konjugace 3. rozpad spojení Hfr F-

132
Hfr konjugace 4. Integrace části přenesené DNA do chromozómu příjemce

133
Další konjugace Existuje ještě řada dalších procesů výměny genetické informace mezi bakteriemi s různým mechanismem

Podobné prezentace


>")







>")
>")







