Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
ZKOUŠKOVÉ TERMÍNY 8.2. 1.3. 18.1. Vždy středa 9:00, posluchárna B2
Písemný test, kombinace výběrových a volných odpovědí

2
Mutace a opravy DNA (kapitola 23, 24)
Molekulární biologie 10. Mutace a opravy DNA (kapitola 23, 24)
")
3
MUTACE DNA změny genetického materiálu daného organizmu (dědičné u potomků dané buňky) pouze mutace v zárodečných buňkách povedou k dědičným změnám, ostatní mutace ovlivní pouze daného jedince většina mutací není letální, ani se na fenotypu neprojeví. Některé mutace dokonce přinášejí organizmu výhodu. vetšina mutací je recesivních, efekt vykompenzován druhou alelou Každý člověk nese v genomu škodlivých recesivních alel, z toho 8 letálních. Kdyby člověk existoval jako haploid, byl by dávno mrtvý! K tomu má každý člověk v genomu asi změn, které jsou ale neškodné

4
MUTACE DNA Osud mutací: mutace opravena
mutace spustí programovanou buněčnou smrt (apoptózu) mutace ignorována Dopad mutace na funkci genu: žádný mutace mírně pozmění aktivitu nebo expresi genu, ale zachová jeho funkci mutace úplně změní funkci genu nebo se gen přestane vyrábět Tichá mutace – neutrální, nemá vliv na funkci daného genu, bez fenotypu, např. ve třetí pozici kodonu, v intronech, v intergenové DNA Nulová mutace – úplně chybí produkt daného genu, loss of function Substituce báze Inserce Delece Inverse Duplikace Translokace uvnitř genu nebo v regulačních sekvencích
 mutace ignorována. Dopad mutace na funkci genu: žádný. mutace mírně pozmění aktivitu nebo expresi genu, ale zachová jeho funkci. mutace úplně změní funkci genu nebo se gen přestane vyrábět. Tichá mutace – neutrální, nemá vliv na funkci daného genu, bez fenotypu, např. ve třetí pozici kodonu, v intronech, v intergenové DNA. Nulová mutace – úplně chybí produkt daného genu, loss of function. Substituce báze. Inserce. Delece. Inverse. Duplikace. Translokace. uvnitř genu nebo v regulačních sekvencích.")
5
TYPY MUTACÍ Substituce báze (bodová mutace)
")
6
Srpková anemie (thalasemie) – bodová mutace v genu pro hemoglobin
kyselina glutamová valin V populaci se rozsirila, protoze dava vetsi rezistenci na malarii

7
Missense mutace – změní kódovanou aminokyselinu v proteinu
neutrální mutace (tichá) (nahrazení aminokyselinou s podobnými vlastnostmi) radikální mutace (nahrazení aminokyselinou s velmi rozdílnými vlastnostmi, vliv na strukturu a tím funkci proteinu) Aminokyseliny konzervované u rozdíných organizmů budou dost možná v aktivním místě proteinu a jejich mutace bude mít závažnější důsledky než mutace ostatních aminokyselin.
 (nahrazení aminokyselinou. s podobnými vlastnostmi) radikální mutace. (nahrazení aminokyselinou. s velmi rozdílnými vlastnostmi, vliv na strukturu a tím funkci proteinu) Aminokyseliny konzervované u rozdíných organizmů budou dost možná v aktivním místě proteinu a jejich mutace bude mít závažnější důsledky než mutace ostatních aminokyselin.")
8
Kondicionální mutace – efekt záleží na podmínkách, ve kterých organizmus žije
např. teplotně sensitivní mutace – projeví se jen při určitých teplotách Mutace v melaninu, která ho dělá aktivnějšímv chladnějších částech těla.

9
Nonsense mutace = většinou efekt jako nulová mutace
Nonsense mutace – zavedení předčasného stop kodonu v mRNA např. UCG UAG ser stop Prokaryota – protein je syntetizován až do předčasného stop kodonu, pak uvolněn, ale protože většinou není správně složený, je degradován Eukaryota – nonsense mediated mRNA decay, zničí se rovnou mRNA Nonsense mutace = většinou efekt jako nulová mutace

10
TYPY MUTACÍ Delece Delece může buď naktivovat geny nebo i jejich expresi zvýšit (delece vazebného místa pro represor v regulační oblasti) Delece jedné či několika bazí – frameshift ve čtecím rámci nebo vliv na splicing, vliv na vazbu transkripčních faktorů atd.

11
Frameshift -1 Frameshift -2 Frameshift -3

12
TYPY MUTACÍ Inzerce Inzerce mobilního genetického elementu (transposony, retrotransposony), virů, nebo chybou DNA polymerázy zastavení exprese zvýšení exprese

13
Translokace Duplikace
TYPY MUTACÍ Inverze Translokace Duplikace

14
Chemické – chemické látky, těžké kovy, kouř, volné radikály…
VZNIK MUTACÍ Spontánní mutace chyby v DNA replikaci, spontánní chemické změny na DNA (s nízkou frekvencí, se zvyšující se teplotou frekvence roste) Indukované mutace Mutageny: vlivy, který svým působením zvyšují pravděpodobnost mutace v DNA Chemické – chemické látky, těžké kovy, kouř, volné radikály… Fyzikální – radiace, vysoké teploty Biologické – některé viry, transposony (neintegruji se přesně, excise, nepřesná oprava) Vysoké teploty zvysuji rychlost spontannich deaminaci, depuinaci atd.
 Indukované mutace. Mutageny: vlivy, který svým působením zvyšují pravděpodobnost mutace v DNA. Chemické – chemické látky, těžké kovy, kouř, volné radikály… Fyzikální – radiace, vysoké teploty. Biologické – některé viry, transposony (neintegruji se přesně, excise, nepřesná oprava) Vysoké teploty zvysuji rychlost spontannich deaminaci, depuinaci atd.")
15
EMS (ethyl methane sulphonate)
INDUKOVANÉ MUTACE Chemické mutageny: EMS (ethyl methane sulphonate) alkylační činidlo, přidává ethylové skupiny bazím pro in vivo mutagenezi živých buněk Dusitany (NO2-) přeměna aminoskupin na hydroxyskupiny mutace 5-methyl cytosinu na thymin pro in vitro mutagenezi plazmidů Pri zahrivani dusitanu vznikaji karcinogenni nitrosaminy…. Opekani spekacku… E250 – dusitan sodný přeměna na rakovinotvorné nitrosaminy
 alkylační činidlo, přidává ethylové skupiny bazím. pro in vivo mutagenezi živých buněk. Dusitany (NO2-) přeměna aminoskupin na hydroxyskupiny. mutace 5-methyl cytosinu na thymin. pro in vitro mutagenezi plazmidů. Pri zahrivani dusitanu vznikaji karcinogenni nitrosaminy…. Opekani spekacku… E250 – dusitan sodný. přeměna na rakovinotvorné. nitrosaminy.")
16
Analogy bází 5-bromouracyl inkorporuje se do DNA během replikace jako thymin existuje ve dvou flip-flop stavech, z nichž jeden se páruje s A a jeden s G problém při následné DNA replikaci, kdy slouží jako templát

17
Interkalační činidla acridine orange, acriflavin, ethidium bromid vmezeří se mezi dvě báze DNA DNA polymeráza během replikace pak rozezná tuto látku jako skutečnou bázi a vloží extra nukleotid do DNA, frameshift… Teratogen – látka způsobující vývojové vady embryí (buď jako mutagen nebo jinými mechanizmy)
")
18
Mutace způsobené radiací:
INDUKOVANÉ MUTACE Mutace způsobené radiací: 1) elektromagnetické vlnění s velmi vysokou frekvencí (UV) UV světlo má vlnovou délku nm, báze absorbují nejvíce při vlnové délce 254nm, tvorba thyminových dimérů, problém při replikaci většina UV ze slunce zachycena ozonovou vrstvou, pokud není poškozená… Thyminové diméry thymin thyminový dimer 2) ionizační záření (X-ray, g-ray) přímé poškození DNA = reakcí záření s DNA (hlavně ds zlomy) při styku s vodou či jinými látkami indukují tvorbu iontů a volných radikálů (hlavně OH.) = nepřímé poškození DNA
 elektromagnetické vlnění s velmi vysokou frekvencí (UV) UV světlo má vlnovou délku nm, báze absorbují nejvíce při vlnové délce 254nm, tvorba thyminových dimérů, problém při replikaci. většina UV ze slunce zachycena ozonovou vrstvou, pokud není poškozená… Thyminové diméry. thymin. thyminový dimer. 2) ionizační záření (X-ray, g-ray) přímé poškození DNA = reakcí záření s DNA (hlavně ds zlomy) při styku s vodou či jinými látkami indukují tvorbu iontů a volných radikálů (hlavně OH.) = nepřímé poškození DNA.")
19
INDUKOVANÉ MUTACE .OH .O2- Poškození oxidativním stresem (= volnými radikály = reactive oxygen species = ROS) 8-hydroxyguanin = 8-oxoguanin Hydroxylovy radikal, superoxid, takz dusikate radikaly (NO…) a dalsi… Formace 8-oxoguaninu (= 8-hydroxyguanin) párování s A záměna G/C za A/T
 a dalsi… page_id=600. Formace 8-oxoguaninu. (= 8-hydroxyguanin) párování s A. záměna G/C za A/T.")
20
Mutace vzniklé chybami DNA polymerázy
SPONTÁNNÍ MUTACE Mutace vzniklé chybami DNA polymerázy proofreading aktivita DNA polymerázy není dokonalá, s malou frekvencí zůstávají behěm replikace v DNA chyby behěm replikace leading strandu zařadí polymeráza průměně 1 špatnou bázi z 10 milionů zreplikovaných (10-7), u lagging strandu 20x více (protože DNA PolI má má horší proofreading aktivitu). navíc může docházet i k ‘uklouznutí’ polymerázy (polymerase slipping): Polymeraza udela daleko vice chyb, ale vetsina je opravena = vysledkem 1 baze na replikaci celeho genomu zavedení delecí nebo inzercí
, u lagging strandu 20x více (protože DNA PolI má má horší proofreading aktivitu). navíc může docházet i k ‘uklouznutí’ polymerázy (polymerase slipping): Polymeraza udela daleko vice chyb, ale vetsina je opravena = vysledkem 1 baze na replikaci celeho genomu. zavedení delecí nebo inzercí.")
21
SPONTÁNNÍ MUTACE Mutace vzniklé chybami DNA polymerázy proofreading aktivita DNA polymerázy není dokonalá, s malou frekvencí zůstávají behěm replikace v DNA chyby behěm replikace leading strandu zařadí polymeráza průměně 1 špatnou bázi z 10 milionů zreplikovaných, u lagging strandu 20x více (protože DNA PolI má má horší proofreading aktivitu). navíc může docházet i k ‘uklouznutí’ polymerázy (polymerase slipping): Většina mutací hned opravena, výsledkem je méně než jedna fixovaná mutace po jednom buněčném dělení. Polymeraza udela daleko vice chyb, ale vetsina je opravena = vysledkem 1 baze na replikaci celeho genomu
. navíc může docházet i k ‘uklouznutí’ polymerázy (polymerase slipping): Většina mutací hned opravena, výsledkem je méně než jedna fixovaná mutace po jednom buněčném dělení. Polymeraza udela daleko vice chyb, ale vetsina je opravena = vysledkem 1 baze na replikaci celeho genomu.")
22
SPONTÁNNÍ MUTACE Mutace kvůli párování podobných úseků DNA a následné rekombinaci přímé repetice na DNA rekombinace (mezi dvěma totožnými nebo velmi podobnými sekvencemi) DELECE a DUPLIKACE DELECE
 DELECE a DUPLIKACE. DELECE.")
23
obrácené repetice na DNA
INVERZE

24
SPONTÁNNÍ MUTACE Mutace kvůli tautomerizaci bazí každá báze existuje jako keto a enol forma – tautomery ve vzájemné rovnováze, ale značně převažují keto formy pokud při replikaci báze zrovna v enol formě, bude zařazena v nesprávném párování

25
záměna C za T SPONTÁNNÍ MUTACE
Mutace kvůli spontánní chemické instabilitě bazí C, A a G mohou pomalu spontánně ztrácet své aminoskupiny = DEAMINACE 100 bazí za den v každé eukaryotní buňce! záměna C za T Cytosin je horkým místem mutací v DNA ! deaminace G a A mnohem vzácnější

26
5000 bazí za den v každé eukaryotní buňce!
SPONTÁNNÍ MUTACE Mutace kvůli spontánní chemické instabilitě bazí A a G se mohou spontánně hydrolyzovat od DNA kostry = DEPURINACE 5000 bazí za den v každé eukaryotní buňce!

27
Poškození neenzymatickou metylací
SPONTÁNNÍ MUTACE Poškození neenzymatickou metylací S malou frekvencí může donor metylačních skupin (S-adenosyl methionin) spontánně metylovat báze adenin 3-methyladenin 8-hydroxyguanin = 8-oxoguanin
 spontánně metylovat báze. adenin. 3-methyladenin. 8-hydroxyguanin = 8-oxoguanin.")
28
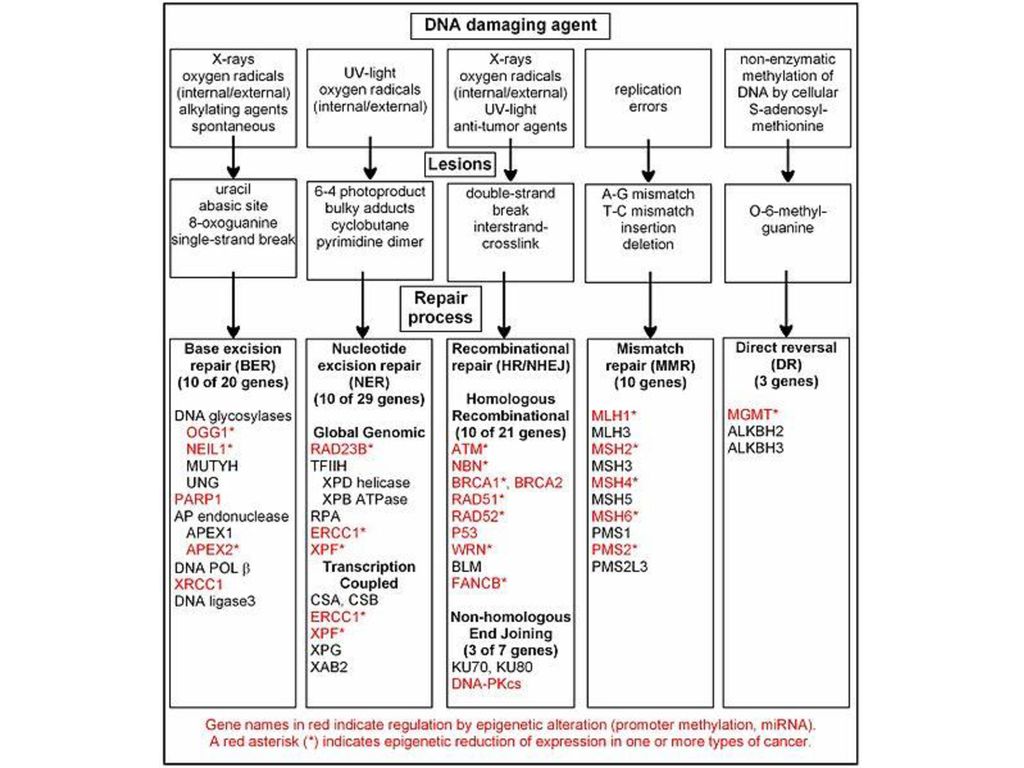
OPRAVY DNA Prokaryota

29
Chyby vzniklé nesprávnou replikací DNA
Proofreading aktivita DNA polymeráz se snaží eliminovat špatné párování během replikace Mismatch v párování bazí, jejich chemické modifikace nebo výskyt analogů bazí vede k distorci DNA Mismatch repair system (MMR) a nucleotide excision repair system (NER) detekují tyto změny struktury DNA spíše než specifické chemické změny O reparaci v rámci proofreading aktivity Pol se v této přednášce nezminujeme… Jak reparační systémy poznají, která ze dvou nesprávně spárovaných bazí je ta správná?
 a nucleotide excision repair system (NER) detekují tyto změny struktury DNA spíše než specifické chemické změny. O reparaci v rámci proofreading aktivity Pol se v této přednášce nezminujeme… Jak reparační systémy poznají, která ze dvou nesprávně spárovaných bazí je ta správná")
30
Jak reparační systémy poznají, která ze dvou nesprávně spárovaných bazí je ta správná?
Je třeba umět rozeznat při replikaci parentální DNA vlákno od nově replikovaného Bakteriální DNA metylázy metylují parentální vlákno DNA dam (DNA adenine methylase) - GATC dcm (DNA cytosine methylase) – CCAGG nebo CCTGG 6-methyladenin a 5-methylcytosin se párují správně, neindukují reparační odpověd’
 - GATC. dcm (DNA cytosine methylase) – CCAGG nebo CCTGG. 6-methyladenin a 5-methylcytosin se párují správně, neindukují reparační odpověd’")
31
dam a dcm metylázy metylují i nové vlákno
několik minut po replikaci je nová DNA šroubovice hemimetylovaná nastupují reparační enzymy a hledají chyby v nemetylovaném vlákně U Eukaryot strand discrimination signal jsou zlomy v DNA po Okazakiho fragmentech, co dalsiho? dam a dcm metylázy metylují i nové vlákno

32
DNA mismatch repair system
(MMR system) rozpoznání chybné báze pomocí MutL , MutS a MutH proteinů + rozpoznání starého a nového vlákna zavedení zlomu v novém DNA vlákně odstranění úseku DNA kolem chybného párování dosyntetozování správné báze DNA polymerázou III (nebo d u eukaryot)
 rozpoznání chybné báze pomocí MutL , MutS a MutH proteinů. + rozpoznání starého a nového vlákna. zavedení zlomu v novém DNA vlákně. odstranění úseku DNA kolem chybného párování. dosyntetozování správné báze DNA polymerázou III (nebo d u eukaryot)")
33
Nucleotide excision repair system (NER system)
nejčastější systém oprav poškozené DNA rozpoznává změny struktury DNA, ale méně senzitivní než MMR (změny struktury musejí být více nápadné) především opravy DNA poškozené UV = thyminové dimery, kroslinkované báze rozpoznání chybné báze pomocí uvrA,B,C proteinů zavedení dvou zlomů okolo T=T odstranění úseku DNA kolem chybného párování a dosyntetizování správné báze DNA polymerázou I
 především opravy DNA poškozené UV = thyminové dimery, kroslinkované báze. rozpoznání chybné báze pomocí uvrA,B,C proteinů. zavedení dvou zlomů okolo T=T. odstranění úseku DNA kolem chybného párování a. dosyntetizování správné báze DNA polymerázou I.")
34
Base excision repair system (BER system)
rozeznává specifické chemické změny v DNA, která se neprojeví na změně struktury DNA opravuje báze, které se normálně v DNA nevyskytují (není pochyb o tom, že jsou špatné) Deaminace adeninu na hypoxantin, guaninu na xanthin, cytosinu na uracil: DNA glykosyláza odštěpí chybnou bázi, vznikne místo bez báze (AP místo) AP endonukleáza štěpí kostru DNA DNA polymeráza I dostaví mezeru
 Deaminace adeninu na hypoxantin, guaninu na xanthin, cytosinu na uracil: DNA glykosyláza odštěpí chybnou bázi, vznikne místo bez báze (AP místo) AP endonukleáza štěpí kostru DNA. DNA polymeráza I dostaví mezeru.")
35
Oxidace guaninu na 8-oxoguanin
(při oxidativním stresu): MutT fosfatáza odstraňuje fosfátové skupiny z volných oxoGTP nukleotidů, aby se nemohly inkorporovat do DNA MutM glycosyláza odstraňuje oxoG z DNA (pokud se páruje správně s C) MutY glycosyláza odstraňuje A z DNA pokud se páruje s oxoG 8-hydroxyguanin = 8-oxoguanin Vzniklé AP místo dostavěné DNA polymerázou I
: MutT fosfatáza odstraňuje fosfátové skupiny z volných oxoGTP nukleotidů, aby se nemohly inkorporovat do DNA. MutM glycosyláza odstraňuje oxoG z DNA. (pokud se páruje správně s C) MutY glycosyláza odstraňuje A z DNA. pokud se páruje s oxoG. 8-hydroxyguanin = 8-oxoguanin. Vzniklé AP místo dostavěné DNA polymerázou I.")
36
Very short patch repair system
Thymin vzniklý spontánní deaminací 5-methyl cytosinu (ne během replikace DNA) je horkým místem mutací (změna CG na TA při následné replikaci) u E.coli je 5-methylcytosin hlavně v místech metylovaných dam a dcm pokud se T vyskytne v sekvencích rozeznávaných dam a dcm metylázami (CCAGG a CCTGG), je odstraněn Vsr endonukleázou Sebevražedné demetylázy Direct reversal (DR) Metylové skupiny na kyslíku u O6-methylguaninu a O4-methylthyminu odstraněny sebevražednými enzymy, které metylovou skupinu přenesou na sebe. Báze je tím rovnou opravena, ale enzym lze použít pouze jednou. Ostatní metylované báze odstraněny DNA glycosylázami
 je horkým místem mutací (změna CG na TA při následné replikaci) u E.coli je 5-methylcytosin hlavně v místech metylovaných dam a dcm. pokud se T vyskytne v sekvencích rozeznávaných dam a dcm metylázami (CCAGG a CCTGG), je odstraněn Vsr endonukleázou. Sebevražedné demetylázy. Direct reversal (DR) Metylové skupiny na kyslíku u O6-methylguaninu a O4-methylthyminu odstraněny sebevražednými enzymy, které metylovou skupinu přenesou na sebe. Báze je tím rovnou opravena, ale enzym lze použít pouze jednou. Ostatní metylované báze odstraněny DNA glycosylázami.")
37
Photoreaktivační systém
oprava thyminových dimerů (opravy také pomocí NER systému, Uvr proteiny) Přímá oprava, rozštěpení dimeru, bez použití polymerázy Fotolyáza – absorbuje viditelné světlo o vlnové délce nm (modré) a jeho energii používá rozštěpení thyminového dimeru = fotoreaktivace U bakterií, kvasinek, některých obratlovců, ale ne u člověka
 Přímá oprava, rozštěpení dimeru, bez použití polymerázy. Fotolyáza – absorbuje viditelné světlo o vlnové délce nm (modré) a jeho energii používá rozštěpení thyminového dimeru = fotoreaktivace. U bakterií, kvasinek, některých obratlovců, ale ne u člověka.")
38
Opravy pomocí rekombinace
Ne všechny chyby DNA se podaří odstranit před průběhem replikace. Pokud mutace brání postupu DNA polymerázy (například přítomnost thyminových dimerů), polymeráza odpadne a začne syntézu opět o kousek dál Vznikne jednořetězcová mezera v replikovaném chromozomu RecA – rekombinační protein vázající ssDNA Rekombinace mezi ‘zdravým’ vláknem na druhém chromozomu a vláknem s mezerou Mezera dostavena podle zdravého vlákna, thyminový dimer sice neopravený, ale oba chromozomy zreplikovány bez mezer
, polymeráza odpadne a začne syntézu opět o kousek dál. Vznikne jednořetězcová mezera. v replikovaném chromozomu. RecA – rekombinační protein vázající ssDNA. Rekombinace mezi ‘zdravým’ vláknem na druhém chromozomu a vláknem s mezerou. Mezera dostavena podle zdravého vlákna, thyminový dimer sice neopravený, ale oba chromozomy zreplikovány bez mezer.")
39
SOS reparační systém Pokud je DNA poškozená na mnoha místech a během replikace vzniká hodně jednovláknových oblastí, SOS systém umožní průběh replikace dosyntetizováním těchto úseků, i když to pravděpodobně zavede mnoho mutací RecA se aktivuje vazbou na ssDNA lexA dimer blokuje expresi SOS genů RecA rozštípe LexA represor lexA je zničen, spustí se exprese SOS genů např. DNA polymeráza V, nemá proofreading activitu, takže můž replikovat i thyminové dimery a místa bez bází (AP místa) Pouze u bakterií, pro Eukaryota příliš nebezpečné replikovat buňky s mnoha mutacemi (rakovina), raději apoptoza nebo inhibice dělení
 Pouze u bakterií, pro Eukaryota příliš nebezpečné replikovat buňky s mnoha mutacemi (rakovina), raději apoptoza nebo inhibice dělení.")
40
Oprava spřažená s transkripcí (transcription coupled repair)
Pokud je DNA hodně poškozená, může bránit průběhu transkripce, RNA polymeráza se zastaví, začne oprava templátového vlákna Bakterie: TRCF protein rozpoznává zastavenou polymerázu (transcription repair coupling factor) Eukaryota: TFIIH rozvolňuje DNA během transkripce, pokud’narazí na poškozenou DNA, naváže proteiny excision repair systému (NER)
 Eukaryota: TFIIH rozvolňuje DNA během transkripce, pokud’narazí na poškozenou DNA, naváže proteiny excision repair systému (NER)")
41
Opravy u Eukaryot Systémy podobné jako u prokaryot, ale méně prozkoumané Spontaneous instability

42
DNA damage checkpoints
Při každém dělení se na přechodech G1/S a G2/M a také v S kontroluje intaktnost DNA ATM kináza – detekuje dvojřetězcové zlomy ATR kináza – detekuje zastavené replikační vidličky Fosforylují celou řadu proteinů vedoucích k zastavení buněčného cyklu nebo apoptose, např. p53

43
Opravy dvojřetězcových zlomů
Non homologous end joining Zlomy dsDNA ionizačním zářením, chemicky nebo po vyštěpení transpozonů eukaryota Vazba Ku proteinů na konce dsDNA Vazba DNA-PK (DNA protein kinasy) Fosforylace XRCC4, díky tomu navázání ligázy a spojení DNA Může spojit omylem i DNA, která k sobě nepatří, chromozomální translokace dsDNA zlomy možno opravit též homologní rekombinací (viz dále)
 Fosforylace XRCC4, díky tomu navázání ligázy a spojení DNA. Může spojit omylem i DNA, která k sobě nepatří, chromozomální translokace. dsDNA zlomy možno opravit též homologní rekombinací (viz dále)")
45
Defekty v reparačních systémech způsobují dědičná onemocnění

46
Problémy s opravami DNA vedou ke zvýšené náchylnosti k rakovině a
k předčasnému stárnutí Xeroderma pigmentosum mutace v genech pro excision repair system, vysoká senzitivita kůže k UV záření autosomálně recesivní incidence 1: v Evropě, 1: v Japonsku rakovina kůže okolo 8.roku života Dědičný non-polypósní kolorektální karcinom 5-7% případů kolorektálního karcinomu autosománě dominantní projevy relativně brzo (průměr 44 let) mutace v mismatch repair systému (MLH1, MSH2) zvýšená náchylnost i k jiným typům rakoviny
47
Wernerův syndrom Předčasné stárnutí způsobené mutací ve Wnr genu – specifické helikázy používané při opravách DNA

48
Dědičná rakovina prsu a vaječníků
BRCA1 a BRCA2 gen (breast cancer A1 a A2) opravují zlomy v dsDNA autosomálně dominantní dědičnost (lze zdědit i od otce) 40% nositelů vyvine rakovinu prsu do věku 70 let (ženy) Náchylnost u k rakovine prsu u muzu mnohem vyssi nez by bylo normalne, ale porad to da jen asi 1% muzu s BRCA1 mutaci.
 opravují zlomy v dsDNA. autosomálně dominantní dědičnost (lze zdědit i od otce) 40% nositelů vyvine rakovinu prsu do věku 70 let (ženy) Náchylnost u k rakovine prsu u muzu mnohem vyssi nez by bylo normalne, ale porad to da jen asi 1% muzu s BRCA1 mutaci.")
49
Horká místa mutací Ne všechna místa v genomu jsou stejně náchylná k mutacím, některá místa mají frekvenci daleko vyšší. Vetšinou místa výskytu 5-methylcytosinu, který občas spontánně deaminuje na thymin a páruje se pak s A místo s G.

50
Fenotyp vzniklý určitou mutací je vrácen do normálního stavu
Reverze fenotypu Fenotyp vzniklý určitou mutací je vrácen do normálního stavu Šance, že by daný nukleotid spontánně zmutoval zpět na původní nukleotid je velmi malá Mnohem častěji je reverze fenotypu způsobena jinou mutací, která vyruší účinek první = supresorová mutace (ať už ve stejném genu nebo v jiném)
")
51
Supresorové tRNA nabité tRNA, které ale mají zmutovaná antikodon, takže rozeznává STOP kodon (např. tRNA pro glycin, kde antikodon GAC zmutoval na AUC) Mohou částečně obnovit expresi genů s předčasným stop kodonem, běžné u prokaryot a kvasinek Pouze pokud existuje více tRNA pro tutéž aminokyselinou, jinak by mutace kodonu byla letální Důsledkem je prodloužení i normálních proteinů, což může být problém.
 Mohou částečně obnovit expresi genů s předčasným stop kodonem, běžné u prokaryot a kvasinek. Pouze pokud existuje více tRNA pro tutéž aminokyselinou, jinak by mutace kodonu byla letální. Důsledkem je prodloužení i normálních proteinů, což může být problém.")
52
Konec

54
Přestávka

55
REKOMBINACE

56
zlom chromozomu a znovuspojení
Eukaryota Rekombinace Výměna genetického materiálu mezi chromozomy nebo nebo mezi jinými molekulami DNA U eukaryot při meioze, při opravách DNA, u bakterií při konjugaci plazmidu homologní chromozómy synapse zlom chromozomu a znovuspojení crossing over – zlom v DNA a její spojení s jinou molekulou (zde dvojitý crossing over)
")
57
Rekombinace především během meiozy
Lepto, zygo atd = stadia profaze prvniho deleni, po S fazi, pak metafaze, anafaze, telofaze… Musí se zavést zlomy v dsDNA = riskantní! Rekombinace během mitózy – k opravě zlomů v dsDNA nebo jednovláknových mezer v dsDNA

58
(u meiozy díky Spo11 nukleáze)
Dvojvláknový zlom (u meiozy díky Spo11 nukleáze) Vznik volného 3’ konce, který se pokrývá Rad51 proteiny Invaze 3’ vlákna do sesterské chromatidy, přechodný vznik triple DNA a následně D-smyčky Dostavění a ligace zbylých konců, vznik Holliday junction 2 teorie, jak oprava double stranded break: DSBR: double strand break repair (Holiday junction model) SDSA: synthesis-dependent strand annealing Následuje další štěpení a separace vláken, může a nemusí vést k výměně celých ramen na chromozomu:
 Vznik volného 3’ konce, který se pokrývá Rad51 proteiny. Invaze 3’ vlákna do sesterské chromatidy, přechodný vznik triple DNA a následně. D-smyčky. Dostavění a ligace zbylých konců, vznik Holliday junction. 2 teorie, jak oprava double stranded break: DSBR: double strand break repair (Holiday junction model) SDSA: synthesis-dependent strand annealing. Následuje další štěpení a separace vláken, může a nemusí vést k výměně celých ramen na chromozomu:")
59
Holliday junction se může otáčet do dvou izomerních stavů
Patch recombinant Vymění se pouze část jednoho vlákna resolváza (rekombináza) – štěpí a liguje DNA v Holliday junction True recombinant Vymění se celý zbytek chromozomu
 – štěpí a liguje DNA v Holliday junction. True recombinant. Vymění se celý zbytek chromozomu.")
60
Crossing over / video

61
rekombinace DNA během meiózy
Genová konverze během meiozy Dvě alely téhož genu, R a r Přeměna jedné alely genu v druhou během rekombinace a následné pravy DNA rekombinace DNA během meiózy Pokud meioza proběhne před opravou DNA, vytvoří se normálně dvě R a dvě r Jinak ale reparační systémy opraví DNA náhodně, buď podle jedné alely nebo podle druhé Někdy se DNA opraví tak, že vzniknou 4 alely téhož genu, druhá alela zanikne r,r,r,r R,R,R,R

62
invaze jednořetězcového vlákna
Prokaryota 1. zlom v dsDNA a invaze jednořetězcového vlákna 2. RecBCD komplex nasedá na zlomy dsDNA 3. Postupuje po DNA, dokud nenarazí na Chi sekvenci (GCTGGTGG, časté) 4. RecC odštěpí jeden z řetězců DNA Zlomy v dsDNA = většinou ne zlomy v genomické DNA, ale cizorodá DNA vniklá do buňky při transformaci, transdukci viry, konjugaci 5. RecBC postupně odhaluje ssDNA s volným 3’koncem, ta se pokrývá RecA proteiny:
 4. RecC odštěpí jeden z řetězců DNA. Zlomy v dsDNA = většinou ne zlomy v genomické DNA, ale cizorodá DNA vniklá do buňky při transformaci, transdukci viry, konjugaci. 5. RecBC postupně odhaluje ssDNA s. volným 3’koncem, ta se pokrývá. RecA proteiny:")
63
ssDNA vlákno s volným 3’koncem se pokrývá RecA proteiny
Zlomy v dsDNA = většinou ne zlomy v genomické DNA, ale v cizorodé DNA vniklé do buňky při transformaci, transdukci viry, konjugaci ssDNA vlákno s volným 3’koncem se pokrývá RecA proteiny … a atakuje dsDNA homologní DNA molekuly Dočasné vytvoření triple helixu Vytěsněné ssDNA vlákno se pak spojí s ssDNA druhé molekuly

64
Homologní rekombinace - mezi dvěma velmi podobnými skevencemi
Nehomologní (místně specifická) rekombinace – mezi jinak nepodobnými sekvencemi, ale iniciováno na krátkém úseku homologie rozpoznávaném specifickými proteiny
 rekombinace – mezi jinak nepodobnými sekvencemi, ale iniciováno na krátkém úseku homologie rozpoznávaném specifickými proteiny.")
65
Místně specifická rekombinace (nehomologní)
Pouze krátké místo homologie mezi dvěma jinak rozdínými molekulami; pro rekombinaci musí být toto místo rozpoznáno specifickými proteiny např. integrace bakteriofágu lambda do chromozomu E.coli attP místa v bakteriofágovi, attB místa v bakterii, pouze oblast attO je uplně homologní Integráza z bakteriofágu štěpí dsDNA

66
See WIKI for a nice overview of repair!
Site directed mutageness, str 763, nezminuju… Mutace pri maturaci protilatek…

Podobné prezentace






>")








.>")




