Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Genom je dynamický

2
Mobilní elementy jako významná část lidského genomu

3
Mobilní elementy

4
Mobilní elementy představují ca necelou polovinu lidského genomu.
20 až > kopií

6
RETROELEMENTY RNA svět -----> DNA svět
význam reverzní transkripce (při vzniku DNA i dnes) retroelementy jsou relikty RNA světa (struktura, replikační mechanizmy, všudypřítomnost) úloha v evoluci genomů, RT je nejčastější gen kódující protein v lidském genomu
 retroelementy jsou relikty RNA světa (struktura, replikační mechanizmy, všudypřítomnost) úloha v evoluci genomů, RT je nejčastější gen kódující protein v lidském genomu.")
7
Retroelementy Retropozice
jsou sekvence DNA nebo RNA obsahující gen pro enzym reverzní transkriptázu (přepis RNA do DNA). Širší definice zahrnuje veškeré sekvence vzniklé reverzní transkripcí RNA do DNA. Po genomech se šíří procesem retropozice. Retropozice je přesun genetického materiálu z jednoho místa v genomu do místa jiného prostřednictvím RNA intermediátu. Má vždy duplikativní charakter.
. Širší definice zahrnuje veškeré sekvence vzniklé reverzní transkripcí RNA do DNA. Po genomech se šíří procesem retropozice. Retropozice. je přesun genetického materiálu z jednoho místa v genomu do místa jiného prostřednictvím RNA intermediátu. Má vždy duplikativní charakter.")
8
Retropozice

9
Reverzní transkripce - nasednutí primeru tRNA a extenze
- odstranění RNA oblasti R a U5 - první přeskok a extenze - odstranění většiny RNA RNázouH - zbyde primer a extenze - odstranění zbytku virové RNA a tRNA - druhý přeskok a dosyntetizování

10
Typy retroelementů

11
Endogenní retroviry • Struktura podobná retrovirům.
• Pozůstatky dávných retrovirových infekcí. • HERV (human endogenous retrovirus).
.")
12
Retroviry - obecné schéma
pol gag LTR env LTR > PR RT RNázaH INT > PBS PPT gag, pol, env geny LTR ... dlouhé koncové repetice PBS .... místo vazby primeru PR doména kódující proteázu INT .... doména kódující integrázu RT doména kódující reverzní transkriptázu PPT polypurinový úsek > přímé repetice LTR U3 R U5 U3 ... jedinečná sekvence na 3‘ konci R repetitivní sekvence U5 ... jedinečná sekvence na 5‘ konci krátká obrácená repetice krátká obrácená repetice promotor polyadenylační signál zesilovače transkripce

13
Retrotranspozony - obecné schéma
(a) Typ Ty1-copia: gag pol LTR LTR > > PBS PR INT RT RNázaH PPT gag, pol, env geny LTR ... dlouhé koncové repetice PBS .... místo vazby primeru PR doména kódující proteázu INT .... doména kódující integrázu RT doména kódující reverzní transkriptázu PPT polypurinový úsek > přímé repetice (b) Typ Ty3-gypsy: pol gag LTR env? LTR > > PBS PR RT RNázaH INT PPT
 Typ Ty1-copia: gag. pol. LTR. LTR. > > PBS. PR INT RT RNázaH. PPT. gag, pol, env geny. LTR ... dlouhé koncové repetice. PBS .... místo vazby primeru. PR doména kódující proteázu. INT .... doména kódující integrázu. RT doména kódující reverzní transkriptázu. PPT polypurinový úsek. > přímé repetice. (b) Typ Ty3-gypsy: pol. gag. LTR. env LTR. > > PBS. PR RT RNázaH INT. PPT.")
14
Retrotranspozony • Struktura podobná retrovirům.
• Obsahují dlouhé koncové repetice (LTR) nesoucí regulační sekvence a promotory. • Ty1-copia, Ty-3gypsy. • U všech eukaryot. Hojné především u rostlin. U obratlovců méně časté. U bobu setého cca 1 milion kopií elementu Ty1-copia, tvoří polovinu genomu.
 nesoucí regulační sekvence a promotory. • Ty1-copia, Ty-3gypsy. • U všech eukaryot. Hojné především u rostlin. U obratlovců méně časté. U bobu setého cca 1 milion kopií elementu Ty1-copia, tvoří polovinu genomu.")
15
Retropozony (polyA, nonLTR retroelementy)
(a) LINE (L1): ORF1 ORF2 5’UTR 3‘UTR > > poly(A) EN RT PPT 1 kb ORF protein vážící RNA EN doména kódující endonukleázu RT doména kódující reverzní transkriptázu 5‘UTR netranslatovaná oblast na 5‘ konci 3‘UTR netranslatovaná oblast na 3‘ konci PPT polypurinový úsek poly(A) polyadeninový úsek > přímé repetice (b) SINE (sekvence Alu): > > levý monomer poly(A) pravý monomer poly(A) 100 bp
 LINE (L1): ORF1. ORF2. 5’UTR. 3‘UTR. > > poly(A) EN RT. PPT. 1 kb. ORF protein vážící RNA. EN doména kódující endonukleázu. RT doména kódující reverzní transkriptázu. 5‘UTR netranslatovaná oblast na 5‘ konci. 3‘UTR netranslatovaná oblast na 3‘ konci. PPT polypurinový úsek. poly(A) polyadeninový úsek. > přímé repetice. (b) SINE (sekvence Alu): > > levý monomer. poly(A) pravý monomer. poly(A) 100 bp.")
16
Retropozony • Nemají LTR. Poly(A) na 3’ konci.
• U všech eukaryot. Nejčastější retrolelementy u člověka a savců. • LINE (Long interspersed nuclear elements) kódují vlastní reverzní transkriptázu. • SINE (Short interspersed nuclear elements) Nemají vlastní reverzní transkriptázu. Jejich šíření závislé na přítomnosti LINE elementů. • Nejznámější SINE je Alu sekvence: nejhojnější retroelement u lidí (1,5 milionů kopií, 13% genomu). Vznikla pravděpodobně z genu pro tRNA 7SLRNA (podobně jako geny pro tRNA nese vnitřní promotor přepisovaný RNA polymerázou III).
 kódují vlastní reverzní transkriptázu. • SINE (Short interspersed nuclear elements) Nemají vlastní reverzní transkriptázu. Jejich šíření závislé na přítomnosti LINE elementů. • Nejznámější SINE je Alu sekvence: nejhojnější retroelement u lidí. (1,5 milionů kopií, 13% genomu). Vznikla pravděpodobně z genu pro tRNA 7SLRNA (podobně jako geny pro tRNA nese vnitřní promotor přepisovaný RNA polymerázou III).")
17
Včleňování LINE elementů
Twin priming: mechanizmus tvorby inverzí TPRT (target-primed RT)
")
18
LINE elementy v genomu člověka
- vliv na velikost genomu u primátů (struktura) - místa homologií pro genovou konverzi a rekombinaci - včleňuje se do genu a narušují kódující sekvence, nemoci (funkce) - přeskupování oblastí, exon shufling, zkráceny na 5’ konci - nedosyntetizovány - zacelují místa chromosomových zlomů (reparace) - nedávná aktivita u primátů, rozdíly mezi druhy i uvnitř druhů (evoluce) Včleňování mechanizmem TPRT (target primed reverse transcription)
 - místa homologií pro genovou konverzi a rekombinaci. - včleňuje se do genu a narušují kódující sekvence, nemoci (funkce) - přeskupování oblastí, exon shufling, zkráceny na 5’ konci - nedosyntetizovány. - zacelují místa chromosomových zlomů (reparace) - nedávná aktivita u primátů, rozdíly mezi druhy i uvnitř druhů (evoluce) Včleňování mechanizmem TPRT (target primed reverse transcription)")
19
LINE elementy a inaktivace chromosomu X
- 2x větší hustota LINE-1 (L1) elementů na chromosomu X - nejvíce v místě centra inaktivace chromosomu X (Xq13-Xq21) - geny, které unikají inaktivaci (10%) obsahují méně L1 elementů - L1 slouží k šíření inaktivačního signálu z centra (Xist gen) - po připojení autosomu ke chromosomu X se signál šíří po autosomu do 100Mb - přenesení genu Xist na autosom vede k šíření signálu – Xist RNA pokryje i autosom - L1 jako vazebná místa Xist RNA!!! - u drozofily za šíření signálu kompenzace dávky genu (zvýšení exprese samčího X) také zodpovídají RNA-proteinové komplexy - L1 elementy jako zesilovače/přenašeče (booster, way station) šíření inaktivačního signálu - zejména zvýšená hustota L1 mladších 100 mil let – vznik inaktivace chromosomu X (XCI) v linii savců před oddělením Metatheria a Eutheria
 elementů na chromosomu X. - nejvíce v místě centra inaktivace chromosomu X (Xq13-Xq21) - geny, které unikají inaktivaci (10%) obsahují méně L1 elementů. - L1 slouží k šíření inaktivačního signálu z centra (Xist gen) - po připojení autosomu ke chromosomu X se signál šíří po autosomu do 100Mb. - přenesení genu Xist na autosom vede k šíření signálu – Xist RNA pokryje i autosom. - L1 jako vazebná místa Xist RNA!!! - u drozofily za šíření signálu kompenzace dávky genu (zvýšení exprese samčího X) také zodpovídají RNA-proteinové komplexy. - L1 elementy jako zesilovače/přenašeče (booster, way station) šíření inaktivačního signálu. - zejména zvýšená hustota L1 mladších 100 mil let – vznik inaktivace chromosomu X (XCI) v linii savců před oddělením Metatheria a Eutheria.")
20
Chromosom X – křižovatka pohybu genů
1. Únik genů z chromosomu X je ovlivněn jeho inaktivací (Xi): - meiotická inaktivace pohlavních chromosomů (MSCI) – sex chromosomu kondenzovány v samčí meióze, „sex body“, „XY body“, aby nedocházelo k rekombinaci mezi nehomologickými chromosomy, geny na XY tedy transkripčně suprimovány - proto backup genů na autosomech, přenos retropozicí (compensatory hypothesis) 2. Přenos genů na chromosom X: - přednostní včleňování i selekce - L1 elementy i retropseudogeny, L1 mašinérie - recesivní geny výhodné pro samce a dominantní geny výhodné pro samice A A A X A A A
: - meiotická inaktivace pohlavních chromosomů (MSCI) – sex chromosomu kondenzovány v samčí meióze, „sex body , „XY body , aby nedocházelo k rekombinaci mezi nehomologickými chromosomy, geny na XY tedy transkripčně suprimovány. - proto backup genů na autosomech, přenos retropozicí (compensatory hypothesis) 2. Přenos genů na chromosom X: - přednostní včleňování i selekce. - L1 elementy i retropseudogeny, L1 mašinérie. - recesivní geny výhodné pro samce a dominantní geny výhodné pro samice. A. A. A. X. A. A. A.")
21
Alu elementy jsou nejhojnější retroelementy lidského genomu
odvozeny z 7SL RNA genu kódujícího podjednotku signální rozpoznávací částice (přenos proteinů přes membrány a začleňování do membrán) - Alu inzerce – u každého 200 narozeného jedince
 - Alu inzerce – u každého 200 narozeného jedince.")
22
Alu elementy tvoří ~10% lidského genomu (>1 milion).
5% lidských alternativních exonů je odvozeno z Alu elementů.

23
Alu elementy a alternativní sestřih
% lidských genů má alternativní sestřih rozdíl mezi člověkem a myší: genů, fenotypové rozdíly způsobeny druhově-specifickým alterantivním sestřihem exony – konstitutivní (konzervativní) a alternativní alternativní exony – majoritní a minoritní formy
 a alternativní. alternativní exony – majoritní a minoritní formy.")
24
Vznik alternativních exonů
duplikace exonů, inzerce transposonu nebo retroviru, mutace existující intronové sekvence, z Alu elementů (5% lidských alternativních exonů)
")
25
Retroelementy prokaryot
Jsou velmi staré. Kódují jediný RT protein s jedinou enzymatickou aktivitou. Ostatní aktivity (proteáza, integráza, endonukleáza) u retrotransposonů a retrovirů byly získány od hostitele jako onkogeny retrovirů. RT prokaryot provádí syntézu nezávislou na primeru podobně jako RNA polymeráza (předchůdce RT). Prokaryotické RT jsou podobné RdRP (RNA-directed RNA polymerase), RT eukaryotických retroelementů jsou méně příbuzné. Telomeráza, starý eukaryotický enzym, je podobná prokaryotickým RT a RT retroposonů.
 u retrotransposonů a retrovirů byly získány od hostitele jako onkogeny retrovirů. RT prokaryot provádí syntézu nezávislou na primeru podobně jako RNA polymeráza (předchůdce RT). Prokaryotické RT jsou podobné RdRP (RNA-directed RNA polymerase), RT eukaryotických retroelementů jsou méně příbuzné. Telomeráza, starý eukaryotický enzym, je podobná prokaryotickým RT a RT retroposonů.")
26
Retroelementy prokaryot
Typy 1. Retrony 2. Retroplasmidy 3. Retrointrony (introny II. skupiny)
")
27
Retrony - primitivní retroelementy bakterií
(b) msDNA: (a) retron: RT msr msd msr ... gen kódující RNA složku msd ... gen kódující DNA složku RT gen pro reverzní transkriptázu
 msDNA: (a) retron: RT. msr. msd. msr ... gen kódující RNA složku. msd ... gen kódující DNA složku. RT gen pro reverzní transkriptázu.")
28
Retrony • Bakteriální retroelementy.
• Chiméry tvořené molekulami DNA a RNA.

29
Další primitivní retroelementy
Mitochondriální (retro)plazmidy dsDNA plazmidy, v mitochondriích hub, kódují RT, replikace procesem transkripce-reverzní transkripce, homologie s tRNA, sekundární struktury na 3‘konci, hairpiny. Introny II. skupiny U bakterií a v organelách hub, řas a rostlin, kódují RT, samosestřih, „reverzní sestřih“ do RNA bez intronu a reverzní transkripce. Telomery U eukaryot, kompenzace zkracování chromozomů, telomeráza - homologie s RT, vlastní RNA templát.
plazmidy. dsDNA plazmidy, v mitochondriích hub, kódují RT, replikace procesem transkripce-reverzní transkripce, homologie s tRNA, sekundární struktury na 3‘konci, hairpiny. Introny II. skupiny. U bakterií a v organelách hub, řas a rostlin, kódují RT, samosestřih, „reverzní sestřih do RNA bez intronu a reverzní transkripce. Telomery. U eukaryot, kompenzace zkracování chromozomů, telomeráza - homologie s RT, vlastní RNA templát.")
30
DNA transpozony • Hojné především u prokaryot.
• Přesunují se mechanismem „cut and paste“ nebo „copy-paste“. • Kódují enzym transpozázu. Často navíc také geny pro rezistenci proti antibiotikům. Lemované obrácenými repeticemi. • IS elementy a Tn elementy u prokaryot. P elementy u drozofily. Ac a Ds elementy u kukuřice (Barbara McClintock).
.")
31
Charakteristické rysy mobilních DNA elementů
• Nesou si vlastní regulační sekvence (promotory, enhancery). Nejsou závislé na přítomnosti cizích promotorů v místě integrace. Mohou naopak ovlivňovat transkripci sousedních genů. • Většinou v genomu pouze malé množství celých nepoškozených elementů. Většinu tvoří nefunkční, zkrácené či jinak poškozené elementy. Ale i poškozené elementy se mohou kopírovat pokud v genomu alespoň několik nepoškozených kopií, které dodávají potřebné enzymy.
. Nejsou závislé na přítomnosti cizích promotorů v místě integrace. Mohou naopak ovlivňovat transkripci sousedních genů. • Většinou v genomu pouze malé množství celých nepoškozených elementů. Většinu tvoří nefunkční, zkrácené či jinak poškozené elementy. Ale i poškozené elementy se mohou kopírovat pokud v genomu alespoň několik nepoškozených kopií, které dodávají potřebné enzymy.")
32
Vznik a evoluce retroelementů
Původní genomy byly tvořeny molekulami RNA myšlenka RNA jako předchůdce DNA všudypřítomnost RNA (probírali jsme v tématu relikty světa RNA) objev ribozymu - uložení i realizace informace reverzní transkriptáza, homologie s RNA replikázami a telomerázami všudypřítomnost retroelementů, různá struktura a mechanizmus replikace, tRNA a sekundární struktury při převedení informace z RNA do proteinů i do DNA
 objev ribozymu - uložení i realizace informace. reverzní transkriptáza, homologie s RNA replikázami a telomerázami. všudypřítomnost retroelementů, různá struktura a mechanizmus replikace, tRNA a sekundární struktury při převedení informace z RNA do proteinů i do DNA.")
33
Význam mobilních DNA elementů
Dřívější představa Představují pouze zbytečné sekvence „junk DNA“. Parazitická DNA, která genomu jen škodí, nepřináší žádný užitek. Negativní vliv mobilních DNA elementů na hostitele • Inzerční mutageneze. • Ovlivnění genové exprese sousedních genů (zvýšení exprese genů díky silným promotorům či naopak snížení exprese genů v důsledku heterochromatinizace). • Ektopická rekombinace mezi elementy v různých částech genomu může vést ke vzniku chromosomových přestaveb či rozsáhlých delecí či duplikací. • Způsobují zvětšování genomu. Způsobuje energenickou zátěž pro buňku. • U člověka 1% mutací kvůli přeskokům DNA elementů, u drosofily až 50%
. • Ektopická rekombinace mezi elementy v různých částech genomu může vést ke vzniku chromosomových přestaveb či rozsáhlých delecí či duplikací. • Způsobují zvětšování genomu. Způsobuje energenickou zátěž pro buňku. • U člověka 1% mutací kvůli přeskokům DNA elementů, u drosofily až 50%")
34
Význam mobilních DNA elementů
Dnešní představa Může docházet ke vzájemnému přizpůsobení transpozonů a jejich hostitele. V některých případech mohou transpozony poskytnout i užitečné nové funkce pro svého hostitele (domestikace transpozonů). Činí genom dynamičtějším.
. Činí genom dynamičtějším.")
35
Molekulární domestikace transpozonů
• Náhrada činnosti telomerázy u Drosophila melanogaster (non- LTR retrotranspozony Het-A, TART, TAHRE). • TERT – telomerická reverzní transkriptáza (proteinová enzymatická komponenta telomerázy), příbuzná RT od L1 • CENP-B protein – váže se na centromery, váže mikrotubuly dělícího vřeténka. blízce příbuzný transpozáze pogo elementu u drozofily (případně Tigger elementu u člověka) • Syncytin – odvozen od env z endogenního retroviru HERV –W. Důležitý pro utváření placenty. • Geny pro miRNA. • Alu a L1 elementy obsahují mnoho CpG míst, které mohou být metylovány - Alu a L1 jsou pravděpodobně hlavními zdroji CpG ostrovů v lidském genomu.
. • TERT – telomerická reverzní transkriptáza (proteinová enzymatická komponenta telomerázy), příbuzná RT od L1. • CENP-B protein – váže se na centromery, váže mikrotubuly dělícího vřeténka. blízce příbuzný transpozáze pogo elementu u drozofily (případně Tigger elementu u člověka) • Syncytin – odvozen od env z endogenního retroviru HERV –W. Důležitý pro utváření placenty. • Geny pro miRNA. • Alu a L1 elementy obsahují mnoho CpG míst, které mohou být metylovány - Alu a L1 jsou pravděpodobně hlavními zdroji CpG ostrovů v lidském genomu.")
36
Pozitivní vliv mobilních DNA elementů na hostitele
Regulace mutační rychlosti v závislosti na stresu • Transpozony mohou být aktivovány v odpovědi na nějaké vnější podmínky –stres, UV záření, teplotu, radioaktivní záření, zranění, infekci patogenem, polyploidizaci, mezidruhovou hybridizaci • Vznik nových mutací, mezi nimiž mohou být nové varianty schopné daný stres přežít • Adaptivní odpověď, strategie pro zvýšení variability a následnou selekci Tam1 , Tam3 u hledíku (1000x větší aktivita při 15°C) Reme1 u melounu (aktivace UV)
 Reme1 u melounu (aktivace UV)")
37
DNA mobilní elementy činí genom dynamičtějším
• Podporují duplikaci genů. Díky přítomnosti reverzní ranskriptázy vznikají v genomu pseudogeny. Důležitý proces při evoluci nových genů. • Umožňují přestavby genomu (translokace, inverze). • „Exon shuffling“ pomocí Helitronů. Helitrony = DNA transpozony, replikativní transpozice pomocí mechanizmu valivé kružnice (obsahují různé fragmenty genů – možnost vzniku chimérických genů)
. • „Exon shuffling pomocí Helitronů. Helitrony = DNA transpozony, replikativní transpozice pomocí. mechanizmu valivé kružnice. (obsahují různé fragmenty genů – možnost vzniku chimérických genů)")
38
Hledání společného předka všech retroelementů
příbuznost retroelementů, evoluční strom progenitor - jednoduchá struktura a všudypřítomnost dvě alternativní hypotézy: (a) retrony (nejjednodušší, codon usage - včleněny dávno) (b) retropozony (největší diverzita sekvencí RT, všudypřítomné)
 retrony (nejjednodušší, codon usage - včleněny dávno) (b) retropozony (největší diverzita sekvencí RT, všudypřítomné)")
39
Dvě hlavní evoluční větve retroelementů
první větev: retroelementy bez LTR, jednodušší struktura druhá větev: retroelementy s LTR, složitější struktura, příbuznější jsou retroviry, Ty3-gypsy a caulimoviry, vzdálenější jsou Ty1-copia a hepadnaviry

40
Evoluce retroelementů
gen pro RNA replikázu gag + INT gag RT INT RT první větev RNA virus LTR bakteriální genom mitochond. plazmid intron Hepadnaviry druhá větev Retropozony env ? RT RT Retrotranspozony Ty1-copia Introny II. skupiny Retrony RT Mitochondriální plazmidy RNA virus Retrotranspozony Ty3-gypsy Retroviry Caulimoviry

41
Chromozomální distribuce retroelementů
Vyskytují se na všech chromozomech (hybridizace in situ). Existují místa s vyšší koncentrací retroelementů i bez nich. Retroelementy jsou přítomny v heterochromatinu i euchromatinu, hřbitovy RE. Sex chromozomy – zjištěna akumulace na chromozomu Y u D. miranda, Cannabis sativa, Marchantia polymorpha .
. Existují místa s vyšší koncentrací retroelementů i bez nich. Retroelementy jsou přítomny v heterochromatinu i euchromatinu, hřbitovy RE. Sex chromozomy – zjištěna akumulace na chromozomu Y u D. miranda, Cannabis sativa, Marchantia polymorpha .")
42
Inzerce retroelementů do již existujících

43
Inzerční specificita a inzerce retroelementů do heterochromatinu
- Inzerce není náhodná (retroviry) - Role IN – targed domain (TD) - Interakce s proteiny chromatinu - inženýrství (nové specifity) - genová terapie integráza heterochromatin
 - Role IN – targed domain (TD) - Interakce s proteiny chromatinu. - inženýrství (nové specifity) - genová terapie. integráza. heterochromatin.")
44
Metoda „transposon display“ – detekce nových inzercí transposonů
- Indukce transpozice stresem - Štěpení genomické DNA - PCR okolní oblasti - Nové bandy

45
Funkce retroelementů I. Koadaptace retroelementů a hostitele
Alternativní hypotézy - parazité x významný činitel v evoluci, jsou slučitelné, koadaptace, vliv na hostitele. Pravidlo 3K: konflikt-kompromis-kooperace (ekologie genomu). Mutabilita – stochastické, ale regulované. Místo inzerce retroelementu do genomu - exony, introny, regulační sekvence, LTR, RE, sekvenční specificita. Regulace počtu kopií retroelementů v genomu - obranné mechanizmy hostitele, metylace, rekombinace.
. Mutabilita – stochastické, ale regulované. Místo inzerce retroelementu do genomu - exony, introny, regulační sekvence, LTR, RE, sekvenční specificita. Regulace počtu kopií retroelementů v genomu - obranné mechanizmy hostitele, metylace, rekombinace.")
46
II. Negativní vliv retroelementů na hostitele
Sobecká a parazitická DNA. Choroby (hemofilie, rakovina), L1, Alu. Příčinou inzerce nebo rekombinace (mezi Alu). III. Pozitivní vliv retroelementů na hostitele Přestavby genomu, rekombinace. Duplikace, genové rodiny, vznik nových genů (SETMAR aj.). Telomery drozofily. Imunitní systém.
, L1, Alu. Příčinou inzerce nebo rekombinace (mezi Alu). III. Pozitivní vliv retroelementů na hostitele. Přestavby genomu, rekombinace. Duplikace, genové rodiny, vznik nových genů (SETMAR aj.). Telomery drozofily. Imunitní systém.")
47
Poškození genomu transposony
Rekombinační přestavby Antisense RNA Inzerční mutageneze Umlčení způsobené metylací Chimerická mRNA

48
Helitrony Jsou nedávno objevené transposony.
Využívají mechanismus otáčející se kružnice.

49
Jsou rostlinné ekvivalenty lidských Alu.
MITE elementy Jsou rostlinné ekvivalenty lidských Alu. Neautonomní elementy (master = DNA TE Mariner). Jsou malé, bp. Obrácené koncové repetice (TIR) - konzervativní bp. AT-bohaté, tvoří sekundární struktury DNA (hairpins). Preference cílového místa TA(A). Asociace s geny v intronech, poblíž 5‘ nebo 3‘ konců genů.
. Jsou malé, bp. Obrácené koncové repetice (TIR) - konzervativní bp. AT-bohaté, tvoří sekundární struktury DNA (hairpins). Preference cílového místa TA(A). Asociace s geny v intronech, poblíž 5‘ nebo 3‘ konců genů.")
50
RYCHLÁ AMPLIFIKACE TRANSPOSONů V EVOLUCI HOMINIDů

51
Korelace mezi expanzemi transposonů a oddělováním větví savců

52
Korelace mezi expanzemi transposonů a oddělováním větví savců
Alu a LINE-1 existují v našem genomu po dlouhou dobu ~80 MY ~150 MY

53
Lidský genom je plný starých transposonů zatímco transposony v jiných genomech jsou mladší
Srovnání stáří transpozonů v eukaryotických genomech

54
Různá místa včlenění retroelementů u různých větví primátů
L1HS480 lokus člověk Retroviry PTERV1: - u Afrických (šimpanz a gorila) - chybí u člověka a Asijských (orangutan, gibon) lemuři
 - chybí u člověka a Asijských (orangutan, gibon) lemuři.")
55
Obranné mechanismy šíření DNA mobilních elementů
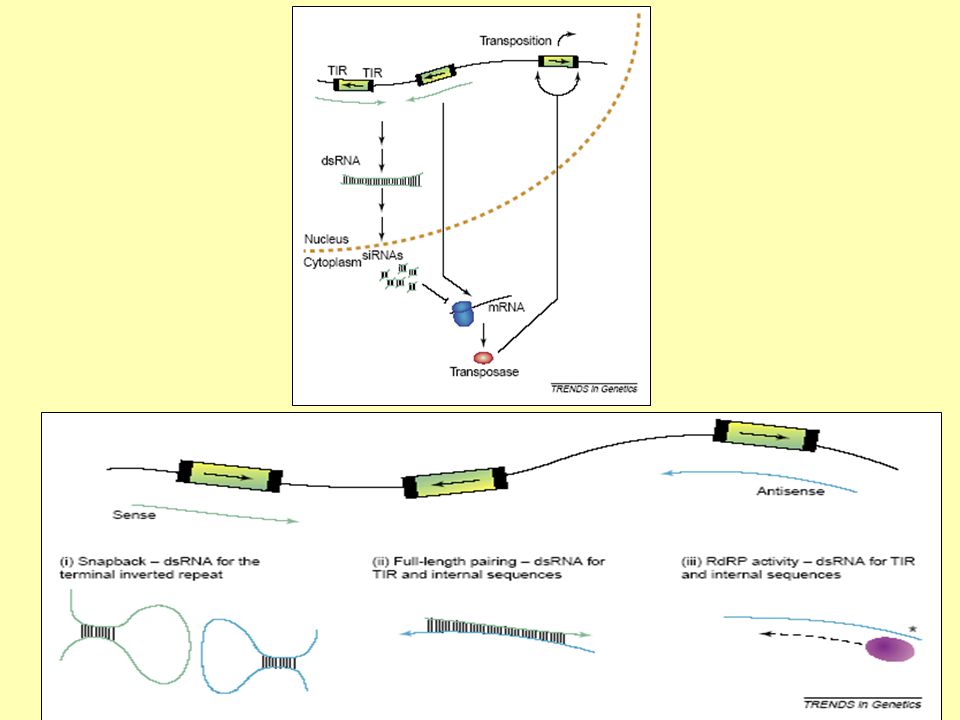
• Umlčování transpozonů pomocí metylace cytosinu či RNA interference. • Odstraňování transpozonů pomocí ektopické rekombinace. • Preferenční včleňování transpozonů do míst, kde neškodí (heterochromatin, jiné transpozony). Ale: Některé elementy Např. MITE elementy, Helitrony (eukaryotické DNA transpozony) se vyskytují převážně v blízkosti genů či uvnitř intronů. • Omezení transpozice na germinální linii.
. Ale: Některé elementy Např. MITE elementy, Helitrony (eukaryotické DNA transpozony) se vyskytují převážně v blízkosti genů či uvnitř intronů. • Omezení transpozice na germinální linii.")
56
UMLČOVÁNÍ TRANSPOSONŮ
Metylace DNA Metylace histonu H3 (lyzin 9) Deacetylace histonů RNAi
 Deacetylace histonů. RNAi.")
57
Umlčování transposonů je transkripční i posttranskripční

58
Transkripční umlčování – metylace promotorů TE.
Posttranskripční umlčování – sekvenčně specifická degradace RNA v cytoplazmě.

59
Umlčování transposonů mechanizmem RNAi
Caenorhabditis elegans a Drosophila nemají metylaci DNA. Umlčování transposonů je zajišťováno pomocí RNAi.

61
Tvorba heterochromatinu
• Transpozony často umlčeny a heterochromatinizovány pomocí DNA metylace a RNA interference. • Transpozony bývají důležitou součástí heterochromatinu centromer. • Inaktivace chromosomu X v somatických buňkách savců (důležité LINE).
.")
62
TANDEMOVÉ REPETICE

63
Evoluce tandemových repeticí
Evoluce v koncertu (concerted evolution) Genová konverze Molekulární tah (molecular drive) Nerovnoměrný crossing-over Evoluce satelitních sekvencí – skládání ze segmentů Genová konverze - interchromosomální - intrachromosomální
 Genová konverze. Molekulární tah (molecular drive) Nerovnoměrný crossing-over. Evoluce satelitních sekvencí – skládání ze segmentů. Genová konverze. - interchromosomální. - intrachromosomální.")
64
Satelity mohou vznikat i z retroelementů

65
PROMISKUITNÍ DNA

66
Genomové sekvence prozrazují, že spousta DNA z organel soustavně bombardovala jádro od doby, kdy organely vznikly. Nedávné experimenty ukázaly, že DNA přechází z organel do jádra v rozsahu, který byl dříve nepředstavitelný. Endosymbiotický transfer genů je všudypřítomný, kontinuální a přirozený proces, který prostupuje dynamiku jaderné DNA. Tento vytrvalý příliv organelové DNA zrušil autonomii organel a zvýšil komplexitu jádra.

67
Organelové genomy – pozůstatky prokaryot
(i) chloroplast kb proteinů progenitor - cyanobacteria (Synechocystis) 3.6 Mb 3000 proteinů (ii) mitochondrie 6-400 kb 3-67 proteinů progenitor - alpha-proteobacteria (Mesorhizobium loti) 7 Mb 6 700 proteinů
 chloroplast kb proteinů. progenitor - cyanobacteria (Synechocystis) 3.6 Mb proteinů. (ii) mitochondrie kb proteinů. progenitor - alpha-proteobacteria (Mesorhizobium loti) 7 Mb proteinů.")
68
? ? sinice proteobakterie mitochondrie chloroplast jádro
~ 4000 kb ~ 4000 genů ~ 3000 kb ~ 3000 genů mitochondrie chloroplast ? ~ 150 kb ~ 100 genů kb* ~ 60 genů ? jádro rostlinná buňka

69
Endosymbiotická evoluce a strom života

70
Endosymbiotický genový přenos: - transport genů, reimport proteinů

71
Mechanizmy genového přenosu
(i) Přenos velkých kusů DNA (“bulk DNA” hypothesis) intergenové spacery, introns experimenty u kvasinek >100kb (ii) Přenos prostřednictvím cDNA (“cDNA intermediates”) přenesená DNA je sestřižena a editována rekombinace sestřižené mtDNA s nesestřiženou mtDNA heterogenita velikostí mtDNA
 Přenos velkých kusů DNA ( bulk DNA hypothesis) intergenové spacery, introns. experimenty u kvasinek. >100kb. (ii) Přenos prostřednictvím cDNA ( cDNA intermediates ) přenesená DNA je sestřižena a editována. rekombinace sestřižené mtDNA s nesestřiženou mtDNA. heterogenita velikostí mtDNA.")
72
Proč některé geny zůstávají v organelách
(i) Hydrophobicita, hydrofóbní proteiny jsou těžko importovány do organel. (ii) Řízení redoxního stavu, organely řídí expresi genů, které kódují komponenty jejich elektronového transportu, jejich lokalizace je výhodnější v organelách.
 Hydrophobicita, hydrofóbní proteiny jsou těžko importovány do organel. (ii) Řízení redoxního stavu, organely řídí expresi genů, které kódují komponenty jejich elektronového transportu, jejich lokalizace je výhodnější v organelách.")
73
Zmenšení genomů u organel a parazitů
Proč některé geny zůstávají v organelách Zmenšení genomů u organel a parazitů Parazité specializace na intracelulární prostředí - ztráta genů Organely - export genů do jádra hostitele - import produktů

74
Některé geny se přenášejí do jádra dříve jiné později
Nejdříve – regulační funkce (sigma factor of RNApolymerase, gamma subunit of ATPase) Poslední – translace - respirace
 Poslední – translace. - respirace.")
75
Kam se přenesená DNA integruje
Nejsou důkazy preferovaných sekvencí nebo částí chromosomů. Sekvenční proměnlivost promiskuitní DNA >95% identity nasvědčuje velké obměně organelových sekvencí. Faktory vedoucí k degeneraci sekvencí Asexualita, poškozující zplodiny metabolismu, selekce na malé genomy. Kompenzující faktory (u rostlin) Polyploidie, reparace DNA.
 Polyploidie, reparace DNA.")
Podobné prezentace





>")
>")













