Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
SJEDNOCENÍ (integrace) METABOLISMU
Metabolismus je sestaven z s vysoce efektivně propojených drah. Každý tělesný orgán má jedinečný metabolický profil. Jídlo a hladovění indukují metabolické změny. Výběr energetického zdroje během cvičení je dán jeho intenzitou a trváním. Ethanol ovlivňuje energetický metabolismus v játrech.

2
ATP je univerzální energetické platidlo.
A) Svalová kontrakce, aktivní transport, zesílení signálů a biosyntézy. B) Hydrolýza ATP mění rovnovážný poměr mezi produkty a reaktanty ve spřažené reakci faktorem 108. Thermodynamicky nevýhodné reakce jsou tak posouvány ve prospěch produktů.
 Svalová kontrakce, aktivní transport, zesílení signálů a biosyntézy. B) Hydrolýza ATP mění rovnovážný poměr mezi produkty a reaktanty ve spřažené reakci faktorem 108. Thermodynamicky nevýhodné reakce jsou tak posouvány ve prospěch produktů.")
3
Společným meziproduktem většiny těchto reakcí je acetylCoA.
ATP je produkován při oxidaci molekul obsahujících energii jako jsou glukosa, mastné kyseliny a aminokyseliny. Společným meziproduktem většiny těchto reakcí je acetylCoA. Citrátový cyklus, oxidatívní fosforylace. Glukosa je také oxidována v glykolýze – pouze 2 ATP, kdežto v citrátovém cyklu 30 ATP.

4
NADPH je hlavním donorem elektronů při reduktivních biosyntézách.
Většinu potřebného NADPH poskytuje pentosafosfátová dráha. Biomolekuly jsou syntetizovány z malých atomových jednotek. Katabolické reakce poskytují stavební jednotky pro biosyntézy. Např. C2 – acetylCoA je prekurzorem biosyntézy mastných kyselin, prostaglandinů, ketolátek a cholesterolu.

5
Nejčastější motivy metabolické regulace.
Allosterické interakce – aktivity enzymů katalyzujících klíčové reakce, spíše než dostupnost substrátů je regulačním prvkem metabolické dráhy. Jako příklad. fosfofruktokinasa při glykolýze a acetylCoAkarboxylasa při syntéze mastných kyselin.

6
Kovalentní modifikace
Kovalentní modifikace. Řada enzymů je vedle allosterické regulace ještě regulována fosforylací nebo adenylací, proč ?? Příklad – glykogenfosforylasa je fosforylcí aktivována, kdežto glykogensynthasa deaktivována.

7
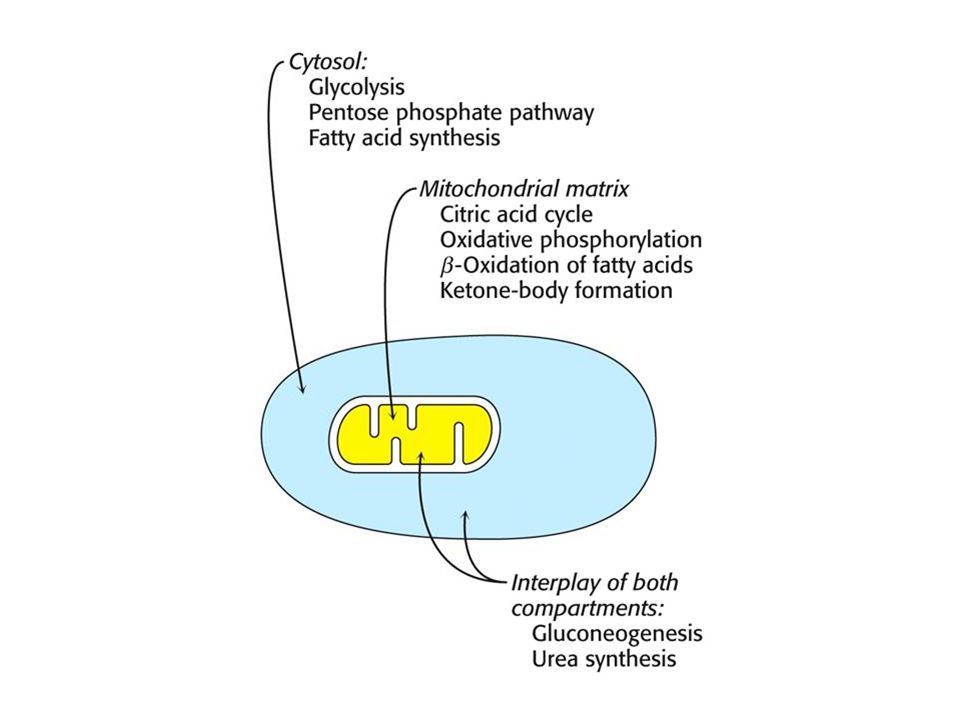
Hladina enzymů. Rychlost syntézy a odbourávání mnoha regulačních enzymů je ovlivňována hormony. Kompartmentace – metabolický model eukaryotních buněk je významně ovlivňován přítomností kompartmentů. Osud mnoha molekul závisí na jejich přítomnosti v mitochondrii nebo v cytoplasmě. Příklad – mastné kyseliny jsou transportovány do matrix v době potřeby energii, kdežto mastné kyseliny v cytoplasmě jsou esterifikovány nebo transportovány.

9
Hlavní metabolické dráhy a kontrolní místa. GLYKOLÝZA

10
Rychlost glykolýzy závisí na poměru ATP/AMP (potřebě energie) a stavebních jednotek což signalizuje hladina citrátu. V játrech je nejdůležitějším regulátorem aktivity fosfofruktokinasy fruktosa-2,6-bisfosfát. Hladina fruktosa-2,6-bisfosfátu je dána aktivitou kinasy, která ji tvoří z fruktosa-6-fosfátu a fosfatasy, která hydrolyzuje fosfát z 2-OH skupiny. V době, kdy je hladina glukosy v krvi nízká, glukagonová kaskáda aktivuje fosfatasu a inhibuje kinasu. Glykolýza je zpomalována a glukosa je použita prostřednictvím krve v jiných orgánech.

11
Citrátový cyklus a oxidativní fosforylace
Respirační kontrola – donory elektronů (NADH a FADH2) jsou reoxidovány a vraceny zpět do cyklu jen za situace, kdy je ADP simultánně fosforylován na ATP. Dostatek ATP snižuje aktivitu dvou enzymů cyklu – isocitrátdehydrogenasy a a-oxoglutarátdehydrogenasy.
 jsou reoxidovány a vraceny zpět do cyklu jen za situace, kdy je ADP simultánně fosforylován na ATP. Dostatek ATP snižuje aktivitu dvou enzymů cyklu – isocitrátdehydrogenasy a. a-oxoglutarátdehydrogenasy.")
12
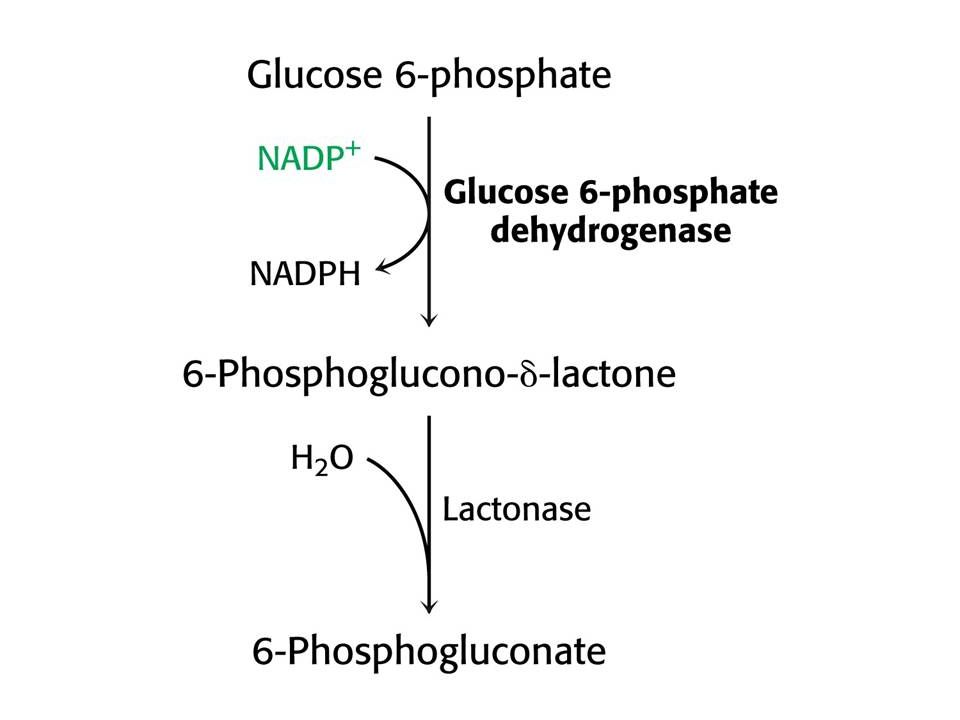
Pentosafosfátová dráha.
Probíhá v cytoplasmě. Oxidační a neoxidační fáze. Klíčový enzym – glukosa-6-fosfátdehdrogenasa. Aktivita závisí na hladině NADP+. Neoxidační fáze – převádí tři ribosy na dvě hexosy a jednu triosu – vstup pentos do glykolýzy.

14
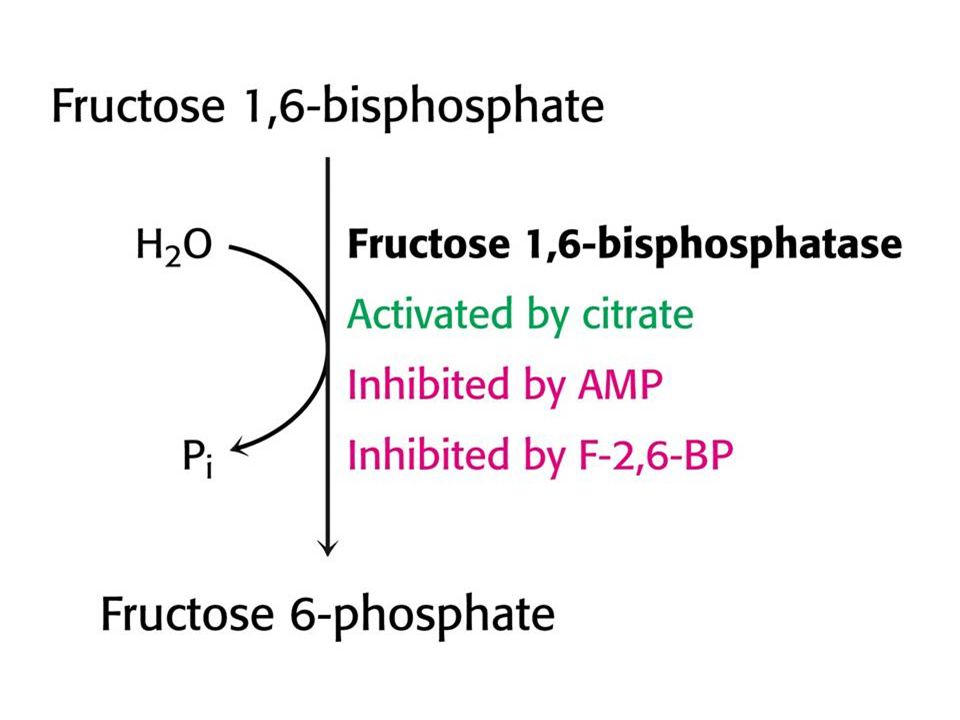
Glukoneogeneze. Syntéza sacharidů v játrech a ledvinách z necukerných prekurzorů – laktát, glycerol a aminokyseliny. Hlavním vstupem je pyruvát karboxylován na oxaloacetát v matrix. Oxaloacetát je v cytoplasmě metabolizován na fosfoenolpyruvát. Důležité jsou dva hydrolytické stupně – obchází ireversibilní stupně glykolýzy. . Glukoneogeneze a glykolýza jsou recipročně regulované dráhy. Jedna nebo druhá !! Např. AMP inhibuje a citrát aktivuje fruktosa-1,6-bisfosfatasu (glukoneogeneze). Obě molekuly mají opačný vliv na fosfofruktokinasu (klíčový enzym glykolýzy). Fruktosa-2,6-bisfosfát inhibuje fruktosa-1,6-bisfosfatasu. Dostatek glukosy – vysoká hladina F-2,6-BP inhibuje glukoneogenezi a aktivuje glykolýzu.
. Obě molekuly mají opačný vliv na fosfofruktokinasu (klíčový enzym glykolýzy). Fruktosa-2,6-bisfosfát inhibuje fruktosa-1,6-bisfosfatasu. Dostatek glukosy – vysoká hladina F-2,6-BP inhibuje glukoneogenezi a aktivuje glykolýzu.")
16
Syntéza a degradace glykogenu
Glykogenfosforylasa – tvorba glukosa-1 –P a izomerace na glukosa-6-fosfát. Syntéza glykogenu – aktivovaná glukosa – UDP-glukosa. UDP-glukosa se tvoří z glukosa-1-fosfátu a UTP. Degradace glykogenu a syntéza jsou koordinováně kontrolovány hromonálně řízenou kaskádou tak, že fosforylasa je aktivní kdy synthasa je inaktivní a naopak.

17
Granule glykogenu v jaterní buňce

18
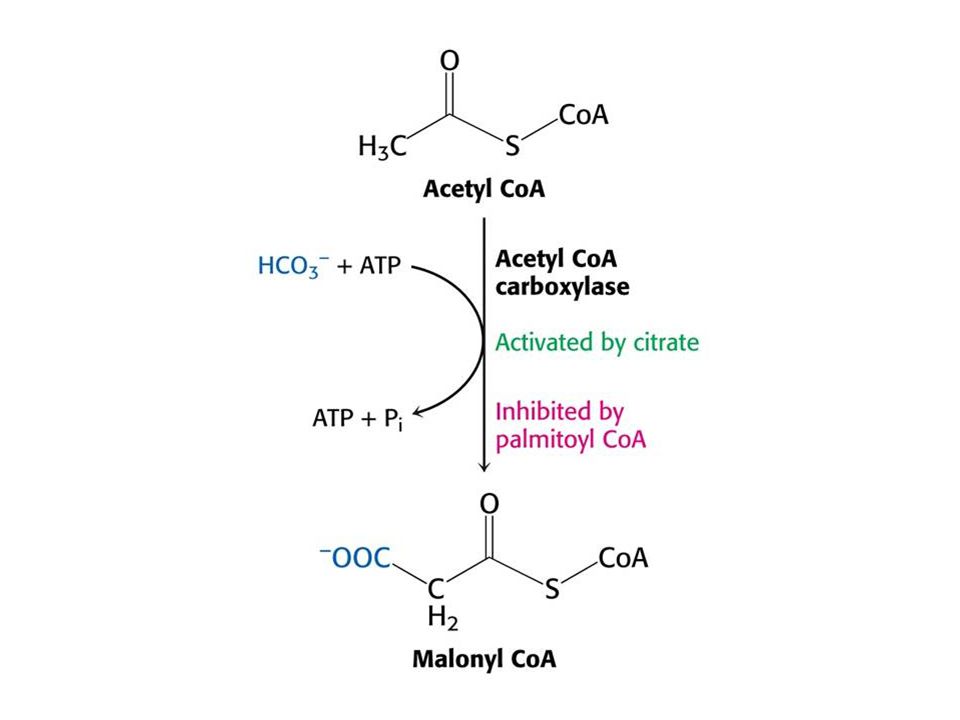
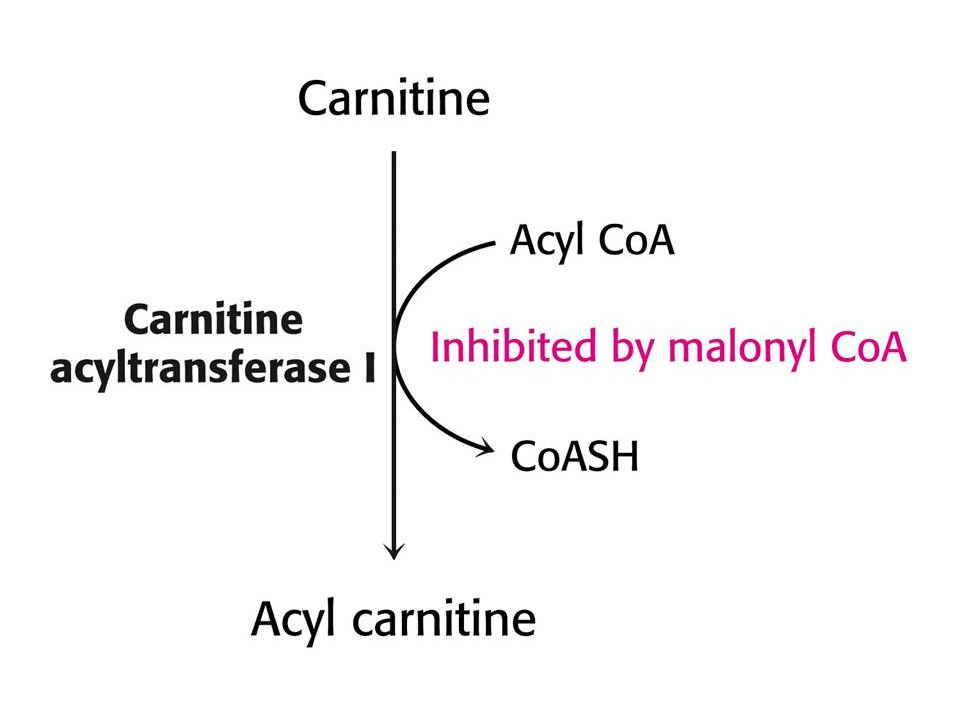
Syntéza a odbourávání mastných kyselin.
Syntéza v cytoplasmě, degradace v matrix. Citrát v cytoplasmě aktivuje acetylCoAkarboxylasu. Při vysoké hladině ATP a acetylCoA, roste hladina citrátu, která urychluje syntézu mastných kyselin. Při odbourávání mastných kyselin hraje klíčovou roli hladina NAD+ a FAD (podobně jako u CC). MalonylCoA inhibuje tvorbu acylkarnitinu karnitinacyltransferasou 1 a tím odbourávání mastných kyselin.
. MalonylCoA inhibuje tvorbu acylkarnitinu karnitinacyltransferasou 1 a tím odbourávání mastných kyselin.")
21
Klíčové metabolity: Glukosa-6-fosfát, Pyruvát a AcetylCoA.
Glukosa-6-P:

22
Pyruvát a acetylCoA.

23
Každý tělesný orgán má svůj jedinečný metabolický profil.
MOZEK. Jediným zdrojem energie pro mozek je glukosa. Výjimku tvoří delší doba hladovění. Mozek denně spotřebuje 120g glukosy (60% celkové spotřeby těla). Větší díl této energie se spotřebuje na transport na membránách vedoucí k vytvoření Na+ K+ membránového potenciálu pro přenos nervových vzruchů. Glukosa je do mozku transportována glukosovým transportérem GLUT3. Koncentrace glukosy v mozku je 1 mM. Při delším hladovění nahrazují glukosu ketolátky !!!
. Větší díl této energie se spotřebuje na transport na membránách vedoucí k vytvoření Na+ K+ membránového potenciálu pro přenos nervových vzruchů. Glukosa je do mozku transportována glukosovým transportérem GLUT3. Koncentrace glukosy v mozku je 1 mM. Při delším hladovění nahrazují glukosu ketolátky !!!")
25
Sval. Hlavním zdrojem energie pro sval jsou glukosa, mastné kyseliny a ketolátky.
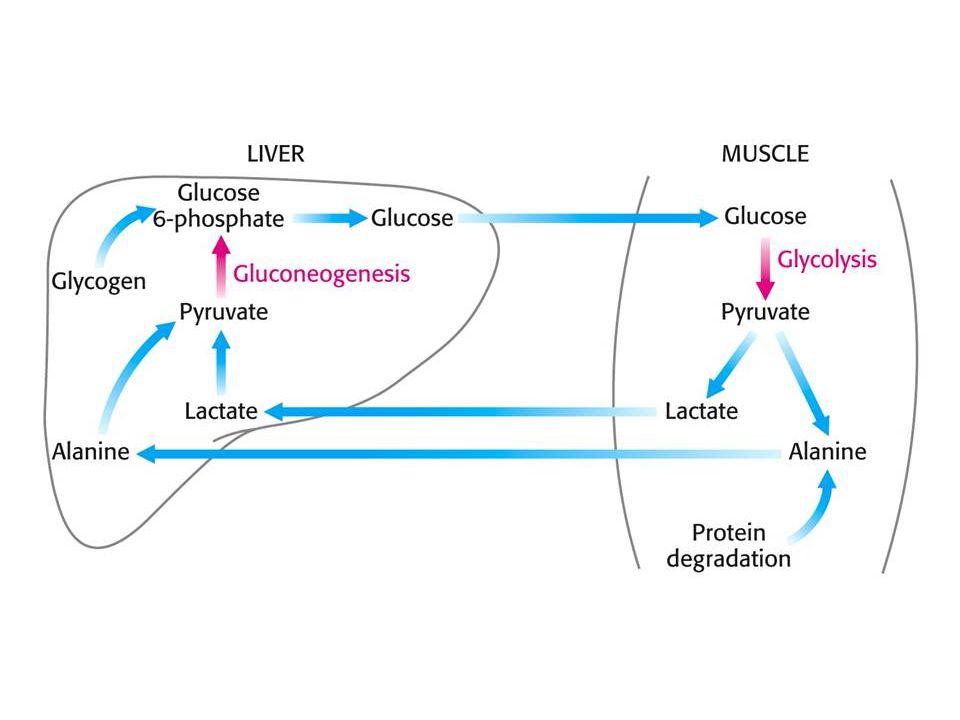
Svaly mají velkou zásobu glykogenu – kJ. Sval, stejně jako mozek, nemá glukosa-6-fosfatasu a neexportuje glukosu. Při intenzívní svalové práci je rychlost glykolýzy větší než CC a vzniklý pyruvát je převáděn na laktát. Tento přesun – Coriho cyklus- přenáší část metabolismu svalu do jater. Část pyruvátu je transaminována na Ala !!! Sval nemůže převést amoniak na močovinu ! Ve svalu se transaminují větvené aminokyseliny Pro odpočívající sval jsou hlavním zdrojem energie mastné kyseliny (85 % potřebné energie). Srdeční sval je bohatý na mitochondrie a nemá zásoby glykogenu. POUZE AEROBNÍ !! Hlavním zdrojem energie srdečního svalu jsou mastné kyseliny (také acetoacetát + laktát). Srdeční sval preferuje acetoacetát před glukosou.
. Srdeční sval je bohatý na mitochondrie a nemá zásoby glykogenu. POUZE AEROBNÍ !! Hlavním zdrojem energie srdečního svalu jsou mastné kyseliny (také acetoacetát + laktát). Srdeční sval preferuje acetoacetát před glukosou.")
27
Adiposní tkáně. Skladovací místo triacyglycerolů. Typický 70 kg muž má 15 kg triacylglycerolů ( kJ energie). Mastné kyseliny se syntetizují u člověka v játrech. Poté jsou esterifikovány s glycerolem a transportovány jako triacylgylceroly do adiposních tkání VLDL. Na povrchu adiposních buněk jsou hydrolyzovány lipoproteinlipasami. Mastné kyseliny vstupují do adiposních buněk. Tam jsou znova esterifikovány. Glycerol se tvoří glykolýzou z glukosy. K ukládání triacylglycerolů je tedy nutná glukosa !!! Při nedostatku glukosy jsou mastné kyseliny transportovány do plasmy. Hlavním ukazatelem pro uvolnění mastných kyselin do plasmy je hladina glycerolu uvnitř adiposních buněk.

28
Syntéza a degradace triacylglycerolů v adiposních buňkách
Syntéza a degradace triacylglycerolů v adiposních buňkách. Mastné kyseliny jsou do adiposních buněk transportovány ve formě triacylglycerolů obsažených ve VLDL.

29
Ledviny. Hlavním úkolem ledvin je vylučování moči, která slouží jako nosič odpadních metabolických produktů a udržování osmolarity tělesných tekutin. Denně se vylučuje 1 až 2 L moči. Krevní plasma je 60x denně filtrována. Reabsorbce spotřebuje velké množství energie - na váhu mají ledviny asi 0, 5% tělesné hmotnosti, spotřebují 10 % kyslíku z buněčné respirace. Glukosa je reabsorbována Na+-glukosovým přenašečem. Během hladovění jsou ledviny místem glukoneogeneze (polovina krevní glukosy).
.")
30
JÁTRA Metabolická aktivita jater je důležitá pro zásobování energií mozku, svalů, a ostatních periferních orgánů. Játra odebírají 2/3 glukosy z krve a všechny zbývající monosacharidy. Glukosa-6-fosfát je ukládána jako glykogen (v játrech kJ), ostatní je metabolizována na acetylCoA – tvorba mastných kyselin, cholesterolu a žlučových kyselin. Glukoneogeneze a uvolňování glukosy z glykogenu zásobuje krev a udržuje hladinu glukosy. Játra také hrají centrální roli při regulaci metabolismu lipidů. Při dostatku energie jsou mastné kyseliny z potravy a syntetizované v játrech uvolňovány do krve formou VLDL(very low density lipoproteins). Při hladovění převádí játra mastné kyseliny na ketolátky. Klíčovým krokem je možnost vstupu mastných kyselin do matrix. Reguluje hladina malonylCoA – pokud je ho dostatek, neprobíhá b-oxidace a tvorba ketolátek. Mastné kyseliny jsou transportovány do adiposních buněk.
, ostatní je metabolizována na acetylCoA – tvorba mastných kyselin, cholesterolu a žlučových kyselin. Glukoneogeneze a uvolňování glukosy z glykogenu zásobuje krev a udržuje hladinu glukosy. Játra také hrají centrální roli při regulaci metabolismu lipidů. Při dostatku energie jsou mastné kyseliny z potravy a syntetizované v játrech uvolňovány do krve formou VLDL(very low density lipoproteins). Při hladovění převádí játra mastné kyseliny na ketolátky. Klíčovým krokem je možnost vstupu mastných kyselin do matrix. Reguluje hladina malonylCoA – pokud je ho dostatek, neprobíhá b-oxidace a tvorba ketolátek. Mastné kyseliny jsou transportovány do adiposních buněk.")
31
Játra shromažďují aminokyseliny – priorita biosyntéza proteinů.
Km aminokyselin pro aminoacyltRNAsynthetasu je nižší než pro enzymy katabolismu aminokyselin. Při katabolismu se první odděluje dusík. Játra uvolňují 20 až 30g močoviny denně. Játra nedokáží odstraňovat dusík z větvených aminokyselin – transaminace probíhá ve svalech. Zdrojem vlastní energie jater jsou a-oxokyseliny. Účelem glykolýzy v játrech je tvorba stavebních jednotek pro biosyntézu. Játra také nedokáží využít acetoacetát jako zdroj energie – nemají transferasu nutnou k aktivaci acetoacetátu na acetylCoA.

34
Jídlo a hladovění vyvolávají metabolické změny.
Budeme sledovat cyklus nasycení a noční hladovění: Nejdůležitějším úkolem metabolismu je udržet glukosovou homeostázu – konstantní hladinu glukosy v krvi. a) Nasycenost. Glukosa a aminokyseliny jsou transportovány ze střeva do krve. Tuky z potravy jsou transportovány do krve cestou lymfatického systému ve formě chylomikronů. Zvýšená hladina glukosy vede k uvolnění insulinu, který je signálem nasycení. Stimuluje skladování energetických látek a syntézu proteinů. Insulin iniciuje proteinkinasovou kaskádu – stimulace syntézu glykogenu ve svalech i játrech a potlačuje glukoneogenezi v játrech. Urychluje také glykolýzu v játrech, která vede ke zvýšení syntézy mastných kyselin.
 Nasycenost. Glukosa a aminokyseliny jsou transportovány ze střeva do krve. Tuky z potravy jsou transportovány do krve cestou lymfatického systému ve formě chylomikronů. Zvýšená hladina glukosy vede k uvolnění insulinu, který je signálem nasycení. Stimuluje skladování energetických látek a syntézu proteinů. Insulin iniciuje proteinkinasovou kaskádu – stimulace syntézu glykogenu ve svalech i játrech a potlačuje glukoneogenezi v játrech. Urychluje také glykolýzu v játrech, která vede ke zvýšení syntézy mastných kyselin.")
35
Jak je odstraněna nadbytečná glukosa po nasycení ?
Insulin urychluje vstup glukosy do jater – GLUT2. V játrech se zvyšuje tvorba glukosa-6-fosfátu rychleji než v krvi, pouze zde je glukokinasa (porovnej KM s hexokinasou) nasycena glukosou. Tvoří se glykogen. Vysoká hladina glukosy vede k její vazbě na fosforylasu a. Vzniklý adukt je substrátem fosfatasy, která ji přemění na fosforylasu b , která neodbourává glykogen. Vysoká hladina insulinu po nasycení podporuje vstup glukosy do svalů a adiposní tkáně. Insulin podporuje absorbci větvených aminokyselin ve svalu. Dále podporuje proteosyntézu a inhibuje odbourávání proteinů.
 nasycena glukosou. Tvoří se glykogen. Vysoká hladina glukosy vede k její vazbě na fosforylasu a. Vzniklý adukt je substrátem fosfatasy, která ji přemění na fosforylasu b , která neodbourává glykogen. Vysoká hladina insulinu po nasycení podporuje vstup glukosy do svalů a adiposní tkáně. Insulin podporuje absorbci větvených aminokyselin ve svalu. Dále podporuje proteosyntézu a inhibuje odbourávání proteinů.")
36
Počáteční stav hladovění.
Několik hodin po jídle hladina glukosy klesne. Dochází k sekreci glukagonu. Glukagon signalizuje hladovění. Hlavním orgánem působnosti glukagonu jsou játra. Stimuluje štěpení glykogenu, inhibuje syntézu, spuštěním hormonální kaskády vedoucí k fosforylaci a aktivaci fosforylasy a inhibici syntézy glykogenu. Inhibuje také syntézu mastných kyselin snížením produkce pyruvátu a snížením aktivity actylCoAkarboxylasy (je ponechána v nefosforylovaném stavu). Glukagon stimuluje glukoneogenezi v játrech a blokuje glykolýzu snížením hladiny F-2,6-bisP. Všechny reakce glukagonu jsou zprostředkovány proteinkinasou aktivovanou cAMP. Uvolněná glukosa se dostává krví do periferních tkání. Snížená spotřeba glukosy svaly a adiposní tkání vede také ke zvýšení hladiny glukosy v krvi.
. Glukagon stimuluje glukoneogenezi v játrech a blokuje glykolýzu snížením hladiny F-2,6-bisP. Všechny reakce glukagonu jsou zprostředkovány proteinkinasou aktivovanou cAMP. Uvolněná glukosa se dostává krví do periferních tkání. Snížená spotřeba glukosy svaly a adiposní tkání vede také ke zvýšení hladiny glukosy v krvi.")
37
Když klesne hladina glukosy, využívají svaly i játra mastné kyseliny jako zdroj energie.
Závěr: Hladina glukosy v krvi na 80mg/dl se udržuje třemi faktory: a) Mobilizací glykogenu a uvolňováním glukosy do krve. b) Uvolňováním mastných kyselin z adiposních tkání. c) U jater a svalů - posunem využívané energie od glukosy k mastným kyselinám
 Mobilizací glykogenu a uvolňováním glukosy do krve. b) Uvolňováním mastných kyselin z adiposních tkání. c) U jater a svalů - posunem využívané energie od glukosy k mastným kyselinám.")
38
Jaký je efekt silné snídaně ?
Tuk je utilizován stejně jako za stavu nasycení. Jinak je to s glukosou. Játra z počátku neabsorbují glukosu z krve – ponechávají ji pro periferní tkáně. Navíc jsou ve stavu glukoneogeneze. Nově syntetizovaná glukosa v první řadě nahrazuje vyčerpaný glykogen. Pokud hladina glukosy v krvi dále roste, játra doplnila glykogen, začínají nadbytečnou glukosu využívat k syntéze mastných kyselin (glycerol).
.")
39
Metabolická adaptace při prodlouženém hladovění minimalizuje degradaci proteinů.
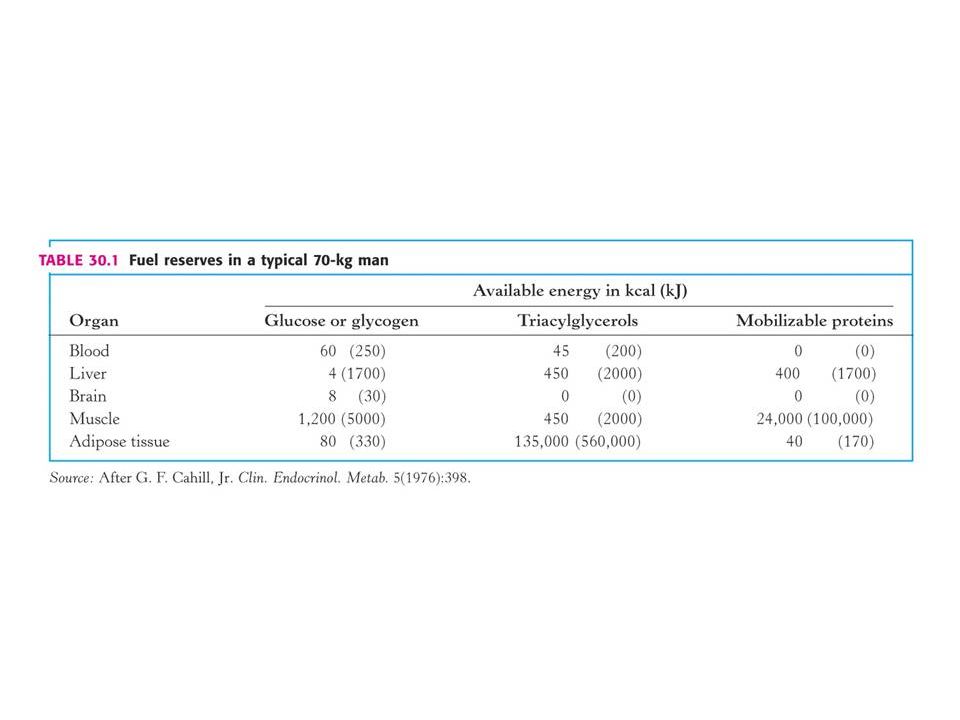
Jaká je adaptace metabolismu na půst až k hladovění ? Typický 70 kg muž má celkové energetické rezervy kJ. Energetická potřeba na 24 hod činí kolem kJ (závisí na tělesné aktivitě). Zásoba energie je na 1 až 3 měsíce. POZOR! Zásoby sacharidů se vyčerpanjí za jeden den ! I při hladovění musí být minimální hladina glukosy v krvi 2, 2 mM ( 40 mg/dl). Prvořadou funkcí metabolismu při hladovění je zásobit glukosou mozek a červené krvinky. Jediným zdrojem energie jsou triacylglyceroly Jediným potenciálním zdrojem glukosy aminokyseliny (odhlédneme-li od minortního množství glycerolu z triacylglyceorolu).
. Zásoba energie je na 1 až 3 měsíce. POZOR! Zásoby sacharidů se vyčerpanjí za jeden den ! I při hladovění musí být minimální hladina glukosy v krvi 2, 2 mM ( 40 mg/dl). Prvořadou funkcí metabolismu při hladovění je zásobit glukosou mozek a červené krvinky. Jediným zdrojem energie jsou triacylglyceroly Jediným potenciálním zdrojem glukosy aminokyseliny (odhlédneme-li od minortního množství glycerolu z triacylglyceorolu).")
40
Výběr zdroje energie během hladovění
Výběr zdroje energie během hladovění. Hladiny ketolátek a mastných kyselin se zvyšují, hladina glukosy klesá.

41
Proteiny nejsou skladovány !
Odbouráváním ztrácí funkci. Druhou prioritou metabolismu při hladovění je ochrana proteinů, což se uskutečňuje posunem odběru energie od glukosy k mastným kyselinám a ketolátkám ! Metabolické změny během prvního dne hladovění jsou shodné s nočním hladověním. Dominantními metabolickými pochody je mobilizace triacylglycerolů v adiposních tkáních glukoneogeneze v játrech. Játra získávají energii pro svoji vlastní potřebu oxidací mastných kyselin z adiposních tkání. Roste koncentrace acetylCoA a citrátu, což vypíná glykolýzu. Spotřeba glukosy svaly klesá (nízká hladina insulinu), mastné kyseliny vstupují volně.
, mastné kyseliny vstupují volně.")
42
Svaly přesunují zdroj energie od glukosy k mastným kyselinám.
b-oxidace mastných kyselin ve svalech zastavuje převod pyruvátu na acetylCoA, protože acetylCoA stimuluje fosforylaci pyruvátdehydrogenasového komplexu, který zůstává inaktivní. Pyruvát, laktát a alanin se transportují do jater – resyntéza glukosy (+ glycerol z triacylglycerolů). Pro glukoneogenezi se využívají i aminokyseliny. Nejdříve se odbourávají proteiny, které mají rychlý obrat, jako střevní epithel a sekrety pankreatu. Svalové proteiny se odbourávají až nakonec (dlouhé hladovění) – odbouráváním ztrácí funkci (srdeční sval).
. Pro glukoneogenezi se využívají i aminokyseliny. Nejdříve se odbourávají proteiny, které mají rychlý obrat, jako střevní epithel a sekrety pankreatu. Svalové proteiny se odbourávají až nakonec (dlouhé hladovění) – odbouráváním ztrácí funkci (srdeční sval).")
43
Jak se zabrání ztrátě svalstva ?
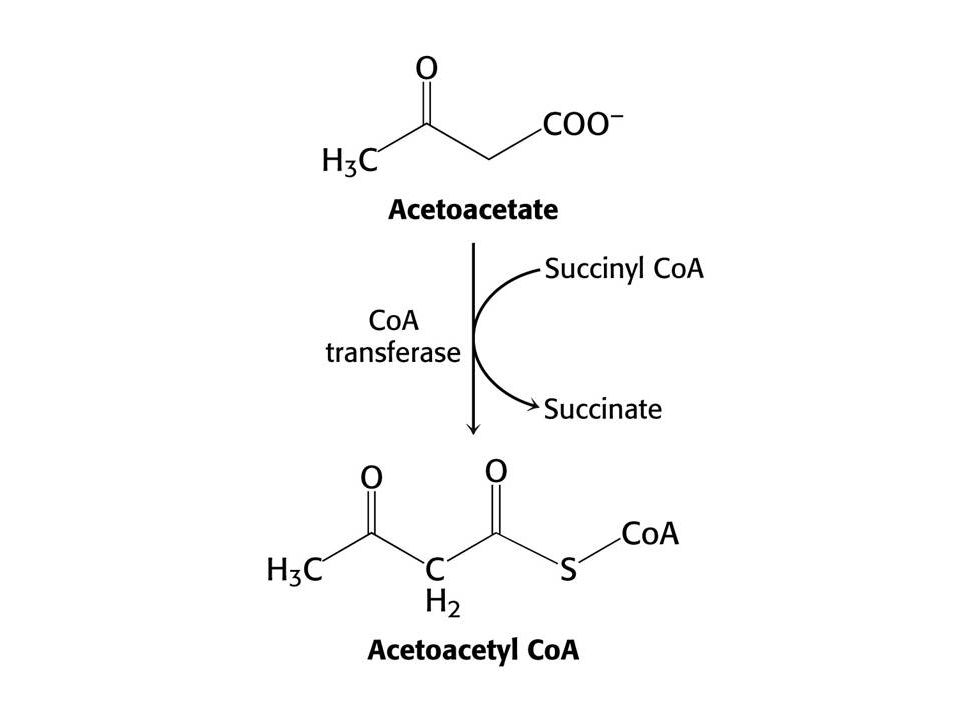
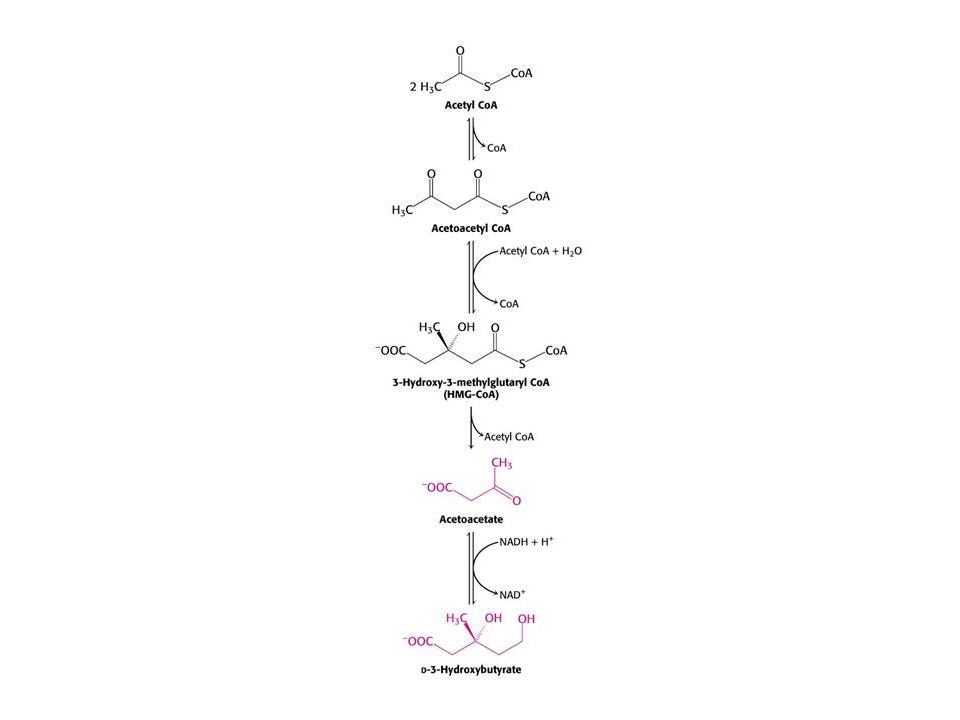
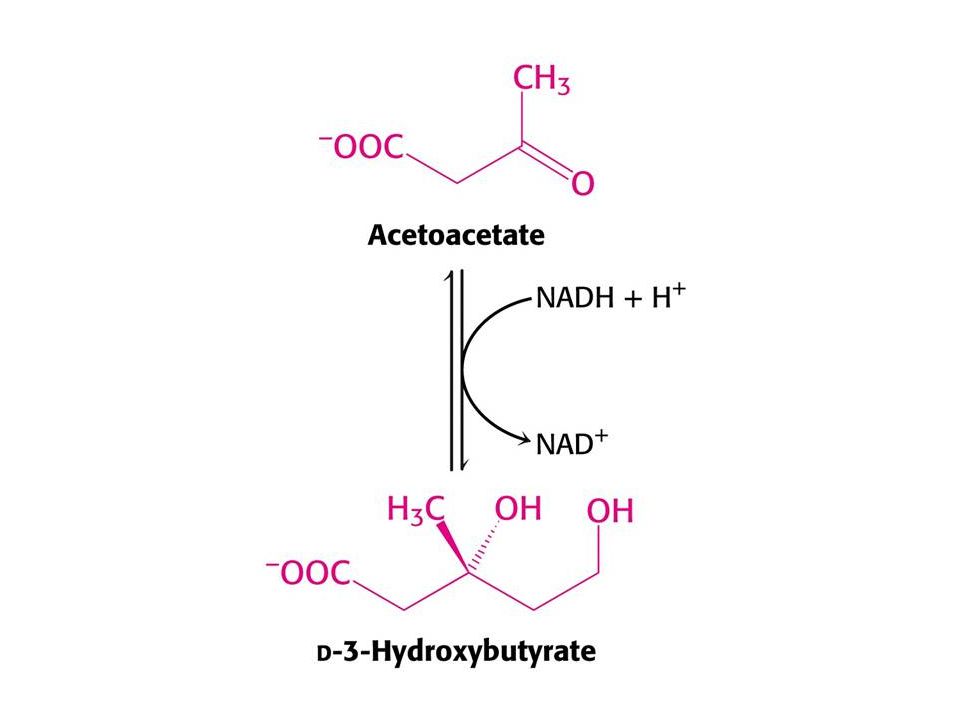
Po 3 dnech hladovění produkují játra velké množství acetoacetátu a D-3-hydroxybutyrátu. Syntéza z acetylCoA roste – CC cyklus nestačí zpracovat acetylCoA. Glukoneogeneze snižuje hladinu oxaloacetátu. V této chvíli se mozek orientuje na acetoacetát. Po třech dnech až 1/3 potřeby energie mozku je z acetoacetátu. Taktéž srdce využívá ketolátky. Po několika týdnech se stávají ketolátky hlavním zdrojem energie mozku. K využití acetoacetátu je nutný přenos CoA z sukcinylCoA.

44
Vstup ketolátek do CC.

45
Ketolátky jsou ekvivalentem mastných kyselin, které prochází bariérou do mozku.
Mozek potřebuje denně jen 40 g glukosy ve srovnáni se 120 g na začátku hladovění. Takto se velmi snižuje potřeba glukosy. Je odbouráváno méně svalstva než v prvním dnu hladovění. Odbourává se jen 20 g svalstva ve srovnání s prvními dny hladovění – 75 g. To je důležité pro přežití. Po vyčerpání zásob triacylglycerolů – jediným zdrojem energie jsou proteiny. Jejich odbourávání se zrychluje – smrt nastane ztrátou funkce srdce, jater a ledvin.

46
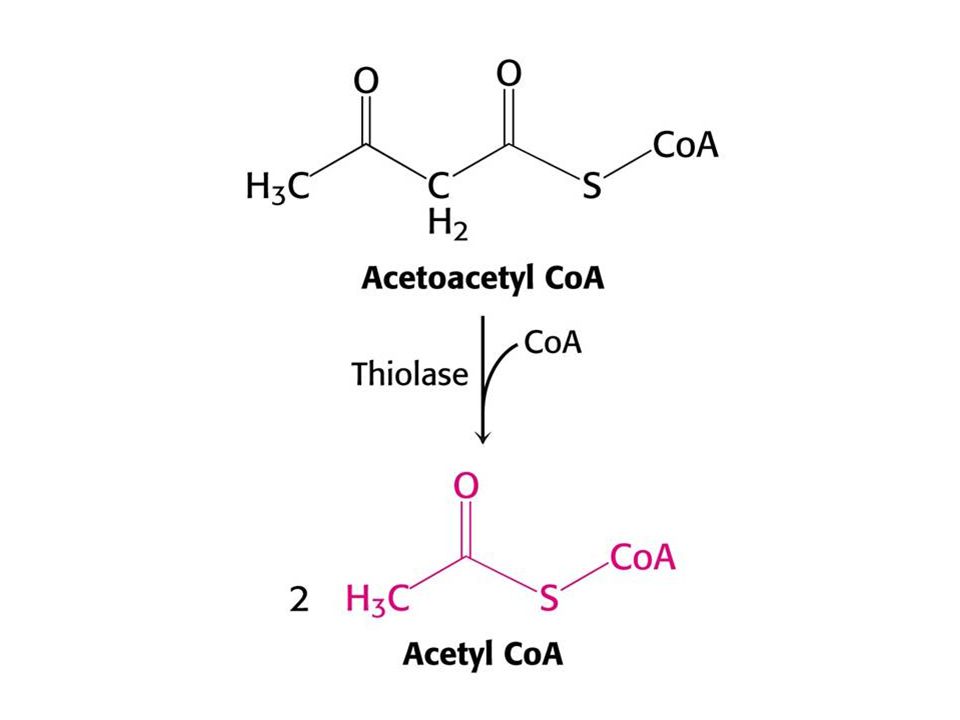
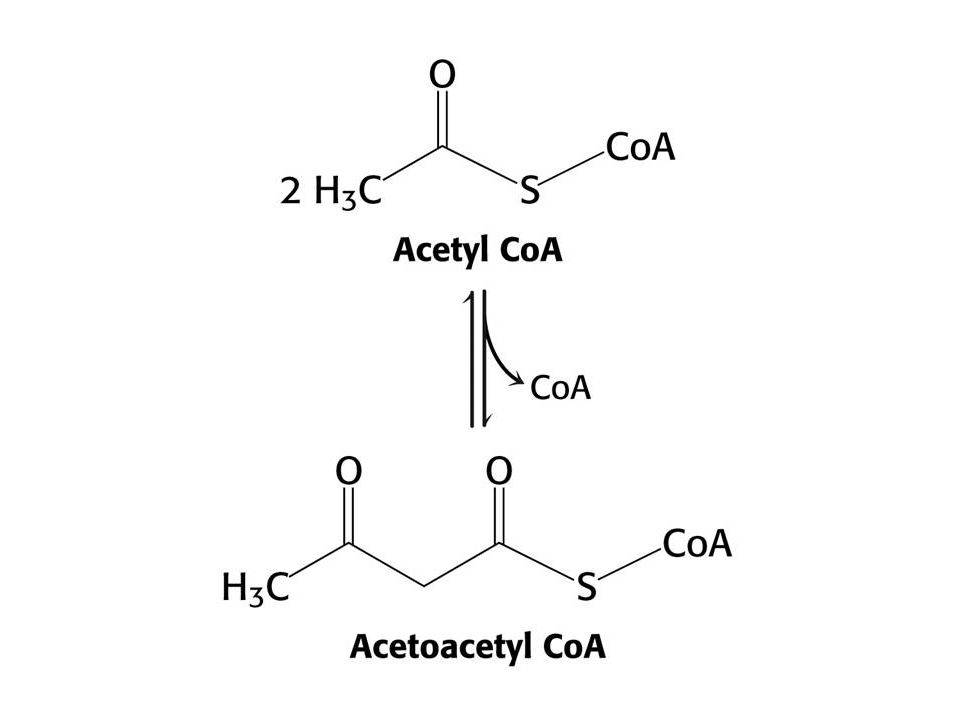
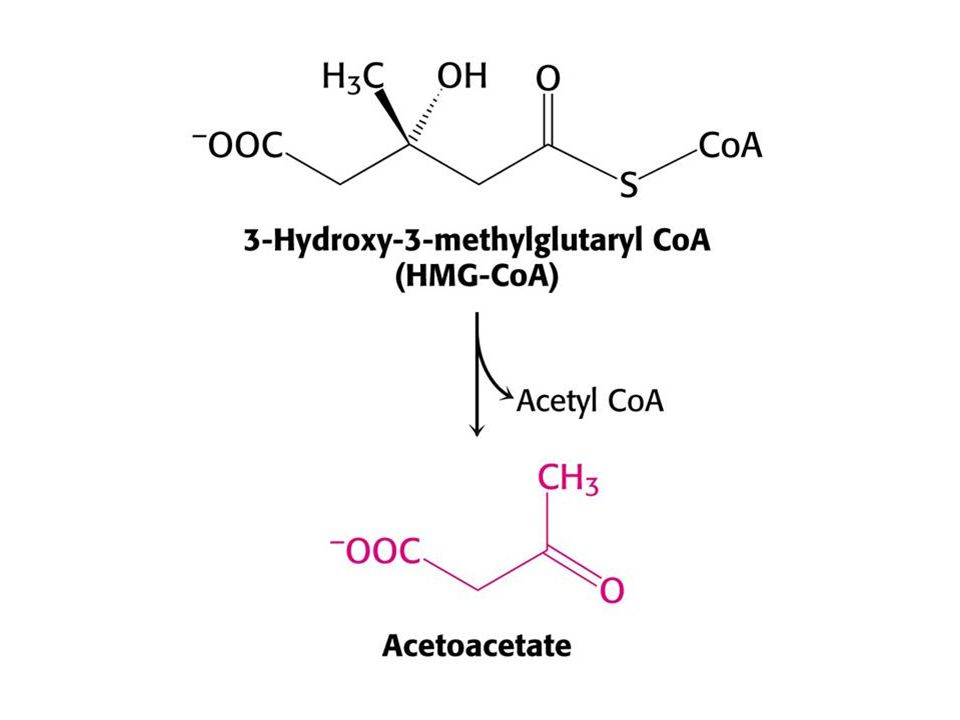
Syntéza ketolátek v játrech.

53
Metabolický zmatek při diabetu způsobený relativní insulinovou nedostatečností a nadbytkem glukagonu. Diabetes mellitus – cukrovka – nadprodukce glukosy z jater, nemožnost jejího využití v jiných orgánech. Výskyt v 5 % populace. Diabetes typu I – insulin dependent diabetes (IDDM), autoimunní destrukce b-buněk pankreatu. Obvykle se projeví před 20 rokem života. Diabetes typu II – non-insulin-dependent (NIDDM). Normální nebo dokonce vyšší hladiny insulinu v krvi, ale poškozené receptory na povrchu buněk.
, autoimunní destrukce b-buněk pankreatu. Obvykle se projeví před 20 rokem života. Diabetes typu II – non-insulin-dependent (NIDDM). Normální nebo dokonce vyšší hladiny insulinu v krvi, ale poškozené receptory na povrchu buněk.")
54
Typ I – bez insulinu, glukagon vyšší než normální hladina, diabetik hladoví přesto, že má vysokou hladinu cukru v krvi. Játra jsou ve stavu glukoneogeneze a ketogeneze. Glukagon způsobuje snížená hladiny F-2,6-BP v játrech. Glykolýza je inhibována. Vysoký poměr glukagon/insulin podporuje štěpení glykogenu. Glukosa se objevuje i v moči (mellitus). Vylučuje se tak i voda - diabetici pociťují hlad i žízeň. Nedostatek glukosy v buňkách vede k nadbytku acetylCoA (b-oxidace), který vzhledem k nedostatku oxaloacetátu nemůže do CC. Tvoří se ketolátky. Vysoké koncentrace ketolátek přesáhnou kapacitu ledvin a tím poruší acidobazickou rovnováhu. Při neléčeném diabetu dochází ke komatu snížením pH krve (acidosa) a dehydrataci. V dechu i moči diabetiků se objevuje také aceton – produkt dekarboxylace acetoacetátu.
, který vzhledem k nedostatku oxaloacetátu nemůže do CC. Tvoří se ketolátky. Vysoké koncentrace ketolátek přesáhnou kapacitu ledvin a tím poruší acidobazickou rovnováhu. Při neléčeném diabetu dochází ke komatu snížením pH krve (acidosa) a dehydrataci. V dechu i moči diabetiků se objevuje také aceton – produkt dekarboxylace acetoacetátu.")
55
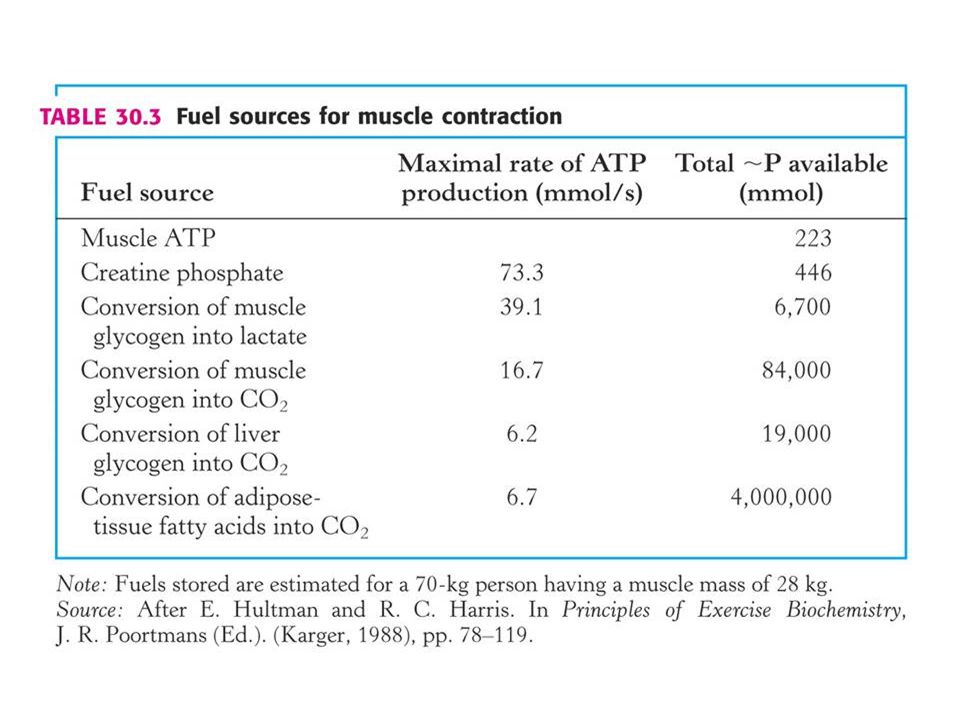
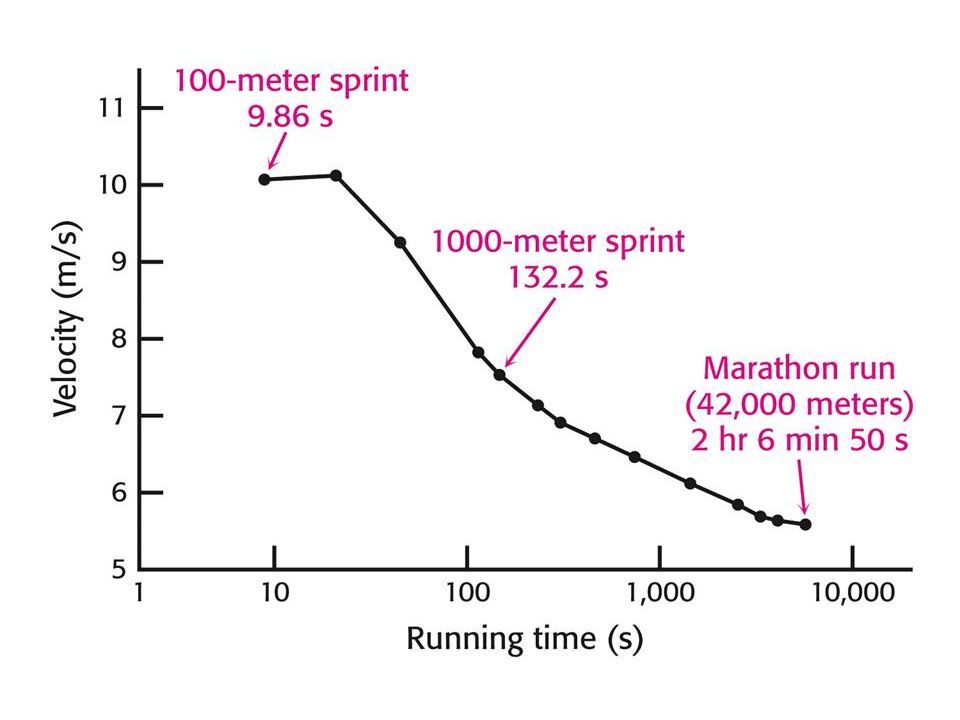
Výběr zdroje energie při fyzické zátěži (cvičení) je dán intenzitou a délkou zátěže.
Sprit a maratonský běh jsou zásobovány z různých zdrojů energie. Sprint (100 m) ze zásob ATP, kreatifosfátu a anaerobní glykolýzy. Během 10 s sprintu klesne hladina ATP ve svalech z 5, 2 na 3, 7 mM a kreatinfosfátu z 9, 1 na 2, 6 mM. Při anaerobní glykolýze vzroste hladina laktátu v krvi z 1, 6 na 8, 3 mM, což vede ke snížení pH v krvi ze 7, 42 na 7, 24 mM. Při běhu na m (132 s) je nutný další zdroj energie. Nastupuje aerobní proces – oxidativni fosforylace. To je pomalý proces a proto nemůže být u tohoto běhu stejná rychlost jako u sprintu.
 ze zásob ATP, kreatifosfátu a anaerobní glykolýzy. Během 10 s sprintu klesne hladina ATP ve svalech z 5, 2 na 3, 7 mM a kreatinfosfátu z 9, 1 na 2, 6 mM. Při anaerobní glykolýze vzroste hladina laktátu v krvi z 1, 6 na. 8, 3 mM, což vede ke snížení pH v krvi ze 7, 42 na 7, 24 mM. Při běhu na m (132 s) je nutný další zdroj energie. Nastupuje aerobní proces – oxidativni fosforylace. To je pomalý proces a proto nemůže být u tohoto běhu stejná rychlost jako u sprintu.")
56
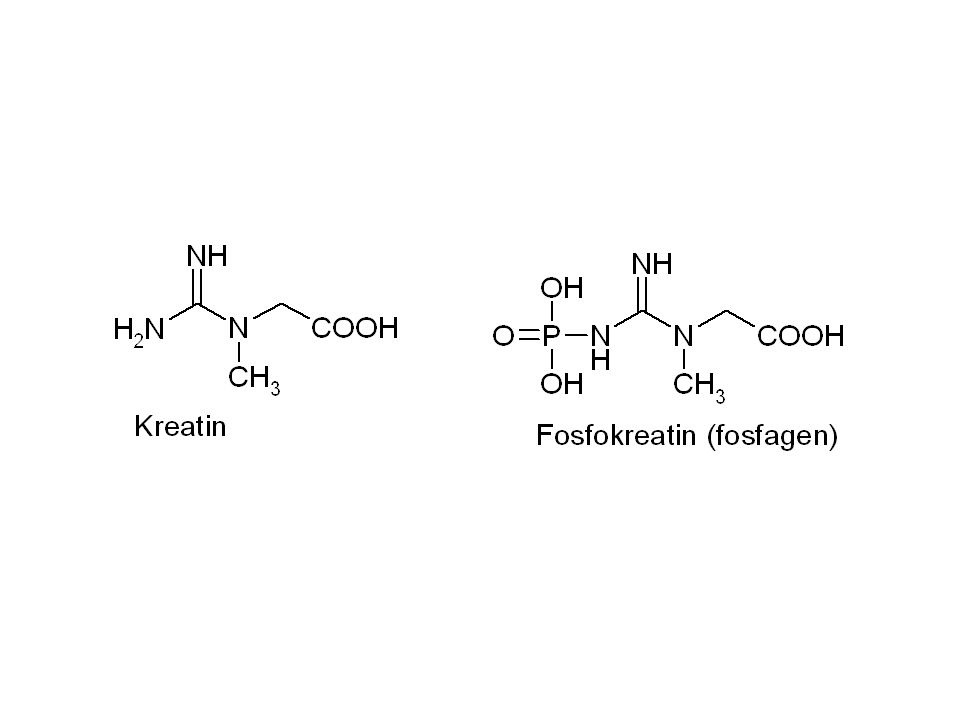
Úloha kreatinfosfátu ve svalech
Kreatinfosfát(KP) + ADP kreatinkinasa ATP + kreatin (K) Při pH 7 je stand. volná energie hydrolýzy KP – 43, 1 kJ/mol. Pro tvorbu ATP z ADP a KP je změna standardní volné energie -12, 6 kJ/mol. Což odpovídá rovnovážné konstantě = 162. Typické koncentrace v odpočívajícím svalu: [ATP] = 4 mM; [ADP]= 0, 013 mM; [KP] = 25 mM a [K] = 13 mM. KP je jediný zdroj regenerace ATP při prvních 4 sek. běhu na 100 m.
 + ADP kreatinkinasa ATP + kreatin (K) Při pH 7 je stand. volná energie hydrolýzy KP – 43, 1 kJ/mol. Pro tvorbu ATP z ADP a KP je změna standardní volné energie. -12, 6 kJ/mol. Což odpovídá rovnovážné konstantě = 162. Typické koncentrace v odpočívajícím svalu: [ATP] = 4 mM; [ADP]= 0, 013 mM; [KP] = 25 mM a [K] = 13 mM. KP je jediný zdroj regenerace ATP při prvních 4 sek. běhu na 100 m.")
58
Při maratonu (42, 2 km) – dochází k zisku energie spoluprací mezi svaly, játry a adiposní tkání. Využívá se jaterní a svalový glykogen, který však nestačí na 2 hodinový běh, kde je nutno vydat 150 molů ATP (veškerý glykogen poskytuje 103 molů ATP). Zapojují se adiposní tkáně – pomalý proces, pomalejší než degradace glykogenu a 10 x pomalejší než zisk energie z kreatinfosfátu. Pro dálkové běhy je důležitá tvorba ATP z mastných kyselin. Pokud by se získávala energie pouze takto, trval by maraton 6 hodin. Elitní běžci spotřebovávají asi stejné množství mastných kyselin a glykogenu (průměrná rychlost 5, 5 m/s).
.")
59
Jak se vytvoří optimální směs těchto dvou energetických zdrojů ?
Nízká hladina sacharidů v krvi a vysoký poměr glukagon / insulin mobilizují mastné kyseliny z adiposní tkáně. Mastné kyseliny vstupují do svalů, kde proběhne b-oxiace na acetylCoA a poté na CO2. Zvýšená hladina acetylCoA snižuje aktivitu pyruvátdehydrogenasového komplexu – brzdí se tak přeměna pyruvátu na acetylCoA. Proces tak snižuje vstup acetylCoA do CC. Glukosa se šetří. Simultánní odběr energie z obou zdrojů tak zabraňuje totální spotřebě glykogenu před počátkem odbourávání mastných kyselin.

62
ETHANOL mění energetický metabolismu v játrech.
Biochemické základy zdravotních potíží způsobených ethanolem. Ethanol je metabolizován játry dvěma drahami. PRVNÍ: V cytoplasmě, alkoholdehydrogenasou, na ethanal, a NADH + H+. V matrix, aldehyddehydrogenasou, na acetát, NADH + H+. Vysoká hladina NADH vede k hromadění laktátu (redukcí pyruvátu). Důsledkem je hypoglykemie a laktátová acidosa. Nadbytek NADH inhibuje také oxidaci mastných kyselin.
. Důsledkem je hypoglykemie a laktátová acidosa. Nadbytek NADH inhibuje také oxidaci mastných kyselin.")
63
Naopak, nadbytek NADH vede k syntéze mastných kyselin.
V játrech se akumulují tracylglyceridy – steatosa jater (ztučnění). DRUHÁ: Ethanolem indukovaný mikrosomální ethanol-oxidující systém (MEOS). Je to dráha obsahující cytochrom-P450. Dochází k tvorbě ethanalu a následně acetátu za současné oxidace NADPH na NADP+. Reakce zahrnuje kyslík a vede tak k tvorbě aktivních forem kyslíku, které poškozují tkáň. Vyčerpává se NADPH a antioxidant glutathion nemůže být regenerován.
. DRUHÁ: Ethanolem indukovaný mikrosomální ethanol-oxidující systém (MEOS). Je to dráha obsahující cytochrom-P450. Dochází. k tvorbě ethanalu a následně acetátu za současné oxidace NADPH na NADP+. Reakce zahrnuje kyslík a vede tak k tvorbě aktivních forem kyslíku, které poškozují tkáň. Vyčerpává se NADPH a antioxidant glutathion nemůže být regenerován.")
64
Efekty dalších metabolitů ethanolu:
Jaterní mitochondrie převádí acetát na acetylCoA za spotřeby ATP (až na AMP + 2 Pi) enzymem thiokinasou (aktivuje krátké mastné kyseliny). Další odbourání acetylCoA CC není možné (inhibice dvou enzymů NADH, kterých ?). Konsekvence: Tvoří se ketolátky, uvolňují se do krve, snižují pH. Tento proces je neúspěšný a proto se odbourávání zastaví na ethanalu, který vytváří kovalentní vazbu s mnoha důležitými funkčními skupinami proteinů. Vysoká konzumace ethanolu vede k značnému poškození jaterní tkáně ethanalem. Stupně následků konzumace ethanolu: 1. Steatoza jater, 2. Alkoholická hepatitida, 3. Cirhosa.
 enzymem thiokinasou (aktivuje krátké mastné kyseliny). Další odbourání acetylCoA CC není možné (inhibice dvou enzymů NADH, kterých ). Konsekvence: Tvoří se ketolátky, uvolňují se do krve, snižují pH. Tento proces je neúspěšný a proto se odbourávání zastaví na ethanalu, který vytváří kovalentní vazbu s mnoha důležitými funkčními skupinami proteinů. Vysoká konzumace ethanolu vede k značnému poškození jaterní tkáně ethanalem. Stupně následků konzumace ethanolu: 1. Steatoza jater, 2. Alkoholická hepatitida, 3. Cirhosa.")
65
Alcohol Metabolism (sung to the tune of Amazing Grace) Peter A
Alcohol Metabolism (sung to the tune of Amazing Grace) Peter A. Rubenstein When ethanol is oxidized An aldehyde‘s produced. Because a hydride‘s freed as well An NAD‘s reduced. The ADH contains a zinc Within its active site. It orients the alcohol And helps to hold it tight. When methanol is drunk instead Formaldehyde is made. When it‘s transformed into formate One‘s vision soon may fade. The ethanol‘s metabolized Within the liver cell Which helps detoxify the blood And thereby keeps us well. When at a party or a bar Keep this thought in your head.
 Peter A. Rubenstein. When ethanol is oxidized. An aldehyde‘s produced. Because a hydride‘s freed as well. An NAD‘s reduced. The ADH contains a zinc. Within its active site. It orients the alcohol. And helps to hold it tight. When methanol is drunk instead. Formaldehyde is made. When it‘s transformed into formate. One‘s vision soon may fade. The ethanol‘s metabolized. Within the liver cell. Which helps detoxify the blood. And thereby keeps us well. When at a party or a bar. Keep this thought in your head.")
66
Domácí úkol 1. Vyslovte hlavní důsledky deficitu každého z následujících enzymů: a) Hexokinasa v adiposní tkáni b) Glukosa-6-fosfatasa v játrech. c) Karnitinacyltransferasa I v kosterních svalech d) Glukokinasa v játrech. e) Thiolasa v mozku f) Kinasa v játrech syntetizující fruktosa-2,6-bisfosfát
 Glukosa-6-fosfatasa v játrech. c) Karnitinacyltransferasa I v kosterních svalech. d) Glukokinasa v játrech. e) Thiolasa v mozku. f) Kinasa v játrech syntetizující fruktosa-2,6-bisfosfát.")
67
2. Respirační kvocient (RQ)
2. Respirační kvocient (RQ). Klasický metabolický index je definován jako objem uvolněného CO2 / objem spotřebovaného O2 a) Vypočtěte hodnotu RQ kompletní oxidace glukosy a tripalmitoylglycerolu. b) Co vypovídá měření RQ o příspěvku těchto různých zdrojů energie během intenzívního cvičení ? (Odbourávání proteinů zanedbejte). 3. Porovnej výtěžek vody při kompletní oxidaci 1 g glukosy s výtěžkem vody z 1 g tripalmitoylglycerolu. Dejte výsledky do souvislosti s evolučním výběrem obsahu velbloudího hrbu. 4. Příjem velkého množství glukosy před startem maratonu by mohlo být způsobem jak zvýšit energetickou zásobu organismu. Zkušení běžci takto nečiní. Jaký je biochemický důvod vyhnout se tomuto způsobu zisku energie před startem ? (uvažuj vliv glukosy na hladinu insulinu).
. Klasický metabolický index je definován jako objem uvolněného CO2 / objem spotřebovaného O2. a) Vypočtěte hodnotu RQ kompletní oxidace glukosy a tripalmitoylglycerolu. b) Co vypovídá měření RQ o příspěvku těchto různých zdrojů energie během intenzívního cvičení (Odbourávání proteinů zanedbejte). 3. Porovnej výtěžek vody při kompletní oxidaci 1 g glukosy s výtěžkem vody z 1 g tripalmitoylglycerolu. Dejte výsledky do souvislosti s evolučním výběrem obsahu velbloudího hrbu. 4. Příjem velkého množství glukosy před startem maratonu by mohlo být způsobem jak zvýšit energetickou zásobu organismu. Zkušení běžci takto nečiní. Jaký je biochemický důvod vyhnout se tomuto způsobu zisku energie před startem (uvažuj vliv glukosy na hladinu insulinu).")
68
5. Cyklisté na Tour der France (více než km během tří týdnů) spotřebují asi kcal energie, což činí cca kcal/den. (odpočívající muž cca kcal/den). A) Při úvaze, že energetický výtěžek z ATP je 12 kcal/mol a ATP má mol. hmotnost 503 vypočti, kolik ATP spotřebuje cyklista při Tour. B) čistý ATP lze zakoupit za 150 USdol. za gram. Kolik by stála energie ATP spotřebovaná při Tour, kdyby ji bylo nutné zakoupit ? 6. Hormon glukagon je signálem hladovění a inhibuje glykolýzu v játrech. Jaký užitek pro organismus má tato inhibice energii produkující dráhy ?
 Při úvaze, že energetický výtěžek z ATP je 12 kcal/mol a ATP má mol. hmotnost 503 vypočti, kolik ATP spotřebuje cyklista při Tour. B) čistý ATP lze zakoupit za 150 USdol. za gram. Kolik by stála energie ATP spotřebovaná při Tour, kdyby ji bylo nutné zakoupit 6. Hormon glukagon je signálem hladovění a inhibuje glykolýzu v játrech. Jaký užitek pro organismus má tato inhibice energii produkující dráhy")
70
Pohodové Vánoce a mnoho životních a studijních úspěchů v roce 2007.

Podobné prezentace









.>")












