Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Mendelistická genetika
FOTO Lenka Hanusová, 2013

2
Pojem mendelismus - vysvětluje dědičnost kvalitativních znaků a jejich chování v následující populaci - Je základem tzv. hybridologické analýzy – analýzy křížení

3
Hybridizace Základní studijní a pracovní metoda hybridologické analýzy
Křížení (hybridizace) = (obvykle cílené) pohlavní rozmnožování dvou vybraných jedinců opačného pohlaví s rozdílnými genotypy Cílem hybridizace: Genetický výzkum Šlechtitelský záměr
 = (obvykle cílené) pohlavní rozmnožování dvou vybraných jedinců opačného pohlaví s rozdílnými genotypy. Cílem hybridizace: Genetický výzkum. Šlechtitelský záměr.")
4
Základní pojmy Gen – úsek DNA se specifickou funkcí.
Strukturní gen – úsek DNA nesoucí genetickou informaci pro polypeptidový řetězec. Alela – varianta genu (odlišující se od ostatních fenotypovým projevem). Genotyp – konkrétní sestava alel v jednom genu, nebo více genech, nebo u jedince. Determinuje fenotypové možnosti nositele. Fenotyp – soubor projevených vnějších znaků a vlastností jedince.
. Genotyp – konkrétní sestava alel v jednom genu, nebo více genech, nebo u jedince. Determinuje fenotypové možnosti nositele. Fenotyp – soubor projevených vnějších znaků a vlastností jedince.")
5
Základní pojmy Kvalitativní znaky – často kódovány geny velkého účinku – tzv. major geny. Znak kóduje eden nebo několik málo genů. Monohybrid (Aa) je kříženec (heterozygot) vzniklý spojením homozygotních rodičů odlišných v jednom znaku (genu). Monohybridismus – sledování jednoho kvalitativního znaku. Dihybrid (Aa, Bb) je kříženec (heterozygot) vzniklý spojením rodičů homozygotů odlišných ve dvou znacích Dihybridismus – sledování dvou znaků současně. Tri-polyhybridismus – sledování 3 a více znaků současně.
 je kříženec (heterozygot) vzniklý spojením homozygotních rodičů odlišných v jednom znaku (genu). Monohybridismus – sledování jednoho kvalitativního znaku. Dihybrid (Aa, Bb) je kříženec (heterozygot) vzniklý spojením rodičů homozygotů odlišných ve dvou znacích. Dihybridismus – sledování dvou znaků současně. Tri-polyhybridismus – sledování 3 a více znaků současně.")
6
Podvojné založení dědičnosti
Gen může mít u jedince 2 různé formy – alely. Alely téhož genu jsou uloženy na stejném místě (lokusu) homologních chromozomů Jedinec od každého rodiče dostane 1 alelu Kvalitativně shodné alely = homozygotní genotyp (AA, aa) Dominantní homozygot – AA Recesivní homozygot - aa Kvalitativně rozdílné alely = heterozygotní genotyp (Aa)
 homologních chromozomů. Jedinec od každého rodiče dostane 1 alelu. Kvalitativně shodné alely = homozygotní genotyp (AA, aa) Dominantní homozygot – AA. Recesivní homozygot - aa. Kvalitativně rozdílné alely = heterozygotní genotyp (Aa)")
7
Vztahy mezi alelami dominance – dominantní alela převládá nad ostatními a projeví se vždy ve fenotypu recesivita –recesivní alela je překryta účinkem dominantní formy, ve fenotypu se projeví pouze v homozygotním stavu neúplná dominance – obě alely se ve fenotypu projeví současně kodominance – alely se projeví ve fenotypu nezávisle na sobě (krevní skupiny) superdominance – heterozygotní konstituce je aktivnější než obě homozygotní
 superdominance – heterozygotní konstituce je aktivnější než obě homozygotní.")
8
Značení P – rodičovská generace (z lat. parentes)
F – generace potomků (z lat. filius, filia) číselný index označující pořadí F1 – první filiální generace po křížení rodičů F2 – druhá filiální generace vzniklá křížením hybridů F1 B1 – výsledek křížení hybrida F1 – s některou rodičovskou variantou (B – backcross)
 číselný index označující pořadí. F1 – první filiální generace po křížení rodičů. F2 – druhá filiální generace vzniklá křížením hybridů F1. B1 – výsledek křížení hybrida F1 – s některou rodičovskou variantou (B – backcross)")
9
Johann Gregor Mendel (1822 – 1884)
Zakladatel moderní genetiky Uplatnil experimentální přístup 1866 – přednáška Experimenty v křížení rostlin „dědičný faktor“ Volná kombinovatelnost vloh a segregace

10
Rostlinu nehodnotil jako celek, sledoval na rostlině konkrétní alernativu (vlohu) vybraného znaku.
 vybraného znaku.")
11
Mendelův postup 1. krok – vytvoření rodičovských čistých linií
2. krok – tvorba F1 generace 3. krok – tvorba F2 generace samoopylením rostlin generace F1 - Vedle dominantního znaku se objevil i znak recesivní v poměru 3 : 1 (fenotypový štěpný poměr)
")
12
Mendelova pravidla dědičnosti
1. uniformita F1 generace 2. identita reciprokých křížení 3. čistota vloh a štěpení 4. volná kombinovatelnost vloh

13
Zápis křížení Mendelistický čtverec
Rodičovská generace P: AA x aa genotyp matky genotyp otce genomy gamet P generace: A a Generace hybridů F1: Aa genotyp hybridů genomy gamet F1 generace: A; a F1: Aa x Aa genomy samčích gamet Generace hybridů F2 A a genomy samičích gamet A a

14
1. pravidlo o uniformitě hybridů F1
x P: BB x bb gamety: B B b b potomci F1 Bb Bb Bb Bb

15
2. pravidlo o identitě reciprokých kříženců
P: bb x BB gamety: b b B B potomci F1 Bb Bb Bb Bb

16
3. pravidlo - čistota vloh a štěpení: segregace
geny heterozygota se předávají další generaci v „čisté“ podobě segregace je důsledkem redukčního dělení gamet Aa x Aa A a A a genomy samčích gamet generace hybridů F2 A a AA Aa aa A a genotypy jedinců F2 genomy samičích gamet tj. genotypový štěpný poměr AA : 2Aa : 1aa fenotypový štěpný poměr dominantní : 1 recesivní

17
Štěpné poměry v F2 generaci při monohybridním štěpení – úplná dominance
F1: Aa Gamety: A, a (A – 50% pravděpodobnost vzniku, a – 50% pravděpodobnost vzniku) F2: Aa x Aa A 0,55 a A 0,5 AA 0,5*0,5 = 0,25 Aa a 0,5 aa Genotypový štěpný poměr v F2: 1 (AA) : 2 (Aa) : 1 (aa) Fenotypový štěpný poměr v F2: 3 (dominantní); A-) : 1 (recesivní, aa)
 F2: Aa x Aa. A. 0,55. a. A 0,5. AA. 0,5*0,5 = 0,25. Aa. a 0,5. aa. Genotypový štěpný poměr v F2: 1 (AA) : 2 (Aa) : 1 (aa) Fenotypový štěpný poměr v F2: 3 (dominantní); A-) : 1 (recesivní, aa)")
18
Zpětné křížení důkaz, že heterozygot monohybrid tvoří 2 druhy gamet v poměru 1 : 1 křížení hybrida F1 generace s některým z homozygotních rodičů nebo s jedincem nesoucím rodičovský genotyp

19
Zpětné testovací křížení
Zaměřeno na odhalení genotypu testovaného jedince – analýza křížení Jedná se o křížení recesivního homozygota a testovaného jedince s dominantním fenotypem, ale pro nás neznámým genotypem Genotyp zjistíme na základě vlastností výsledných potomků (kříženců)
")
20
x P1: bb BB F1 : Bb x x B1 BB x Bb B1 bb x Bb BB Bb Bb bb

21
Intraalelické interakce
Úplná dominance – projev dominantní alely převládá nad ostatními a v fenotypu se projeví vždy. Neúplná dominance – heterozygoti jsou fenotypově odlišní od rodičovských homozygotů, znak je vyjádřen intermediálně Kodominance – alely se projeví ve fenotypu heterozygota samostatně, nezávisle na sobě

22
Neúplná dominance P WW ww F1 Ww x 1 WW Ww ww

23
4. pravidlo o volné kombinovatelnosti vloh
Sledujeme více než 1 gen rozchod alel různých genů nezávisle na sobě při gametogenezi, tj. lokusy na různých chromozómových párech - vzniká tolik typů gamet, kolik je možných kombinací, tedy monohybrid 2 typy gamet 1 : 1 dihybrid 4 typy gamet 1 : 1 : 1 : 1

24
Dihybridní křížení G – žlutý W - kulatý g - zelený w - svraštělý
P : GGWW x ggww nebo GGww x ggWW GW GW gw gw Gw Gw gW gW F1 : GgWw gamety: GW Gw gW gw

25
F2:Mendelistický čtverec
GW Gw gW gw GGWW GGWw GgWW GgWw GGww Ggww ggWW ggWw ggww genotypový štěpný poměr 1: 2 : 1 : 2 : 4 : 2 : 1 : 2 : 1 fenotypový štěpný poměr : 3: 3 : 1 šlechtitelské novinky úhlopříčka homozygotů úhlopříčka heterozygotů

26
Zpětné křížení ggww x GgWw GGWW x GgWw ggWW x GgWw GGww x GgWw
genotypový štěpný poměr 1 : 1 : 1 : 1 fenotypový štěpný poměr 1 žlutý kulatý 1 fenotyp genotypový štěpný poměr 1 : 1 : 1 : 1 fenotypový štěpný poměr 1 : 1 : 1 : fenotypy ggWW x GgWw GGww x GgWw genotypový štěpný poměr 1 : 1 : 1 : 1 fenotypový štěpný poměr žlutý kulatý : 1 zelený kulatý 2 fenotypy genotypový štěpný poměr 1 : 1 : 1 : 1 fenotypový štěpný poměr 1 žlutý kulatý : 1žlutý svraštělý 2 fenotypy

27
Polyhybridní křížení rodiče se odlišují ve více než dvou znacích počty gamet: monohybrid 2, dihybrid 4, trihybrid 8, tetrahybrid 16, polyhybrid 2n n = počet znaků, v kterých je heterozygotní odvození frekvence kombinací – rozvětvovací metoda

28
Trihybridní křížení P1: AABBCC X aabbcc gamety ABC abc F1 AaBbCc
AbC Abc aBC aBc abC abc

29
Fenotypy trihybridů v F2
A nebo a B nebo b C nebo c štěpný poměr 3/4C – (3/4)(3/4)(3/4)ABC = 27/64 A-B-C- 3/4B 1/4c – (3/4)(3/4)(1/4)ABc = 9/64 A-B-cc ¾ A 3/4C – (3/4)(1/4)(3/4)AbC = 9/64 A-bbC- 1/4b 1/4c – (3/4)(1/4)(1/4)Abc = 3/64 A-bbcc 3/4C – (1/4)(3/4)(3/4)aBC = 9/64 aaB-C- 3/4B 1/4c – (1/4)(3/4)(1/4)aBc = 3/64 aaB-cc 1/4 a 3/4C – (1/4)(1/4)(3/4)abC = 3/64 aabbC- 1/4b 1/4c – (1/4)(1/4)(1/4)abc = 1/64 aabbcc
(3/4)(3/4)ABC = 27/64 A-B-C- 3/4B. 1/4c – (3/4)(3/4)(1/4)ABc = 9/64 A-B-cc. ¾ A. 3/4C – (3/4)(1/4)(3/4)AbC = 9/64 A-bbC- 1/4b. 1/4c – (3/4)(1/4)(1/4)Abc = 3/64 A-bbcc. 3/4C – (1/4)(3/4)(3/4)aBC = 9/64 aaB-C- 3/4B. 1/4c – (1/4)(3/4)(1/4)aBc = 3/64 aaB-cc. 1/4 a. 3/4C – (1/4)(1/4)(3/4)abC = 3/64 aabbC- 1/4b. 1/4c – (1/4)(1/4)(1/4)abc = 1/64 aabbcc.")
30
Výpočet genomů gamet A nebo a B nebo b C nebo c štěpný poměr
0,5 C – (0,5)(0,5)(0,5)ABC = 0,125 ABC 0,5 B 0,5 c - (0,5)(0,5)(0,5)ABc = 0,125 ABc 0,5 A 0,5 C – (0,5)(0,5)(0,5)AbC = 0,125 AbC 0,5 b 0,5 c – (0,5)(0,5)(0,5)Abc = 0,125 ABC 0,5 C – (0,5)(0,5)(0,5)aBC = 0,125 aBC 0,5 B 0,5 c – (0,5)(0,5)(0,5)aBc = 0,125 aBc 0,5 a 0,5 C – (0,5)(0,5)(0,5)abC = 0,125 abC 0,5 b 0,5 c – (0,5)(0,5)(0,5)abc = 0,125 abc
(0,5)(0,5)ABC = 0,125 ABC. 0,5 B. 0,5 c - (0,5)(0,5)(0,5)ABc = 0,125 ABc. 0,5 A. 0,5 C – (0,5)(0,5)(0,5)AbC = 0,125 AbC. 0,5 b. 0,5 c – (0,5)(0,5)(0,5)Abc = 0,125 ABC. 0,5 C – (0,5)(0,5)(0,5)aBC = 0,125 aBC. 0,5 B. 0,5 c – (0,5)(0,5)(0,5)aBc = 0,125 aBc. 0,5 a. 0,5 C – (0,5)(0,5)(0,5)abC = 0,125 abC. 0,5 b. 0,5 c – (0,5)(0,5)(0,5)abc = 0,125 abc.")
31
Trihybridní křížení AaBbCc x AaBbCc
0,5 a AA 0,25 Aa aa B 0,5 b BB 0,25 Bb bb C 0,5 c CC 0,25 Cc cc Fenotypový poměr A- : a- : 1 0,75 : 0,25 Fenotypový poměr A- : a- : 1 0,75 : 0,25 Fenotypový poměr A- : a- : 1 0,75 : 0,25 S jakou pravděpodobností vznikne jedinec s fenotypem A-bbC-? 0,75*0,25*0,75 = 0, %

32
Zobecnění pro F2 generaci
počet vzorec n = 1 n = 2 n = 3 n = 4 gametických kombinací 2n 2 4 8 16 nejmenší úplná generace 4n 64 256 genotypů 3n 3 9 27 81 homozygotů homoz. rekombinací 2n – 2 - 6 14 heterozygotů 22n – 2n 12 56 240 fenotypů

33
Štěpení v F2 generaci při úplné dominanci
fenotypový štěpný poměr (3 + 1)1 3 : 1 (3 + 1)2 9 : 3 : 3 : 1 (3 + 1)3 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1 (3 + 1)4 87 : 27 : 27 : 27 : 27 : 9 : 9 : 9 : 9 : 3 : 3 : 3 : 3 : 1
1. 3 : 1. (3 + 1)2. 9 : 3 : 3 : 1. (3 + 1)3. 27 : 9 : 9 : 9 : 3 : 3 : 3 : 1. (3 + 1)4. 87 : 27 : 27 : 27 : 27 : 9 : 9 : 9 : 9 : 3 : 3 : 3 : 3 : 1.")
34
Nejmenší velikost úplné F2
teoretická při pravděpodobném výskytu homozygota 95 % 99 % 99,9 % 4 11 16 24 47 72 107 64 134 297 442

35
Dědičnost kvalitativních znaků není náhodná, ale pravidelná.
Všechny štěpné poměry v genetice jsou založeny na zákonech velkých čísel. Štěpný poměr je poměr statistický, tj. uplatní se jen při dostatečném počtu potomků, (sta – tisíce). Je-li počet potomků malý, štěpný poměr se od ideálních teoretických čísel více či méně liší.
. Je-li počet potomků malý, štěpný poměr se od ideálních teoretických čísel více či méně liší.")
36
χ2 test test pro ověření shody skutečných a teoretických štěpných poměrů d – rozdíl mezi skutečným a očekávaným počtem potomků ve třídách e – očekávaný počet potomků ve třídách

37
Podmínky platnosti MZ 1 gen = 1 znak
Geny neleží na pohlavních chromozomech (autozomální dědičnost) Každý gen leží na jiném chromozomu.
 Každý gen leží na jiném chromozomu.")
38
Vazba vloh

39
Thomas Hunt Morgan (1866 – 1945) americký genetik a embryolog
pokusy s octomilkou (D. melanogaster)
")
40
Morganova pravidla geny jsou na chromozómu uspořádány lineárně za sebou počet vazbových skupin odpovídá haploidnímu počtu chromozómů Výjimky: přeskupování genů nebo jejich částí Překřížení a rekombinace během meiózy (několik vazbových skupin na 1 chromozomu)
")
41
Geny na chromozomu a) vazbová skupina b) syntenní skupina
 vazbová skupina b) syntenní skupina")
42
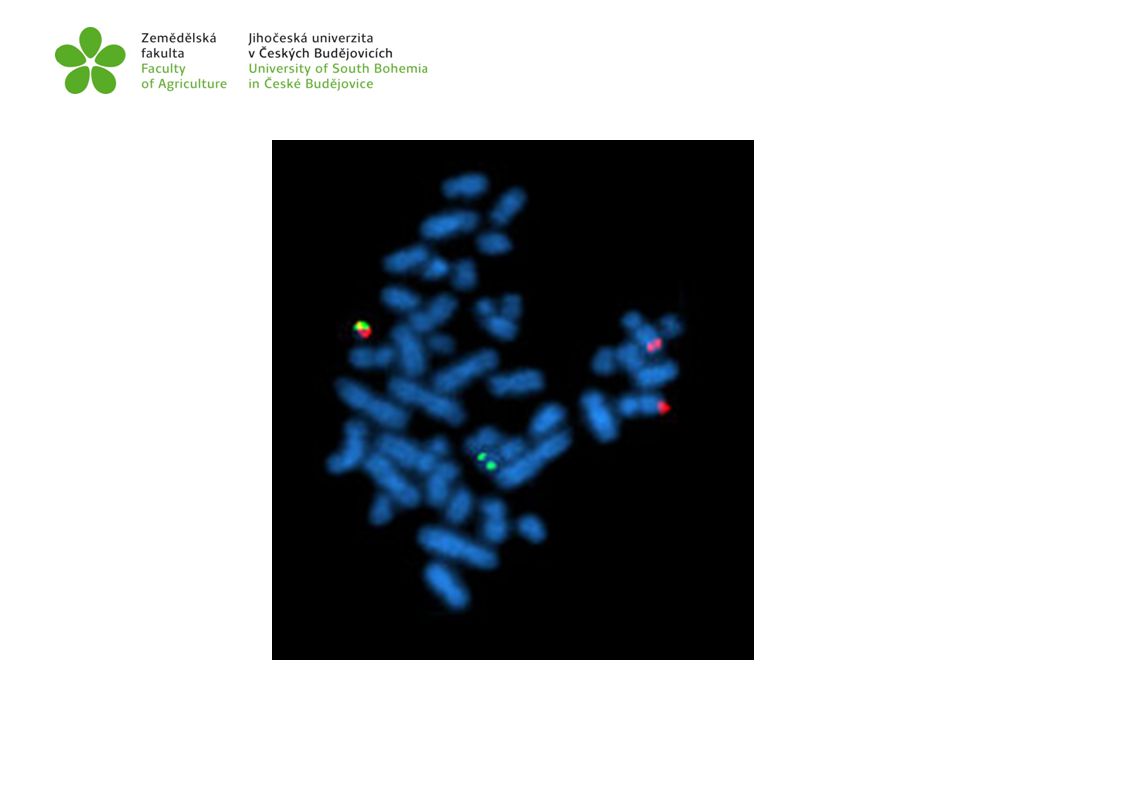
Syntenní skupina geny lokalizované na stejném chromozómu jsou vzdáleny natolik, že se chovají jako nezávislé pravděpodobnost rekombinací je 50% (vzdálenost mezi geny > než 50cM) Prokazujeme ji hybridologickou analýzou (genetické mapování) nebo cytogenetickými a molekulárně-genetickými metodami (in situ hybridizace, FISH)
 Prokazujeme ji hybridologickou analýzou (genetické mapování) nebo. cytogenetickými a molekulárně-genetickými metodami (in situ hybridizace, FISH)")
44
Vazbová skupina blízké umístění podíl rekombinací < 50% geny se nemusí přenášet společně Pravděpodobnost, že se geny nepřenesou společně, je pravděpodobností výskytu crossing-overů.

45
Druhy vazby geny zůstávají stále ve stejné vazbové skupině - mezi chromozomy neprobíhá c. o. Úplná: geny mohou přecházet z jednoho homolog. chromozomu na druhý - mezi chromozomy probíhá c. o. - vznik rekombinantních gamet, jejichž frekvence je vždy nižší než frekvence rodičovských, nerekombinovaných gamet Neúplná:

46
Crosing over v meiotické profázi I
Příčina neúplné vazby Crosing over v meiotické profázi I

47
CROSSING-OVER párování homologických chromozomů (bivalent)
chromozomy s rekombinovanými chromatidami

48
DVOJITÝ CROSSING-OVER

49
Pravděpodobnost θ (theta)
podíl rekombinovaných gamet ze všech sledovaných 0.00 při úplné vazbě bez rekombinace vzdálenost genů je velká - volná kombinovatelnost

50
Vzdálenost mezi geny cM centimorgan
1 cM je pravděpodobnost rekombinací mezi lokusy 1% chromozóm č. 21 = 0,5 M (50 cM) chromozóm č. 1 = 2 M (200 cM) lidský haploidní genom = 30M = 3 000cM
 chromozóm č. 1 = 2 M (200 cM) lidský haploidní genom = 30M = 3 000cM.")
51
Rekombinace zvyšuje počet geneticky různých gamet
rekombinace, náhodný rozchod chromozómů a chromatid a náhodné spojení gamet vytváří prakticky nevyčerpatelný počet kombinací

52
Vazbová fáze cis (coupling)
na jednom chromozómu alely dominantní, na druhém recesívní P: AB ab x F1: AB ab

53
Vazbová fáze cis (coupling)
AB ab A B A B a b a b

54
Vznikají gamety rodičovské AB, ab rekombinované Ab, aB
Vazbová fáze cis Vznikají gamety rodičovské AB, ab rekombinované Ab, aB A B A b a B a b

55
Vazbová fáze trans (repulsion)
na jednom chromozómu alela dominantní a recesívní, na druhém recesívní a dominantní P: Ab aB x F1: Ab aB

56
Vazbová fáze trans (repulsion)
Ab aB A b A b a B a B

57
Vznikají gamety rodičovské Ab, aB rekombinované AB, ab
Vazbová fáze trans Vznikají gamety rodičovské Ab, aB rekombinované AB, ab A B A b a B a b

58
Vazbová fáze cis Zpětné křížení AB/ab X ab/ab gamety: AB Ab aB ab ab
genotypy: AB/ab (a1) Ab/ab (a2) aB/ab (a3) ab/ab(a4) pokud existuje úplná vazba mezi lokusy „A“ a „B“, je na 100 potomků AB/ab (a1) Ab/ab (a2) aB/ab (a3) ab/ab (a4)
 Ab/ab (a2) aB/ab (a3) ab/ab(a4) pokud existuje úplná vazba mezi lokusy „A a „B , je na 100 potomků. AB/ab (a1) Ab/ab (a2) aB/ab (a3) ab/ab (a4)")
59
Při úplné vazbě je na 100 potomků
Vazbová fáze trans Zpětné křížení Ab/aB x aa/bb gamety: AB Ab aB ab ab Při úplné vazbě je na 100 potomků AB/ab (a1) Ab/ab (a2) aB/ab (a3) ab/ab (a4)
 Ab/ab (a2) aB/ab (a3) ab/ab (a4)")
60
AaBb (a1) = 25 Aabb (a2) = 25 aaBb (a3) = 25 aabb (a4) = 25
Bez vazby AaBb (a1) = 25 Aabb (a2) = 25 aaBb (a3) = 25 aabb (a4) = 25
 = 25 Aabb (a2) = 25 aaBb (a3) = 25 aabb (a4) = 25.")
61
A – nafialovělá barva aleuronu kukuřičného zrna
a - žlutá barva B – kulaté zrno b - hranaté zrno Testovací křížení AaBb x aabb, úplná vazba: cis: AB/ab x aabb 50% zrn nafialovělých kulatých (a1) AB/ab 50% zrn žlutých hranatých (a4) ab/ab žádné fialové hranaté ani žluté kulaté trans: Ab/aB 50% zrn nafialovělých hranatých (a2) Ab/ab 50% zrn žlutých kulatých (a3) aB/ab žádné fialové kulaté ani žluté hranaté
 AB/ab. 50% zrn žlutých hranatých (a4) ab/ab. žádné fialové hranaté ani žluté kulaté. trans: Ab/aB. 50% zrn nafialovělých hranatých (a2) Ab/ab. 50% zrn žlutých kulatých (a3) aB/ab. žádné fialové kulaté ani žluté hranaté.")
62
Neúplná vazba výskyt rekombinantů, jejichž frekvence je vždy nižší než frekvence rodičovských nerekombinovaných genotypů; % rekombinant zjistíme z dvojnásobných testovacích křížení: AaBb x aabb

63
Musíme znát vazbovou fázi!
Morganovo číslo p % podíl rekombinant ze všech informativních potomků Musíme znát vazbovou fázi!

64
Musíme znát vazbovou fázi!
Batesonovo číslo c Kolikrát častěji vznikají gamety rodičovské, než rekombinované Musíme znát vazbovou fázi!

65
Určení vazbové fáze z genotypu potomků zpětného křížení, zpětná dedukce genotypů rodičů a prarodičů

66
převaha nerekombinant a1, a4, tzn. fáze cis
AaBb x aabb AaBb (a1) Aabb (a2) aaBb (a3) aabb (a4) převaha nerekombinant a1, a4, tzn. fáze cis
 Aabb (a2) aaBb (a3) aabb (a4) převaha nerekombinant a1, a4, tzn. fáze cis.")
67
AaBb x aabb AaBb (a1) Aabb (a2) aaBb (a3) aabb (a4) převaha nerekombinant a2, a3 , tj. fáze trans

68
Pořadí genů na chromozomu lze určit analýzou dvojitých crossing – overů:
tříbodový test

69
A B A B C

70
Lineární vztah mezi % rekombinant a vzdáleností v cM je do cca cM. Kvůli dvojitým a vícenásobným crossing overům se vzdálenosti v genomu jakoby prodlužují.

71
Interference Jsou rekombinace na jednom chromozómu na sobě nezávislé? a . b Frekvence dvojitého c.-o. = [%] 100 a - frekvence jednoduchého c.-o. v jedné oblasti b - frekvence jednoduchého c.-o. v druhé oblasti Př.: 23,84 . 6,16 / 100 = 1,47 % ale skutečná frekvence dvoj. c.-o. = 0,72%

72
Tj. potlačení vzniku dalšího c. -o. na chromozómu, kde již jeden c. -o
Tj. potlačení vzniku dalšího c.-o. na chromozómu, kde již jeden c.-o. proběhl. Příčina – mechanické schopnosti chromozómu překřížit se na více místech. Další příčiny, protože interference se vyskytuje i mezi geny vzdál. více než 30 cM.

73
skutečný podíl dvoj. c.-o.
Interference se vyjadřuje jako skutečný podíl dvoj. c.-o. Koef. koincidence = teor. podíl dvoj. c.-o. 0,72 / 1,47 = 0,49

74
Délkové a rekombinační hodnoty jsou však zkresleny také nerovnoměrnou frekvencí rekombinací v různých částech genomu, v jednotlivých chromozomech a jejich segmentech. Existují tzv. horká místa (hot spots) s častou tvorbou chiasmat, na druhé straně dlouhé segmenty cM, v nichž je suprese rekombinací a pokud nastanou, mají často patologické následky, např. T/t genový komplex u myší, projevující se řadou malformací a infertilitou.
 s častou tvorbou chiasmat, na druhé straně dlouhé segmenty cM, v nichž je suprese rekombinací a pokud nastanou, mají často patologické následky, např. T/t genový komplex u myší, projevující se řadou malformací a infertilitou.")
75
Nevýhody výpočtu Morganova čísla odstraňuje metoda lod skóre:
lze určit existenci vazby a souč. její sílu výsledky segregace potomků z různých křížení lze jednoduše akumulovat k vyhodnocení stačí dvougenerační rodokmen (P a F1), v P nemusí být známa vazbová fáze pro důkaz vazby stačí minimální počet potomků potomci nemusí pocházet jen z dvojnásobných testovacích křížení, tj. AaBb x aabb, ale i z jiných typů, jednoduché nebo dvojnás. intercrossy AaBb x Aabb; AaBb x AaBb bez dominance či s dominancí v jedné nebo obou alelách.
, v P nemusí být známa vazbová fáze. pro důkaz vazby stačí minimální počet potomků. potomci nemusí pocházet jen z dvojnásobných testovacích křížení, tj. AaBb x aabb, ale i z jiných typů, jednoduché nebo dvojnás. intercrossy AaBb x Aabb; AaBb x AaBb bez dominance či s dominancí v jedné nebo obou alelách.")
76
θ = 0, úplná vazba θ = 0,01-0,20 úzká (těsná) vazba θ = 0,21-0,35 středně těsná vazba θ = 0,36-0,49 volná vazba θ = 0, volná kombinovatelnost

77
Lod skóre - statistická metoda pro detekci vazby z poměru dvou pravděpodobností Log logaritmus odds šance pro převahu pravděpodobnosti skóre je zde poměr skutečných nebo hypotetických rekombinant k nerekombinantám

78
Lod skóre Výsledek Lod skóre z jednoho typu křížení (páření, rodiny) se sčítá s dalšími Z1, Z2, Z3… SZ. Je-li akumulované Lod 3 pak je pravděpodobnost 1000 : 1 ve prospěch vazby. Jestliže akumulované Lod -2, jsou geny volně kombinovatelné s pravděpodobností 1:100. Je-li vypočtená hodnota mezi -2 a 3, nelze učinit rozhodnutí, je nutné pokračovat v analýzách.

79
Vazbová nerovnováha (disekvilibrium)
nerovnoměrná frekvence 4 možných kombinací (AB, Ab, aB, ab) v populaci častější výskyt kombinací 2 alel příčiny: evoluční výhoda (pouze pro cis) rovnováha dosud nebyla navozena pro krátký evoluční interval
 v populaci. častější výskyt kombinací 2 alel. příčiny: evoluční výhoda (pouze pro cis) rovnováha dosud nebyla navozena pro krátký evoluční interval.")
80
Biologický význam vazby
uchování stálé sestavy funkčně spřízněných genů evoluční konzervace genových rodin (funkční geny + pseudogeny) funkční záloha – změnou podmínek prostředí mohou být pseudogeny „zapnuty“
 funkční záloha – změnou podmínek prostředí mohou být pseudogeny „zapnuty")
81
Biologický význam rekombinace
zvyšování proměnlivosti při uchování vazbové lokalizace genů vznik nových genotypů

82
Biologický význam vazby a rekombinace
Pro schopnost populace přizpůsobit svůj genofond změněným podmínkám má význam i uspořádání genů na chromozomech. Na určitou vlastnost působí geny XYZ. Je-li optimální fenotyp intermediární, je optimální genotyp heterozygotní XxYyZz. Potom je výhodná velmi silná vazba, protože nejvýhodnějším genotypem je XyZ/xYz (trans), zaručuje heterozygotnost, suboptimální genotypy vznikají s mnohem menší pravděpodobností než při volné kombinovatelnosti vloh, zachovává se maximální genetická variabilita.
, zaručuje heterozygotnost, suboptimální genotypy vznikají s mnohem menší pravděpodobností než při volné kombinovatelnosti vloh, zachovává se maximální genetická variabilita.")
83
Dojde-li k takové změně prostředí, že optimální bude homozygotní genotyp XYZ/XYZ nebo xyz/xyz, je vazba méně výhodná, neboť k přechodu na vazbovou fázi cis XYZ/xyz je nutný crossing-ower. Mutací vzniklé alely tedy musí být rekombinacemi zapojeny do genofondu populace.

84
Genové interakce

85
Působení genů Gen Znak

86
G1 G2 G3 G4 Gn Znak 1 Znak 2 G1 Znak 1 Znak 2 např.: aa Monomerie
Polymerie G1 Znak Znak 2 Pleiotropie např.: aa Znak 1 (barva) Znak 2 (plodnost) (srpk. krvinky) (anémie)
 Znak 2 (plodnost) (srpk. krvinky) (anémie)")
87
Genové interakce - interalelické
- intraalelické - interalelické A a intraalelické - typ dědičnosti, tj. dominance, neúplná dominance, kodominance, superdominance A a B b interalelické - genové interakce - typ genových interakcí

88
Dihybridní křížení P AABB x aabb F1 AaBb F2 9 A-B- : 3 A-bb : 3 aaB-
♂ ♀ 9 A-B- : 3 A-bb : aaB- : aabb

89
Interakce bez změny štěpného poměru
A-B- ořechovitý hřebínek A-bb růžicovitý hřebínek aaB- hráškovitý hřebínek aabb listovitý hřebínek Značka r : 3 růžicovitý : hráškovitý : listovitý 9 ořechovitý

90
Kompenzace A-bb prohnutí aaB- prohnutí A-B- kompenzace 10 : 3 : 3
(9 A-B- + 1 aabb) : 3 (aaB-) : 3 (A-bb )
 : 3. (aaB-) : 3. (A-bb )")
91
Inhibice A červená, a bílá barva peří B (I) inhibitor, BA : 3 červená
(9 A-B- + 3 aaB- + 1 aabb) : 3 červená (A-bb)
 : 3 červená. (A-bb)")
92
Inhibice A červená, a bílá barva peří B (I) inhibitor, BA
Inhibice je příčinou jevu, kdy stejný znak se někdy dědí jako dominantní, jindy jako recesívní. U subpopulací (plemen, odrůd apod.), u nichž se vyskytuje inhibitor B (I), se bílá barva dědí jako dominantní (genotypy --BB). U plemen, u nichž se inhibitor nevyskytuje, se bílá barva dědí jako recesívní (genotypy A-bb červená barva, aabb bílá).
, u nichž se vyskytuje inhibitor B (I), se bílá barva dědí jako dominantní (genotypy --BB). U plemen, u nichž se inhibitor nevyskytuje, se bílá barva dědí jako recesívní (genotypy A-bb červená barva, aabb bílá).")
93
Dominantní epistáze W – bílá barva srsti u psů B – černá b - hnědá
Epistáze W nad B WB Wb wB wb WWBB WWBb WwBB WwBb WWbb Wwbb wwBB wwBb wwbb 12 (9 W-B- + 3 W-bb) bílá : 3 (wwB-) černá : 1 aabb hnědá
 bílá. : 3 (wwB-) černá. : 1 aabb. hnědá.")
94
Recesivní epistáze Králíci: A - divoké zbarvení (aguti)
a - černá barva c - blokuje tvorbu pigmentu Epistáze cc nad A; a CA Ca cA ca CCAA CCAa CcAA CcAa CCaa Ccaa ccAA ccAa ccaa 9 (C-A-) aguti : 3 (C-aa) černá : (cc--) albín
 aguti. : 3 (C-aa) černá. : 4 (cc--) albín.")
95
Komplementární faktory (dvojitá recesívní epistáze)
aa B- bb A- A-B- normální slyšení aaB- A-bb hluchota aabb 9 : 7 (3aaB- + 3 A-bb + 1 aabb) slyšící neslyšící
 slyšící. neslyšící.")
96
Duplicitní faktory nekumulativní s dominancí
A,B alely pro opeření běháků a, b běháky neopeřují 15 (9 A-B- + 3 A B-) : (aabb) opeřené běháky neopeřené běháky
 : 1 (aabb) opeřené běháky. neopeřené běháky.")
97
Duplicitní faktory kumulativní s dominancí
A-bb B-aa A-B- – diskové plody dýně tmavohnědé obilky ječmene aabb – podlouhlé dýně světlé obilky ječmene kulaté dýně světle hnědé obilky ječmene 9 (A-B-) diskové : (3 A-bb + 3 aaB-) kulaté : 1 (aabb) podlouhlé
 diskové. : 6 (3 A-bb + 3 aaB-) kulaté. : 1 (aabb) podlouhlé.")
98
Duplicitní faktory kumulativní bez dominance
Zbarvení obilek pšenice, barva kůže lidí aj. Účinek dominantních alel se sčítá bez ohledu na příslušnost k alelickému páru. ADITIVITA Dominantní alela = aktivní alela Štěpný poměr 1 4 : 4 3 : 6 2 : 4 1 : 1 ZESVĚTLOVÁNÍ Aktivních alel

99
n počet zúčastněných alel
Duplicitní faktory kumulativní bez dominance (a + b)n n počet zúčastněných alel (a + b)4 1a4 + 4a3b + 6a2b2 + 4ab3 + 1b4 Štěpný poměr počet alel
n. n počet zúčastněných alel. (a + b)4. 1a4 + 4a3b + 6a2b2 + 4ab3 + 1b4. Štěpný poměr počet alel.")
100
délka uší u králíků Triplicitní kumulativní faktory bez dominance
geny L1, L2, L3 genotyp l1l1l2l2l3l3 fenotyp 10 cm 1 aktivní alela = 2cm genotyp L1L1L2L2L3L3 fenotyp 22 cm

101
Triplicitní kumulativní faktory bez dominance
(délka uší u králíků) P L1L1L2L2L3L x l1l1l2l2l3l3 22 cm cm F1 L1l1L2l2L3l3 16cm F2 štěpný poměr : : : : : : akt. alel cm 1 6 22 6 5 20 15 4 18 20 3 16 15 2 14 6 1 12 1 10
 P L1L1L2L2L3L3 x l1l1l2l2l3l3. 22 cm 10 cm. F1 L1l1L2l2L3l3. 16cm. F2. štěpný poměr : : : : : : akt. alel. cm")
102
Triplicitní kumulativní faktory bez dominance
(a + b)n (a + b)6 1a6+6a5b+15a4b2+20a3b3+15a2b4+6ab5+1b6
n. (a + b)6. 1a6+6a5b+15a4b2+20a3b3+15a2b4+6ab5+1b6.")
103
n počet zúčastněných alel
Štěpný poměr - distribuce fenotypů Rozvinutý binom (1+1)n Pascalův trojúhelník n počet zúčastněných alel (1+1)0 (1+1)1 (1+1)2 (1+1)3 (1+1)4 (1+1)5 (1+1)6 (1+1)7 (1+1)8 (1+1)9 (1+1)10 1
n. Pascalův trojúhelník. n počet zúčastněných alel. (1+1)0. (1+1)1. (1+1)2. (1+1)3. (1+1)4. (1+1)5. (1+1)6. (1+1)7. (1+1)8. (1+1)9. (1+1)")
104
Počet fenotypů 2n Počet rozdílných genotypů 3n n = počet alelických párů 210 = = 220 = =

105
Normální rozdělení četností
Distribuce fenotypů při vysokém počtu zúčastněných lokusů Normální rozdělení četností y Počet jedinců x x Hodnota znaku

106
U polygenní dědičnosti je fenotyp kromě genů ovlivněn prostředím, rozdělení fenotypů je kontinuální.

107
Znaky Kvalitativní Kvantitativní proměnlivost vliv vnějšího prostředí
dědičnost účinek genů metody studia

108
Geny - základní účinek - modifikující účinek - interakce - pleiotropie

109
Expresivita genu - intenzita projevu.

110
Dále vliv prostředí, stavu organismu aj.

111
Nestejná penetrance, tj. frekvence projevu.

112
Modifikující účinek genů
pleiotropní efekt neměřitelný Geny se základním účinkem Geny modifikátory intezifikátory supresory různá exprese genů expresivita

113
Mutace

114
Základní dělení mutací
Podle oblasti genomu, kterou postihuje: A) genomové B) chromozomové = chromozomální aberace C) genové = bodové
 genomové. B) chromozomové = chromozomální aberace. C) genové = bodové.")
115
Genomové mutace stav, kdy dojde ke zvýšení nebo snížení počtu celých chromozomů, respektive chromozomálních sad Aneuploidie Euplodie

116
Genomové mutace

117
Genomové mutace

118
Chromozomální aberace
A) balancované B) nebalancované 1) duplikace 2) delece 3) inzerce 4) inverze 5) translokace
 balancované. B) nebalancované. 1) duplikace. 2) delece. 3) inzerce. 4) inverze. 5) translokace.")
119
Chromozomální aberace
6) izochromozom 7) fragmentace 8) ring chromozom 9) marker chromozom
 izochromozom. 7) fragmentace. 8) ring chromozom. 9) marker chromozom.")
120
Chromozomální aberace

121
Genové mutace Na úrovni vlákna DNA 1) adice 2) delece 3) substituce
a) tranzice b) transverze
 tranzice. b) transverze.")
122
Mutace kódující sekvence
A) mutace neměnící smysl (samesense, silent mutation) B) mutace měnící smysl (missense mutation) C) nesmyslné mutace (nonsense mutation)
 mutace neměnící smysl (samesense, silent mutation) B) mutace měnící smysl (missense mutation) C) nesmyslné mutace (nonsense mutation)")
123
Dělení podle způsobu vzniku
1) spontánní 2) indukované – mutageny Dělení mutagenů: A) fyzikální B) chemické C) biologické
 spontánní. 2) indukované – mutageny. Dělení mutagenů: A) fyzikální. B) chemické. C) biologické.")
124
Další typy dělení Podle typu zasažené buňky: Somatické Gametické
Podle životaschopnosti mutanta: Vitální Podmíněně letální Letální

125
Další typy dělení Podle konečného účinku na organismus:
Pozitivní – užitečné Neutrální Negativní - škodlivé

126
Reverzibilita mutace Obnovení původního genotypu 2 způsoby:
Pomocí zpětné mutace Pomocí supresorové mutace

127
Mechanismy opravy mutací
1) fotoreaktivace 2) excizní oprava 3) oprava chybného párování metylací 4) SOS odpověď 5) postreplikační oprava
 fotoreaktivace. 2) excizní oprava. 3) oprava chybného párování metylací. 4) SOS odpověď. 5) postreplikační oprava.")
128
Děkuji za pozornost.

Podobné prezentace