Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
eukaryogeneze

2
Vznik eukaryot Canfieldův oceán - anoxický a sulfidický Vznik života
Eukaryotické lipidy Stopy sterolů Nárůst O2 v atmosféře Nárůst O2 v oceánech Bilateria Vznik eukaryot

3
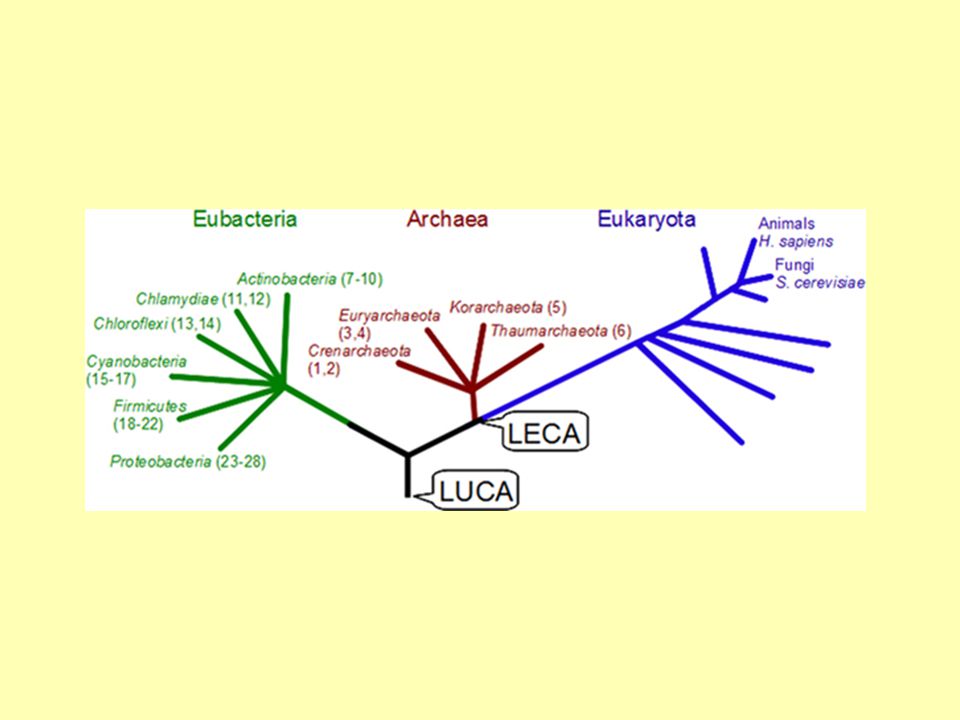
LUCA FECA LECA

4
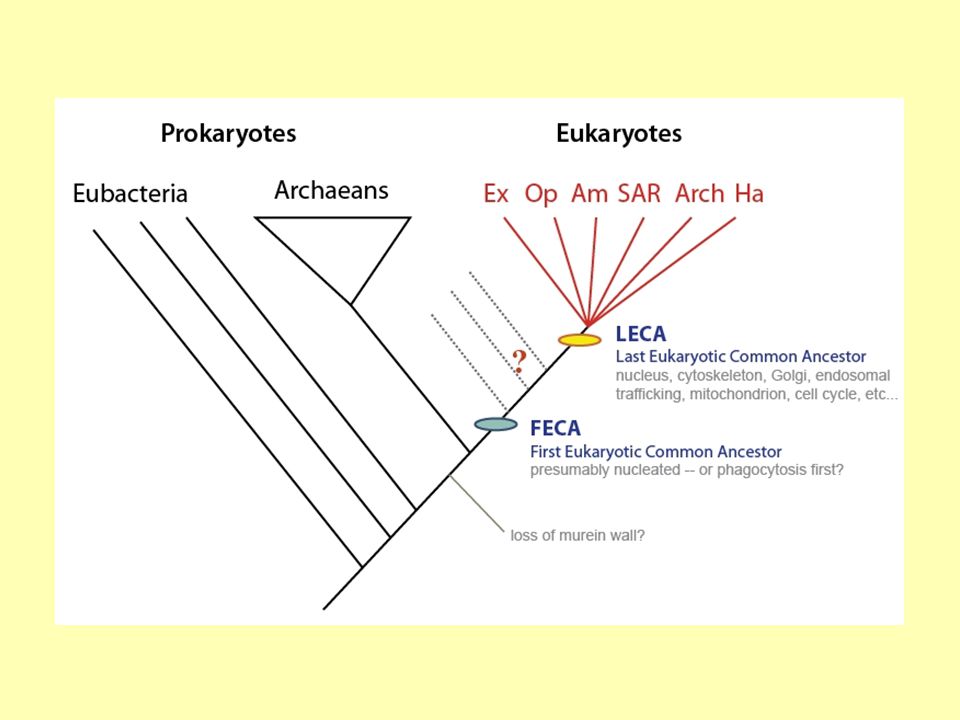
Co je LUCA již víme. FECA je First Eukaryotic Common Ancestor
Co je LUCA již víme. FECA je First Eukaryotic Common Ancestor. LECA je Last Eukaryotic Common Ancestor.

7
Poslední společný předek všech eukaryot LECA
Odhaduje se, že existuje 75 odlišných linií eukaryot. Většina z nich jsou protista. Velikost genomu kolísá od 8.2 megabází (Mb) u Babesia bovis do 112,000–220,050 Mb u Prorocentrum micans (flagellata, patří do protist). To naznačuje, že genom eukaryot prodělal během evoluce značné změny. Poslední společný předek všech eukaryot LECA byl zřejmě fagotrofní protist s jádrem, min. jednou centriolou a ciliem (řasinka, brva, u čl. je např. na epitelu dýchacích cest, ve vejcovodu), fakultativně s aerobní mitochondrií, sexuálním rozmnožováním (meióza, splynutí gamet), cystou, buněčnou stěnou z chitinu a/nebo celulózy a možná s peroxizómy. Pozdější endosymbióza vedla k výskytu plastidů v některých liniích. Viz dále.
 u Babesia bovis do 112,000–220,050 Mb u Prorocentrum micans (flagellata, patří do protist). To naznačuje, že genom eukaryot prodělal během evoluce značné změny. Poslední společný předek všech eukaryot LECA. byl zřejmě fagotrofní protist s jádrem, min. jednou centriolou a ciliem (řasinka, brva, u čl. je např. na epitelu dýchacích cest, ve vejcovodu), fakultativně s aerobní mitochondrií, sexuálním rozmnožováním (meióza, splynutí gamet), cystou, buněčnou stěnou z chitinu a/nebo celulózy a možná s peroxizómy. Pozdější endosymbióza vedla k výskytu plastidů v některých liniích. Viz dále.")
8
Dva typy buněk Endoplazmatické Bičík Jádro retikulum Mitochondrie DNA
Ribozómy Cytoplasmatická membrána Golgiho komplex Bičík Cytoplazmatická membrána a stěna Miller 11th©Brooks-cole

9
Dva typy buněk Endoplazmatické Jádro retikulum Mitochondrie
Cytoplasmatická membrána Golgiho komplex Bičík Miller 11th©Brooks-cole

10
Taxony, které mají buňku Obvyklá velikost Typ jádra DNA
Prokaryotické buňky Eukaryotické buňky Taxony, které mají buňku Bakterie, archea Protista, houby, rostliny, živočichové Obvyklá velikost ca 1 – 10 mm ca 10 – 100 mm Typ jádra Pouze nukleární region, není pravé jádro Jádro oddělené dvojitou nukleární membránou DNA Obv. cirkulární Segmentovaná do lineárních molekul, komplex s histony, v chromozomech Syntéza RNA a proteinů V cytoplazmě RNA v jádře, proteiny v cytoplazmě Ribozomy 50S + 30S 60S + 40S Organely a membránové struktury Velmi málo vnitřních struktur Organely přítomny, buňky silně organizovány vnitřními membránami a cytoskeletem Pohyb buňky Bičík (flagela) z flagelinu Bičík a řasinky (flagela a cilia) obsahující mikrotubuly, lamelipodia a filopodia obsahující aktin (někdy) Mitochondrie Nemá (v pravém smyslu) Obvykle mnoho (některé buňky jedna nebo chybí) Chloroplasty Nemá U řas a rostlin Organizace Obvykle samostatné buňky Jednobuněčné, kolonie, mnohobun. organismy Buněčné dělení Prosté dělení Mitóza (někdy pučení) a meióza
 z flagelinu. Bičík a řasinky (flagela a cilia) obsahující mikrotubuly, lamelipodia a filopodia obsahující aktin (někdy) Mitochondrie. Nemá (v pravém smyslu) Obvykle mnoho (některé buňky jedna nebo chybí) Chloroplasty. Nemá. U řas a rostlin. Organizace. Obvykle samostatné buňky. Jednobuněčné, kolonie, mnohobun. organismy. Buněčné dělení. Prosté dělení. Mitóza (někdy pučení) a meióza.")
11
Tři domény 16S (18S) rRNA
 rRNA")
12
Membrány ARCHEA BACTERIA

13
Eukaryota jsou chiméra
Eubaktérie Archebaktérie Membrány a enzymy energetického metabolismu a mnohé další geny Zejména metabolismus DNA a genová exprese LUCA

14
Eukaryotické novinky Mitochondrie a plastidy (semiautonomní organely)
Cytoskeleton, centrioly, bičíky a s tím spojené molekulární motory Jádro Endomembránový systém (ER, Golgiho komplex, lysozómy) Fagocytóza Organely typu microbodies (např. peroxizómy) Odlišná kontrola buněčného cyklu a mitóza Pohlavní rozmnožování (meióza, splývání gamet a jader) Genom ve formě lineárních chromozómů představující samostatné replikony Znaky na genomu (introny, repetitivní oblasti…) a další…
 Fagocytóza. Organely typu microbodies (např. peroxizómy) Odlišná kontrola buněčného cyklu a mitóza. Pohlavní rozmnožování (meióza, splývání gamet a jader) Genom ve formě lineárních chromozómů představující samostatné replikony. Znaky na genomu (introny, repetitivní oblasti…) a další…")
15
LECA a FECA LECA – Last Eukaryotic Common Ancestor
Mezi FECA a LECA uběhla spousta času a došlo k výrazným proměnám buňky. FECA – First Eukaryotic Common Ancestor

16
Důležitá je zejména endosymbióza s mitochondriemi.

17
Kulík nilský Pluvianus aegyptius

18
Mitochondrie a plastidy
Semiautonomní organely: Dvě (plastid i více ) membrány Vlastní genom a aparát genové exprese Nevznikají de novo Matrix Kristy Jsou pozůstatkem pohlcené bakterie Granum Granum Stroma Stroma Teorie seriální endosymbiózy (SET) Lumen thylakoidu Membrána thylakoidu
 membrány. Vlastní genom a aparát genové exprese. Nevznikají de novo. Matrix. Kristy. Jsou pozůstatkem pohlcené bakterie. Granum. Granum. Stroma. Stroma. Teorie seriální endosymbiózy (SET) Lumen thylakoidu. Membrána thylakoidu.")
19
Z čeho vznikla mitochondrie
Dvojitá membrána (OBĚ) membrány pocházejí z baktérie, mají stejné složení, přítomnost porinů Mitochondrie vznikla jednou. Nejbližšími žijícími příbuznými mitochondrií jsou alfa-proteobaktérie. Která skupina archebakterií to byla není jasné. Rhodospirillum Pelagibacter ubique Rickettsie - značně redukovaní vnitrobuněční paraziti. Ale současné rickettsie dělí od mitochondriálního předka milióny? miliardy? let samostatné evoluce. Předek mohl vypadat jinak.
 membrány pocházejí z baktérie, mají stejné složení, přítomnost porinů. Mitochondrie vznikla jednou. Nejbližšími žijícími příbuznými mitochondrií jsou alfa-proteobaktérie. Která skupina archebakterií to byla není jasné. Rhodospirillum. Pelagibacter ubique. Rickettsie - značně redukovaní vnitrobuněční paraziti. Ale současné rickettsie dělí od mitochondriálního předka milióny miliardy let samostatné evoluce. Předek mohl vypadat jinak.")
20
Mitochondriální genom
Prošel výraznou redukcí Současné a-proteobakterie 834 genů (parazitická Rickettsia), 4000 genů (Rhodospirillum) Mitochondrie (0) genů, většinou 12-20 Reclinomonas americana Chromera velia Anaerobní homology mitochondrie Ztráta genů - nepotřebné geny jsou zmutovány a postupně deletovány. Přenos do jádra -obdoba lateral gene transfer, proces stále probíhá 1 gen za 105 generací umělého systému v kvasince S. cerevisiae. U větších genomů jsou kódující oblasti vyznačeny červeně.
, 4000 genů (Rhodospirillum) Mitochondrie 67-2 (0) genů, většinou Reclinomonas. americana. Chromera. velia. Anaerobní homology. mitochondrie. Ztráta genů - nepotřebné geny jsou zmutovány a postupně deletovány. Přenos do jádra -obdoba lateral gene transfer, proces stále probíhá 1 gen za 105 generací umělého systému v kvasince S. cerevisiae. U větších genomů jsou kódující oblasti vyznačeny červeně.")
21
Genů mitochondriálního původu je velký počet

22
Transformace mitoch. proteomu
Mitochondriální proteom obsahuje několik stovek (Plasmodium) až 3000 (obratlovci) proteinů Proto-mitochondrie ztráta Přesměrování ~ 70% z protomitochondrie zisk Moderní mitochondrie ~ 15% pochází čas proteiny
 až 3000 (obratlovci) proteinů. Proto-mitochondrie. ztráta. Přesměrování. ~ 70% z protomitochondrie. zisk. Moderní mitochondrie. ~ 15% pochází. čas. proteiny.")
23
Transport metabolitů (např. ATP)
Je zprostředkováván rodinou proteinů na vnitřní mitochondriální membráně. Tyto proteiny jsou eukaryotického původu. Pohled z cytosolické strany do transpotního kanálu, ve kterém se právě nachází molekula ADP.

24
Selekční výhoda mitochondrie
Efektivnější metabolismus Rozdíl v energetickém zisku a efektivitě energetického metabolismu je obrovský (2 ATP x 36ATP). To se ale týká dnešní mitochondrie! V okamžiku, kdy hostitel pohltil bakterii nebyly ustanoveny mechanismy transportu látek a hlavně ATP. Mohla už v tuto chvíli přinášet hostiteli energetický užitek?
. To se ale týká dnešní mitochondrie! V okamžiku, kdy hostitel pohltil bakterii nebyly ustanoveny mechanismy transportu látek a hlavně ATP. Mohla už v tuto chvíli přinášet hostiteli energetický užitek")
25
Selekční výhoda mitochondrie
Ochrana před toxickým kyslíkem (Ox-Tox model) Předek mitochondrie snižoval ve svém okolí koncentraci kyslíku, což mohlo být přitažlivé pro anaerobního hostitele. Teprve poté, když hostitel symbionta ovládl, začal mu přinášet výhody ve formě ATP. O2 O2 O2 O2 O2 O2 O2 O2 O2 O2 O2 O2 O2 O2 O2 O2 O2
 Předek mitochondrie snižoval ve svém okolí koncentraci kyslíku, což mohlo být přitažlivé pro anaerobního hostitele. Teprve poté, když hostitel symbionta ovládl, začal mu přinášet výhody ve formě ATP. O2. O2. O2. O2. O2. O2. O2. O2. O2. O2. O2. O2. O2. O2. O2. O2. O2.")
26
Selekční výhoda mitochondrie
Ochrana před toxickým kyslíkem (Ox-Tox model) Předek mitochondrie snižoval ve svém okolí koncentraci kyslíku, což mohlo být přitažlivé pro anaerobního hostitele. Až když hostitel symbionta ovládl, začal mu přinášet výhody ve formě ATP. O2 ATP O2 O2 O2
 Předek mitochondrie snižoval ve svém okolí koncentraci kyslíku, což mohlo být přitažlivé pro anaerobního hostitele. Až když hostitel symbionta ovládl, začal mu přinášet výhody ve formě ATP. O2. ATP. O2. O2. O2.")
27
Selekční výhoda mitochondrie
Žádná nebyla, mitochondrie byl parazit. Mitochondrie byla původně parazitická baktérie, jejíž infekci buňka přežila a baktérii si zotročila.

28
Selekční výhoda mitochondrie
Metabolické symbiózy Mezi předkem mitochondrie a hostitelská buňka docházelo nejprve k oboustranně prospěšné výměně metabolitů – vodíku (vodíková hypotéza), substrátů a produktů fotosyntézy (hypotéza fotosyntetické mitochondrie). Později došlo k pohlcení předka mitochondrie. Výměna metabolitů
, substrátů a produktů fotosyntézy (hypotéza fotosyntetické mitochondrie). Později došlo k pohlcení předka mitochondrie. Výměna metabolitů.")
29
Všechna současná eukaryota mají mitochondrii
Archezoa

30
Různé formy mitochondrie
Klasické mitochondrie Anaerobní mitochondrie loss of genome Hydrogenosomy Nemají oxydativní fosforylaci – dýchací řetězec produkují ATP a koncovým metabolitem je H2 Mitosomy Neprodukují ATP Obsahují proteiny typické pro mitochondrie Většinou zapojeny do biogeneze Fe-S klastrů Někdy funkce neznámá

31
LECA měl mitochondrii LECA FECA

32
Vznik cytoskeletu PROKARYOTA EUKARYOTA FtsZ, TubZ a RepX Tubulin
MukB ? Myosin MreB, ParM, Aktin FtsA,

33
Vznik cytoskeletu PROKARYOTA EUKARYOTA Tubulin Tubulin
Verrucomicrobia (pravděpodobně laterální přenos z eukaryot) Nitrosoarcheum (Archeon z kmene Thaumarcheota) Crenaktin Aktin Některá Crenarcheota, Aigarcheota a Korarcheota
 Nitrosoarcheum (Archeon z kmene Thaumarcheota) Crenaktin Aktin. Některá Crenarcheota, Aigarcheota a Korarcheota.")
34
Vznik bičíku Prokaryotický bičík Eukaryotický bičík Vnější páry
mikrotubulů Dyneinová raménka Středový pár mikrotubulů Radiální paprsky Dyneinová raménka Průřez bičíkem Plazmatická membrána Bičíky Průřez bazálním tělískem Triplety Bazální tělísko Eukaryotický bičík Pohyb bičíku

35
Vznik bičíku - endogenní
Centriola Centriola je homologická s bazálním tělískem bičíku a obě struktury se zapojují do utváření dělícího vřeténka. Mitóza je pro buňku důležitější než bičík, a proto se dá předpokládat, že vznikla dříve. Bičík mohl vzniknout protažením centrioly na druhou stranu. Bičíkové „motory“ dyneiny jsou odvozeny od cytoplasmatických.

36
Vznik bičíku – endosymbiózou - ne
Mixotricha paradoxa Ze spirochét – pohyb pomocí spirochet je znám u některých prvoků (Parabasala). Spirochety mají nějaké tubuly, ty ale nejsou z tubulinu, a nemají ani dynein. Z verrucomicrobií – mají tubuliny a vytvářejí něco jako tubuly, ale nehýbou jimi, zřejmě nemají dynein. Žijí na povrchu některých nálevníků jakoa epixenozómy. Splynutím epixenozómu s buňkou, redukcí jeho genomu a vystřelovacího aparátu a prodloužením tubulinů mohl vzniknout bičík.
. Spirochety mají nějaké tubuly, ty ale nejsou z tubulinu, a nemají ani dynein. Z verrucomicrobií – mají tubuliny a vytvářejí něco jako tubuly, ale nehýbou jimi, zřejmě nemají dynein. Žijí na povrchu některých nálevníků jakoa epixenozómy. Splynutím epixenozómu s buňkou, redukcí jeho genomu a vystřelovacího aparátu a prodloužením tubulinů mohl vzniknout bičík.")
37
Vznik bičíku - endosymbiózou
Mixotricha paradoxa Ze spirochét – pohyb pomocí spirochet je znám u některých prvoků (Parabasala). Spirochety mají nějaké tubuly, ty ale nejsou z tubulinu, a nemají ani dynein. Z verrucomicrobií – mají tubuliny a vytvářejí něco jako tubuly, ale nehýbou jimi, zřejmě nemají dynein. Žijí na povrchu některých nálevníků jako epixenozómy. Splynutím epixenozómu s buňkou, redukcí jeho genomu a vystřelovacího aparátu a prodloužením tubulinů mohl vzniknout bičík.
. Spirochety mají nějaké tubuly, ty ale nejsou z tubulinu, a nemají ani dynein. Z verrucomicrobií – mají tubuliny a vytvářejí něco jako tubuly, ale nehýbou jimi, zřejmě nemají dynein. Žijí na povrchu některých nálevníků jako epixenozómy. Splynutím epixenozómu s buňkou, redukcí jeho genomu a vystřelovacího aparátu a prodloužením tubulinů mohl vzniknout bičík.")
38
LECA FECA LECA měl: mitochondrii bičík a cytoskelet
Bezbičíkatá eukaryota (některé améby, většina hub) nejspíš nemají bičík druhotně (ztráta). FECA
 nejspíš nemají bičík druhotně (ztráta). FECA.")
39
Vznik endomembránového systému
Membránová dědičnost DNA

40
Vznik endomembránového systému
Membránová dědičnost DNA

41
Vznik endomembránového systému

42
Vznik endomembránového systému
ER Jádro

43
Vznik endomembránového systému
ER Jádro

44
Vznik endomembránového systému
Proto-Golgi- fago-lysozóm ER Jádro

45
Vznik endomembránového systému
Endozóm Lysozóm Golgi ER Jádro

46
Vznik endomembránového systému
Endozóm Lysozóm Golgi ER Jádro

47
Vznik endomembránového systému
Endozóm Lysozóm Golgi ER Jádro

48
Vznik endomembránového systému
Eukaryota bez Golgi diplomonády retortamonády Endozóm Lysozóm Golgi oxymonády heterolobosea ER archaméby mikrosporidie Jádro

49
LECA FECA LECA měl: mitochondrii bičík a cytoskelet
endomembránový systém LECA FECA

50
Výhody obaleného jádra
Endozóm Lysozóm Golgi Produkce radikálů v cytoplasmě ER UV a kyslíkové radikály Invaze intronů pravděpodobně z mitochondrie Jádro

51
STRUKTURA GENOMU ER Jádro
Endozóm Lysozóm Golgi STRUKTURA GENOMU ER Změnila se pravděpodobně spolu se vznikem mitózy, meiózy a pohlavního rozmnožování Jádro

52
LECA FECA LECA měl: mitochondrii bičík a cytoskelet
endomembránový systém lineární chromozomy, mitózu, sex, introny bakteriální typ fosfolipidů LECA FECA

53
Endogenně z endoplasmatického retikula
Vznik peroxizómu Endosymbioticky z G+ baktérie Endogenně z endoplasmatického retikula Peroxisom G+ baktérie Peroxisom Eukaryota bez peroxisomů Modifikované peroxisomy Glykosomy Glyoxysomy Diplomonády Retortamonády Mikrosporidie Oxymonády Parabasala Archaméby Trypanosomy Rostliny

54
Endogenně z endoplasmatického retikula
Vznik peroxizómu Endosymbioticky z G+ baktérie Endogenně z endoplasmatického retikula Peroxisom G+ baktérie Peroxisom Eukaryota bez peroxisomů Modifikované peroxisomy Glykosomy Glyoxysomy Diplomonády Retortamonády Mikrosporidie Oxymonády Parabasala Archaméby Trypanosomy Rostliny

55
Peroxisomy Dříve označované mikrotělíska, buněčné organely ovoidního tvaru, ohraničeny jednoduchou membránou (viz elektronogram). Velikost kolem 0,5 µm (0,1-1,2 µm), přibl. stejně jako primární lyzosomy (Ly viz elektronogram). Homogenní matrix peroxisomů obsahuje enzymy oxidující substrát a redukující O2 a H2O2. Dále obs. katalázu, která rozkládá peroxid vodíku na vodu a kyslík. U některých živ. druhů (ne u člověka) je v matrix krystalický nukleotid. Peroxisomy nejsou konstantní organelou všech buněk. Vyskytují se především v hepatocytech a v buňkách ledvinných tubulů. Funkcí je především ochrana buňky před účinkem peroxidu vodíku. Peroxisomy se tvoří za účasti endoplazmatického retikula, přičemž syntéza proteinů probíhá na ribosomech. Peroxisomy se mohou dělit.
. Velikost kolem 0,5 µm (0,1-1,2 µm), přibl. stejně jako primární lyzosomy (Ly viz elektronogram). Homogenní matrix peroxisomů obsahuje enzymy oxidující substrát a redukující O2 a H2O2. Dále obs. katalázu, která rozkládá peroxid vodíku na vodu a kyslík. U některých živ. druhů (ne u člověka) je v matrix krystalický nukleotid. Peroxisomy nejsou konstantní organelou všech buněk. Vyskytují se především v hepatocytech a v buňkách ledvinných tubulů. Funkcí je především ochrana buňky před účinkem peroxidu vodíku. Peroxisomy se tvoří za účasti endoplazmatického retikula, přičemž syntéza proteinů probíhá na ribosomech. Peroxisomy se mohou dělit.")
56
Eukaryotní buňka 1. jadérko, 2. jádro, 3. ribozom, 4. váček, 5. hrubé endoplaztmatické retikulum, 6. golgiho aparát, 7. cytoskelet, 8. hladké endoplazmtické retikulum, 9. mitochondrie, 10. vacuola, 11. cytozol, 12. lysozom, 13. centriol.

57
Lysozomy Sférické membránové organely vyskytující se u mnoha eukaryontních organismů. Obsahují přes 50 typů enzymů (kyselé hydrolázy), rozklad extra- i intracelulárního materiálu – sacharidů, proteinů, lipidů, nukleových kyselin. Autofagii využívá buňka k recyklaci vlastních opotřebovaných struktur. Vznikají vět. oddělením se z hladkého endoplazmatického retikula nebo Golgiho aparátu. Lysozomy: primární – váčky s enzymy, bez materiálu; sekundární – podstatně větší, vznikají splynutím primárního lysozomu s fagozomem; terciární – obsahují zbytky materiálu, který už nejde rozložit, např. lipofuscin.
, rozklad extra- i intracelulárního materiálu – sacharidů, proteinů, lipidů, nukleových kyselin. Autofagii využívá buňka k recyklaci vlastních opotřebovaných struktur. Vznikají vět. oddělením se z hladkého endoplazmatického retikula nebo Golgiho aparátu. Lysozomy: primární – váčky s enzymy, bez materiálu; sekundární – podstatně větší, vznikají splynutím primárního lysozomu s fagozomem; terciární – obsahují zbytky materiálu, který už nejde rozložit, např. lipofuscin.")
58
Onemocnění na podkladě defektu lyzosomů:
Vrozená – genetické poruchy jejich enzymů či regulačních proteinů, dochází k postupnému hromadění materiálu, zejm. v CNS a makrofázích. Asi 40 chorob, frekvence 1 / 100 000 porodů, většinou neléčitelné. Fabryho choroba– tato je léčitelná podáváním chybějící α-galaktosidázy. Niemann-Pickova choroba. Tay-Sachsova choroba. Gaucherova choroba. Pompeho choroba. Získaná – většinou jen relativní poruchy, z nadbytku přijímaného materiálu. Při dně jsou membrány lysozomů rozrušovány krystalky kyseliny močové, to vede k uvolnění enzymů, které se projeví kloubním zánětem.

59
Zde se odehrála většina
LECA měl: mitochondrii bičík a cytoskelet endomembránový systém lineární chromozomy, mitózu, sex, introny bakteriální typ fosfolipidů LECA Zde se odehrála většina eukaryogeneze FECA nevíme, zda měl peroxisom

60
JAK VYPADAL FECA? LECA FECA

61
Některá prokaryota jakoby měla k eukaryotům blíže
Eubakteriální skupina PVC (Planctomycetes, Verrucomicrobia a Chlamydiae) U některých zástupců se vyskytují následující znaky typické pro eukaryota: Kompartmentalizace buňky Proteinový plášť na membráně a cosi podobné endocytóze Kondenzovaný chromatin Množí se pučením Syntetizují steroly Obsahují lipidy podobné archeím Tubulin Gemmata obscuriglobus
 U některých zástupců se vyskytují následující znaky typické pro eukaryota: Kompartmentalizace buňky. Proteinový plášť na membráně a cosi podobné endocytóze. Kondenzovaný chromatin. Množí se pučením. Syntetizují steroly. Obsahují lipidy podobné archeím. Tubulin. Gemmata obscuriglobus.")
62
Některá prokaryota jakoby měla k eukaryotům blíže
Archeální superskupina TACK (Thaumarchaeota, Crenarchaeota, Korarchaeota, Aigarchaeota) U některých zástupců se vyskytují následující znaky typické pro eukaryota: Crenactin (homolog actinu) Tubulin Kompatmentalizace Ubiquitin Velká podobnost enzymů genové exprese
 U některých zástupců se vyskytují následující znaky typické pro eukaryota: Crenactin (homolog actinu) Tubulin. Kompatmentalizace. Ubiquitin. Velká podobnost enzymů genové exprese.")
63
Fagotrofický vznik eukaryot (Neomura)
ARCHAEA EUKARYOTA Hypertermofilie – nová buněčná stěna jiné lipidy Flexibilní povrch – vznik endom. systému, cytoskeletu a fagocytózy NEOMURA Mureinová stěna nahrazena N-glykoproteiny EUBACTERIA Steroly H1 histon Proteazóm Gram-pozitivní Ztráta vnější membrány LUCA Gram-negativní

64
Fagotrofický vznik eukaryot (Neomura)
ARCHAEA EUKARYOTA Další eukaryotické inovace - mitóza, introny, sex atd. Je možné, že invaze nových virů a snaha uniknout před antibiotiky vedla k vzniku dost odlišného aparátu replikace, transkripce a translace Hypertermofílie – nová buněčná stěna jiné lipidy Mitochondrie Flexibilní povrch – vznik endom. systému, cytoskeletu a fagocytózy NEOMURA Mureinová stěna nahrazena N-glykoproteiny EUBACTERIA α-proteobakterie Steroly H1 histon Proteazóm Gram-pozitivní Ztráta vnější membrány LUCA Gram-negativní

65
Fagotrofický vznik eukaryot
Pozdní „příchod“ mitochondrie EUKARYOTA EUBACTERIA ARCHEA Mitochondrii pozřel až téměř dokonalý eukaryot schopný fagocytózy

66
Fagotrofický vznik eukaryot (Neomura)
Je v souladu s chimerickým původem genomu eukaryot: „archeální“ geny euk. zdědila od společného předka archeí a eukaryot, „eubakteriální“ geny od mitochondriálního endosymbionta. Vysvětluje mnoho biochemických vlastností eukaryot (přítomnost sterolů, chitinu, H1 histonu, proteazómu atd.), jako zděděné vlastnosti po Gram pozitivních baktériích (skupina Actinobacteria). Nepředpokládá málo pravděpodobné události (splývání prokaryotických buněk), jako hypotéza které budou následovat.
, jako zděděné vlastnosti po Gram pozitivních baktériích (skupina Actinobacteria). Nepředpokládá málo pravděpodobné události (splývání prokaryotických buněk), jako hypotéza které budou následovat.")
67
Fagotrofický vznik eukaryot (Neomura)
ALE Vznik fagocytózy u prokaryotické buňky bez mitochondrie je problematický. Fagocytóza je energeticky náročný proces a mohl navíc vzniknout jen u buněk s velkým objemem. Předek eukaryot vytvářel většinu svého ATP z protonového gradientu na plazmatické membráně. Je proto těžké si představit, že by se mohl výrazně zvětšit a ještě na plazmatické membráně provozovat fagocytózu. H2 H2 H2 H2 ADP H2 ATP ADP H2 H2 ATP ADP ADP H2 H2 H2 ATP ATP H2 H2

68
Vodíková hypotéza Ke vzniku eukaryot došlo v anaerobním prostředí
H2 produkující alfa-proteobaktérie Metanogenní archebaktérie H2 (Schopná aerobního i anaerobního metabolismu) CH4 H2 Ke vzniku eukaryot došlo v anaerobním prostředí – první eukaryot byl fakultativní anaerob
 CH4. H2. Ke vzniku eukaryot došlo v anaerobním prostředí. – první eukaryot byl fakultativní anaerob.")
69
Vodíková hypotéza Brzký „příchod“ mitochondrie
EUKARYOTA EUBACTERIA ARCHEA Mitochondrie byla spouštěčem eukaryogeneze

70
Vodíková hypotéza PRO Navrhuje selekční tlak pro vznik mitochondrie (H2) Obdobnou endosymbiózu lze najít v přírodě (hydrogenosomy nálevníků a metanogeni).
.")
71
Vodíková hypotéza PROTI
Nahrazení archeálních lipidů v membráně eukaryotickými je obtížně představitelné.

72
Vznik eukaryot Došlo k němu jen jednou.
LECA byl už pokročilý eukaryot, neznáme mezistupně eukaryogeneze. Genom eukaryot je chiméra eubakteriálních a archeálních genů, membrány jsou eubakteriální. Eukaryogeneze mohla probíhat pozvolna a hnací silou mohl být vynález fagotrofie nebo pohlavního rozmnožování. Nebo mohla být eukaryogeneze odstartovaná náhlou a velmi nepravděpodobnou událostí – fúzí archeona a eubakterie, při které vznikla mitochondrie. První eukaryot mohl být aerob ale i fakultativní anaerob.

73
Podle tradiční představy se bakterie postupně přeměnily na primitivní eukaryotickou buňku se všemi znaky současných eukaryot, ale bez mitochondrie. Tuto evoluci zřejmě spustila katastrofická ztráta buněčné stěny.

74
Domény: - Bacteria - Archea (bez jádra) Eucaryota (s jádrem), součástí domény jsou eukaryota s jádrem, ale bez mitochondrie, kterou buď ztratily, nebo mají jen jejich pozůstatky např. mitozomy. Ty mají někt. mt funkce, ale ne aerobní dýchání. Dříve považovaná za skupinu Arechozoa.
, součástí domény jsou eukaryota s jádrem, ale bez mitochondrie, kterou buď ztratily, nebo mají jen jejich pozůstatky např. mitozomy. Ty mají někt. mt funkce, ale ne aerobní dýchání. Dříve považovaná za skupinu Arechozoa.")
75
Přes 1000 druhů primitivních eukaryot nemá mitochondrie
Přes 1000 druhů primitivních eukaryot nemá mitochondrie. Některé ji měly a ztratily ji, jiné ji nikdy neměly, protože se od hlavní linie eukaryot oddělily dříve, než je eukaryota získala. ????????????????????????????

Podobné prezentace
.>")









>")








