Stáhnout prezentaci
1
2. ALLOSTERIE Aspartáttranskarbamoylasa jako příklad allosterického enzymu. Allosterické enzymy se neřídí kinetikou Michaelise a Mentenové. Modely allosterie: Symetrický model ( Jacques Monod, Jeffries Wyman a Jean-Pierre Changeux – podle autorů zkracovaný jako MWC model). Angl. concerted (přel. současný) Sekvenční model (Daniel Koshland). Conceptual insights – Stryer, Chapter 10. Hemoglobin je příkladem neenzymového allosterického proteinu
. Angl. concerted (přel. současný) Sekvenční model (Daniel Koshland). Conceptual insights – Stryer, Chapter 10. Hemoglobin je příkladem neenzymového allosterického proteinu.")
2
Enzym Aspartáttranskarbamoylasa (EC 2. 1. 3
Enzym Aspartáttranskarbamoylasa (EC ) je allostericky inhibován konečným produktem metabolické cesty cytidintrifosfátem (CTP). Inhibice zpětnou vazbou. ATCasa katalyzuje kritický stupeň kondenzace Asp s karbamoylfosfátem při syntéze pyrimidinových nukleotidů.
 je allostericky inhibován konečným produktem metabolické cesty cytidintrifosfátem (CTP). Inhibice zpětnou vazbou. ATCasa katalyzuje kritický stupeň kondenzace Asp s karbamoylfosfátem při syntéze pyrimidinových nukleotidů.")
3
Cytidintrifosfát (CTP) inhibuje ATCasu
Cytidintrifosfát (CTP) inhibuje ATCasu. Podobnost mezi substrátem a produktem není žádná.
 inhibuje ATCasu. Podobnost mezi substrátem a produktem není žádná.")
4
Od slide 6 po slide 13 je prezentován způsob objasnění struktury ATCasy. Není podstané pro pochopení principu allosterie !! Modifikace cysteinových vedlejších řetězců (ATCasy). Oddělení podjednotek chromatografií na iontoměniči nebo centrifugací.
. Oddělení podjednotek chromatografií na iontoměniči nebo centrifugací..")
5
Centrifugační studie ATCasy.
Centrifugace v gradientu sacharosy. Sedimentační koeficient nativního enzymu je 11,6 S. Dvě podjednotky 2,8 S a 5,8 S. Svedberg (zkratka S či Sv), též Svedbergova jednotka, je odvozená jednotka fyzikální veličiny známé jako sedimentační koeficient. Tato veličina udává čas, za který proběhne sedimentace dané makromolekuly při její ultracentrifugaci. Platí: 1 Svedberg = 1 S = sekundy Jak od sebe oddělíme obě podjednotky a jak lze odstranit p-hydroxymerkuribenzoát ? Větší podjednotka (c) je katalytická, ale není inhibována CTP. Složena ze tří řetězců po 34 kD se označuje c3. Regulační podjednotka (r) váže CTP. Složena ze dvou řetězců po 17 kD je označena r2. Nativní enzym je složen ze dvou katalytických trimerů a tří regulačních dimerů (2 c3 + 3 r2 = c6r6).
, též Svedbergova jednotka, je odvozená jednotka fyzikální veličiny známé jako sedimentační koeficient. Tato veličina udává čas, za který proběhne sedimentace dané makromolekuly při její ultracentrifugaci. Platí: 1 Svedberg = 1 S = sekundy. Jak od sebe oddělíme obě podjednotky a jak lze odstranit. p-hydroxymerkuribenzoát Větší podjednotka (c) je katalytická, ale není inhibována CTP. Složena ze tří řetězců po 34 kD se označuje c3. Regulační podjednotka (r) váže CTP. Složena ze dvou řetězců po 17 kD je označena r2. Nativní enzym je složen ze dvou katalytických trimerů a tří regulačních dimerů (2 c3 + 3 r2 = c6r6).")
6
Ultracentrifugační studie ATCasy
Ultracentrifugační studie ATCasy. A) Sedimentační rychlost nativní ATCasy. B) Enyzm po působení p-hydroxymerkuribenzoanu. Enzym disocioval na regulační (r) a katalytické podjednotky (c).
 Sedimentační rychlost nativní ATCasy. B) Enyzm po působení p-hydroxymerkuribenzoanu. Enzym disocioval na regulační (r) a katalytické podjednotky (c).")
7
Kvarterní struktura ATCasy. Vpravo je zjednodušený pohled
Kvarterní struktura ATCasy. Vpravo je zjednodušený pohled. Katalytické trimery jsou dva (druhý je zakrytý). Regulační dimery jsou tři. 2 c3 + 3 r2 = c6r6
. Regulační dimery jsou tři. 2 c3 + 3 r2 = c6r6.")
8
Strukturní podobnost mezi reakčním produktem ATCasy N-fosfonokarbamoylaspartátem a PALA (bisubstrátový analog reakčního meziproduktu). Krystalizace ACTasy za přítomnosti PALA - lokalizace aktivního místa.

9
Aktivní místo ATCasy-trimer
Aktivní místo ATCasy-trimer. V aktivním místě, které obsahuje hlavně vedlejší skupiny z jedné podjednotky, ale také část druhé podjednotky (zeleně), je vázán PALA.
, je vázán PALA.")
10
ATCasa existuje ve dvou konformacích: kompaktní , relativně inaktivní označovaná jako T (tense) a uvolněná (relaxed) označovaná R. PALA stabilizuje R stav. ATCasa splňuje kritéria symetrického modelu.

11
Cytidintrifosfát (CTP) stabilizuje T stav
Cytidintrifosfát (CTP) stabilizuje T stav. CTP se váže na regulační podjednotky ATCasy.
 stabilizuje T stav. CTP se váže na regulační podjednotky ATCasy.")
12
T a R stavy v rovnováze. I v situaci bez přítomnosti substrátu existuje ATCasa v rovnováze obou stavů. Bez substrátu převažuje T nad R stavem faktorem 200.

13
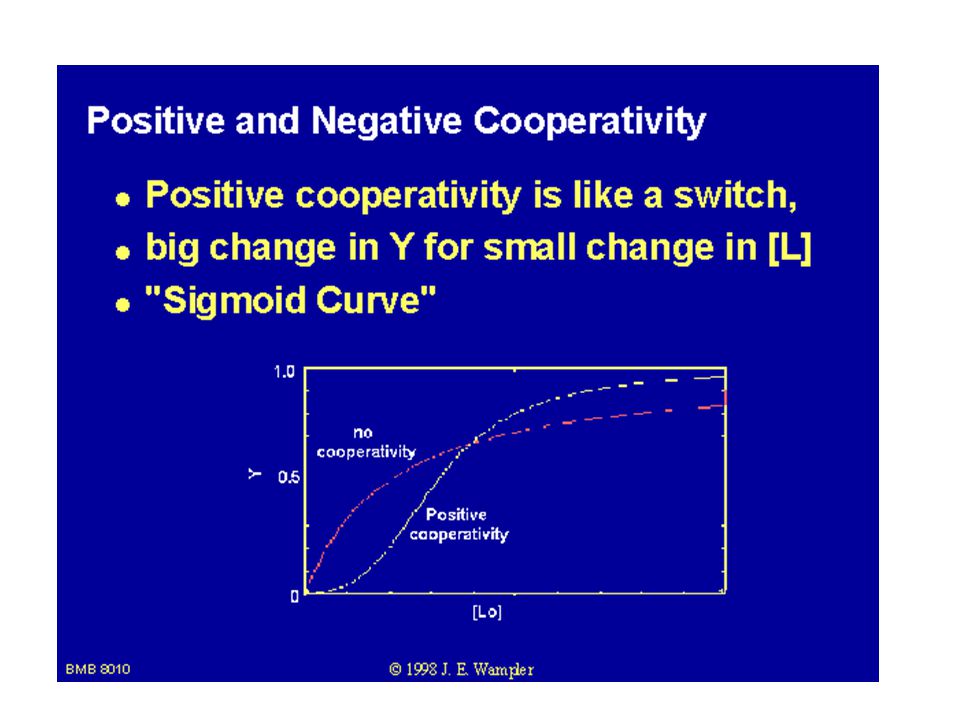
ATCase vykazuje sigmoidní kinetiku
ATCase vykazuje sigmoidní kinetiku. Základní podmínka allosterie proteinu: a) Kvartérní struktura (složen z podjednotek). b) Závislost rychlosti enzymové reakce na koncentraci substrátu je esovitá křivka – sigmoidní tvar.
 Kvartérní struktura (složen z podjednotek). b) Závislost rychlosti enzymové reakce na koncentraci substrátu je esovitá křivka – sigmoidní tvar.")
14
Tvorbu sigmoidní křivky lze interpretovat jako směs dvou Michaelis-Mentenovských enzymů, jeden s vysokou hodnotou KM odpovídající T stavu a druhý s nízkou hodnotou KM odpovídající R stavu. S růstem koncentrací substrátu se rovnováha posunuje od T k R stavu.

15
Vliv CTP na kinetiku ATCasy
Vliv CTP na kinetiku ATCasy. CTP je allosterický inhibitor, stabilizuje T stav. Znesnadňuje vstup substrátu do aktivního místa.

16
Vliv ATP na kinetiku ATCasy. ATP je alosterický aktivátor
Vliv ATP na kinetiku ATCasy. ATP je alosterický aktivátor. Stabilizuje stav R – usnadněný vstup substrátu do aktivního místa.

17
Působení substrátů na allosterické enzymy se označuje jako homotropní efekt
(z řečtiny „stejný“). Působení nesubstrátových molekul na allosterické enzymy jako např. CTP a ATP na ATCasu se označuje jako heterotropní efekt. Zkuste uvažovat proč působí ATP jako aktivátor ATCasy (fyziologická úvaha).
. Působení nesubstrátových molekul na allosterické enzymy jako např. CTP a ATP na ATCasu se označuje jako heterotropní efekt. Zkuste uvažovat proč působí ATP jako aktivátor ATCasy (fyziologická úvaha).")
18
Glukokinasa Glukokinasa (EC ), systematický název: ATP:D-glukosa 6-fosfotransferasa je enzym katalyzující fosforylaci glukosy na glukosa-6-fosfát. Glukokinasa je přítomna v buňkách jater, pankreatu, mozku, střevě lidí a mnoha obratlovců. V každém z těchto orgánů hraje významnou roli při regulaci metabolismu sacharidů. Působí jako glukosový sensor. Reaguje na změny hladin glukosy při hladovění nebo nasycení. Glukokinasa (GK) je hexokinasový izozym homologní s dalšími hexokinasami. GK je kódovaná odlišným genem, na rozdíl od hexokinas, má také odlišné kinetické vlastnosti a s tím související různé odlišné funkce. GK má, na rozdíl od ostatních hexokinas, nižší afinitu ke glukose a její aktivita je lokalizována jen v několika typech buněk.
, systematický název: ATP:D-glukosa 6-fosfotransferasa je enzym katalyzující fosforylaci glukosy na glukosa-6-fosfát. Glukokinasa je přítomna v buňkách jater, pankreatu, mozku, střevě lidí a mnoha obratlovců. V každém z těchto orgánů hraje významnou roli při regulaci metabolismu sacharidů. Působí jako glukosový sensor. Reaguje na změny hladin glukosy při hladovění nebo nasycení. Glukokinasa (GK) je hexokinasový izozym homologní s dalšími hexokinasami. GK je kódovaná odlišným genem, na rozdíl od hexokinas, má také odlišné kinetické vlastnosti. a s tím související různé odlišné funkce. GK má, na rozdíl od ostatních hexokinas, nižší afinitu ke glukose a její aktivita je lokalizována jen v několika typech buněk.")
19
Glukokinasa GK mění svoji strukturu v závislosti na koncentraci glukosy. Je saturována při koncentraci glukosy 8 mM/L. GK není inhibována produktem – glukosa-6-fosfátem !!!! GK vykazuje, ač monomer, kooperativitu – allosterické chování !!! GK je monomer 465 aminokyselin o molekulové hmotnosti 50kD. Na molekule enzymu jsou dvě vazebná místa. Jedno pro substráty (aktivní místo) vážící glukosu a MgATP a druhé, pravděpodobně, pro allosterický aktivátor-inhibitor, které dosud nebyly jednoznačně identifikovány. Ostatní hexokinasy jsou dimery. GK – aktivita může být zvyšována nebo redukována v minutových intervalech glukokinasovým regulačním proteinem(GKRP).
 vážící glukosu a MgATP a druhé, pravděpodobně, pro allosterický aktivátor-inhibitor, které dosud nebyly jednoznačně identifikovány. Ostatní hexokinasy jsou dimery. GK – aktivita může být zvyšována nebo redukována. v minutových intervalech glukokinasovým regulačním proteinem(GKRP).")
20
Glukokinasa s v aktivním místě vázanou glukosou

21
Dva modely allosterie. Symetrický model ( Jacques Monod, Jeffries Wyman a Jean-Pierre Changeux – podle autorů zkracovaný jako MWC model). Sekvenční model (Daniel Koshland).
.")
22
Conceptual insights – Chapter 10.

23
Kvantitativní popis modelu MWC
Kvantitativní popis modelu MWC. Frakční aktivita Yf je část aktivních míst s navázaným substrátem úměrná reakční rychlosti. a je poměr [ S ] / disociační konstantě S s enzymem ve stavu R; L je poměr koncentrací enzymu ve stavu T k enzymu ve stavu R. Vazba regulátorů ATP a CTP mění hodnotu L.

24
Symetrický (MWC) model – kvantitativní formulace.
Mějme enzym s n aktivními místy. Dva stavy enzymu v rovnováze: R – vysoká afinita k substrátu, T - nízká afinita k substrátu. Definujeme c jako poměr afinit obou forem enzymu k substrátu S, měřeno jako disociační konstanty.

25
Frakční aktivita Y, je frakce aktivních míst s navázanými substráty
Frakční aktivita Y, je frakce aktivních míst s navázanými substráty. Je přímo úměrná reakční rychlosti. a je poměr [ S ] ku disociační konstantě S v komplexu s enzymem v R stavu. L je poměr koncentrace enzymu v T stavu ku koncentraci enzymu v R stavu (KR). Vazba regulátorů jako je ATP nebo CTP na ATCasu mění hodnotu L a tím odezvu na koncentraci substrátu.
. Vazba regulátorů jako je ATP nebo CTP na ATCasu mění hodnotu L a tím odezvu na koncentraci substrátu.")
27
Příklad využití kvantitativního modelu s daty ATCasy.
U ATCasy se n = 6. Vynikajícího souhlasu s experimentálními daty se dosahuje při L = 200 a c = 0, 1. Z toho plyne, že za absence vázaného substrátu je posunuta rovnováha na stranu T faktorem 200 ( pouze 1 molekula z 200 je ve stavu R) a afinita R stavu k substrátu je 10x vyšší než ke stavu T. Když se substrát váže do každého aktivního místa, rovnováha se posouvá k R. Např.:Pokud je obsazena polovina aktivních míst substrátem(tři ze šesti) je poměr T/R 1 : 5. Znamená to, že téměř všechny molekuly jsou ve stavu R.
 a afinita R stavu k substrátu je 10x vyšší než ke stavu T. Když se substrát váže do každého aktivního místa, rovnováha se posouvá k R. Např.:Pokud je obsazena polovina aktivních míst substrátem(tři ze šesti) je poměr T/R 1 : 5. Znamená to, že téměř všechny molekuly jsou ve stavu R.")
28
Vliv CTP a ATP se řeší jednoduše změnou hodnoty L.
Při saturaci CTP se hodnota L zvyšuje na 1 250. To znamená, že je třeba daleko více substrátu k posunu rovnováhy do stavu R. Při saturaci ATP klesá hodnota L na 70.

29
SEKVENČNÍ MODEL Daniel KOSHLAND – postupná vazba ligandů (substrátů) na podjednotky proteinu vede k postupnému vzájemnému ovlivňování jednotlivých podjednotek. Možno vysvětlit i negativní kooperaci. U symetrického modelu (MWC) přechází celý enzym z jednoho stavu do druhého (T a R).
 na podjednotky proteinu vede k postupnému vzájemnému ovlivňování jednotlivých podjednotek. Možno vysvětlit i negativní kooperaci. U symetrického modelu (MWC) přechází celý enzym z jednoho stavu do druhého. (T a R).")
30
Jednoduchý sekvenční model tetramerního allosterického enzymu
Jednoduchý sekvenční model tetramerního allosterického enzymu. Postupnou vazbou substrátu na podjednotky se mění konformace okolních ze stavu T na R.

31
Max Perutz (1914 – 2002) a John Kendrew – Nobelova cena za chemii v roce 1962.
Max Perutz pracoval na rentgenové struktuře Hb více než 30 let na Univerzitě v Cambridge (UK). Struktura o rozlišení 2, 8 Å byla získána až v roce 1968. John Kendrew navazoval na M. Perutze, Objasnil strukturu velrybího myoglobinu – rentgenostrukturní analýzou, metodou isomorfní záměny. Těžký atom jako Hg2+ (bohatý na elektrony) se musí vázat na protein, tak, aby nedošlo ke změně struktury. Dochází k podstatné změně v intenzitě odrazu, ale ne v pozici elektronové hustoty.
. Struktura o rozlišení 2, 8 Å byla získána až v roce John Kendrew navazoval na M. Perutze, Objasnil strukturu velrybího myoglobinu – rentgenostrukturní analýzou, metodou isomorfní záměny. Těžký atom jako Hg2+ (bohatý na elektrony) se musí vázat na protein, tak, aby nedošlo. ke změně struktury. Dochází k podstatné změně v intenzitě odrazu, ale ne v pozici elektronové hustoty.")
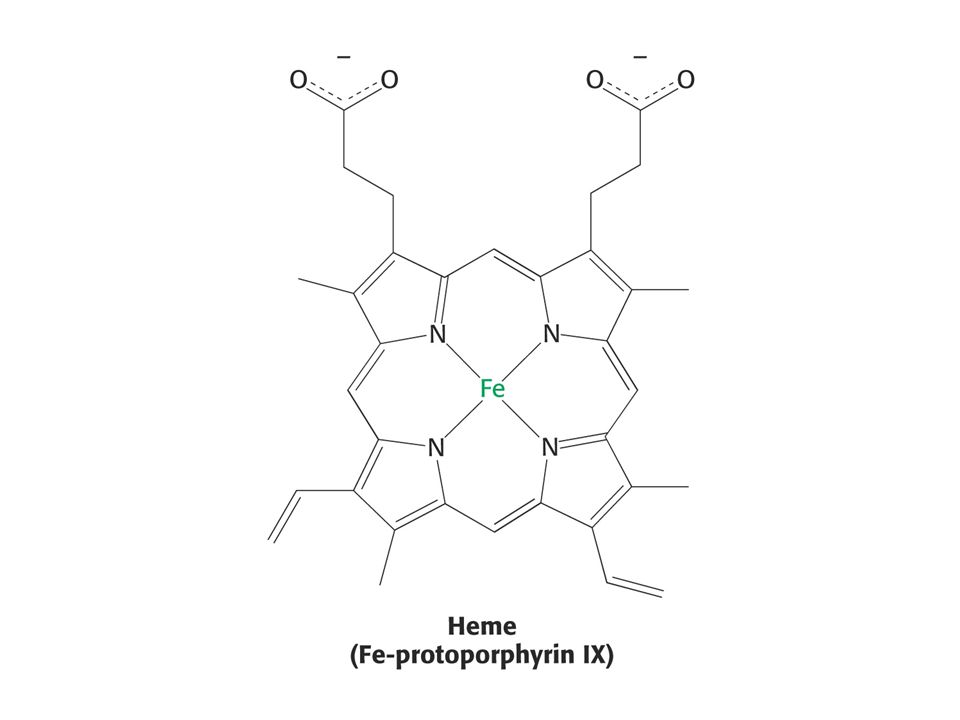
32
Kvartérní struktura hemoglobinu – dva řetězce a a dva b tvoří pár ab dimerů – heterotetramer a2b2.

33
Kooperativní zvýšení transportu kyslíku u hemoglobinu
Kooperativní zvýšení transportu kyslíku u hemoglobinu. Kooperativita mezi vazebnými místy pro kyslík vede k zásobení tkání větším množstvím kyslíku (1, 7x) než by mohl nekooperativní protein. (pO2 je parciální tlak kyslíku).
 než by mohl nekooperativní protein. (pO2 je parciální tlak kyslíku).")
34
Jednotky tlaku 1 torr = 1 mm Hg ≈ 133,322 Pa
Pascal (značka Pa) je základní jednotka tlaku. Udává, jak velká síla (v Newtonech) působí na jednotkovou plochu (1 m2), tzn. je ekvivalentní N/m². 1 hPa = 100 Pa = 1 mb (milibar) 1 Pa = 1 N/m2 1 hPa = 100Pa 1 kPa = 1000 Pa 760 torrů ≈ 101,325 hPa 1 atm = Pa = 101,325 kPa = 0, MPa = 760 torr Parciální tlak plynu ve směsi je tlak, který by tento plyn vykazoval, pokud by byl v celém objemu sám.
 je základní jednotka tlaku. Udává, jak velká síla (v Newtonech) působí na jednotkovou plochu (1 m2), tzn. je ekvivalentní N/m². 1 hPa = 100 Pa = 1 mb (milibar) 1 Pa = 1 N/m2 1 hPa = 100Pa 1 kPa = 1000 Pa. 760 torrů ≈ 101,325 hPa. 1 atm = Pa = 101,325 kPa = 0, MPa = 760 torr. Parciální tlak plynu ve směsi je tlak, který by tento plyn vykazoval, pokud by byl v celém objemu sám.")
35
Krevní oběh – srdce a plíce
Krevní oběh – srdce a plíce. Formy Hb: HbO2 = oxyhemoglobin, deoxyhemoglobin, Mb = svalový myoglobin.

36
Karbonátanhydratasa EC 4.2.1.1 (metaloenzym – Zn2+) Zkratka CA.
 Zkratka CA.")
37
Aktivní místo živočišné a-CA (Zn2+ je vázán na tři imidazolové zbytky His)
")
38
Reakce katalyzované CA.
CO2 + H2O HCO H+ (za situace vysoké koncentrace CO2 v tkáních) Velmi rychlá reakce – 104 až 106 obratů za sekundu (číslo přeměny). Limitováno jen difůzí !!! Zpětná reakce: HCO H+ H2CO3 CO2 + H2O (v plících a nefronech ledvin za stavu nízké koncentrace CO2 , v rostlinách).
 Velmi rychlá reakce – 104 až 106 obratů za sekundu (číslo přeměny). Limitováno jen difůzí !!! Zpětná reakce: HCO3- + H+ H2CO3 CO2 + H2O (v plících a nefronech ledvin za stavu nízké koncentrace CO2 , v rostlinách).")
39
Funkce CA Primární funkcí CA u živočichů je udržování rovnováhy mezi CO2 a hydrgenuhličitanem za účelem udržování acidobazické rovnováhy v krvi a tkáních. Další funkcí je role CA při transportu CO2 z tkání. Existuje 14 různých izoforem CA u savců. Rostliny obsahují odlišnou formu nazvanou β-karbonátanhydratasa, která je z hlediska evoluce odlišný enzym, ale podílí se na stejné reakci a využívá atom zinku v aktivním místě. U rostlin je úkolem CA zvyšovat koncentraci CO2 v chloroplastech za účelem zvýšení rychlosti karboxylace katalyzované RuBisCem. Reakce fotosyntézy – RuBisCo využívá pouze CO2 a ne uhličitou kyselinu nebo hydrogenuhličitan.

40
Mechanismus působení CA
Prosthetická skupina Zn 2+ v aktivním místě je koordinována třemi His. Čtvrtá koordinační pozice je obsazena molekulou vody. Dochází k polarizaci vazby vodík – kyslík. Kyslík má negativní náboj a tím je vazba s vodíkem zeslabena. Čtyři His kolem vody jako substrátu váží proton, což vede k k uvolnění hydroxidové skupiny vázané na Zn. Příklad acidobazické katalýzy. Aktivní místo také obsahuje specifickou kapsu pro CO2 v těsné blízkosti hydroxidové skupiny. Na elektrony bohatý OH – se váže na oxid uhličity za tvorby hydrogenuhličitanu.

42
Pozice Fe2+ v deoxyhemoglobinu.

43
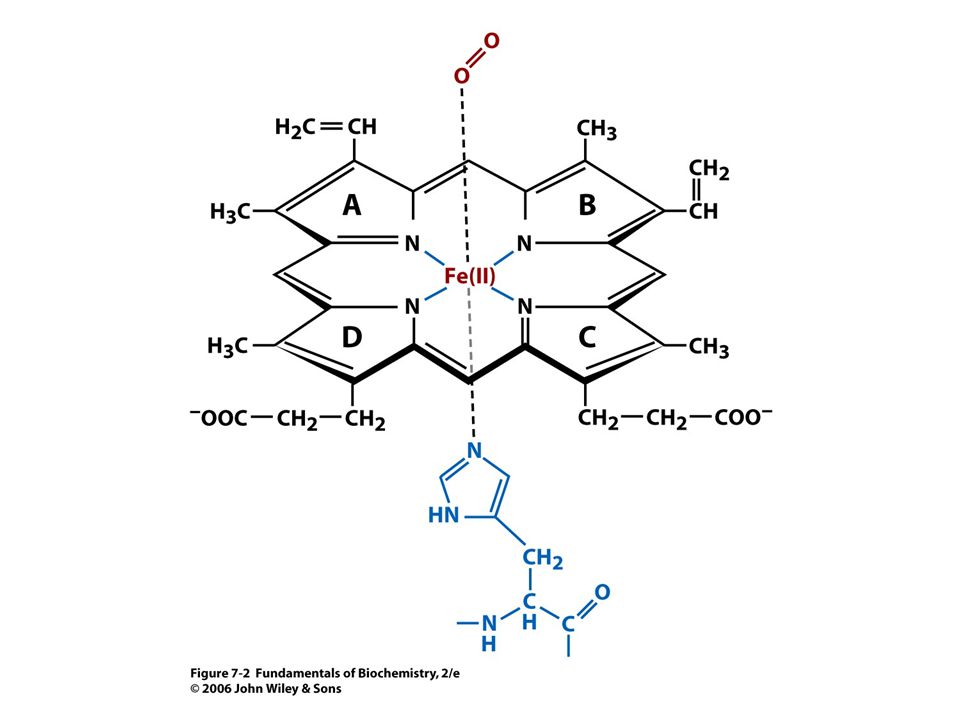
Vazba kyslíku vede ke strukturním změnám
Vazba kyslíku vede ke strukturním změnám. Atom Fe je vtažen po vazbě kyslíku do roviny hemu. S ním je vtažen i proximální His.

44
Konformační změny v hemoglobinu
Konformační změny v hemoglobinu. Pohyb Fe2+ po vazbě kyslíku vtáhne do kruhu proximální His, který jako součást a helixu ovlivní svým C koncem vztah mezi ab páry vedoucí k dalším strukturním změnám. Porovnáni se strukturou deoxy (šedá).
.")
45
Přechod ze stavu T do stavu R u hemoglobinu
Přechod ze stavu T do stavu R u hemoglobinu. Po vazbě kyslíku na pár a1 b1 se a1 b1 pár pootočí vůči druhému a2 b2 páru o 15o.

46
Posun mezi řetězci b2 a a1 hemoglobinu po vazbě kyslíku
Posun mezi řetězci b2 a a1 hemoglobinu po vazbě kyslíku. Stavy R vázán kyslík a T bez kyslíku.

47
Symetrický model allosterie
Symetrický model allosterie. V jedné kvarterní struktuře nemohou existovat vedle sebe formy T a R.

48
Sekvenční model allosterie
Sekvenční model allosterie. Ligand po vazbě na jednu podjednotku progresívně vyvolává konformační změny sousedních podjednotek.

49
Síť iontových párů v deoxyhemoglobinu
Síť iontových párů v deoxyhemoglobinu. C-koncové skupiny (Arg 141a a His 146b) v T stavu. Jsou vytvářeny intra a interjednotkové vazby. Energií vznikající vazbou O2 – Fe jsou vazby přerušeny.
 v T stavu. Jsou vytvářeny intra a interjednotkové vazby. Energií vznikající vazbou O2 – Fe jsou vazby přerušeny.")
50
Síť iontových párů v deoxyhemoglobinu
Síť iontových párů v deoxyhemoglobinu. C-koncové skupiny (Arg 141a a His 146b) v T stavu. Jsou vytvářeny intra a interjednotkové vazby. Energií vznikající vazbou O2 – Fe jsou vazby přerušeny.
 v T stavu. Jsou vytvářeny intra a interjednotkové vazby. Energií vznikající vazbou O2 – Fe jsou vazby přerušeny.")
51
Který z modelů je ten, který lze aplikovat na kooperativní vazbu kyslíku na hemoglobin (Hb) ?
Žádný z obou modelů nelze na Hb čistě aplikovat. Vyhovuje spíše kombinace obou modelů. Hb se třemi místy obsazenými kyslíkem splňuje kriteria sekvenčního modelu – nachází se v R stavu. Zbývající čtvrté vazebné místo má afinitu ke kyslíku 20x vyšší než plně deoxygenovaný Hb vážící svůj první kyslík. Kriteria sekvenčního modelu však nejsou splněna, protože Hb s kyslíkem navázaným pouze v jednom místě ze čtyř zůstává v T stavu. Tato struktura váže kyslík 3x pevněji než plně deoxygenovaný Hb – což odpovídá symetrickému (MWC) modelu. Závěr: Oba modely reprezentují idealizované limitní stavy, ke kterým se reálný systém blíží.
 modelu. Závěr: Oba modely reprezentují idealizované limitní stavy, ke kterým se reálný systém blíží.")
52
Describe and differentiate between the concerted model and the sequential model of allosteric regulation. How does hemoglobin do both? CONCERTED – MWC (allosteric) - connected enzyme subunits must exist in the same conformation - equilibrium (absense of substrate (or ligand)) favors the T-tense state (4 squares) vs. R-relaxed state (4 circles) - the binding of substrate to one subunits causes all remaining subunits to assume the R state (i.e.) thus enhancing their affinity for substrate SEQUENTIAL (allosteric) - subunits need not exist in same conformation - molecules of substrate bind via induced-fit protocol - conformational changes are not propagates to other subunits - substrate binding causes increased substrate affinity only Hemoglobin is a combination of both it is concerted: 3 sites occupied by oxygen associated with R state it is sequential: 1 site occupied by oxygen associated with T state
 - connected enzyme subunits must exist in the same conformation. - equilibrium (absense of substrate (or ligand)) favors the T-tense state (4 squares) vs. R-relaxed state (4 circles) - the binding of substrate to one subunits causes all remaining subunits to assume the R state (i.e.) thus enhancing their affinity for substrate. SEQUENTIAL (allosteric) - subunits need not exist in same conformation. - molecules of substrate bind via induced-fit protocol. - conformational changes are not propagates to other subunits. - substrate binding causes increased substrate affinity only. Hemoglobin is a combination of both. it is concerted: 3 sites occupied by oxygen associated with R state. it is sequential: 1 site occupied by oxygen associated with T state.")
55
Hillova rovnice, Hillův koeficient
In biochemistry, the binding of a ligand to a macromolecule is often enhanced if there are already other ligands present on the same macromolecule (this is known as Cooperative binding). The Hill coefficient, named for Archibald Vivian Hill, provides a way to quantify this effect. It describes the fraction of the macromolecule saturated by ligand as a function of the ligand concentration; it is used in determining the degree of cooperativeness of the ligand binding to the enzyme or receptor. It was originally formulated by Archibald Hill in 1910 to describe the sigmoidal O2 binding curve of hemoglobin. A coefficient of 1 indicates completely independent binding, regardless of how many additional ligands are already bound. Numbers greater than one indicate positive cooperativity, while numbers less than one indicate negative cooperativity. The Hill coefficient was originally devised to explain the cooperative binding of oxygen to hemoglobin (a system which has a Hill coefficient of 2.8-3). A.V. Hill (1886 – 1977) was an English physiologis, one of the founders of the diverse disciplines of biophysic and operations research. He shared the 1922 Nobel Prize in Physiolgy and Medicine “for his elucidation of the production of heat and mechanical work in muscles.
. The Hill coefficient, named for Archibald Vivian Hill, provides a way to quantify this effect. It describes the fraction of the macromolecule saturated by ligand as a function of the ligand concentration; it is used in determining the degree of cooperativeness of the ligand binding to the enzyme or receptor. It was originally formulated by Archibald Hill in 1910 to describe the sigmoidal O2 binding curve of hemoglobin. A coefficient of 1 indicates completely independent binding, regardless of how many additional ligands are already bound. Numbers greater than one indicate positive cooperativity, while numbers less than one indicate negative cooperativity. The Hill coefficient was originally devised to explain the cooperative binding of oxygen to hemoglobin (a system which has a Hill coefficient of 2.8-3). A.V. Hill (1886 – 1977) was an English physiologis, one of the founders of the diverse disciplines of biophysic and operations research. He shared the 1922 Nobel Prize in Physiolgy and Medicine for his elucidation of the production of heat and mechanical work in muscles.")
56
Taking the reciprocal of both sides, rearranging, inverting again, and then taking the logarithm on both sides of the equation leads to an alternative formulation of the Hill equation (θ = theta): θ - fraction of occupied sites where the ligand can bind to the active site of the receptor protein. [L] - ligand concentration Kd - Apparent dissociation constant derived from the law of mass action (equilibrium constant for dissociation) KA - ligand concentration producing half occupation (ligand concentration occupying half of the binding sites), that is also the microscopic dissociation constant. n - Hill coefficient, describing cooperativity (and many more, depending on the system, in the case of which the Hill equation is used)
 KA - ligand concentration producing half occupation (ligand concentration occupying half of the binding sites), that is also the microscopic dissociation constant. n - Hill coefficient, describing cooperativity (and many more, depending on the system, in the case of which the Hill equation is used)")
57
Hodnoty n a jejich interpretace
When appropriate, the value of the Hill constant describes the cooperativity of ligand binding in the following way: n > 1 - Positively cooperative reaction: Once one ligand molecule is bound to the enzyme, its affinity for other ligand molecules increases. n < 1 - Negatively cooperative reaction: Once one ligand molecule is bound to the enzyme, its affinity for other ligand molecules decreases. n = 1 - Noncooperative reaction: The affinity of the enzyme for a ligand molecule is not dependent on whether or not other ligand molecules are already bound. The Hill equation (as a relationship between the concentration of a compound adsorbing to binding sites and the fractional occupancy of the binding sites) is equivalent to the Langmuir equation.
 is equivalent to the Langmuir equation.")
58
Typický Hillův graf pro Hb. Y = frakce obsazených akt
Typický Hillův graf pro Hb. Y = frakce obsazených akt. míst; L – koncentrace ligandu (kyslík); kd = disociační konstanta kyslík – Hb; n = 2,8
; kd = disociační konstanta kyslík – Hb; n = 2,8.")
59
Leghemoglobin Leghemoglobin je hemoprotein analogický hemoglobinu u motýlokvětých rostlin fixujících dusík ve rhizobiích (dusík fixující baktérie) na kořenech. Leghemoglobin má také červenou barvu. Jedná se o symbiotický proces. Původně se mělo zato, že apoprotein leghemoglobinu syntetizuje rostlina a hem bakterie. Nejnovější poznatky ukazují, že hem je také syntetizován rostlinou. Leghemoglobin má vysokou afinitu ke kyslíku – asi 10 x vyšší než b-řetězec hemoglobinu (Km = 0, 01 mM).
 na kořenech. Leghemoglobin má také červenou barvu. Jedná se o symbiotický proces. Původně se mělo zato, že apoprotein leghemoglobinu syntetizuje rostlina a hem bakterie. Nejnovější poznatky ukazují, že hem je také syntetizován rostlinou. Leghemoglobin má vysokou afinitu ke kyslíku – asi 10 x vyšší než b-řetězec hemoglobinu. (Km = 0, 01 mM).")
60
Leghemoglobin U rostlin infikovaných Rhizobii by přítomnost kyslíku v kořenových nodulech silně redukovala funkci a aktivitu na kyslík citlivé nitrogenasy – enzymu, který fixuje atmosférický dusík. Leghemoglobin pufruje koncentarci volného kyslíku v cytoplasmě infikovaných buněk a brání vstupu kyslíku na nitrogenasu. . Poslední studie ukazují, že leghemoglobin skladuje jen tolik kyslíku kolik je třeba k podpoře respirace v nodulech na několik sekund. Jeho funkcí je tedy také transport kyslíku do respirujících symbiotických bakteriálních buněk, analogicky jak tak činí hemoglobin transportující kyslík do respirujících tkání.

61
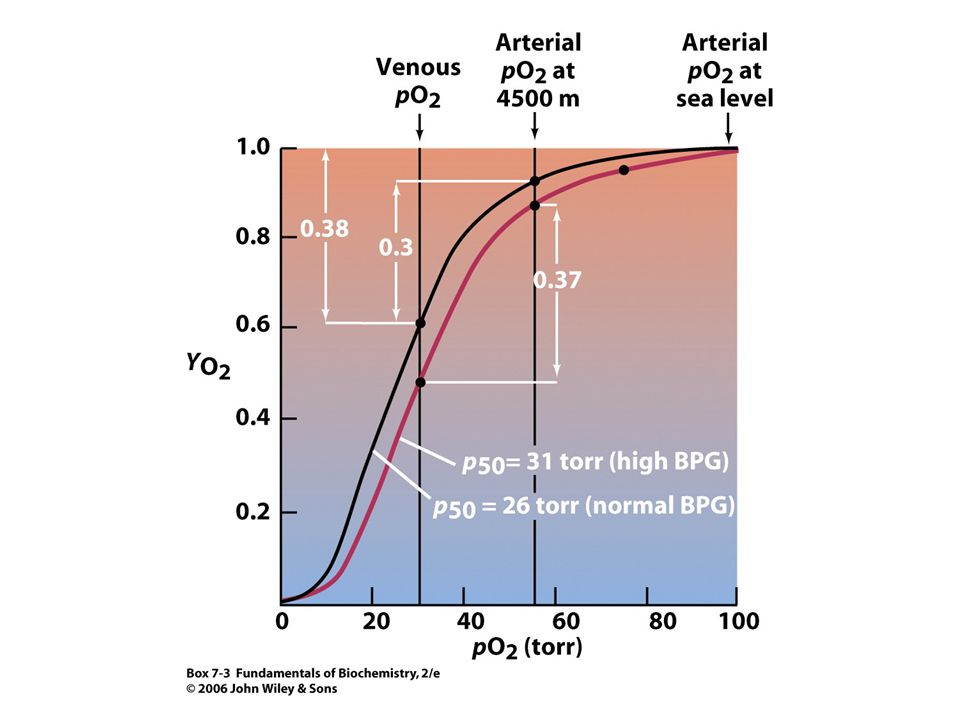
Vliv 2,3-bisfosfoglycerátu (2,3-BPG) na afinitu kyslíku k hemoglobinu.
Bez přítomnosti 2,3-BPG je afinita hemoglobinu ke kyslíku podstatně vyšší a uvolňuje se jen 8 % přenášeného kyslíku do tkání. 2,3-DPG je přítomen v erythrocytech v koncentraci 2 mM (stejná jako konc. hemoglobinu).
.")
62
Způsob vazby 2,3-BPG (náboj-5) na lidský hemoglobin
Způsob vazby 2,3-BPG (náboj-5) na lidský hemoglobin. Váže se do centrální dutiny deoxyhemoglobinu na osm kladně nabitých skupin obou b řetězců.
 na lidský hemoglobin. Váže se do centrální dutiny deoxyhemoglobinu na osm kladně nabitých skupin obou b řetězců.")
63
Proč 2,3-BPG tak silně ovlivňuje afinitu hemoglobinu ke kyslíku ?
2,3-BPG se váže přednostně na deoxyhemoglobin, který stabilizuje. Deoxyhemoglobin je ve stavu T. Přechod do stavu R se děje jen po oddělení 2,3-BPG ! Fyziologický důsledek: Plodový (fetální) hemoglobin se liší od hemoglobinu dospělého člověka. Je to tetramer obsahující dva řetězce a a dva g !! Řetězce g jsou ze 72 % identické s řetězci b u hemoglobinu dospělých. Tato změna však znamená, že řetězce g mají o dva pozitivní náboje méně než b a tím je podstatně snížena afinita fetálního hemoglobinu k 2,3-BPG a tím zvýšena afinita ke kyslíku ve srovnání s hemoglobinem mateřským. To umožňuje bezproblémový přenos kyslíky z matky na plod.
 hemoglobin se liší od hemoglobinu dospělého člověka. Je to tetramer obsahující dva řetězce a a dva g !! Řetězce g jsou ze 72 % identické s řetězci b u hemoglobinu dospělých. Tato změna však znamená, že řetězce g mají o dva pozitivní náboje méně než. b a tím je podstatně snížena afinita fetálního hemoglobinu. k 2,3-BPG a tím zvýšena afinita ke kyslíku ve srovnání s hemoglobinem mateřským. To umožňuje bezproblémový přenos kyslíky z matky na plod.")
64
Inositol – inositolhexafosfát(IP6), fytinová kyselina.
, fytinová kyselina.")
65
Afinita ke kyslíku u fetálních erythrocytů.

66
Bohrův efekt. Rychle metabolizující tkáně, jako svalstvo, mají velké požadavky na kyslík a produkují velké množství H+ a CO2. Jak H+, tak CO2 jsou heterotropní efektory hemoglobinu zvyšující uvolňování kyslíku. Afinita hemoglobinu ke kyslíku klesá při vstupu hemoglobinu do oblastí s nižším pH. Transport z plic (pH 7, 4), parciální tlak kyslíku 100 torrů do aktivního svalu s pH 7, 2 a parciálního tlaku kyslíku 20 torrů vede k uvolnění 77 % celkové přenášené kapacity. Pokud by nedošlo k poklesu pH, tak by se uvolnilo pouze 66 %. Ve tkáních, kde je vyšší koncentrace CO2 klesá afinita hemoglobinu ke kyslíku. Za přítomnosti CO2 o parciálním tlaku 40 torrů se uvolní z hemoglobinu 90 % přenášeného kyslíku. Vliv H+ a CO2 na regulaci vazby kyslíku na hemoglobin se nazývá Bohrův efekt. Podle Christiana Bohra, který jev popsal v roce 1904.
, parciální tlak kyslíku 100 torrů do aktivního svalu s pH 7, 2 a parciálního tlaku kyslíku 20 torrů vede k uvolnění 77 % celkové přenášené kapacity. Pokud by nedošlo k poklesu pH, tak by se uvolnilo pouze 66 %. Ve tkáních, kde je vyšší koncentrace CO2 klesá afinita hemoglobinu ke kyslíku. Za přítomnosti CO2 o parciálním tlaku 40 torrů se uvolní z hemoglobinu 90 % přenášeného kyslíku. Vliv H+ a CO2 na regulaci vazby kyslíku na hemoglobin se nazývá Bohrův efekt. Podle Christiana Bohra, který jev popsal v roce")
67
Vliv pH a koncentrace CO2 na afinitu kyslíku k hemoglobinu.

68
Chemický základ Bohrova efektu.
V deoxyhemoglobinu tvoří tři aminokyseliny dvě iontové vazby stabilizující T stav. Tvorba jedné z vazeb závisí na přítomnosti dalšího protonu na His b 146. To umožňuje vazbu na Asp 94.

69
CO2 stabilizuje deoxyhemoglobin reakcí s terminální aminoskupinou za tvorby karbamátu. Změna náboje vede k pevnější vazbě mezi ab dimery a tím ke stabilizaci T stavu.

70
Hemoglobin s navázanými CO2 a H+ vstupuje do plic, kde uvolní CO2 a H+ a naváže kyslík.
Hemoglobin se však podílí pouze 14 % na celkovém transportu oxidu uhličitého a protonů. Jak CO2, tak H+ jsou transportovány krví jako hydrogenuhličitan (HCO3-), který se tvoří spontánně nebo enzymově karbonátanhydrasou.
, který se tvoří spontánně nebo enzymově karbonátanhydrasou.")
71
Glykovaný hemoglobin (HbA1)
Neenzymová glykace na terminální NH2 skupině (Val) β-řetězce Glykovaná frakce asi 5% celkového množství hemoglobinu (úměrná koncentraci glukózy v krvi) Měřením hladiny HbA1 lze získat informace o průběhu diabetes mellitus (odráží hladinu glukózy za posledních několik týdnů); Cukr CHO + NH2 CH2 Protein Cukr CH N CH2 Protein Cukr CH2 NH CH2 Protein Schiffova base Amadoriho přesmyk Glykovaný protein
 β-řetězce. Glykovaná frakce asi 5% celkového množství hemoglobinu (úměrná koncentraci glukózy v krvi) Měřením hladiny HbA1 lze získat informace o průběhu diabetes mellitus (odráží hladinu glukózy za posledních několik týdnů); Cukr CHO + NH2 CH2 Protein. Cukr CH N CH2 Protein. Cukr CH2 NH CH2 Protein. Schiffova base. Amadoriho přesmyk. Glykovaný protein.")
72
Přepínání hemoglobinu

73
MYOGLOBIN

75
Vazba kyslíku na myoglobin – myoglobin je zpoloviny nasycen kyslíkem při tlaku kyslíku (pO2) 2, 8 torru.
 2, 8 torru.")
76
Fyziologická role myoglobinu.
Rolí myoglobinu je usnadňovat difůzi kyslíku ve svalech při intenzívní svalové činnosti. Rychlost difůze kyslíku z krevních kapilár do tkání je limitována jeho nízkou rozpustností ve vodě (10-4 M v krvi). Myoglobin zvyšuje efekt rozpustnosti kyslíku ve svalových buňkách. Myoglobin jako „sklad“ kyslíku je důležitý u mořských savců jako je např. velryba, jejichž koncentrace myoglobinu ve svalech je 10x vyšší než u suchozemských savců. Myši u kterých byl vyřazen („knocked out“) gen pro myoglobin, mají světlejší svalstvo, ale jinak jsou normální. Myoglobin není pro život suchozemských savců nezbytný. V mozku byl nedávno popsán protein charakteru myoglobinu nazvaný neuroglobin. Funkcí je podpora koncentrace O2 v nervových tkáních, které jsou metabolicky velmi aktivní. Mozek reprezentuje asi 2% hmotnosti lidského těla, ale spotřebovává 20% získaného kyslíku.
. Myoglobin zvyšuje efekt rozpustnosti kyslíku ve svalových buňkách. Myoglobin jako „sklad kyslíku je důležitý u mořských savců jako je např. velryba, jejichž koncentrace myoglobinu ve svalech je 10x vyšší než u suchozemských savců. Myši u kterých byl vyřazen („knocked out ) gen pro myoglobin, mají světlejší svalstvo, ale jinak jsou normální. Myoglobin není pro život suchozemských savců nezbytný. V mozku byl nedávno popsán protein charakteru myoglobinu nazvaný neuroglobin. Funkcí je podpora koncentrace O2 v nervových tkáních, které jsou metabolicky velmi aktivní. Mozek reprezentuje asi 2% hmotnosti lidského těla, ale spotřebovává 20% získaného kyslíku.")
77
Adaptace na výšku. Atmosférický tlak klesá s výškou. Tlak kyslíku ve výšce m je 110 torrů, tj. 70 % hodnoty tlaku na hladině moře. Tlak kyslíku pod 85 torrů je neslučitelný se životem. Adaptace na výšku je spojena se zvýšením množství Hb v erythrocytech a zvýšením počtu erythrocytů. Tento komplexní proces obvykle zahrnuje několik týdnů. Mnohdy stačí pouze jeden den k dosažení dostatečné adaptace. Tento efekt spočívá v rapidním zvýšení množství BPG syntetizovaného v erythrocytech (obvykle ze 4 mM na 8 mM; BPG nemůže procházet membránou erythrocytu). Zvýšená koncentrace BPG způsobuje, že se křivka saturace Hb kyslíkem posunuje oproti křivce na hladině moře do pozice znamenající nižší afinitu !! Na hladině moře je rozdíl pO2 mezi tepennou a žilní krví 70 torrů a Hb uvolňuje 38 % vázaného kyslíku. Když obsah vázaného kyslíku v tepnách klesne na 55 torrů, to je ve výšce m, uvolňuje Hb jen 30 % vázaného kyslíku. Adaptace na výšku (pokles vázaného kyslíku v plících, zvýšení množství kyslíku uvolněného v tkáních) znamená, že Hb přenáší prakticky normální množství kyslíku.
. Zvýšená koncentrace BPG způsobuje, že se křivka saturace Hb kyslíkem posunuje oproti křivce na hladině moře do pozice znamenající nižší afinitu !! Na hladině moře je rozdíl pO2 mezi tepennou a žilní krví 70 torrů a Hb uvolňuje 38 % vázaného kyslíku. Když obsah vázaného kyslíku v tepnách klesne na 55 torrů, to je ve výšce m, uvolňuje Hb jen 30 % vázaného kyslíku. Adaptace na výšku (pokles vázaného kyslíku v plících, zvýšení množství kyslíku uvolněného v tkáních) znamená, že Hb přenáší prakticky normální množství kyslíku.")
78
Koncentrace BPG se zvyšuje v erythrocytech mnohem rychleji než proběhne syntéza Hb (erythrocyty nemají jádro a proto nemohou syntetizovat proteiny). Hladina BPG je mnohem citlivější regulace přenosu kyslíku než změna respirační rychlosti. Hyperventilace, další rychlá odezva na výšku, vede k respirační alkalose. Hyperventilace urychluje vydechování CO2 a způsobuje respirační alkalosu. Alkalosa (pH větší než 7, 6) a acidosa (pH menší než 7, 1) jsou následky nerovnováhy krevních ústojů. Čím je způsobována acidosa a jak se dá potlačit ?? Alkalosa se dá potlačit NaCl nebo KCl (proč?) nebo vdechováním vzduchu obohaceneého CO2.
 a acidosa (pH menší než 7, 1) jsou následky nerovnováhy krevních ústojů. Čím je způsobována acidosa a jak se dá potlačit Alkalosa se dá potlačit NaCl nebo KCl (proč ) nebo vdechováním vzduchu obohaceneého CO2.")
79
Obyvatelé And a Himalájí mají vysokou kapacitu plic, vysokou hladinu Hb a často také větší pravou srdeční komoru což znamená zvýšení objemu čerpané krve. Na rozdíl od adaptace lidí na výšku, většina savců žijících ve vysokých výškách (lamy), má geneticky pozměněný Hb vyznačující se vyšší afinitou ke kyslíku. K adaptaci na výšku vedou dvě cesty: zvýšení i snížení p50 Hb.
, má geneticky pozměněný Hb vyznačující se vyšší afinitou ke kyslíku. K adaptaci na výšku vedou dvě cesty: zvýšení i snížení p50 Hb.")
81
Abnormální hemoglobiny

82
Methemoglobin Také ferrihemoglobin, zkr. metHb, hemoglobin obsahující trojmocné železo (Fe3+ ), není schopen přenášet kyslík. V erythrocytech vzniká za fyziologických podmínek v malém množství přímou oxidací železa kyslíkem; je zde enzymově zpětně redukován na Fe2+ formu. Větší množství metHb vzniká následkem otrav (acetanilid, chlornany, dusitany atd.) nebo vrozených poruch syntézy hemoglobinu. Může se patologicky objevovat v krevní plasmě (methemoglobinemie) nebo v moči (methemoglobinurie). Způsobuje modrání kůže, obecně, sliznic (cyanóza). Talasemie - disproporce tvorby řetězců a a b hemoglobinu. Některé vzácnější formy hereditárních anémií vyskytující se v dospělé populaci v ČR a SR – β−talasemie a nestabilní hemoglobinové varianty V. Divoký, S. Walczysková, D. Pospíšilová, M. Priwitzerová, S. Takáčová, I. Kostelecká, M. Divoká, Š. Rožmanová, M. Jarošová, J. Čermák, K. Indrák a Česko-slovenská kooperativní skupina pro diagnostiku hemoglobinopatií
, není schopen přenášet kyslík. V erythrocytech vzniká za fyziologických podmínek v malém množství přímou oxidací železa kyslíkem; je zde enzymově zpětně redukován na Fe2+ formu. Větší množství metHb vzniká následkem otrav (acetanilid, chlornany, dusitany atd.) nebo vrozených poruch syntézy hemoglobinu. Může se patologicky objevovat v krevní plasmě (methemoglobinemie) nebo v moči (methemoglobinurie). Způsobuje modrání kůže, obecně, sliznic (cyanóza). Talasemie - disproporce tvorby řetězců a a b hemoglobinu. Některé vzácnější formy hereditárních anémií vyskytující se v dospělé populaci v ČR a SR – β−talasemie a nestabilní hemoglobinové varianty. V. Divoký, S. Walczysková, D. Pospíšilová, M. Priwitzerová, S. Takáčová, I. Kostelecká, M. Divoká, Š. Rožmanová, M. Jarošová, J. Čermák, K. Indrák a Česko-slovenská kooperativní skupina pro diagnostiku hemoglobinopatií.")




 ústojný systém krve. V plasmě je ústojný systém dvojice hydrogenuhličitan / kyselina uhličitá: H 2 CO 3 H + + HCO.>")

















