Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Neuropřenašeče: serotonin, acetylcholin, histamin
Neurochemie: Neuropřenašeče: serotonin, acetylcholin, histamin 5. přednáška

2
V lidském mozku ještě další malé neuropřenašeče: serotonin, acetylcholin a histamin
serotonin a histamin = monoaminy díky přítomnosti jedné aminoskupiny všechny monoaminy uvolňovány z relativně malého počtu neuronů široce projikujících v rámci celé neuraxis acetylcholin není monoamin - bývá k nim přiřazován proto, že je rovněž uvolňován z nevelkého počtu neuronů (receptory spřažené s G-proteiny) oproti monoaminům nehomogenní skupina: serotonin z tryprofanu, acetylcholin složený z cholinu a acetyl CoA, histamin z histidinu SEROTONIN (5-hydroxytryptamin, 5-HT) 90% celkového tělesného serotoninu se nachází v enterochromafinních buňkách střeva, kde regulují střevní motilitu. Zbylých 10% produkují serotoninergní neurony CNS. za fyziologických stavů = regulace nálady, spánku, ovlivňování respirace, termoregulace či vasokonstrikce (poslední tři hlavně cestou 5-HT7 Rs) etiogeneze celé řady onemocnění, počínaje migrenickými bolestmi hlavy a konče depresemi velké množství subtypů (role 5-HT je velmi různorodá a její patofyziologie vysoce zajímavá)
 oproti monoaminům nehomogenní skupina: serotonin z tryprofanu, acetylcholin složený z cholinu a acetyl CoA, histamin z histidinu. SEROTONIN (5-hydroxytryptamin, 5-HT) 90% celkového tělesného serotoninu se nachází v enterochromafinních buňkách střeva, kde regulují střevní motilitu. Zbylých 10% produkují serotoninergní neurony CNS. za fyziologických stavů = regulace nálady, spánku, ovlivňování respirace, termoregulace či vasokonstrikce (poslední tři hlavně cestou 5-HT7 Rs) etiogeneze celé řady onemocnění, počínaje migrenickými bolestmi hlavy a konče depresemi. velké množství subtypů (role 5-HT je velmi různorodá a její patofyziologie vysoce zajímavá)")
3
Syntéza serotoninu serotonin (5-hydroxytryotamin) – indolový skelet, který nese na uhlíku 5 hydroxyskupinu a na konci uhlíkatého řetězce je vybaven terminální aminoskupinou syntetizován z aminokyseliny tryptofanu tryptofan aktivně transportován přes hematoencefalickou bariéru následně hydroxylován pomocí tryptofanhydroxylasay (TPH) vzniklý 5-hydroxytryptofan dekarboxylován na serotonin dekarboxylasou aromatických kyselin (AADC) v šišince následuje ještě konverze serotoninu na melatonin pomocí hydroxyindol-O-methyltransferasy
 – indolový skelet, který nese na uhlíku 5 hydroxyskupinu a na konci uhlíkatého řetězce je vybaven terminální aminoskupinou syntetizován z aminokyseliny tryptofanu tryptofan aktivně transportován přes hematoencefalickou bariéru následně hydroxylován pomocí tryptofanhydroxylasay (TPH) vzniklý 5-hydroxytryptofan dekarboxylován na serotonin dekarboxylasou aromatických kyselin (AADC) v šišince následuje ještě konverze serotoninu na melatonin pomocí 5-hydroxyindol-O-methyltransferasy.")
4
Tryptofanhydroxylasa
TPH = tzv. rate-limiting enzymem syntetické dráhy serotoninu (a melatoninu) obsahuje ion dvojmocného železa a tvoří homotetramer se doménami k každé podjednotce ke své funkci vyžaduje tetrahydrobiopterin a kyslík v lidské tkáni a ve všech savčích tkáních existují dvě isoformy, TPH1 a TPH2 každá kódována jiným genem (ležícím na chromosomech 11 a 12) ) sekvenční homologie je 71% N-koncová doména TPH2 je o 46 aminokyselinových zbytků delší TPH2 je predominantně exprimována v serotoninergních neuronech mozku a střevní tkáni TPH1 se vyskytuje v ostatních částech CNS (např. v šišince), v intestinální mukose, enterochromafinních buňkách, kožních buňkách či mastocytech
 obsahuje ion dvojmocného železa a tvoří homotetramer se 3 doménami k každé podjednotce ke své funkci vyžaduje tetrahydrobiopterin a kyslík v lidské tkáni a ve všech savčích tkáních existují dvě isoformy, TPH1 a TPH2 každá kódována jiným genem (ležícím na chromosomech 11 a 12) ) sekvenční homologie je 71% N-koncová doména TPH2 je o 46 aminokyselinových zbytků delší TPH2 je predominantně exprimována v serotoninergních neuronech mozku a střevní tkáni TPH1 se vyskytuje v ostatních částech CNS (např. v šišince), v intestinální mukose, enterochromafinních buňkách, kožních buňkách či mastocytech.")
5
Regulace hladiny serotoninu
TPH: krátkodobé i dlouhodobé regulace TPH jsou v podstatě identické s regulacemi tyrosinhydroxylasy TPH může být aktivována proteinkinasou A a některými Ca2+/kalmodulin-dependentními proteinkinasami, nejspíše CaM-kinasou II promotor jejího genu může být aktivován pomocí cAMP na CRE (cAMP-response element) p-chlorofenylalanin (PCPA) irrverzibilně blokuje TPH a způsobují dlouhodbou depleci serotoninu Další cesty regulace: halogenované amfetaminy (fenfluramin) indukují rychlý výlev serotoninu z neuronů narušují skladování serotoninu ve váčcích ten se pak uvolňuje do cytoplasmy následně transportován do synaptické štěrbiny serotoninovým transportérem, fungujícím reverzně oproti své normální přenosové funkci dlouhodobá expozice těmto látkám může způsobit depleci serotoninu z neuronů a s ní spojenou down-regulaci TPH a serotoninového transportéru
 p-chlorofenylalanin (PCPA) irrverzibilně blokuje TPH a způsobují dlouhodbou depleci serotoninu. Další cesty regulace: halogenované amfetaminy (fenfluramin) indukují rychlý výlev serotoninu z neuronů narušují skladování serotoninu ve váčcích ten se pak uvolňuje do cytoplasmy následně transportován do synaptické štěrbiny serotoninovým transportérem, fungujícím reverzně oproti své normální přenosové funkci dlouhodobá expozice těmto látkám může způsobit depleci serotoninu z neuronů a s ní spojenou down-regulaci TPH a serotoninového transportéru.")
6
Regulace hladiny serotoninu - tryptofan
vyloučení tryptofanu z potravy vede k více než 90% snížení hladiny serotoninu v mozku u pacientů zotavujících se z deprese vyvolá deplece tryptofanu z potravy návrat depresivních syndromů... ... ale jen v té skupině, která byla léčena selektivními inhibitory zpětného vychytávání serotoninu (SSRI), nikoliv jinými typy antidepresiv potravní deplece tryptofanu ovšem nevedla k nástupu depresivních syndromů u pacientů s obsesivně-kompulzivními poruchami, ani u zdravých jedinců ztráta serotoninu per se nemůže způsobit depresi u jedinců, kteří již touto poruchou netrpěli úroveň syntézy serotoninu se zvýší po orálním podání L-tryptofanu (TPH není za normálních podmínek saturována) až 7g dávky tryptofanu vyvolaly mírný sedativní efekt a lehce zvýšily hladiny hormonů podvěsku mozkového (prolaktin), které serotonin reguluje kontaminované komerční preparáty tryptofanu způsobily např. vážnou formu oesinofilního- myalgického syndromu. odbouráván monoaminooxidasani. serotoninergní neurony exprimují MAOB s nižší afinitou k serotoninu než MAOA tyto neurony obsahují cytoplasmatickou zásobu serotoninu primární úkol MAOB asi je degradovat stopové aminy – potenciální falešné neuropřenašeče extracelulárně degradován pomocí MAOA jiného původu produktem oxidace je kyselina 5-hydroxyindoloctová (5-HIAA) hladina 5-HIAA koreluje s projevy impulsivního násilí Degradace serotoninu
, nikoliv jinými typy antidepresiv potravní deplece tryptofanu ovšem nevedla k nástupu depresivních syndromů u pacientů s obsesivně-kompulzivními poruchami, ani u zdravých jedinců ztráta serotoninu per se nemůže způsobit depresi u jedinců, kteří již touto poruchou netrpěli úroveň syntézy serotoninu se zvýší po orálním podání L-tryptofanu (TPH není za normálních podmínek saturována) až 7g dávky tryptofanu vyvolaly mírný sedativní efekt a lehce zvýšily hladiny hormonů podvěsku mozkového (prolaktin), které serotonin reguluje kontaminované komerční preparáty tryptofanu způsobily např. vážnou formu oesinofilního- myalgického syndromu. odbouráván monoaminooxidasani. serotoninergní neurony exprimují MAOB s nižší afinitou k serotoninu než MAOA tyto neurony obsahují cytoplasmatickou zásobu serotoninu primární úkol MAOB asi je degradovat stopové aminy – potenciální falešné neuropřenašeče extracelulárně degradován pomocí MAOA jiného původu produktem oxidace je kyselina 5-hydroxyindoloctová (5-HIAA) hladina 5-HIAA koreluje s projevy impulsivního násilí. Degradace serotoninu.")
7
Funkční neuroanatomie
lidský mozek = několik set tisíc serotoninergních neuronů lokalizovány jen do několika málo jader mozkového kmene dvě rostrální a dvě kaudální (dorsální) nuclei raphe leží ve středové ose mozkového kmene mezi středním mozkem a prodlouženou míchou ovlivňují míšní dráhy zapojené do přenosu bolestivých signálů a aktivitu míšních interneuronů a motoneuronů rostrální jádra inervují téměř celý mozek společně s projekcemi z locus coeruleus tvoří část ARAS MDMA (extáze) extáze = (RS)-1-(benzo[d][1,3]dioxol-5-yl)-N-methylpropan-2-amin neurony dosrálního raphe jsou k působení extáze citlivější než jiné serotoninergní neurony v univerzitních kampusech od 80. let jako klubová droga ligand serotoninového 5-HT2 receptoru vykazuje vysokou afinitu pro 5-HT1 receptory zlepšuje náladu a alteruje percepci vedlejší účinky: tachykardie, hypertermie, agitace a úzkostné ataky MDMA ničí serotoninergní neurony v neuronu vyvolává oxidační stres podobně jako MPTP na neuronech dopaminergních trvalé poškození serotoninergních neuronů (ztráta euforigenních efektů vyvolaných MDMA může být u jejích konzumentů vyvolána jejich počínající ztrátou)
 nuclei raphe leží ve středové ose mozkového kmene mezi středním mozkem a prodlouženou míchou ovlivňují míšní dráhy zapojené do přenosu bolestivých signálů a aktivitu míšních interneuronů a motoneuronů rostrální jádra inervují téměř celý mozek společně s projekcemi z locus coeruleus tvoří část ARAS. MDMA (extáze) extáze = (RS)-1-(benzo[d][1,3]dioxol-5-yl)-N-methylpropan-2-amin neurony dosrálního raphe jsou k působení extáze citlivější než jiné serotoninergní neurony v univerzitních kampusech od 80. let jako klubová droga ligand serotoninového 5-HT2 receptoru vykazuje vysokou afinitu pro 5-HT1 receptory zlepšuje náladu a alteruje percepci vedlejší účinky: tachykardie, hypertermie, agitace a úzkostné ataky MDMA ničí serotoninergní neurony v neuronu vyvolává oxidační stres podobně jako MPTP na neuronech dopaminergních trvalé poškození serotoninergních neuronů (ztráta euforigenních efektů vyvolaných MDMA může být u jejích konzumentů vyvolána jejich počínající ztrátou)")
8
Skladování, uvolňování, zpětné vychytávání serotoninu
do synaptických váčků přenášen pomocí sejného přenašeče monoaminů (VMAT) jako katecholaminy i plnění serotoninových váčků blokují a dlouhodobou synaptickou depleci serotoninu vyvolávají stejná farmka, např. reserpin nebo tetrabenazin uvolňován standardně synapticky v synaptické štěrbině jej může potkat jeden z následujích osudů: 1) difúze ze synaptické štrbiny 2) katabolizace pomocí MAO 3) aktivace presynaptických receptorů 4) aktivace postsynaptických receptorů a 5) jeho zpětné vychytání presynaptickým serotoninovým transportérem. Serotoninový transportér (SERT) lidský serotoninový transportér (SERT) je kódován jedním genem protein délky 630 aminokyselin, obsahující 2 glykosylační místa a 8 serin-threoninových míst fosforylačních jeho homologie s transportéry pro dopamin a noradrenalin je asi 48% a má i podobnou strukturu (12 TM segmentů, N- a C- koncové intracelulární smyčky)
 jako katecholaminy i plnění serotoninových váčků blokují a dlouhodobou synaptickou depleci serotoninu vyvolávají stejná farmka, např. reserpin nebo tetrabenazin. uvolňován standardně synapticky v synaptické štěrbině jej může potkat jeden z následujích osudů: 1) difúze ze synaptické štrbiny. 2) katabolizace pomocí MAO. 3) aktivace presynaptických receptorů. 4) aktivace postsynaptických receptorů a. 5) jeho zpětné vychytání presynaptickým serotoninovým transportérem. Serotoninový transportér (SERT) lidský serotoninový transportér (SERT) je kódován jedním genem protein délky 630 aminokyselin, obsahující 2 glykosylační místa a 8 serin-threoninových míst fosforylačních jeho homologie s transportéry pro dopamin a noradrenalin je asi 48% a má i podobnou strukturu (12 TM segmentů, N- a C- koncové intracelulární smyčky)")
9
Serotoninový transportér (SERT)
exprese SERT mRNA je nejvýraznější v ncl. raphe stopovatelná je také v dorzálním a mediánním jádře v jiných jádrech mozkového kmene se SERT mRNA nevyskytuje SERT protein ovšem detekovatelný v celém CNS díky rozsáhlým projekcím serotoninergních neuronů nejvíce v neostriatu, amygdale, septu, substantia nigra a mozkové kůře lokalizován na terminálách a axonálnách varikozitách látky inhibující SERT prodlužují serotoninergní signalizaci s vysokou afinitou se na SERT váží mnohá SSRI antidepresiva (fluoxetin, sertalin, fluvoxamin, paroxetin nebo citalopram) k léčbě depresí a panických poruch ve vysokých dávkách slouží i k léčbě obsesivně-kompulzivních poruch i v terapii posttraumatikcého stresu či generalizované úzkostné poruchy afinitu k SERT vykazují i některá tricyklická antidepresiva (clomipramin) SERT i DAT se se zhruba stejnou afinitou váže kokain a inhibuje je amfetamin naopak transportér pro serotonin a dopamin stimuluje a potencuje výlev dopaminu i serotoninu účinek kokainu i amfetaminu je ale v případě dopaminergní transmise daleko výraznější než v případě serotoninu nástup účinku inhibitorů SERT dlouhodobý (v řádech týdnů) působení nejspíše zahrnuje změnu v expresi neuronálních genů, promotor lidského SERT genu obsahuje regulační sekvence pro AP-1, AP-2, SP-1 a CRE expresi SERT mRNA zřejmě modulují i antidepresiva (pacienti s depresi vykazují vyšší výskyt SERT ve středním mozku)
 k léčbě depresí a panických poruch ve vysokých dávkách slouží i k léčbě obsesivně-kompulzivních poruch i v terapii posttraumatikcého stresu či generalizované úzkostné poruchy afinitu k SERT vykazují i některá tricyklická antidepresiva (clomipramin) SERT i DAT se se zhruba stejnou afinitou váže kokain a inhibuje je amfetamin naopak transportér pro serotonin a dopamin stimuluje a potencuje výlev dopaminu i serotoninu účinek kokainu i amfetaminu je ale v případě dopaminergní transmise daleko výraznější než v případě serotoninu. nástup účinku inhibitorů SERT dlouhodobý (v řádech týdnů) působení nejspíše zahrnuje změnu v expresi neuronálních genů, promotor lidského SERT genu obsahuje regulační sekvence pro AP-1, AP-2, SP-1 a CRE expresi SERT mRNA zřejmě modulují i antidepresiva (pacienti s depresi vykazují vyšší výskyt SERT ve středním mozku)")
10
Serotoninové receptory
rodina 5-HT receptorů poměrně široká identifikováno nejméně 14 receptorových subtypů každý z nich se specifickou strukturou, farmakologií a distribučním vzorcem nomenklatura 5-HT receptorů hybridem mezi původní nomenklaturou farmakologickou a novější nomenklaturou „klonovací“ 13 x 5-HT receptorů = receptory spřažené s G- proteiny jeden (5-HT3) = ligandem ovládaný napěťový kanál neselektivní pro ionty sodíku a draslíku (event. Ca2+) aktivace 5-HT3 R vede k rychlým a přechodným depolarizacím pentamer, buď monomer 5-HT3A nebo heteromer 5-HT3A + 5-HT3B
 = ligandem ovládaný napěťový kanál neselektivní pro ionty sodíku a draslíku (event. Ca2+) aktivace 5-HT3 R vede k rychlým a přechodným depolarizacím pentamer, buď monomer 5-HT3A nebo heteromer 5-HT3A + 5-HT3B.")
11
Serotoninové receptory

12
Serotoninové receptory

13
Serotoninové autoreceptory
5-HT1A = somatodendritický autoreceptor (výskyt na tělech a dendtritech serotoninergních neuronů aktivace 5-HT1A receptoru vede k poklesu aktivity neuronu a snížení syntézy serotoninu aktivace presynaptického 5-HT1D autoreceptoru (a 5-HT1B autoreceptoru u hlodavců) lokálně snižuje syntézu a výlev neuropřenašeče 5-HT1A a 5-HT1D autoreceptory vysoce strukturně podobné oba spřaženy s Gi proteinem (inhibiční působení = cestou aktivace dovnitř usměrňujících draslíkových kanálů a inhibice napěťově ovládaných kanálů vápníkových) Farmakologie serotoninového receptoru pro své jednotlivé subtypy specifickými ligandy množství látek s definovanou klinickou aplikací parciální agonisté 5-HT1A receptoru = v terapii generalizované úzkostné poruchy (GAD): buspiron a gepiron méně účinné než tato dvě farmaka jsou benzodiazepiny, které ale nezpůsobují závislost, takže jsou rovněž v terapii GAD používány agonista 5-HT1A receptoru sumatriptan, který částečně působí i na 5-HT1D receptor je od roku 1993 užíván jako vysoce efektivní látka v léčbě migrény všechny známé halucinogeny = parciální agonisté 5-HT2A receptorů ; potentní antagonisté 5-HT2A receptorů = mnohá antipsychotika (clozapin, risperidon, olanzapin); zároveň anatagonisté D2 dopaminergních receptorů schizofrenie = vysoce selektivní antagonisté 5-HT2A receptorů (MDL , v testech)
 lokálně snižuje syntézu a výlev neuropřenašeče 5-HT1A a 5-HT1D autoreceptory vysoce strukturně podobné oba spřaženy s Gi proteinem (inhibiční působení = cestou aktivace dovnitř usměrňujících draslíkových kanálů a inhibice napěťově ovládaných kanálů vápníkových) Farmakologie serotoninového receptoru. pro své jednotlivé subtypy specifickými ligandy množství látek s definovanou klinickou aplikací parciální agonisté 5-HT1A receptoru = v terapii generalizované úzkostné poruchy (GAD): buspiron a gepiron méně účinné než tato dvě farmaka jsou benzodiazepiny, které ale nezpůsobují závislost, takže jsou rovněž v terapii GAD používány agonista 5-HT1A receptoru sumatriptan, který částečně působí i na 5-HT1D receptor je od roku 1993 užíván jako vysoce efektivní látka v léčbě migrény všechny známé halucinogeny = parciální agonisté 5-HT2A receptorů ; potentní antagonisté 5-HT2A receptorů = mnohá antipsychotika (clozapin, risperidon, olanzapin); zároveň anatagonisté D2 dopaminergních receptorů schizofrenie = vysoce selektivní antagonisté 5-HT2A receptorů (MDL , v testech)")
14
Farmakologie serotoninového receptoru
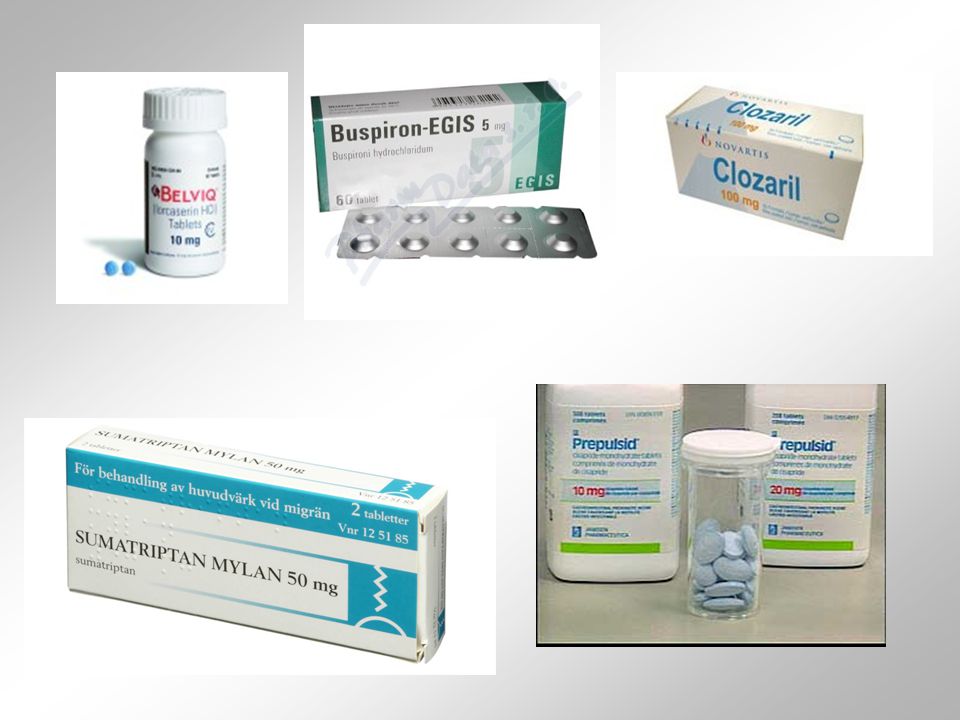
5-HT2C agonista lorcaserin: dokončená klinická studie fáze III, od 2012 v USA k léčbě obezity asi 30% pacientů vykazovalo 5% úbytek váhy při 10 mg b.i.d., s minimem vedlejších účinků (Lorqess) 5-HT4 agonista cisaprid: zvyšuje motilitu GIT díky stimulaci výlevu ACh v tlustém střevě (prokinetická terapie GER: Prepulsid) nová generace parciálních agonistů v terapii srdečních arytmií, neurodegenerativních onemocnění či inkontinence 5-HT2 ligandy 5-HT7: selektivní antagonisté potenciálně v terapii poruch spánku - při podání a počátku spánkové periody zkracují dobu REM spánku 5-HT4 ligandy 5-HT7 ligandy
 5-HT4 agonista cisaprid: zvyšuje motilitu GIT díky stimulaci výlevu ACh v tlustém střevě (prokinetická terapie GER: Prepulsid) nová generace parciálních agonistů v terapii srdečních arytmií, neurodegenerativních onemocnění či inkontinence. 5-HT2 ligandy. 5-HT7: selektivní antagonisté potenciálně v terapii poruch spánku - při podání a počátku spánkové periody zkracují dobu REM spánku. 5-HT4 ligandy. 5-HT7 ligandy.")
16
Farmakologie serotoninového receptoru
antagonisté 5-HT3 receptorů = antiemetika, užíváni k minimalizaci nausey a zvracení vyvolaného chemoterapií (ondansetron, granisetron) na zvířecích modelech prokázány také jejich anxiolytické a paměť posilující účinky agonista 5-HT4 receptoru metoclopramid (který mj. také antagonizuje D2 dopaminergní receptory) = používán k potlačování nausey a zvracení agonisté 5-HT5 receptorů silná antiemetika antagonisté 5-HT6 a 5-HT7 receptorů (aripiprazol) = využíváni jako atypická antipsychotika (clozapin, risperidon) a antidepresiva v léčbě schizofrenií, bipolárních poruch a deprese, od roku 2008 se v USA používají i v terapii autismu Sloučeniny, o kterých se vědělo, že účinkují na jiných receptorových cílech, se ukázaly i jako ligandy 5-HT receptorů: tricyklická antidepresiva (amitriptylin), která blokují též noradrenalinové receptory, antagonizují 5-HT6 a 5-HT7 receptory SSRI fluoxetin antagonizuje i 5-HT2C receptory pozorování jsou přínosná pro rozvoj nových antidepresiv Neselektivni 5-HT agonista = LSD (1943, ingesce při syntéze – Albert Hofman)
 na zvířecích modelech prokázány také jejich anxiolytické a paměť posilující účinky. agonista 5-HT4 receptoru metoclopramid (který mj. také antagonizuje D2 dopaminergní receptory) = používán k potlačování nausey a zvracení agonisté 5-HT5 receptorů silná antiemetika. antagonisté 5-HT6 a 5-HT7 receptorů (aripiprazol) = využíváni jako atypická antipsychotika (clozapin, risperidon) a antidepresiva v léčbě schizofrenií, bipolárních poruch a deprese, od roku 2008 se v USA používají i v terapii autismu. Sloučeniny, o kterých se vědělo, že účinkují na jiných receptorových cílech, se ukázaly i jako ligandy 5-HT receptorů: tricyklická antidepresiva (amitriptylin), která blokují též noradrenalinové receptory, antagonizují 5-HT6 a 5-HT7 receptory SSRI fluoxetin antagonizuje i 5-HT2C receptory pozorování jsou přínosná pro rozvoj nových antidepresiv. Neselektivni 5-HT agonista = LSD (1943, ingesce při syntéze – Albert Hofman)")
17
ACETYLCHOLIN (ACh) acetylcholin (ACh) = díky pracím Dalea, Loewiho, Feldberga a dalších v první třetině minulého století prvním objeveným neuropřenašečem nejdříve nazván „vagusstoff“ diky svému působení v rámci parasympatiku poměrně záhy byl popsán i jako neuropřenašeč nervosvalového spojení a CNS syntetizován reverzibilní reakcí, při níž cholinacetyltransferasa (ChAT) přenáší acetylovou skupinu z acetylkoenzymu A na choli za vzniku acetylcholinu rychlost reakce je limitována dostupností cholinu cholin je transportován do terminál buď volný, nebo vázaný na membránové fosfolipidy většina ACh syntetizována v nervových zakončeních bohatých na mitochondrie (producenty acetylkoenzymu A) a cholin synaptické působení ACh je ukončeno acetylcholinesterasou (AChE) hydrolyzuje ACh na acetát a cholin AChE = nesmírně výkonný enzym schopný hydrolyzovat 1000 molekul ACh za vteřinu na jednu molekulu enzymu v cytoplasmě a vnější cytoplasmatické membráně, takže je schopna hydrolyzovat ACh extra- i intracelulárně HS-CoA + + Cholinacetyltransferasa (Acetylcholinesterasa) -S-CoA
 = díky pracím Dalea, Loewiho, Feldberga a dalších v první třetině minulého století prvním objeveným neuropřenašečem nejdříve nazván „vagusstoff diky svému působení v rámci parasympatiku poměrně záhy byl popsán i jako neuropřenašeč nervosvalového spojení a CNS syntetizován reverzibilní reakcí, při níž cholinacetyltransferasa (ChAT) přenáší acetylovou skupinu z acetylkoenzymu A na choli za vzniku acetylcholinu rychlost reakce je limitována dostupností cholinu cholin je transportován do terminál buď volný, nebo vázaný na membránové fosfolipidy většina ACh syntetizována v nervových zakončeních bohatých na mitochondrie (producenty acetylkoenzymu A) a cholin synaptické působení ACh je ukončeno acetylcholinesterasou (AChE) hydrolyzuje ACh na acetát a cholin AChE = nesmírně výkonný enzym schopný hydrolyzovat 1000 molekul ACh za vteřinu na jednu molekulu enzymu v cytoplasmě a vnější cytoplasmatické membráně, takže je schopna hydrolyzovat ACh extra- i intracelulárně. HS-CoA + + Cholinacetyltransferasa. (Acetylcholinesterasa) -S-CoA.")
18
Acetylcholinesterasa „v akci“
Řetězce: A, B, C, D EC: Klasifikace: Hydrolasa Acetylcholinesterasa „v akci“ Tzv. asymetrická forma (1 000 kDa): 4 katalytické jednotky / řetězec „ocásek“ kolagenové struktury (3 polypeptidové řetězce) zakotvený v bazální membráně nervosvalového spojení AChE – „dokonalý“ enzym ! (40) 80 ms ! na molekulu
: 4 katalytické jednotky / řetězec. „ocásek kolagenové struktury (3 polypeptidové řetězce) zakotvený v bazální membráně nervosvalového spojení. AChE – „dokonalý enzym. ! (40) 80 ms ! na molekulu.")
19
anticholinesterasy = inhibují AChE způsobují extracelulární akumulaci acetylcholinu – tedy efekt shodný s nadměrnou stimulaci acetylcholinergních zakončení NS reverzibilní inhibitory (fysostigmin či neostigmin) = inhbiice AChE po dobu cca 4 h klinicky užívány v léčbě glaukomu, myastenie gravis, dysfunkce hladké svaloviny močového měchýře a střev aj. neostigmin nemůže procházet hematoencefalickou bariérou (kvartérní amoniová sůl) v CNS využívány anticholinesterasy jako tacrin nebo donepezil zvyšují hladinu ACh u pacientů s Alzheimerovou chorobou (jen mírně terapeuticky účinné) irreverzibilní anticholinesterasy fosforylují AChE a kompletně blokují odbourávání ACh pro obnovení funkce synapse nutná syntéza nových enzymových molekul insekticidy, při požití pro člověka vysoce toxické hlavní třída nervových plynů (sarin, tabun či soman) každá z těchto látek může způsobit úmrtí do 5 minut od expozice primární příčinou smrti je respirační selhání, kterému předchází celá řada autonomních příznaků a kognitivních poruch Sarin neostigmin fysostigmin Tabun

20
1. vznik fosforylenzymu -HF + H-O-Ser-ACE Ser-AChE 2. vznik karbamylenzymu + H-O-Ser-ACE Ser-AChE Co s tím? Antidota (pyridinaloxin, bispyridin) Atropine is not an actual antidote for organophosphate poisoning. However, by blocking the action of acetylcholine at muscarinic receptors, atropine also serves as a treatment for poisoning by organophosphate insecticides and nerve gases, such as Tabun (GA), Sarin (GB) and Soman (GD). Troops that are likely to be attacked with chemical weapons often carry autoinjectors with atropine and obidoxime, which can be quickly injected into the thigh. Atropine is often used in conjunction with Pralidoxime chloride. Atropine is given as a treatment for SLUDGE (Salivation, Lacrimation, Urination, Diaphoresis, Gastrointestinal motility, Emesis) symptoms caused by organophosphate poisoning. Another mnemonic is DUMBBELSS, which stands for Diarrhea, Urination, Miosis, Bradycardia, Bronchoconstriction, Excitation (as of muscle in the form of fasciculations and CNS), Lacrimation, Salivation, and Sweating (only sympathetic innervation using Musc receptors Ser-AChE + + H-O-Ser-AChE
 Atropine is not an actual antidote for organophosphate poisoning. However, by blocking the action of acetylcholine at muscarinic receptors, atropine also serves as a treatment for poisoning by organophosphate insecticides and nerve gases, such as Tabun (GA), Sarin (GB) and Soman (GD). Troops that are likely to be attacked with chemical weapons often carry autoinjectors with atropine and obidoxime, which can be quickly injected into the thigh. Atropine is often used in conjunction with Pralidoxime chloride. Atropine is given as a treatment for SLUDGE (Salivation, Lacrimation, Urination, Diaphoresis, Gastrointestinal motility, Emesis) symptoms caused by organophosphate poisoning. Another mnemonic is DUMBBELSS, which stands for Diarrhea, Urination, Miosis, Bradycardia, Bronchoconstriction, Excitation (as of muscle in the form of fasciculations and CNS), Lacrimation, Salivation, and Sweating (only sympathetic innervation using Musc receptors. Ser-AChE. + + H-O-Ser-AChE.")
21
Cholinergní dráhy v mozku
rychlá excitační nikotinická synapse pro CNS poměrně atypická odpovědí zprostředkované spíše ACh receptory muskarinovými ( víc) ve striatu jako „interní“ neuropřenašeč: skupina tonicky aktivních neuronů (TANs) interneuronů s hladkými dendrity v jádrech roztroušených v celém mozku jejich axony inervují většinu oblastí CNS hlavní cholinergní vstupy do kortexu a hippokampu z jader na spodině předního mozku, zejména z jader septa a ncl. basalis typické rozvětvené difusní projekce inervující kůru mozkovou, hippokampus, amygdalu, thalamus a mozkový kmen léze ncl. basalis snižuje hladinu acetylcholintransferasy v mozkové kůře o víc než 50 velké cholinergní neurony inervující oblast striata jsou důležité pro extrarapyramidovou kontrolu motoriky a některé formy implicitní paměti u parkinsonických pacientů vede ztráta dopaminergní inervace striata k prohloubení motorických obtíží antagonisté muskarinových acetylcholinovcýh receptorů (hexefenidyl, benzotropin) tyto příznaky zmírňují účastní se procesů učení, kognice a vzniku paměťových stop tvorba paměťových stop ovšem není zprostředkována pouze cholinergními dráhami; ty jsou především modulačními vstupními cestami ke kortikálním a hippokampálním neuronům ACh může na pyramidové buňky kůry mozkové působit excitačně nebo inhibičně excitace působením ACh na M1 receptory na vlastních pyramidových buňkách inhibice navázáním ACh na M2 receptory GABAergních interneuronů.
 ve striatu jako „interní neuropřenašeč: skupina tonicky aktivních neuronů (TANs) interneuronů s hladkými dendrity. v jádrech roztroušených v celém mozku jejich axony inervují většinu oblastí CNS hlavní cholinergní vstupy do kortexu a hippokampu z jader na spodině předního mozku, zejména z jader septa a ncl. basalis typické rozvětvené difusní projekce inervující kůru mozkovou, hippokampus, amygdalu, thalamus a mozkový kmen léze ncl. basalis snižuje hladinu acetylcholintransferasy v mozkové kůře o víc než 50 velké cholinergní neurony inervující oblast striata jsou důležité pro extrarapyramidovou kontrolu motoriky a některé formy implicitní paměti u parkinsonických pacientů vede ztráta dopaminergní inervace striata k prohloubení motorických obtíží antagonisté muskarinových acetylcholinovcýh receptorů (hexefenidyl, benzotropin) tyto příznaky zmírňují účastní se procesů učení, kognice a vzniku paměťových stop tvorba paměťových stop ovšem není zprostředkována pouze cholinergními dráhami; ty jsou především modulačními vstupními cestami ke kortikálním a hippokampálním neuronům ACh může na pyramidové buňky kůry mozkové působit excitačně nebo inhibičně excitace působením ACh na M1 receptory na vlastních pyramidových buňkách inhibice navázáním ACh na M2 receptory GABAergních interneuronů.")
22
Skladování a výlev koncentrován ve váčcích v presynaptickém zakončení transport do váčků inhibuje např. vesamicol k výlevu dochází klasicky po depolarizaci a vtoku vápníku do nervového zakončení selektivních toxinů inhibující výlev ACh: botulotoxin A nebo tetanotoxin jsou to zinkové endoproteasy, které štěpí SNARE proteiny a-latratoxin černé vdovy naopak navozuje masivní výlev ACh s následnou nadměrnou postsynaptickou stimulací. receptory pro tyto toxiny = např. neurexin, nebo nově popsaný receptor spřažený s G-proteinem. ACh není zpětně vychytáván ani do glií, ani do nervového zakončení, a není proto recyklován rychle metabolizuje, takže nemůže dojít ani k jeho dufúzi ze synaptické štěrbiny do terminály zpět je s vysokou afinitou přenášen jen cholin, tam využit pro syntézu nového neuropřenašeče transport cholinu je primárním regulačním mechanismem ovlivňujícím koncentraci acetylcholinu v synapsi inhibitory cholinového transportu (hemicholinium, vesamicol) způsobí vyčerpání zásob ACh v terminále cholinový transportér leží u člověka na genu SLC5A7.
 způsobí vyčerpání zásob ACh v terminále cholinový transportér leží u člověka na genu SLC5A7.")
23
Acetylcholinové receptory
působí na dvou typech receptorů muskarinové acetylcholinové receptory (mAChRs) = receptory spřažené s G-proteiny receptory nikotinické (nAChRs) = patří do superrodiny ligandem ovládaným iontových kanálů spolu s 5-HT3 glutamátovými, glycinovými a GABA receptory látky působící na acetylcholinové receptory jsou důležitými regulátory funkce parasympatiku Muskarinové acetylcholinové receptory váží muskarin (z muchomůrky Amanita muscaria ) v periferních tkáních, autonomním nervstvu i CNS 5 subtypů mAChRs označovaných jako M1-M5 exprimovaných v mozku odpovědi subtypů se liší podle dalšího systému druhých poslů, iontových kanálů a efektorů M1, M3 a M5 receptory obvykle působí stimulačně nebo inhibičně na fosfatidylinositolový systém M2 a M4 receptory vyvolávají zejména inhibiční odpovědi, a to buď aktivací dovnitř usměrňujících draslíkových kanálů, inhibicí kanálů vápníkových nebo inhibicí adenylylcyklasy. muskarin
 = receptory spřažené s G-proteiny receptory nikotinické (nAChRs) = patří do superrodiny ligandem ovládaným iontových kanálů spolu s 5-HT3 glutamátovými, glycinovými a GABA receptory látky působící na acetylcholinové receptory jsou důležitými regulátory funkce parasympatiku. Muskarinové acetylcholinové receptory. váží muskarin (z muchomůrky Amanita muscaria ) v periferních tkáních, autonomním nervstvu i CNS 5 subtypů mAChRs označovaných jako M1-M5 exprimovaných v mozku. odpovědi subtypů se liší podle dalšího systému druhých poslů, iontových kanálů a efektorů M1, M3 a M5 receptory obvykle působí stimulačně nebo inhibičně na fosfatidylinositolový systém M2 a M4 receptory vyvolávají zejména inhibiční odpovědi, a to buď aktivací dovnitř usměrňujících draslíkových kanálů, inhibicí kanálů vápníkových nebo inhibicí adenylylcyklasy. muskarin.")
24
muskarinový acetylcholinový receptor (mAChR)
")
25
Farmakologie muskarinových receptorů
rozsáhlá subtypově selektivní ligandů není mnoho a žádný z nich není využíván v široké klinické praxi obecní muskarinoví agonisté zahrnují přírodní látky (muskarin, pilocarpin nebo arekolin) i syntetické sloučeniny (karbachol, oxotremorin) všichni tito centrálně účinkující agonisté navozují silné slinění a pocení, spojené s korovou excitací občas klinicky využíváni v léčbě glaukomu nebo retence moči účinní při zmírňování Sjögrenova syndromu (choroba charakteristická autoimunitní degenerací slinných žláz) prototypičtí muskarinoví antagonisté = atropin a skopolamin atropin byl využíván kosmeticky jako dilatační agens zornic je užíván v oftalmologické praxi příbuzní antagonisté (benztropin) jsou užívání jako podpůrná léčba parkinsonismu a parkinsonických příznaků navozených antipsychotiky skopolamin podávaný v malých dávkách (obvykle náplastěmi transdermálně) je účinný v prevenci kinetóz Muskarinové receptory také antagonizuje mnoho psychotropních látek (tricyklická antidepresiva, antipsychotika jako chlorpromazin); jejich vazba na mAChRs se projevuje obvykle nepříjemnými vedlejšími účinky léčby (zácpa, sucho v ústech).
 i syntetické sloučeniny (karbachol, oxotremorin) všichni tito centrálně účinkující agonisté navozují silné slinění a pocení, spojené s korovou excitací občas klinicky využíváni v léčbě glaukomu nebo retence moči účinní při zmírňování Sjögrenova syndromu (choroba charakteristická autoimunitní degenerací slinných žláz) prototypičtí muskarinoví antagonisté = atropin a skopolamin atropin byl využíván kosmeticky jako dilatační agens zornic je užíván v oftalmologické praxi příbuzní antagonisté (benztropin) jsou užívání jako podpůrná léčba parkinsonismu a parkinsonických příznaků navozených antipsychotiky skopolamin podávaný v malých dávkách (obvykle náplastěmi transdermálně) je účinný v prevenci kinetóz. Muskarinové receptory také antagonizuje mnoho psychotropních látek (tricyklická antidepresiva, antipsychotika jako chlorpromazin); jejich vazba na mAChRs se projevuje obvykle nepříjemnými vedlejšími účinky léčby (zácpa, sucho v ústech).")
26
nikotinický acetylcholinový receptor (nAChR)
pentamerický protein složený z různých podjednotek každá podjednotka má 4 trans membránové segmenty (M1-M4) M2 segment má a-helikální kon formaci, tvoří vlastní pór kanálu segmenty M1, 3 a 4 zřejmě b-listy, obklopují vnitřní kruh M2 šroubovic propouští Na+, K+ a Ca2+ ionty ligand: 2 molekuly acetylcholinu působící kooperativně nAChR : 2 40 kDa, 461 AA 49 kDa, 493 AA 57 kDa, 506 AA / 65 kDa, 522 AA nAChRs – krásný příklad molekulární evoluce starobylá receptorová rodina – už u diblastik (houby, žahavci) majících jen ento- a ektoderm: ale jen různé a podjednotky (homoligomery, mil. let) triblastika (Coelomata) s mesodermem – vznik svaloviny a potřeba svalového podtypu nAChRs: diferenciace dalších podjednotkových typů (1300 mil. let)
 M2 segment má a-helikální kon- formaci, tvoří vlastní pór kanálu. segmenty M1, 3 a 4 zřejmě b-listy, obklopují vnitřní kruh M2 šroubovic. propouští Na+, K+ a Ca2+ ionty. ligand: 2 molekuly acetylcholinu působící kooperativně. nAChR : 2 40 kDa, 461 AA. 49 kDa, 493 AA. 57 kDa, 506 AA. / 65 kDa, 522 AA. nAChRs – krásný příklad molekulární evoluce. starobylá receptorová rodina – už u diblastik (houby, žahavci) majících jen ento- a ektoderm: ale jen různé a podjednotky (homoligomery, mil. let) triblastika (Coelomata) s mesodermem – vznik svaloviny a potřeba svalového podtypu nAChRs: diferenciace dalších podjednotkových typů (1300 mil. let)")
27
nacházejí se na nervosvalovém spojení, v autonomních gangliích, dření nadledvin i CNS
17 popsaných podjednotek dělí se na svalové a neuronální podtypy na různých chromosomech (2, 8, 15, ale i 1, 4, 11, 20) typická je jejich velmi rychlá desensitizace, nezávislá na dalších proteinech (jako je např. arrestin u metabotropních receptorů) je vnitřní vlastností receptoru rychlost desensitizace regulována fosforylací receptorových podjednotek pomocí proteinkinas A a C k desensitizace vede i déledobá expozice ligandu
 typická je jejich velmi rychlá desensitizace, nezávislá na dalších proteinech (jako je např. arrestin u metabotropních receptorů) je vnitřní vlastností receptoru rychlost desensitizace regulována fosforylací receptorových podjednotek pomocí proteinkinas A a C k desensitizace vede i déledobá expozice ligandu.")
28
208 náhodných kombinací při 5 podjednotkách

29
neuronální nAChRs = pentamery s centrálním pórem většina heteropentamerních, ale obsahují pouze podjednotky a a b v neuronoání tkáni bylo popsáno 8 a podjednotek a 3 b podjednotky typická stechiometrie je 2a : 3b homopentamery z podjednotek a7, které formují funkční kanály

30
svalové nikotinické acetylcholinové receptory jsou složeny z 5 podejdnotek obvyklá podjednotková kombinace dospělého svalového nAChRs je α1, β1, δ, a ε podjednotka v poměru 2:1:1:1 embryonální svalový nAChR vykazuje složení je α1, β1, δ, a g podjednotka, také v poměru 2:1:1:1 místem vazby ligandu (ACh) jsou a podjednotky, z nichž každá váže kooperativně jednu molekulu. myasthenia gravis = vzácná dědičná porucha z řekého ύς (sval), aσθένεια (slabost) a latinského gravis (vážný, těžký autoimunitní choroba popsané už roku 1977 spojena se svalovou slabostí, únavou, zejména v oblasti svalů mimickch, šíjových a pletencových 85% pacinetů má detekovatelné protilátky proti nAChRs cca 65 % nemocných má patologické změny thymu, kdy autoreaktivní B buňky produkují autoprotilátky proti nikotinovým acetylchlonovým receptorům u cca 7 % nemocných jsou detekovatelné protilátky proti tyrosinkinase (Ab anti MuSK) u cca % pacientů je přítomen thymom tvoří se protilátky proti titinu, ryanodinovému receptoru aj. na úrovni nervosvalového spojení dochází ke zvýšení turnoveru ACh receptorů a k jejich větší degradaci tím klesá cholinergní přenos a dochází ke zmíněným svalovým projevům. standardní léčba = podávání inhibitorů AChE, čímž se zvyšuje dostupnost ACh v synaptické štěrbině a šance, že zaktivují zbývající receptory někdy jsou podáváni i glukokortikoidy nebo imunosupresiva, v případě maysthenické krizi se přistupuje k plasmaferéze
, aσθένεια (slabost) a latinského gravis (vážný, těžký autoimunitní choroba popsané už roku 1977 spojena se svalovou slabostí, únavou, zejména v oblasti svalů mimickch, šíjových a pletencových 85% pacinetů má detekovatelné protilátky proti nAChRs cca 65 % nemocných má patologické změny thymu, kdy autoreaktivní B buňky produkují autoprotilátky proti nikotinovým acetylchlonovým receptorům u cca 7 % nemocných jsou detekovatelné protilátky proti tyrosinkinase (Ab anti MuSK) u cca % pacientů je přítomen thymom tvoří se protilátky proti titinu, ryanodinovému receptoru aj. na úrovni nervosvalového spojení dochází ke zvýšení turnoveru ACh receptorů a k jejich větší degradaci tím klesá cholinergní přenos a dochází ke zmíněným svalovým projevům. standardní léčba = podávání inhibitorů AChE, čímž se zvyšuje dostupnost ACh v synaptické štěrbině a šance, že zaktivují zbývající receptory někdy jsou podáváni i glukokortikoidy nebo imunosupresiva, v případě maysthenické krizi se přistupuje k plasmaferéze.")
31
Farmakologie nikotinických acetylcholinových receptorů
ligandů na nAChRs není mnoho svalové a v menší míře i neuronální nAChRs blokuje kurare slabý parciální agonista = sukcinycholin navozuje prodlouženou desensitizaci nAChRs a klinicky je používán během anestezie v gangliích parasympatiku i sympatetických větvích automního nervstva blokují nAChRs hexamethonium a mecamylamin velmi málo antagonistů selektivních pro nějaký subtyp nAChRs asi nejselektivnější dostupný antagonista = methyllykakonitin preferenčně antagonizuje a7 homopentamery selektivní agonista epibatidin (pouze experimentálně (jako antinocicepční agens) na nAChRs se nachází i jiná vazební místa, než jsou ta pro ACh a bungarotoxin na aktivační místo receptoru se váže i fysostigmin, inhibitor acetylcholinesterasy zvyšuje tok iontů receptorovým kanálem jiná modulace nAChRs: fenylcyklidin, chlorpromazin nebo některá anestetika někteří fungují jako nekompetitivní blokátory, které zkracují dobu otevření kanálu nebo urychlují jeho desensitizaci nAChRs inhibují i některé steroidy, vazbou na zatím neznámé místo. nikotin - vysoce návyková látka zvyšuje bdělost, zlepšuje paměť a schopnosti učení se má také určitý antinocicepční efekt, takže agonisté nAChRs by se snad mohli uplatnit v terapii bolesti
 na nAChRs se nachází i jiná vazební místa, než jsou ta pro ACh a bungarotoxin na aktivační místo receptoru se váže i fysostigmin, inhibitor acetylcholinesterasy zvyšuje tok iontů receptorovým kanálem jiná modulace nAChRs: fenylcyklidin, chlorpromazin nebo některá anestetika někteří fungují jako nekompetitivní blokátory, které zkracují dobu otevření kanálu nebo urychlují jeho desensitizaci nAChRs inhibují i některé steroidy, vazbou na zatím neznámé místo. nikotin - vysoce návyková látka zvyšuje bdělost, zlepšuje paměť a schopnosti učení se má také určitý antinocicepční efekt, takže agonisté nAChRs by se snad mohli uplatnit v terapii bolesti.")
33
Syntetické a degradativní dráhy
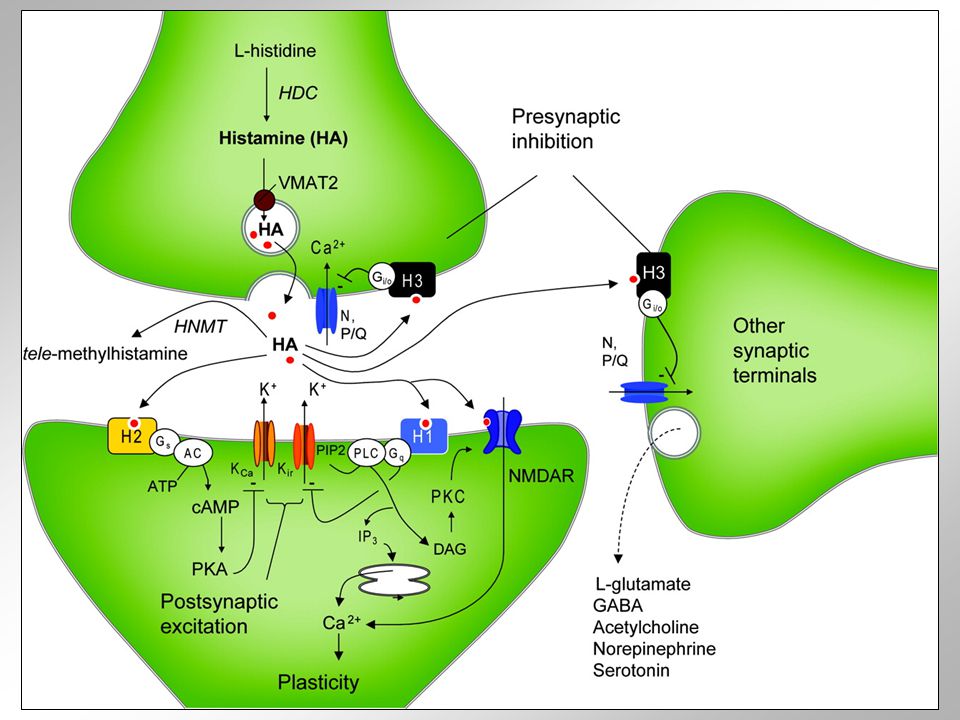
HISTAMIN velmi hojný mimo nervový systém hraje zásadní roli např. v sekreci žaludečních šťáv nebo imunitních odpovědích na alergeny je synapticky uvolňován v rámci celého CNS v CNS účinkuje přinejmenším na třech receptorových subtypech Syntetické a degradativní dráhy vzniká v jednokrokové reakci zahrnující dekarboxylaci histidinu enzymem histidindekarboxylasou proteinem o délce 662 aminokyselin její inhibitor a-fluoromethylhistidin je testován na zvířecích modelech může vznikat i pomocí dekarboxylasy aromatických aminokyselin (AADC) katabolizován na methylhistamin pomocí enzymu histamin methyltransferasy diaminové oxidasy jej eventuálně mohou konvertovat v imidazolacetaldehyd výlev standardní vychytáván zpět do terminály ev. difúze ze synaptické štěrbiny a enzymatická degradce mimo ni
 katabolizován na methylhistamin pomocí enzymu histamin methyltransferasy diaminové oxidasy jej eventuálně mohou konvertovat v imidazolacetaldehyd. výlev standardní vychytáván zpět do terminály ev. difúze ze synaptické štěrbiny a enzymatická degradce mimo ni.")
34
Funkční anatomie těla histaminergních neuronů jsou uložena v ncl. tuberomamillaris hypothalamu jejich axony s bohatými kolaterálami inervují podobně jako u ostatních biogenních aminů téměř všechny části CNS synapse jsou difúzní a jen příležitostně vytvářejí klasické pre- a postsynaptické uspořádání každé jádro obsahuje u člověka asi histaminergních neuronů inervují také gliové buňky a malé cévy a kapiláry zdá se, že regulují obecné aktivity mozku jakou je úroveň stavu bdělosti či energetický metabolismus byl prokázán vztah mezi aktivitou histaminergních neuronů a úrovní bděost nejaktivnější během vigility, zatímco během spánku pomalých vln jejich aktivita úplně vymizí ovlivňuje různé periferní tkán, účastní se např. alergické reakce, odpovědi tkáně na poranění nebo žaludeční sekrece v mozku cestou H1 receptorů depolarizuje cholinergní neurony ncl. basalis (snížením propustnosti jejich membrány pro draslík a aktivací na tetrodotoxin necitlivých sodíkových kanálů) v buňkách ganglion nodosum blokuje přes H1 receptory draslíkové kanály generující následnou hyperpolarizci po akčním potenciálu, což také vede ke zvýšení jejich excitability zapojeni do regulace uvolňování hormonů podvěsku mozkového, percepce bolesti, kontroly příjmu potravu nebo prevence závratí antialergika neprocházejí hematoencefalickou bariérou (bez sedačního efektu) centrálně účinkující antihistaminika ovšem ano; jsou součástí mnoha volně prodejných léků na spaní některá antidepresiva (mianserin, doxepin) nebo antipsychotika (klozapin) = částečně i sedativa (jsou silnými antagonisty H1 receptorů)
 v buňkách ganglion nodosum blokuje přes H1 receptory draslíkové kanály generující následnou hyperpolarizci po akčním potenciálu, což také vede ke zvýšení jejich excitability zapojeni do regulace uvolňování hormonů podvěsku mozkového, percepce bolesti, kontroly příjmu potravu nebo prevence závratí antialergika neprocházejí hematoencefalickou bariérou (bez sedačního efektu) centrálně účinkující antihistaminika ovšem ano; jsou součástí mnoha volně prodejných léků na spaní některá antidepresiva (mianserin, doxepin) nebo antipsychotika (klozapin) = částečně i sedativa (jsou silnými antagonisty H1 receptorů)")
35
Receptory pro histamin
identifikovány 4 třídy receptorů pro histamin proteiny spřažené s G-proteiny

37
H3 receptor

38
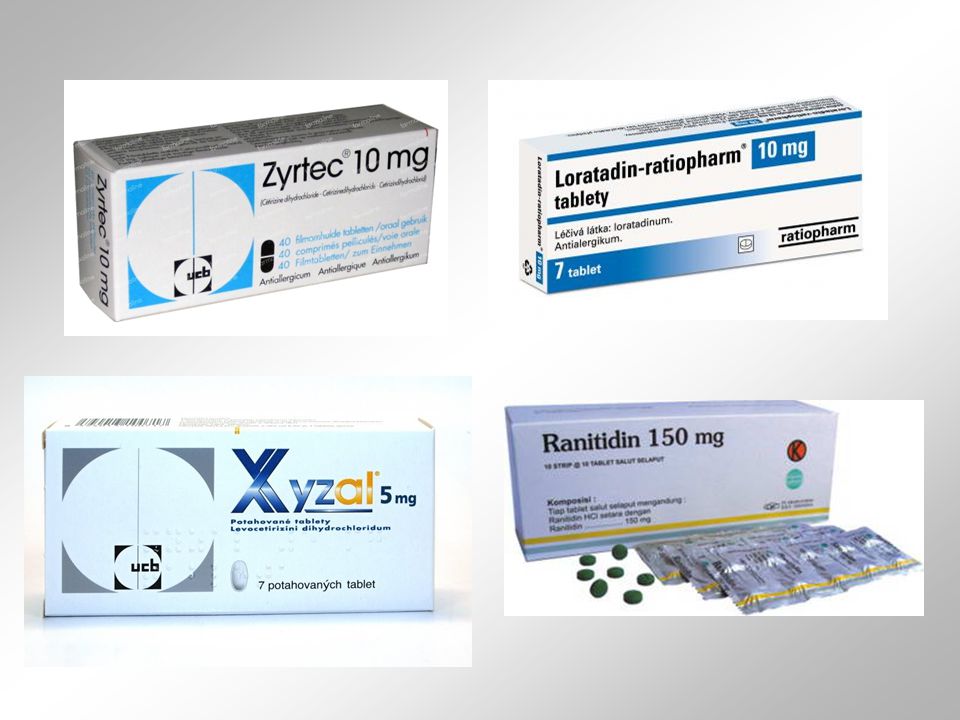
H1 receptor aktivován endogenním histaminem uvolněným z ncl. tuberomamillaris. tyto neurony „pálí“ s frekvencí asi 2 Hz za bdělosti, ve spánku jejich aktivita klesá ke 0.5 Hz ze všech neuronů vykazují nejvíce závislý cyklus „spánek-bdění“ aktivity v kortexu vede aktivace H1 receptorů k inhibici draslíkových kanálů, což má za následek depolarizaci membrány antagonisté H1 receptorů = v léčbě standardních a chladových alergií difenhydramin, meclizin a dimenhydrinát = první generace H1 antagonistů a procházejí skrze hematoencefalickou bariéru (BBB) druhá generace antihistamink (citerizin, loratadin nebo terfenandin) z mozku vylučována a předepisována jako nesedující medikace v léčbě alergií H2 receptor patří mezi ty proteiny spřažené s G-proteiny, které jsou podobny rhodopsinu silným stimulantem produkce cAMP uvolňuje vápník z vnitrobuněčných rezervoárů typický agonista H2 receptoru = betazol (dole) stimuluje sekreci žaludečních šťáv, reguluje motilitu GIT a střevní sekreci zřejmě je zapojen i do regulace buněčného růstu a diferenciace většina ligandů H2 receptorů neprochází BBB mezi antagonisty H2 receptorů patří cimetidin nebo ranitidin u malého počtu pacientů mohou způsobovat stavy zmatenosti
 druhá generace antihistamink (citerizin, loratadin nebo terfenandin) z mozku vylučována a předepisována jako nesedující medikace v léčbě alergií. H2 receptor. patří mezi ty proteiny spřažené s G-proteiny, které jsou podobny rhodopsinu silným stimulantem produkce cAMP uvolňuje vápník z vnitrobuněčných rezervoárů typický agonista H2 receptoru = betazol (dole) stimuluje sekreci žaludečních šťáv, reguluje motilitu GIT a střevní sekreci zřejmě je zapojen i do regulace buněčného růstu a diferenciace většina ligandů H2 receptorů neprochází BBB mezi antagonisty H2 receptorů patří cimetidin nebo ranitidin u malého počtu pacientů mohou způsobovat stavy zmatenosti.")
40
H3 receptor působí v CNS jako presynaptický receptor inhibuje výlev a syntézu histaminu může účinkovat i jako heteroreceptor kontrolující výlev dalších neuropřenašečů gen pro H3 receptor vykazují jen asi 20% homologii s geny pro H1 a H2 receptory existuje asi 6 jeho isoforem typičyí agonisté: (R)-a-methylhistamin navozující spánek nebo látky jako cipralisant, immepip, Imetit, Immethridine či Methimepip H4 receptor exprimován hlavně v kostní dřeni a bílé linii krvinek stimuluje uvolňování neutrofilů z kostní dřeně exprimován i v řadě jiných tkání (střevo, thymus, trachea, játra, plíce aj.) aktivuje chemotaxi mastocytů, zřejmě cestou poklesu hladiny cAMP jeho antagonisté mají protizánětlivé a anatihyperanalgestické účinky immepip (R)-a-methylhistamin
-a-methylhistamin navozující spánek nebo látky jako cipralisant, immepip, Imetit, Immethridine či Methimepip. H4 receptor. exprimován hlavně v kostní dřeni a bílé linii krvinek stimuluje uvolňování neutrofilů z kostní dřeně exprimován i v řadě jiných tkání (střevo, thymus, trachea, játra, plíce aj.) aktivuje chemotaxi mastocytů, zřejmě cestou poklesu hladiny cAMP jeho antagonisté mají protizánětlivé a anatihyperanalgestické účinky. immepip. (R)-a-methylhistamin.")
41
Co si pamatovat z dnešní přednášky
serotin – syntéza, transportér, receptory a jejich farmakologie acetylcholin – syntéza, receptory a jejich farmakologie - histamin – syntéza, receptory a jejich farmakologie

Podobné prezentace








>")











