Stáhnout prezentaci
1
Trávení a metabolismus přírodních látek (základní metabolické procesy)
Prezentace Trávení a Metabolismus se zabývá osudy potravy (tj. především sacharidů, bílkovin a triacylglycerolů) v lidském organismu, tzn. rozkladem potravy na základní stavební jednotky (tj. monosacharidy, aminokyseliny a mastné kyseliny) a dalšími procesy, kterých se tyto základní jednotky zúčastňují. Na prezentaci úzce navazuje prezentace „Citrátový cyklus a dýchací řetězec“. Milada Teplá, Helena Klímová, KUDCH, PřF UK v Praze
 v lidském organismu, tzn. rozkladem potravy na základní stavební jednotky (tj. monosacharidy, aminokyseliny a mastné kyseliny) a dalšími procesy, kterých se tyto základní jednotky zúčastňují. Na prezentaci úzce navazuje prezentace „Citrátový cyklus a dýchací řetězec . Milada Teplá, Helena Klímová, KUDCH, PřF UK v Praze.")
2
Člověk patří mezi chemoorganotrofní aerobní organismy
Chemotrofní organismy získávají energii oxidací živin. Chemotrofní organismy využívají chemické reakce jako zdroj energie. O2 potrava voda CO2 Heterotrofní (organotrofní) organismy mají za hlavní zdroj uhlíku jiné organické látky (tuky, bílkoviny, lipidy). Aerobní organismus potřebuje ke svému životu nezbytně kyslík. Co musí obsahovat naše strava? Bílkoviny Sacharidy Lipidy Vitaminy a minerální látky Vodu stolice moč
 organismy mají za hlavní zdroj uhlíku jiné organické látky (tuky, bílkoviny, lipidy). Aerobní organismus potřebuje ke svému životu nezbytně kyslík. Co musí obsahovat naše strava Bílkoviny. Sacharidy. Lipidy. Vitaminy a minerální látky. Vodu. stolice. moč.")
3
Osud potravy v našem těle
Játra Žlučník Slinivka břišní Řitní otvor Žaludek Tenké střevo Tlusté střevo Ústní dutina Potrava se v ústech žvýká a nakonec se mísí se slinami do vlhké hmoty, které říkáme sousto. Potrava Ústa Na snímku č. 6 až 12 je zanimován osud potravy v našem těle. Na těchto snímcích je stručně popsána trávicí soustava člověka. Červená kulička znázorňuje přijímanou potravu, která vstupuje do ústní dutiny.

4
Osud potravy v našem těle
Játra Žlučník Slinivka břišní Řitní otvor Žaludek Tenké střevo Tlusté střevo Ústní dutina Sousto v hltanu putuje tzv. peristaltickými pohyby až do jícnu. Pozor někdy se stane, že se spustí peristaltika zpětná neboli zvracení. Potrava Ústa Hltan a jícen Svalová stěna jícnu Kontrahovaný sval Sousto (červená kulička) putuje do jícnu. Sousto Relaxovaný sval
 putuje do jícnu. Sousto. Relaxovaný sval.")
5
Osud potravy v našem těle
Játra Žlučník Slinivka břišní Řitní otvor Žaludek Tenké střevo Tlusté střevo Ústní dutina V žaludku se jeho silné svaly stahují, a dovolují tak rozmačkat a rozdrtit obsah uvnitř na lepkavou a blátivou hmotu, které se říká trávenina (chymus). Zde se také naše potrava setkává s celou řadou enzymů. Potrava Ústa Hltan a jícen Žaludek Sousto (červená kulička) vstupuje do žaludku, kde se přeměňuje na tráveninu (chymus).
. Zde se také naše potrava setkává s celou řadou enzymů. Potrava. Ústa. Hltan a jícen. Žaludek. Sousto (červená kulička) vstupuje do žaludku, kde se přeměňuje na tráveninu (chymus).")
6
Osud potravy v našem těle
Játra Žlučník Slinivka břišní Řitní otvor Žaludek Tenké střevo Tlusté střevo Ústní dutina Chymus ve dvanáctníku obsahuje částečně strávenou potravu ze žaludku, trávicí šťávy ze slinivky břišní a žluč ze žlučníku. Potrava Ústa Hltan a jícen Žaludek Dvanáctník

7
Osud potravy v našem těle
Játra Žlučník Slinivka břišní Řitní otvor Žaludek Tenké střevo Tlusté střevo Ústní dutina Tenké střevo je hlavním místem vstřebávání živin. Potrava Ústa Hltan a jícen Žaludek Dvanáctník Tenké střevo Chymus (červená kulička) z dvanáctníku vstupuje do tenkého střeva.
 z dvanáctníku vstupuje do tenkého střeva.")
8
Osud potravy v našem těle
V tlustém střevě dochází ke vstřebávání vody (až 90%). Nestrávené zbytky (vláknina, trávicí šťávy, žluč, buňky střevní výstelky, bakterie) tvoří stolici. 1/3 suché váhy představují bakterie. Játra Žlučník Slinivka břišní Řitní otvor Žaludek Tenké střevo Tlusté střevo Ústní dutina Potrava Ústa Hltan a jícen Žaludek Dvanáctník Tenké střevo V tlustém střevě se nestrávené zbytky přeměňují na stolici (hnědá kulička). Tlusté střevo
. Nestrávené zbytky (vláknina, trávicí šťávy, žluč, buňky střevní výstelky, bakterie) tvoří stolici. 1/3 suché váhy představují bakterie. Játra. Žlučník. Slinivka břišní. Řitní otvor. Žaludek. Tenké střevo. Tlusté střevo. Ústní dutina. Potrava. Ústa. Hltan a jícen. Žaludek. Dvanáctník. Tenké střevo. V tlustém střevě se nestrávené zbytky přeměňují na stolici (hnědá kulička). Tlusté střevo.")
9
Osud potravy v našem těle
V tlustém střevě dochází ke vstřebávání vody (až 90%). Nestrávené zbytky se mění na hnědou hmotu (stolice), jejíž 1/3suché váhy představují bakterie produkující methan. Mezi nestrávené zbytky patří i vláknina. Játra Žlučník Slinivka břišní Řitní otvor Žaludek Tenké střevo Tlusté střevo Ústní dutina Potrava Ústa Hltan a jícen Žaludek Dvanáctník Tenké střevo V tlustém střevě se nestrávené zbytky přeměňují na stolici (hnědá kulička). Tlusté střevo Řitní kanál
. Nestrávené zbytky se mění na hnědou hmotu (stolice), jejíž 1/3suché váhy představují bakterie produkující methan. Mezi nestrávené zbytky patří i vláknina. Játra. Žlučník. Slinivka břišní. Řitní otvor. Žaludek. Tenké střevo. Tlusté střevo. Ústní dutina. Potrava. Ústa. Hltan a jícen. Žaludek. Dvanáctník. Tenké střevo. V tlustém střevě se nestrávené zbytky přeměňují na stolici (hnědá kulička). Tlusté střevo. Řitní kanál.")
10
Trávení Během procesu trávení dochází za pomoci hydrolytických enzymů k rozkladu potravy na jednodušší látky. nové sloučeniny trávení potrava jednodušší látky CO2, H2O, energie

11
Metabolismus Metabolismus jsou všechny chemické procesy, při nichž dochází k přeměně látek (látková výměna) a energií (energetická výměna) v buňkách a živých organismech. Metabolismus v sobě zahrnuje přeměny produktů trávení na odpadní látky, nebo výstavby nových pro život důležitých sloučenin. Co je metabolit? Metabolit je produkt metabolismu.
 a energií (energetická výměna) v buňkách a živých organismech. Metabolismus v sobě zahrnuje přeměny produktů trávení na odpadní látky, nebo výstavby nových pro život důležitých sloučenin. Co je metabolit Metabolit je produkt metabolismu.")
12
Schéma metabolismu Zažívací trubice Sacharidy Bílkoviny Lipidy
Ukládány jako glykogen v játrech Ukládány v pojivové tkáni Ukládány jako tuk v tukové tkáni Trávení Trávení Trávení Monosacharidy Aminokyseliny Mastné kyseliny Pyruvát AcetylCoA Odstranění aminoskupiny Citrátový cyklus Amoniak Energie, oxid uhličitý a voda Moč

13
Anabolismus a katabolismus
Látky chemicky jednodušší + energie Katabolismus syntéza rozklad Látky chemicky složitější Snímek znázorňuje vysvětlení rozdílu mezi anabolickými a katabolickými reakcemi. - Anabolické reakce: tj. ta část metabolismu, při níž se vytvářejí chemicky složitější látky (např. z aminokyselin vznikají bílkoviny). - Katabolické reakce: tj. ta část metabolismu, při níž jsou látky rozkládány na z chemického hlediska jednodušší látky (např. rozklad glukosy na vodu a oxid uhličitý). Anabolické reakce jsou především endergonické (energii spotřebovávají). Katabolické reakce jsou především exergonické (energii uvolňují).
. - Katabolické reakce: tj. ta část metabolismu, při níž jsou látky rozkládány na z chemického hlediska jednodušší látky (např. rozklad glukosy na vodu a oxid uhličitý). Anabolické reakce jsou především endergonické (energii spotřebovávají). Katabolické reakce jsou především exergonické (energii uvolňují).")
14
Rychlost metabolismu Rychlost metabolismu je celkově ovlivňována hormony. Jednotlivé reakce jsou katalyzovány enzymy, a tím jsou urychlovány. Čím je rychlost ovlivněna? Věkem Pohlavím Celkovým stavem organismu (fyzickým i psychickým) Regulace metabolismu se děje změnami enzymové aktivity. Protože prakticky všechny metabolické reakce jsou katalyzovány enzymy, jde o velmi účinný proces, který umožňuje, aby určité metabolické reakce probíhaly rychleji (zvýšení enzymové aktivity) nebo pomaleji (snížení enzymové aktivity). Rychlost metabolismu celkově ovlivňují hormony, které koordinují funkce tkání a orgánů. Hormony jsou produkovány určitými tkáněmi a vyplavovány do krevního oběhu. Krví se dostávají k cílovým orgánům a způsobují tam fyziologické změny, jako je např. řízení metabolických pochodů. Produkce hormonů a činnost enzymů závisí na řadě faktorů jako je věk a pohlaví. Dále záleží na celkovém stavu organismu – jak fyzickém tak psychickém. Při intenzivním cvičení (např. plavání) může rychlost metabolismu vzrůst až patnáctkrát. Sami si můžeme všimnout, že náš tep je značně rychlejší taktéž frekvence nádechu a výdechu. Při stresu je metabolismus rovněž zvýšen, proto se u stresovaných lidí zvýší rychlost vylučování odpadních látek.
 Regulace metabolismu se děje změnami enzymové aktivity. Protože prakticky všechny metabolické reakce jsou katalyzovány enzymy, jde o velmi účinný proces, který umožňuje, aby určité metabolické reakce probíhaly rychleji (zvýšení enzymové aktivity) nebo pomaleji (snížení enzymové aktivity). Rychlost metabolismu celkově ovlivňují hormony, které koordinují funkce tkání a orgánů. Hormony jsou produkovány určitými tkáněmi a vyplavovány do krevního oběhu. Krví se dostávají k cílovým orgánům a způsobují tam fyziologické změny, jako je např. řízení metabolických pochodů. Produkce hormonů a činnost enzymů závisí na řadě faktorů jako je věk a pohlaví. Dále záleží na celkovém stavu organismu – jak fyzickém tak psychickém. Při intenzivním cvičení (např. plavání) může rychlost metabolismu vzrůst až patnáctkrát. Sami si můžeme všimnout, že náš tep je značně rychlejší taktéž frekvence nádechu a výdechu. Při stresu je metabolismus rovněž zvýšen, proto se u stresovaných lidí zvýší rychlost vylučování odpadních látek.")
15
Organismy potřebují stále energii, kterou získávají rozkladem potravy.
Tuto energii spotřebují na endergonické reakce. Kde organismus energii uchovává? v tzv. makroergických sloučeninách ATP se skládá z ribosy a tří zbytků kyseliny trihydrogenfosforečné. Odtržením posledního zbytku kyseliny trihydrogenfosforečné vzniká adenosindifosfát (ADP) a anorganický fosfát (P, někdy též značeno jako Pi) za uvolnění energie: H2O + ATP → ADP + P ∆G ≈ -30 KJ Přebytečná energie z látkového metabolismu naopak umožňuje vznik ATP: ADP + P → ATP + H2O ∆G ≈ 30 KJ ATP patří mezi anhydridové makroergické sloučeniny. Vysoká hodnota energie této sloučeniny je dána elektrostatickými repulsemi mezi záporně nabitými kyslíkovými atomy.
 a anorganický fosfát (P, někdy též značeno jako Pi) za uvolnění energie: H2O + ATP → ADP + P ∆G ≈ -30 KJ. Přebytečná energie z látkového metabolismu naopak umožňuje vznik ATP: ADP + P → ATP + H2O ∆G ≈ 30 KJ. ATP patří mezi anhydridové makroergické sloučeniny. Vysoká hodnota energie této sloučeniny je dána elektrostatickými repulsemi mezi záporně nabitými kyslíkovými atomy.")
16
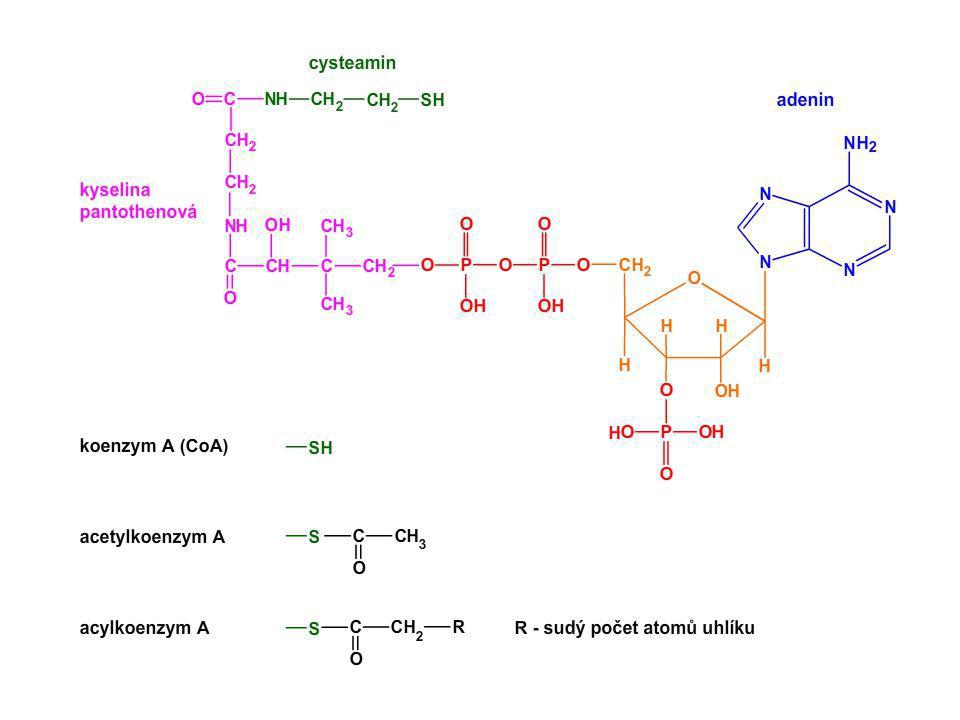
Trávení sacharidů Obrázek znázorňuje čemu podléhají polysacharidy (např. škrob) během procesu trávení. Konečnými produkty jsou jednotlivé monosacharidy (v tomto případě glukosové jednotky). Jednotlivé monosacharidy se dále mohou odbourávat, či sloužit jako stavební produkty. Maltasa = α-D-glukosidasa
 během procesu trávení. Konečnými produkty jsou jednotlivé monosacharidy (v tomto případě glukosové jednotky). Jednotlivé monosacharidy se dále mohou odbourávat, či sloužit jako stavební produkty. Maltasa = α-D-glukosidasa.")
17
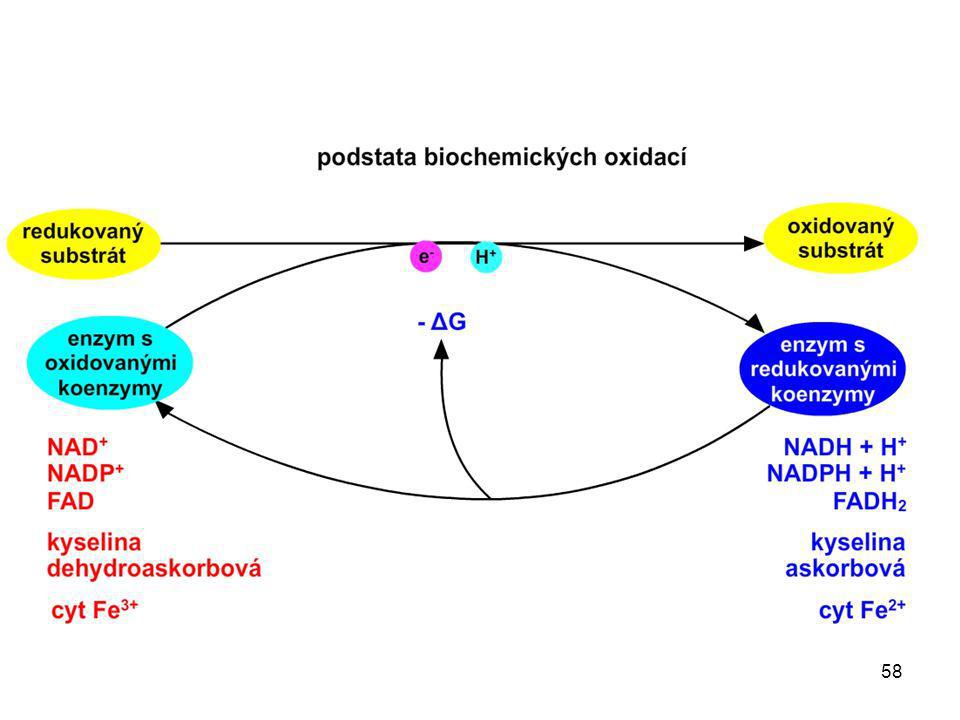
Trávení bílkovin Peptidový řetězec H3N+ COO- aminopeptidasa
karboxypeptidasa endopeptidasa endopeptidasa tripeptidasa Schéma znázorňuje, k čemu dochází během štěpení peptidového řetězce v procesu trávení, a jaké enzymy se na tomto štěpení podílejí. Konečnými produkty jsou jednotlivé aminokyseliny (na schématu jsou aminokyseliny zobrazeny jako barevné kuličky). Jednotlivé aminokyseliny se dále mohou odbourávat, či sloužit jako stavební látky. Rozdělení peptidas na jednotlivé skupiny (karboxypeptidasy, aminopeptidasy …) je možné na středních školách vynechat. dipeptidasa dipeptidasa dipeptidasa
. Jednotlivé aminokyseliny se dále mohou odbourávat, či sloužit jako stavební látky. Rozdělení peptidas na jednotlivé skupiny (karboxypeptidasy, aminopeptidasy …) je možné na středních školách vynechat. dipeptidasa. dipeptidasa. dipeptidasa.")
18
Trávení triacylglycerolů
mastné kyseliny (v tomto případě se jedná o kyselinu stearovou) triacylglycerol monoacylglycerol (diacylglycerol, resp. glycerol) Hydrolytické štěpení triacylglycerolů probíhá v cytoplasmě.
 triacylglycerol. monoacylglycerol. (diacylglycerol, resp. glycerol) Hydrolytické štěpení triacylglycerolů probíhá v cytoplasmě.")
19
Metabolismus sacharidů
Odbourávání monosacharidů probíhá v několika fázích: 1. Fáze glykolýzy 2. Fáze glykolýzy 3. Fáze glykolýzy Kliknutím na jednotlivé fáze glykolýzy přejdete na příslušné snímky. Jednotlivé kroky glykolýzy je možné z učiva středoškolské biochemie vynechat (snímek č. 25 – 27). Glykolýza probíhá v cytoplasmě. 4. Aerobní a anaerobní odbourávání pyruvátu
. Glykolýza probíhá v cytoplasmě. 4. Aerobní a anaerobní odbourávání pyruvátu.")
20
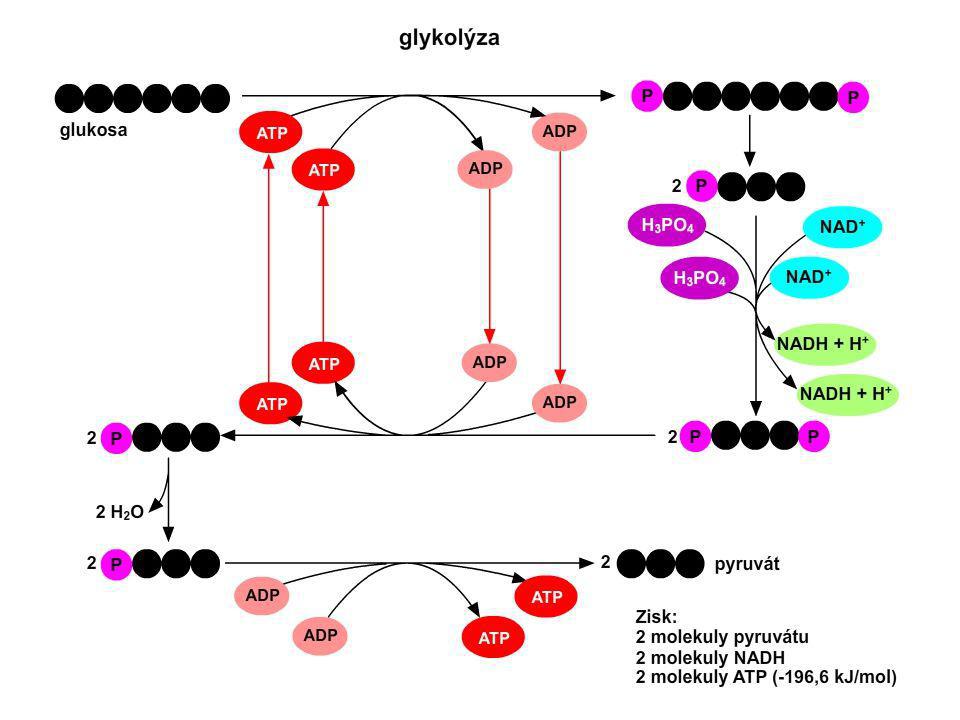
2x glyceraldehyd-3-fosfát
sacharidy Glykolýza - schéma trávení amylasy glukosa 1. fáze glykolýzy ATP ADP glukosa-6-fosfát isomerace fruktosa-6-fosfát 2. fáze glykolýzy ATP ADP fruktosa-1,6-bisfosfát 2x glyceraldehyd-3-fosfát 2x (NAD++P) 2x (NADH+H+) 3. fáze glykolýzy 2x 1,3-bisfosfoglycerát 2x ADP 2x ATP Průběh 1. fáze glykolýzy: A) Výchozí sloučeninou je glykogen: glykogen + H3PO4 → glukosa-1-fosfát enzym: fosforylasa glukosa-1-fosfát → glukosa-6-fosfát enzym: fosfoglukomutasa glukosa-6-fosfát → fruktosa-6-fosfát enzym: glukosafosfátisomerasa B) Výchozí sloučeninou je glukosa: glukosa + ATP → glukosa-6-fosfát + ADP enzym: hexokinasa glukosa-6-fosfát → fruktosa-6-fosfát enzym: glukosafosfátisomerasa Průběh 2. fáze glykolýzy: fruktosa-6-fosfát + ATP → fruktosa-1,6-bisfosfát + ADP enzym: fosfofruktokinasa fruktosa-1,6-bisfosfát → 3-fosfoglyceraldehyd + dihydroxyacetonfosfát enzym: fruktosabisfosfátaldolasa dihydroxyacetonfosfát ↔ 3-fosfoglyceraldehyd enzym: triosafosfátisomerasa Průběh 3. fáze glykolýzy: 3-fosfoglyceraldehyd + NAD+ + H3PO4 → 1,3-bisfosfoglycerát + NADH + H+ enzym: HS-enzym (glyceraldehyd-3-fosfátdehydrogenasa), dehydrogenace 1,3-bisfosfoglycerát + ADP → 3-fosfoglycerát + ATP enzym: fosfoglycerátkinasa, substrátová fosforylace 3-fosfoglycerát → 2-fosfoglycerát enzym: fosfoglycerátfosfomutasa (koenzym: 2,3-bisfosfoglycerát) 2-fosfoglycerát → fosfoenolpyruvát + H2O enzym: enolasa fosfoenolpyruvát + ADP → enolpyruvát + ATP enzym: pyruvátkinasa enolpyruvát ↔ pyruvát 2x 3-fosfoglycerát Celkový zisk: 2 molekuly ATP 2 molekuly NADH 2x 2-fosfoglycerát 2x H2O 2x fosfoenolpyruvát 2x ADP 2x ATP 2x pyruvát
 2x (NADH+H+) 3. fáze glykolýzy. 2x 1,3-bisfosfoglycerát. 2x ADP. 2x ATP. Průběh 1. fáze glykolýzy: A) Výchozí sloučeninou je glykogen: glykogen + H3PO4 → glukosa-1-fosfát enzym: fosforylasa. glukosa-1-fosfát → glukosa-6-fosfát enzym: fosfoglukomutasa. glukosa-6-fosfát → fruktosa-6-fosfát enzym: glukosafosfátisomerasa. B) Výchozí sloučeninou je glukosa: glukosa + ATP → glukosa-6-fosfát + ADP enzym: hexokinasa. glukosa-6-fosfát → fruktosa-6-fosfát enzym: glukosafosfátisomerasa. Průběh 2. fáze glykolýzy: fruktosa-6-fosfát + ATP → fruktosa-1,6-bisfosfát + ADP enzym: fosfofruktokinasa. fruktosa-1,6-bisfosfát → 3-fosfoglyceraldehyd + dihydroxyacetonfosfát enzym: fruktosabisfosfátaldolasa. dihydroxyacetonfosfát ↔ 3-fosfoglyceraldehyd enzym: triosafosfátisomerasa. Průběh 3. fáze glykolýzy: 3-fosfoglyceraldehyd + NAD+ + H3PO4 → 1,3-bisfosfoglycerát + NADH + H+ enzym: HS-enzym (glyceraldehyd-3-fosfátdehydrogenasa), dehydrogenace. 1,3-bisfosfoglycerát + ADP → 3-fosfoglycerát + ATP enzym: fosfoglycerátkinasa, substrátová fosforylace. 3-fosfoglycerát → 2-fosfoglycerát enzym: fosfoglycerátfosfomutasa (koenzym: 2,3-bisfosfoglycerát) 2-fosfoglycerát → fosfoenolpyruvát + H2O enzym: enolasa. fosfoenolpyruvát + ADP → enolpyruvát + ATP enzym: pyruvátkinasa. enolpyruvát ↔ pyruvát. 2x 3-fosfoglycerát. Celkový zisk: 2 molekuly ATP. 2 molekuly NADH. 2x 2-fosfoglycerát. 2x H2O. 2x fosfoenolpyruvát. 2x ADP. 2x ATP. 2x pyruvát.")
22
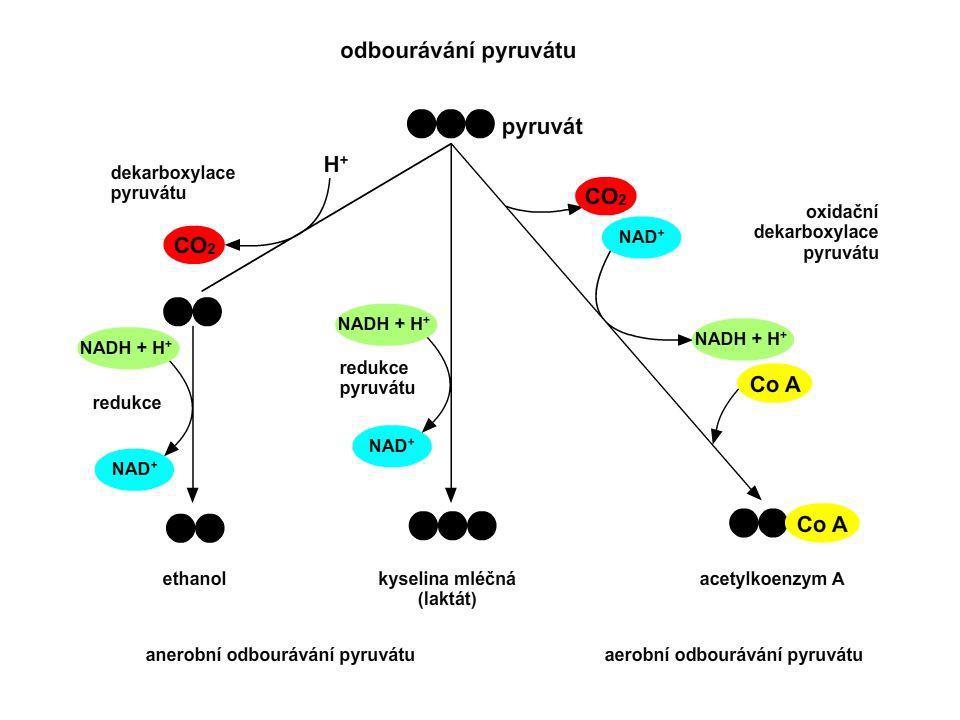
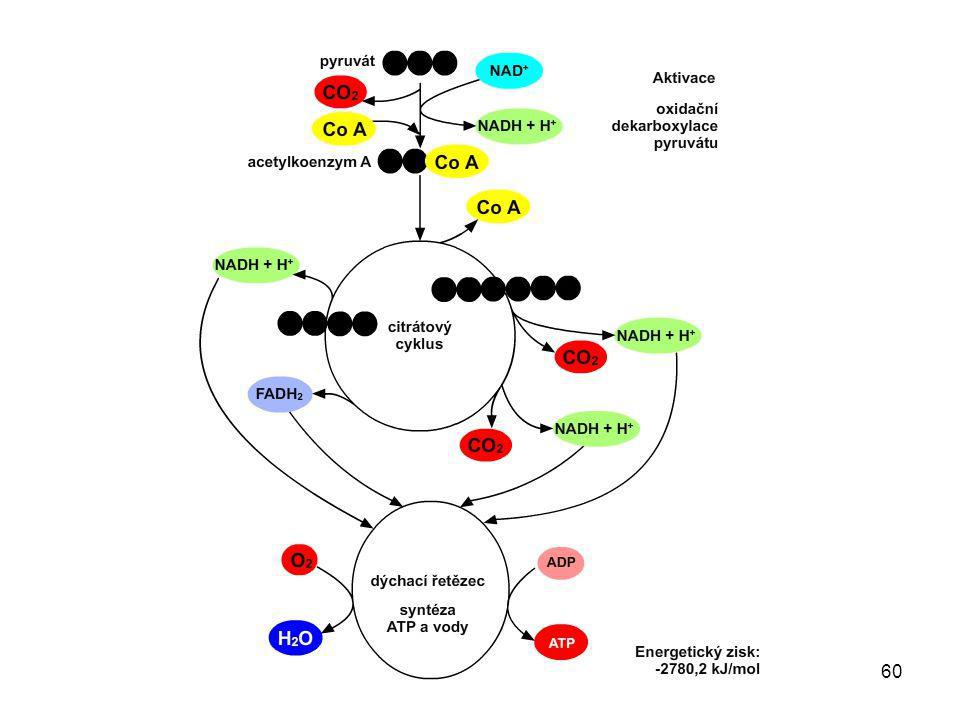
Aerobní odbourávání pyruvátu
1. Aerobní odbourávání pyruvátu (oxidační dekarboxylace pyruvátu) acetylkoenzym A Aerobní odbourávání pyruvátu probíhá v mitochondriích. Acetylkoenzym A vstupuje dále do citrátového cyklu, kde dochází k jeho postupnému odbourávání. Redukovaný koenzym (NADH) přechází do dýchacího řetězce, kde se oxiduje, čímž vzniká energie.
 acetylkoenzym A. Aerobní odbourávání pyruvátu probíhá v mitochondriích. Acetylkoenzym A vstupuje dále do citrátového cyklu, kde dochází k jeho postupnému odbourávání. Redukovaný koenzym (NADH) přechází do dýchacího řetězce, kde se oxiduje, čímž vzniká energie.")
24
Anaerobní odbourávání pyruvátu
2A Tvorba laktátu laktát 2B Alkoholové (ethanolové) kvašení Při cvičení je náš metabolismus mnohem rychlejší, než když jsme v klidu. Tím se kyslík mnohem rychleji spotřebovává a vzniká tak „kyslíkový dluh“. Redukovaný koenzym NADH nemůže být oxidován v dýchacím řetězci. Aby nedocházelo k jeho hromadění, začne se glukosa odbourávat anaerobně. Tímto náhradním procesem dochází k doplňování potřebné energie. Pyruvát se redukuje na laktát za spoluúčasti enzymu laktátdehydrogenasy (LDH).
 kvašení. Při cvičení je náš metabolismus mnohem rychlejší, než když jsme v klidu. Tím se kyslík mnohem rychleji spotřebovává a vzniká tak „kyslíkový dluh . Redukovaný koenzym NADH nemůže být oxidován v dýchacím řetězci. Aby nedocházelo k jeho hromadění, začne se glukosa odbourávat anaerobně. Tímto náhradním procesem dochází k doplňování potřebné energie. Pyruvát se redukuje na laktát za spoluúčasti enzymu laktátdehydrogenasy (LDH).")
26
Aerobní a anaerobní odbourávání - schéma
pyruvát aerobní odbourávání anaerobní odbourávání acetylkoenzym A tvorba laktátu alkoholové kvašení citrátový cyklus + dýchací řetězec CO2, H2O + energie laktát ethanol

27
Metabolismus triacylglycerolů

28
Odbourávání mastných kyselin
Aktivace mastných kyselin koenzym A karboxylová kyselina aktivovaná karboxylová kyselina 1. Fáze β-oxidace enoylkoenzym A 2. Fáze β-oxidace Jednotlivé kroky odbourávání mastných kyselin je možné z učiva středoškolské biochemie vynechat (snímek č. 38 – 41). Důležité jsou tučně zvýrazněné poznámky uvedené na snímku č. 39. Účast enzymů na odbourávání mastné kyseliny: Přeměnu aktivované mastné kyseliny na enoylkoenzym A katalyzuje enzym acylkoenzym-A-dehydrogenasa. Přeměnu enoylkoenzymuA na hydroxyacylkoenzym A katalyzuje enzym enoylkoenzym-A-hydrasa. Dehydrogenaci hydroxyacylkoenzymu A na oxoacylkoenzym A (ketoacylkoenzym A) katalyzuje enzym β-hydroxyacylkoenzym-A-dehydrogenasa. Thiolytické štěpení při kterém vzniká acetylkoenzym A katalyzuje enzym β-ketokinasa. β-oxidace probíhá v matrix mitochondrií, aktivace mastných kyselin probíhá v cytoplasmě. (Zdroj: Sofrová, D. a kol: Biochemie – Základní kurz. Praha: Karolinum, s. 176) hydroxyacylkoenzym A oxoacylkoenzym A
. Důležité jsou tučně zvýrazněné poznámky uvedené na snímku č. 39. Účast enzymů na odbourávání mastné kyseliny: Přeměnu aktivované mastné kyseliny na enoylkoenzym A katalyzuje enzym acylkoenzym-A-dehydrogenasa. Přeměnu enoylkoenzymuA na hydroxyacylkoenzym A katalyzuje enzym enoylkoenzym-A-hydrasa. Dehydrogenaci hydroxyacylkoenzymu A na oxoacylkoenzym A (ketoacylkoenzym A) katalyzuje enzym β-hydroxyacylkoenzym-A-dehydrogenasa. Thiolytické štěpení při kterém vzniká acetylkoenzym A katalyzuje enzym β-ketokinasa. β-oxidace probíhá v matrix mitochondrií, aktivace mastných kyselin probíhá v cytoplasmě. (Zdroj: Sofrová, D. a kol: Biochemie – Základní kurz. Praha: Karolinum, s. 176) hydroxyacylkoenzym A. oxoacylkoenzym A.")
29
Odbourávání mastných kyselin
3. Fáze β-oxidace acetylkoenzym A aktivovaná mastná kyselina zkrácená o dva uhlíky Původní řetězec karboxylové kyseliny se vždy zkracuje o dva uhlíky. Jednotlivé kroky odbourávání mastných kyselin je možné z učiva středoškolské biochemie vynechat (snímek č. 38 – 41). Důležité jsou tučně zvýrazněné poznámky uvedené na snímku č. 39. Účast enzymů na odbourávání mastné kyseliny: Přeměnu aktivované mastné kyseliny na enoylkoenzym A katalyzuje enzym acylkoenzym-A-dehydrogenasa. Přeměnu enoylkoenzymuA na hydroxyacylkoenzym A katalyzuje enzym enoylkoenzym-A-hydrasa. Dehydrogenaci hydroxyacylkoenzymu A na oxoacylkoenzym A (ketoacylkoenzym A) katalyzuje enzym β-hydroxyacylkoenzym-A-dehydrogenasa. Thiolytické štěpení při kterém vzniká acetylkoenzym A katalyzuje enzym β-ketokinasa. β-oxidace probíhá v matrix mitochondrií, aktivace mastných kyselin probíhá v cytoplasmě.
. Důležité jsou tučně zvýrazněné poznámky uvedené na snímku č. 39. Účast enzymů na odbourávání mastné kyseliny: Přeměnu aktivované mastné kyseliny na enoylkoenzym A katalyzuje enzym acylkoenzym-A-dehydrogenasa. Přeměnu enoylkoenzymuA na hydroxyacylkoenzym A katalyzuje enzym enoylkoenzym-A-hydrasa. Dehydrogenaci hydroxyacylkoenzymu A na oxoacylkoenzym A (ketoacylkoenzym A) katalyzuje enzym β-hydroxyacylkoenzym-A-dehydrogenasa. Thiolytické štěpení při kterém vzniká acetylkoenzym A katalyzuje enzym β-ketokinasa. β-oxidace probíhá v matrix mitochondrií, aktivace mastných kyselin probíhá v cytoplasmě.")
30
aktivace mastných kyselin FAD dehydrogenace aktivovaná mastná kyselina
triacylglycerol karboxylová kyselina trávení glycerol glykolýza ATP + CoA-SH aktivace mastných kyselin FAD dehydrogenace aktivovaná mastná kyselina AMP FADH2 H2O hydroxyacylCoA enoylCoA enoylCoA FADH2 NAD+ H2O hydratace NADH + H+ oxoacylCoA hydroxyacylCoA FAD NAD+ dehydrogenace aktivovaná mastná kyselina zkrácená o dva uhlíky NADH + H+ oxoacylCoA CoA-SH acetylCoA citrátový cyklus + dýchací řetězec CO2,H2O + energie

31
aktivace mastných kyselin FAD 1. fáze ß-oxidace
triacylglycerol karboxylová kyselina trávení glycerol glykolýza ATP + CoA-SH aktivace mastných kyselin FAD 1. fáze ß-oxidace aktivovaná mastná kyselina ADP FADH2 H2O hydroxyacylCoA enoylCoA enoylCoA FADH2 NAD+ H2O 2. fáze ß-oxidace NADH + H+ 1. fáze ß-oxidace oxoacylCoA hydroxyacylCoA FAD NAD+ 2. fáze ß-oxidace aktivovaná mastná kyselina zkrácená o dva uhlíky NADH + H+ oxoacylCoA CoA-SH acetylCoA citrátový cyklus + dýchací řetězec 3. fáze ß-oxidace CO2,H2O + energie

32
Rozdíly v energetickém výtěžku
Zisk (počet molekul) Energie uvolněná ATP NADH FADH2 při úplné oxidaci 1 molekuly glukosy 4 10 2 při anerobním odbourávání 1 molekuly glukosy při rozštěpení 1 molekuly kyseliny palmitové 8-2 31 15 Při anaerobním odbourávání jedné molekuly glukosy se uvolní nejméně energie, naopak při úplném rozkladu jedné molekuly kyseliny palmitové se uvolní energie nejvíce. Množství uvolněných molekul ATP se v odborné literatuře liší. ODBOURÁVÁNÍ SACHARIDŮ: Při odbourávání 1 molekuly glukosy na 2 molekuly pyruvátu vznikají v průběhu glykolýzy 2 molekuly ATP a 2 molekuly NADH. Pyruvát se může dále odbourávat buď aerobně nebo anaerobně: Aerobní odbourávání pyruvátu: Pyruvát se může dále odbourávat aerobně na acetylkoenzym A za vzniku 1 molekuly NADH (ze 2 molekul pyruvátu vznikají 2 molekuly NADH). Acetylkoenzym A vstupuje do citrátového cyklu, kde dále dochází ke vzniku 1 molekuly GTP (resp. ATP), 3 molekul NADH a 1 molekuly FADH2 (tj. ze 2 molekul acetylkoenzymu A vznikají 2 molekuly ATP, 6 molekul NADH a 2 molekuly FADH2). Anaerobní odbourávání pyruvátu: Pyruvát se odbourává na laktát za současné spotřeby 1 molekuly NADH (tj. na 2 molekuly pyruvátu spotřebujeme 2 molekuly NADH). ODBOURÁVÁNÍ TRIACYLGLYCEROLŮ: Odbourávání kyseliny palmitové: Při úplném rozštěpení 1 molekuly kyseliny palmitové vzniká 7 molekul FADH2, 7 molekul NADH a 8 molekul acetylkoenzymu A (z 8 molekul acetylkoenzymu A se v citrátovém cyklu získá celkem 8 molekul ATP, 24 molekul NADH a 8 molekul FADH2). 1 molekula ATP se spotřebuje (odpovídá spíše 2 molekulám ATP – ATP se rozkládá až na AMP) při vzniku acylkoenzymu A (aktivace mastných kyselin).
 Energie uvolněná. ATP. NADH. FADH2. při úplné oxidaci 1 molekuly glukosy při anerobním odbourávání 1 molekuly glukosy. při rozštěpení 1 molekuly kyseliny palmitové Při anaerobním odbourávání jedné molekuly glukosy se uvolní nejméně energie, naopak při úplném rozkladu jedné molekuly kyseliny palmitové se uvolní energie nejvíce. Množství uvolněných molekul ATP se v odborné literatuře liší. ODBOURÁVÁNÍ SACHARIDŮ: Při odbourávání 1 molekuly glukosy na 2 molekuly pyruvátu vznikají v průběhu glykolýzy 2 molekuly ATP a 2 molekuly NADH. Pyruvát se může dále odbourávat buď aerobně nebo anaerobně: Aerobní odbourávání pyruvátu: Pyruvát se může dále odbourávat aerobně na acetylkoenzym A za vzniku 1 molekuly NADH (ze 2 molekul pyruvátu vznikají 2 molekuly NADH). Acetylkoenzym A vstupuje do citrátového cyklu, kde dále dochází ke vzniku 1 molekuly GTP (resp. ATP), 3 molekul NADH a 1 molekuly FADH2 (tj. ze 2 molekul acetylkoenzymu A vznikají 2 molekuly ATP, 6 molekul NADH a 2 molekuly FADH2). Anaerobní odbourávání pyruvátu: Pyruvát se odbourává na laktát za současné spotřeby 1 molekuly NADH (tj. na 2 molekuly pyruvátu spotřebujeme 2 molekuly NADH). ODBOURÁVÁNÍ TRIACYLGLYCEROLŮ: Odbourávání kyseliny palmitové: Při úplném rozštěpení 1 molekuly kyseliny palmitové vzniká 7 molekul FADH2, 7 molekul NADH a 8 molekul acetylkoenzymu A (z 8 molekul acetylkoenzymu A se v citrátovém cyklu získá celkem 8 molekul ATP, 24 molekul NADH a 8 molekul FADH2). 1 molekula ATP se spotřebuje (odpovídá spíše 2 molekulám ATP – ATP se rozkládá až na AMP) při vzniku acylkoenzymu A (aktivace mastných kyselin).")
33
Vztah metabolismu triacylglycerolů a sacharidů
potrava tuky sacharidy mastné kyseliny + glycerol monosacharidy pyruvát laktát acetylkoenzym A citrátový cyklus + dýchací řetězec CO2,H2O, energie

34
Citrátový cyklus

35
Jednotlivé kroky citrátového cyklu je možné z učiva středoškolské biochemie vynechat.
Odbouráním 1 acetylového zbytku (do citrátového cyklu přichází vázaný na koenzym A) se přímo získá 1 molekula GTP (GTP se dále přeměňuje na ATP), 3 molekuly NADH a 1 molekula FADH2, dále vznikají 2 molekuly oxidu uhličitého a spotřebuje se 1 molekula vody.
 se přímo získá 1 molekula GTP (GTP se dále přeměňuje na ATP), 3 molekuly NADH a 1 molekula FADH2, dále vznikají 2 molekuly oxidu uhličitého a spotřebuje se 1 molekula vody.")
36
Citrátový cyklus – spustit animaci
Acetylkoenzym A Citrát S-CoA S-CoA Oxalacetát Isocitrát H H 2-oxoglutarát Dýchací řetězec H H Malát H H Černé kuličky znázorňují počet atomů uhlíku v molekule, červené kuličky znázorňují atomy kyslíku a bílé kuličky atomy vodíku. Animace se spustí po kliknutí na animační tlačítko „Citrátový cyklus – spustit animaci“. GTP Fumarát Sukcinylkoenzym A H H Sukcinát

37
Respirace (dýchání) Během dýchání dochází k oxidaci živin, a tím vzniká energie potřebná pro pohon řady biochemických procesů. O2 CO2

38
Vnější a vnitřní respirace
Dýchání (respiraci) můžeme rozdělit na dvě rozdílné části jednoho procesu: Vnitřní respirace Vnější respirace Probíhá v buňkách. Složky potravy jsou odbourávány řadou chemických reakcí, z nichž většina potřebuje kyslík. Tyto reakce uvolňují energii. Dodává kyslík buňkám, a tím umožňuje vnitřní respiraci.
 můžeme rozdělit na dvě rozdílné části jednoho procesu: Vnitřní respirace. Vnější respirace. Probíhá v buňkách. Složky potravy jsou odbourávány řadou chemických reakcí, z nichž většina potřebuje kyslík. Tyto reakce uvolňují energii. Dodává kyslík buňkám, a tím umožňuje vnitřní respiraci.")
39
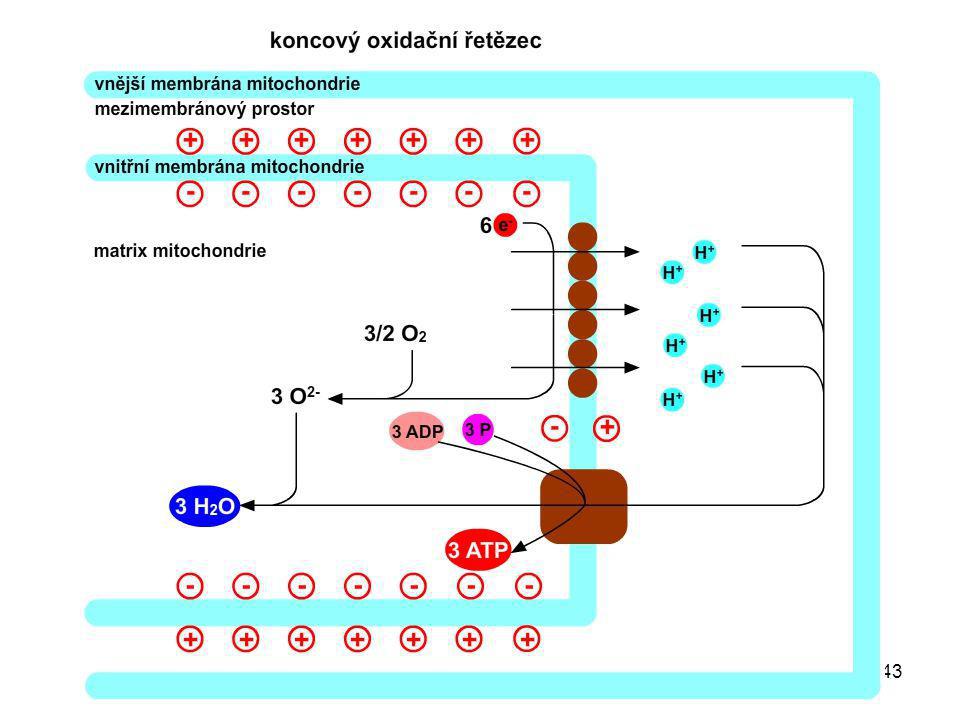
Mitochondrie Dýchací řetězec je lokalizován ve vnitřní mitochondriální membráně. Citrátový cyklus probíhá v matrix mitochondrie. Matrix Vnitřní membrána Mezimembránový prostor Vnější membrána

40
4H+(z matrix) + 4e- + O2 → 2H2O + energie
Dýchací řetězec Dýchací řetězec je souborem reakcí, které ukončují energetické odbourávání monosacharidů, mastných kyselin, glycerolu a aminokyselin. V dýchacím řetězci probíhá oxidace redukovaných koenzymů: NADH + H+ → NAD+ + 2H+ + 2e- FADH2 → FAD + 2H+ + 2e- Protony jsou přenášeny do mezimembránového prostoru. Elektrony jsou přenášeny po sérii akceptorů na elementární kyslík za vzniku vody. 4H+(z matrix) + 4e- + O2 → 2H2O + energie
 + 4e- + O2 → 2H2O + energie.")
41
Dýchací řetězec Dýchací řetězec se skládá ze 4 enzymových komplexů a z enzymu ATP-synthasy. Během přenosu elektronů vzniká energie, kterou využívá každý enzymový komplex k čerpání protonů z matrix do mezimembránového prostoru Při přechodu protonů zpět do matrix vzniká energie, která pohání syntézu ATP z ADP a anorganického fosfátu. Protony roztáčí stopku hlavy ATP-synthasy, a tím dochází k syntéze ATP.

42
Schéma energetického metabolismu v mitochondriích
Vnější mitochondriální membrána Vnitřní mitochondriální membrána H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ H+ ATP - synthasa H+ 2H2O e- H+ H+ H+ O2 ATP Dýchací řetězec 3NAD+, FAD ADP + P 3NADH; FADH2 H+ Citrátový cyklus O2 2CO2 2CO2 Pyruvát (pyruvát je konečným produktem glykolýzy) a mastné kyseliny (ve formě acylkarnitinu) vstupují přes mitochondriální membrány do matrix mitochondrie. V matrix dochází k odbourání na acetylkoenzym A, který vstupuje do citrátového cyklu, kde se váže na kyselinu oxaloctovou za vzniku kyseliny citronové. V citrátovem cyklu dochází ke vzniku 2 molekul oxidu uhličitého, 3 molekul NADH, 1 molekuly FADH2 a 1 molekuly GTP (GTP se přeměňuje na ATP). NADH a FADH2 vstupují dále do dýchacího řetězce, kde se oxidují na NAD+ a FAD. Transport elektronů umožňuje vytvářet elektrochemický protonový gradient – protony jsou přenášeny do mezimembránového prostoru, což způsobí, že koncentrace protonů je v mezimembránovém prostoru vyšší než v matrix (pH v mezimembránovém prostoru je vyšší než pH matrix). Během přenosu protonů zpět do matrix dochází k uvolnění energie, která je využita pro tvorbu ATP z ADP a anorganického fosfátu. Elektrony jsou nakonec přeneseny na kyslík za vzniku vody. Jedná se o kyslík, který dýcháme. Matrix Acetyl-CoA Pyruvát Mastné kyseliny Cytosol (Pomocí přenašeče) Pyruvát Mastné kyseliny
 a mastné kyseliny (ve formě acylkarnitinu) vstupují přes mitochondriální membrány do matrix mitochondrie. V matrix dochází k odbourání na acetylkoenzym A, který vstupuje do citrátového cyklu, kde se váže na kyselinu oxaloctovou za vzniku kyseliny citronové. V citrátovem cyklu dochází ke vzniku 2 molekul oxidu uhličitého, 3 molekul NADH, 1 molekuly FADH2 a 1 molekuly GTP (GTP se přeměňuje na ATP). NADH a FADH2 vstupují dále do dýchacího řetězce, kde se oxidují na NAD+ a FAD. Transport elektronů umožňuje vytvářet elektrochemický protonový gradient – protony jsou přenášeny do mezimembránového prostoru, což způsobí, že koncentrace protonů je v mezimembránovém prostoru vyšší než v matrix (pH v mezimembránovém prostoru je vyšší než pH matrix). Během přenosu protonů zpět do matrix dochází k uvolnění energie, která je využita pro tvorbu ATP z ADP a anorganického fosfátu. Elektrony jsou nakonec přeneseny na kyslík za vzniku vody. Jedná se o kyslík, který dýcháme. Matrix. Acetyl-CoA. Pyruvát. Mastné kyseliny. Cytosol. (Pomocí přenašeče) Pyruvát. Mastné kyseliny.")
44
Enzymové komplexy (zjednodušené schema)
I NADH-CoQ oxidoreduktasa II Sukcinát-CoQ oxidoreduktasa III CoQH2– cytochrom c oxidoreduktasa IV Cytochrom c oxidasa V ATP-synthasa 2H+ H+ 4H+ 4H+ Cyt c Cyt c I 2e- III IV V Koenzym Q 2e- 4e- 2H+ II 2e- O2 + 2H+ 2e- Tento snímek je možné při výuce středoškolské biochemie vynechat či zařadit do semináře. Dýchací řetězec se skládá ze 4 enzymových komplexů a z enzymu ATP-synthasy. Ubichinony (koenzym Q, CoQ) a cytochromy (cyt) slouží jako pohyblivé přenašeče elektronů (koenzym Q i protonů). Během přenosu elektronů vzniká energie, kterou využívá každý enzymový komplex k čerpání protonů (protony pocházejí z vody H2O ↔ H+ + OH-) z matrix do mezimembránového prostoru – vytváří se elektrochemický protonový gradient (koncentrace protonů je v mezimembránovém prostoru daleko vyšší než v matrix). Při přechodu protonů zpět do matrix vzniká energie, která pohání syntézu ATP z ADP a anorganického fosfátu (jedná se o oxidační fosforylaci). Protony roztáčí stopku hlavy ATP-synthasy, a tím dochází k syntéze ATP. Takto popsaný vznik molekuly ATP popisuje tzv. chemiosmotická teorie, za kterou dostal v roce 1978 Peter Mitchell Nobelovu cenu. Viz též snímek č. 20. 4H+ 2H+ 4H+ 2H2O NAD+ FADH2 ADP + P NADH + H+ FAD Matrix H+ ATP
 a cytochromy (cyt) slouží jako pohyblivé přenašeče elektronů (koenzym Q i protonů). Během přenosu elektronů vzniká energie, kterou využívá každý enzymový komplex k čerpání protonů (protony pocházejí z vody H2O ↔ H+ + OH-) z matrix do mezimembránového prostoru – vytváří se elektrochemický protonový gradient (koncentrace protonů je v mezimembránovém prostoru daleko vyšší než v matrix). Při přechodu protonů zpět do matrix vzniká energie, která pohání syntézu ATP z ADP a anorganického fosfátu (jedná se o oxidační fosforylaci). Protony roztáčí stopku hlavy ATP-synthasy, a tím dochází k syntéze ATP. Takto popsaný vznik molekuly ATP popisuje tzv. chemiosmotická teorie, za kterou dostal v roce 1978 Peter Mitchell Nobelovu cenu. Viz též snímek č H+ 2H+ 4H+ 2H2O. NAD+ FADH2. ADP + P. NADH + H+ FAD. Matrix. H+ ATP.")
45
Enzymové komplexy: Komplex I
Komplex I: NADH-CoQ-oxidoreduktasa NADH + H+ + CoQ → NAD+ + CoQH2 2H+ 4H+ I 2Fe2+ 2Fe3+ 2CoQH2 (2e-,2H+) 2Fe3+ 2Fe2+ 2e- 2e- 2CoQH. (1e-,1H+) FMNH2 (2H+,2e-) FMN Matrix 2H+ 2H+ Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Komplex I je NADH-CoQ-oxidoreduktasa, která katalyzuje reakci: NADH + H+ + CoQ → NAD+ + CoQH2. V komplexu I dochází k přenosu dvou protonů a dvou elektronů z koenzymu NADH + H+ na prostetickou skupinu FMN (flavinmononukleotid) za vzniku FMNH2. Protony jsou dále uvolňovány do mezimembránového prostoru a elektrony (po jednom) přecházejí na další prostetickou skupinu typu FeS (kde dochází ke změně oxidačního čísla mezi Fe2+ a Fe3+). Elektrony (opět po jednom) jsou poté společně s dalšími protony z matrix přebírány semichinoidní formou ubichinonu CoQH• (ubisemichinon, mající k dispozici 1 e- a 1 H+) za vzniku ubichinolu CoQH2 (mající již 2 e-, 2 H+) – viz dále komplex III. NAD+ NADH + H+
 2Fe3+ 2Fe2+ 2e- 2e- 2CoQH. (1e-,1H+) FMNH2. (2H+,2e-) FMN. Matrix. 2H+ 2H+ Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Komplex I je NADH-CoQ-oxidoreduktasa, která katalyzuje reakci: NADH + H+ + CoQ → NAD+ + CoQH2. V komplexu I dochází k přenosu dvou protonů a dvou elektronů z koenzymu NADH + H+ na prostetickou skupinu FMN (flavinmononukleotid) za vzniku FMNH2. Protony jsou dále uvolňovány do mezimembránového prostoru a elektrony (po jednom) přecházejí na další prostetickou skupinu typu FeS (kde dochází ke změně oxidačního čísla mezi Fe2+ a Fe3+). Elektrony (opět po jednom) jsou poté společně s dalšími protony z matrix přebírány semichinoidní formou ubichinonu CoQH• (ubisemichinon, mající k dispozici 1 e- a 1 H+) za vzniku ubichinolu CoQH2 (mající již 2 e-, 2 H+) – viz dále komplex III. NAD+ NADH + H+")
46
Enzymové komplexy: Komplex II
Komplex II: Sukcinát-CoQ-oxidoreduktasa FADH2 + CoQ → FAD + CoQH2 4H+ II 2CoQH2 (2e-,2H+) 2CoQH. (1e-,1H+) 2e- FADH2 (2H+,2e-) FAD Matrix 2H+ Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Komplex II je Sukcinát-CoQ-oxidoreduktasa, která katalyzuje reakci: FADH2 + CoQ → FAD + CoQH2. Komplex II je flavoprotein (obsahuje prostetickou skupinu FAD a dvě skupiny typu Fe2S2). V komplexu II dochází k přenosu dvou protonů a dvou elektronů ze sukcinátu na prostetickou skupinu FAD za vzniku FADH2 a fumarátu. Protony nejsou ihned uvolňovány do mezimembránového prostoru, jak tomu bylo v komplexu I, ale společně s elektrony přecházejí na semichinoidní formu ubichinonu CoQH• (mající 1 e- a 1 H+) za vzniku ubichinolu CoQH2 (mající již 2 e-, 2 H+) – viz dále komplex III. Sukcinát -OOC-CH2-CH2-COO- Fumarát -OOC-CH=CH-COO-
 2CoQH. (1e-,1H+) 2e- FADH2. (2H+,2e-) FAD. Matrix. 2H+ Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Komplex II je Sukcinát-CoQ-oxidoreduktasa, která katalyzuje reakci: FADH2 + CoQ → FAD + CoQH2. Komplex II je flavoprotein (obsahuje prostetickou skupinu FAD a dvě skupiny typu Fe2S2). V komplexu II dochází k přenosu dvou protonů a dvou elektronů ze sukcinátu na prostetickou skupinu FAD za vzniku FADH2 a fumarátu. Protony nejsou ihned uvolňovány do mezimembránového prostoru, jak tomu bylo v komplexu I, ale společně s elektrony přecházejí na semichinoidní formu ubichinonu CoQH• (mající 1 e- a 1 H+) za vzniku ubichinolu CoQH2 (mající již 2 e-, 2 H+) – viz dále komplex III. Sukcinát. -OOC-CH2-CH2-COO- Fumarát. -OOC-CH=CH-COO-")
47
Enzymové komplexy: Komplex III
Komplex III: CoQH2–cytochrom-c-oxidoreduktasa CoQH2 + 2 cyt c (Fe3+) → CoQ + 2 cyt c (Fe2+) + 2 H+ 2H+ Cyt c Cyt c Cyt c III IV Cyt b e- Elektron z komplexu I nebo II CoQH. (1e-,1H+) e- e- CoQH2 (2e-,2H+) e- H+ Matrix H+ Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Komplex III je CoQH2–cytochrom-c-oxidoreduktasa, která katalyzuje reakci: CoQH2 + cyt c (Fe3+) → CoQ + cyt c (Fe2+) + 2 H+. Komplex III reoxiduje ubichinol (CoQH2) – produkt komplexu I a II. Ubichinol je oxidován na semichinon (CoQH•) tak, že jeden elektron přechází na cyt b a uvolní se jeden proton do mezimembránového prostoru, druhý elektron se přenese na cyt c (Fe3+ se redukuje na Fe2+) a opět se uvolní jeden proton do mezimembránového prostoru za vzniku ubichinonu (CoQ, k dispozici 0 e- a 0 H+). Na ubichinon (CoQ) se zpátky přenese elektron z cyt b a zároveň se naváže jeden proton z matrix mitochondrie za vzniku ubisemichinonu (CoQH•, mající k dispozici opět 1 e- a 1 H+). Ubisemichinon opět váže elektron z komplexu I (resp. II) a proton za vzniku ubichinolu a celý cyklus se opakuje. Jeden elektron se vždy cyklicky vrací přes cyt b, což má za následek přenos celkem dvou protonů z matrix do mezimembránového prostoru při přenosu jednoho elekronu na cyt c. (detailněji – viz následující snímky, snímky č. 17 a 18). Koenzym Q
 → CoQ + 2 cyt c (Fe2+) + 2 H+ 2H+ Cyt c. Cyt c. Cyt c III. IV. Cyt b. e- Elektron. z komplexu I. nebo II. CoQH. (1e-,1H+) e- e- CoQH2. (2e-,2H+) e H+ Matrix. H+ Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Komplex III je CoQH2–cytochrom-c-oxidoreduktasa, která katalyzuje reakci: CoQH2 + cyt c (Fe3+) → CoQ + cyt c (Fe2+) + 2 H+. Komplex III reoxiduje ubichinol (CoQH2) – produkt komplexu I a II. Ubichinol je oxidován na semichinon (CoQH•) tak, že jeden elektron přechází na cyt b a uvolní se jeden proton do mezimembránového prostoru, druhý elektron se přenese na cyt c (Fe3+ se redukuje na Fe2+) a opět se uvolní jeden proton do mezimembránového prostoru za vzniku ubichinonu (CoQ, k dispozici 0 e- a 0 H+). Na ubichinon (CoQ) se zpátky přenese elektron z cyt b a zároveň se naváže jeden proton z matrix mitochondrie za vzniku ubisemichinonu (CoQH•, mající k dispozici opět 1 e- a 1 H+). Ubisemichinon opět váže elektron z komplexu I (resp. II) a proton za vzniku ubichinolu a celý cyklus se opakuje. Jeden elektron se vždy cyklicky vrací přes cyt b, což má za následek přenos celkem dvou protonů z matrix do mezimembránového prostoru při přenosu jednoho elekronu na cyt c. (detailněji – viz následující snímky, snímky č. 17 a 18). Koenzym Q.")
48
Enzymové komplexy: Komplex III
Komplex III: CoQH2–cytochrom-c-oxidoreduktasa CoQH2 + 2 cyt c (Fe3+) → CoQ + 2 cyt c (Fe2+) + 2 H+ Cyt b Cyt c Elektron z komplexu I nebo II e- H+ CoQH. (1e-,1H+) CoQH2 (2e-,2H+) CoQ e- e- H+ e- H+ CoQH. (1e-,1H+) e- H+ e- e- H+ CoQH. (1e-,1H+) H+ CoQH. (1e-,1H+) e- Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Na tomto snímku je schematicky pomocí animace (pohyblivých kuliček znázorňujících protony a elektrony) znázorněn přenos elektronů a protonů pomocí koenzymu Q. Animace se spustí po kliknutí na animační tlačítko „Spustit animaci“. Matrix H+ H+ H+ H+ Koenzym Q H+ H+ H+ H+ Spustit animaci
 → CoQ + 2 cyt c (Fe2+) + 2 H Cyt b. Cyt c. Elektron. z komplexu I. nebo II. e- H+ CoQH. (1e-,1H+) CoQH2. (2e-,2H+) CoQ. e- e- H+ e- H+ CoQH. (1e-,1H+) e- H+ e- e- H+ CoQH. (1e-,1H+) H+ CoQH. (1e-,1H+) e Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Na tomto snímku je schematicky pomocí animace (pohyblivých kuliček znázorňujících protony a elektrony) znázorněn přenos elektronů a protonů pomocí koenzymu Q. Animace se spustí po kliknutí na animační tlačítko „Spustit animaci . Matrix. H+ H+ H+ H+ Koenzym Q. H+ H+ H+ H+ Spustit animaci.")
49
Enzymové komplexy: Komplex III
Komplex III: CoQH2–cytochrom-c-oxidoreduktasa CoQH2 + 2 cyt c (Fe3+) → CoQ + 2 cyt c (Fe2+) + 2 H+ H+ H+ Cyt c e- H+ e- Elektron z komplexu I nebo II e- CoQH2 (2e-,2H+) CoQH. (1e-,1H+) H+ H+ e- e- e- Cyt b CoQ H+ CoQH. (1e-,1H+) e- Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Na tomto snímku je schematicky pomocí šipek znázorněn přenos elektronů a protonů pomocí koenzymu Q. Obrázek znázorňuje průběh animace, která je k dispozici na předchozím snímku. H+ Matrix H+ Koenzym Q
 → CoQ + 2 cyt c (Fe2+) + 2 H+ H+ H Cyt c. e- H+ e- Elektron. z komplexu I. nebo II. e- CoQH2. (2e-,2H+) CoQH. (1e-,1H+) H+ H+ e- e- e- Cyt b. CoQ. H+ CoQH. (1e-,1H+) e Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Na tomto snímku je schematicky pomocí šipek znázorněn přenos elektronů a protonů pomocí koenzymu Q. Obrázek znázorňuje průběh animace, která je k dispozici na předchozím snímku. H+ Matrix. H+ Koenzym Q.")
50
Enzymové komplexy: Komplex IV
Komplex IV: Cytochrom-c-oxidasa 4 cyt c (Fe2+) H+ → 4 cyt c (Fe3+) + 2 H2O Cyt c Cyt c Cyt c III IV e- e- e- e- 4e- Cyt a Cyt a3 O2 + Matrix Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Komplex IV je cytochrom-c-oxidasa, která katalyzuje reakci: 4 cyt c (Fe2+) H+ → 4 cyt c (Fe3+) + 2 H2O. Elektrony z cytochromu c přebírá cytochrom a; odtud jsou přenášeny na cytochrom a3 (dochází ke změně oxidačního čísla mezi Cu2+ a Cu+). Cytochrom a3 postupně redukuje molekuly kyslíku na oxidové ionty (O2-), které s protony z matrix reagují za vzniku molekuly vody. Hlavní úloha kyslíku, jejž vdechujeme, spočívá tedy v tom, aby udržoval komplex cytochromů v oxidovaném stavu. 4H+ 2 H2O
 H+ → 4 cyt c (Fe3+) + 2 H2O. Cyt c. Cyt c. Cyt c III. IV. e- e- e- e- 4e- Cyt a Cyt a3. O2. + Matrix. Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Komplex IV je cytochrom-c-oxidasa, která katalyzuje reakci: 4 cyt c (Fe2+) H+ → 4 cyt c (Fe3+) + 2 H2O. Elektrony z cytochromu c přebírá cytochrom a; odtud jsou přenášeny na cytochrom a3 (dochází ke změně oxidačního čísla mezi Cu2+ a Cu+). Cytochrom a3 postupně redukuje molekuly kyslíku na oxidové ionty (O2-), které s protony z matrix reagují za vzniku molekuly vody. Hlavní úloha kyslíku, jejž vdechujeme, spočívá tedy v tom, aby udržoval komplex cytochromů v oxidovaném stavu. 4H+ 2 H2O.")
51
Enzymové komplexy Matrix II Sukcinát-CoQ oxidoreduktasa III CoQH2–
cytochrom c oxidoreduktasa I NADH-CoQ oxidoreduktasa IV Cytochrom c oxidasa V ATP-synthasa 4H+ 4H+ 2H+ H+ Cyt c Cyt c Cyt c II III I IV V CoQH. 2e- e- 2e- CoQH. e- 4e- CoQH2 CoQH2 e- e- e- 2H+ 2H+ e- O2 + 2H+ 2H+ 2H+ 4H+ FADH2 Matrix Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. Dýchací řetězec se skládá ze 4 enzymových komplexů a z enzymu ATP-synthasy. Ubichinony (koenzym Q, CoQ) a cytochromy (cyt) slouží jako pohyblivé přenašeče elektronů (koenzym Q i protonů). Během přenosu elektronů vzniká energie, kterou využívá každý enzymový komplex k čerpání protonů (protony pocházejí z vody H2O ↔ H+ + OH-) z matrix do mezimembránového prostoru – vytváří se elektrochemický protonový gradient (koncentrace protonů je v mezimembránovém prostoru daleko vyšší než v matrix). Při přechodu protonů zpět do matrix vzniká energie, která pohání syntézu ATP z ADP a anorganického fosfátu (jedná se o oxidační fosforylaci). Protony roztáčí stopku hlavy ATP-synthasy, a tím dochází k syntéze ATP. Takto popsaný vznik molekuly ATP popisuje tzv. chemiosmotická teorie, za kterou dostal v roce 1978 Peter Mitchell Nobelovu cenu. Dehydrogenací jedné molekuly NADH se celkem přenese 6 protonů do mezimembránového prostoru, dehydrogenací sukcinátu se přenesou jen 4 protony. Mitchell předpokládal, že na vznik jedné molekuly ATP je zapotřebí zpětného přenosu dvou protonů. To odpovídá představě, že oxidací jedné molekuly NADH vznikají 3 molekuly ATP, oxidací jedné molekuly FADH2 vznikají 2 molekuly ATP. Z toho vyplývá, že při odbourávání jedné molekuly acetylkoenzymu A vznikne 12 molekul ATP (1 z GTP, 9 ze tří molekul NADH a 2 z jedné molekuly FADH2). Nejvíce energie tedy vzniká při tvorbě vody v dýchacím řetězci. FAD NADH + H+ 2H2O NAD+ ADP + P H+ Fumarát -OOC-CH=CH-COO- ATP Sukcinát -OOC-CH2-CH2-COO-
 a cytochromy (cyt) slouží jako pohyblivé přenašeče elektronů (koenzym Q i protonů). Během přenosu elektronů vzniká energie, kterou využívá každý enzymový komplex k čerpání protonů (protony pocházejí z vody H2O ↔ H+ + OH-) z matrix do mezimembránového prostoru – vytváří se elektrochemický protonový gradient (koncentrace protonů je v mezimembránovém prostoru daleko vyšší než v matrix). Při přechodu protonů zpět do matrix vzniká energie, která pohání syntézu ATP z ADP a anorganického fosfátu (jedná se o oxidační fosforylaci). Protony roztáčí stopku hlavy ATP-synthasy, a tím dochází k syntéze ATP. Takto popsaný vznik molekuly ATP popisuje tzv. chemiosmotická teorie, za kterou dostal v roce 1978 Peter Mitchell Nobelovu cenu. Dehydrogenací jedné molekuly NADH se celkem přenese 6 protonů do mezimembránového prostoru, dehydrogenací sukcinátu se přenesou jen 4 protony. Mitchell předpokládal, že na vznik jedné molekuly ATP je zapotřebí zpětného přenosu dvou protonů. To odpovídá představě, že oxidací jedné molekuly NADH vznikají 3 molekuly ATP, oxidací jedné molekuly FADH2 vznikají 2 molekuly ATP. Z toho vyplývá, že při odbourávání jedné molekuly acetylkoenzymu A vznikne 12 molekul ATP (1 z GTP, 9 ze tří molekul NADH a 2 z jedné molekuly FADH2). Nejvíce energie tedy vzniká při tvorbě vody v dýchacím řetězci. FAD. NADH + H+ 2H2O. NAD+ ADP + P. H+ Fumarát. -OOC-CH=CH-COO- ATP. Sukcinát. -OOC-CH2-CH2-COO-")
52
Reakce enzymových komplexů
Komplex I: NADH-CoQ-oxidoreduktasa NADH + H+ + CoQ → NAD+ + CoQH2 Komplex II: Sukcinát-CoQ-oxidoreduktasa FADH2 + CoQ → FAD + CoQH2 Komplex III: CoQH2-cytochrom-c-oxidoreduktasa CoQH2 + 2 cyt c (Fe3+) → CoQ + 2 cyt c (Fe2+) + 2H+ Komplex IV: Cytochrom-c-oxidasa Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. 4 cyt c (Fe2+) H+ → 4 cyt c (Fe3+) + 2 H2O Kyslík udržuje komplex cytochromů v oxidovaném stavu (Fe2+ zoxiduje zpátky na Fe3+).
 → CoQ + 2 cyt c (Fe2+) + 2H+ Komplex IV: Cytochrom-c-oxidasa. Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. 4 cyt c (Fe2+) H+ → 4 cyt c (Fe3+) + 2 H2O. Kyslík udržuje komplex cytochromů v oxidovaném stavu. (Fe2+ zoxiduje zpátky na Fe3+).")
53
Koenzym FADH2

54
Koenzym FADH2 V dýchacím řetězci probíhá oxidace redukovaných koenzymů.

55
Koenzym NADH

56
Koenzym NADH V dýchacím řetězci probíhá oxidace redukovaných koenzymů.

57
NADH + H+ + CoQ → NAD+ + CoQH2
Koenzym Q, Koenzym QH2 Obsah probírané látky na snímcích č je nad rámec středoškolského pojetí biochemického učiva. Snímky jsou určeny především pro středoškolské učitele biochemie. NADH + H+ + CoQ → NAD+ + CoQH2 FADH2 + CoQ → FAD + CoQH2

61
Použitá literatura [1] ALBERTS, B. a kol. Základy buněčné biologie. Ústí nad Labem: Espero Publishing, 1997. [2] SOFROVÁ, D. – TICHÁ, M. a kol. Biochemie – základní kurz. Praha: skripta UK, 1993. [3] KARLSON, P. Základy biochemie. Praha: Academia, 1981. [4] BLOOMFIELD, M., M. Chemistry and the Living Organism. Canada: John Wiley & Sons, Inc., 1992. [5] KLOUDA, P. Základy biochemie. Ostrava: nakladatelství Pavel Klouda, 2000. [6] VODRÁŽKA, Z. Biochemie. Praha: Scientia, 1998. [7] McKEE, T. – McKEE, J. Biochemistry: an introduction. United States of America: McGraw-Hill Companies, 1999. Obrázky: Milada Teplá (Adobe flash, ChemSketch)
![Použitá literatura [1] ALBERTS, B. a kol. Základy buněčné biologie. Ústí nad Labem: Espero Publishing,](http://slideplayer.cz/slide/2535830/9/images/61/Pou%C5%BEit%C3%A1+literatura+%5B1%5D+ALBERTS%2C+B.+a+kol.+Z%C3%A1klady+bun%C4%9B%C4%8Dn%C3%A9+biologie.+%C3%9Ast%C3%AD+nad+Labem%3A+Espero+Publishing%2C.jpg "[2] SOFROVÁ, D. – TICHÁ, M. a kol. Biochemie – základní kurz. Praha: skripta UK, [3] KARLSON, P. Základy biochemie. Praha: Academia, [4] BLOOMFIELD, M., M. Chemistry and the Living Organism. Canada: John Wiley & Sons, Inc., [5] KLOUDA, P. Základy biochemie. Ostrava: nakladatelství Pavel Klouda, [6] VODRÁŽKA, Z. Biochemie. Praha: Scientia, [7] McKEE, T. – McKEE, J. Biochemistry: an introduction. United States of America: McGraw-Hill Companies, Obrázky: Milada Teplá (Adobe flash, ChemSketch)")