Stáhnout prezentaci
1
Transkripce u prokaryot a eukaryot (kapitola 11)
Molekulární biologie 3. Transkripce u prokaryot a eukaryot (kapitola 11) (regulace transkripce – zvláštní přednáška později)
 (regulace transkripce – zvláštní přednáška později)")
2
messenger RNA (mRNA) - kóduje informaci pro výrobu proteinů
Transkripce – proces, při kterém je informace z DNA převedena do RNA. DNA pouze uchovává informaci, ale kromě toho sama není zdrojem funkcí nutných pro život buňky. Slouží jako tempát pro výrobu RNA (mRNA i nekódující RNA) a proteinů. RNA messenger RNA (mRNA) - kóduje informaci pro výrobu proteinů nekódující RNA – plní funkci sama o sobě, nekóduje proteiny (tRNA,rRNA,snRNA…) Genová exprese = mRNA + protein
 a proteinů. RNA. messenger RNA (mRNA) - kóduje informaci pro výrobu proteinů. nekódující RNA – plní funkci sama o sobě, nekóduje proteiny (tRNA,rRNA,snRNA…) Genová exprese = mRNA + protein.")
3
RNA vlákno je komplementární k antisense vláknu DNA
antisense vlákno (templátové, nekódující) sense vlákno (netemplátové, kódující) vlákna jsou od sebe dočasně oddělena (denaturují) vyrobeno nové vlákno RNA RNA vlákno je komplementární k antisense vláknu DNA RNA stejná jako sense vlákno DNA, ale z ribonukleotidů a místo T jsou U syntéza vždy ve směru 5’ - 3’ (3’OH ribozy s 5’fosfátem sousedního nukletodu) pouze jedno vlákno se přepisuje (transkribuje) do RNA Pouze malý počet genů je v daný moment transkribováno, většina podléhá přísně časové a tkánově specifické regulaci podle aktuálních potřeb buňky. Houskeeping geny (konstitutivní) – geny nutné pro základní buněčné procesy, exprimovány téměř pořád
 sense vlákno. (netemplátové, kódující) vlákna jsou od sebe dočasně oddělena (denaturují) vyrobeno nové vlákno RNA. RNA vlákno je komplementární. k antisense vláknu DNA. RNA stejná jako sense vlákno DNA, ale z ribonukleotidů a místo T jsou U. syntéza vždy ve směru 5’ - 3’ (3’OH ribozy s 5’fosfátem. sousedního nukletodu) pouze jedno vlákno se přepisuje (transkribuje) do RNA. Pouze malý počet genů je v daný moment transkribováno, většina podléhá přísně časové a tkánově specifické regulaci podle aktuálních potřeb buňky. Houskeeping geny (konstitutivní) – geny nutné pro základní buněčné procesy, exprimovány téměř pořád.")
4
Pouze jedno vlákno je transkribováno pro daný gen, ale někdy může být druhé vlákno užito jako templát pro jiný gen ležící v opačné orientaci a překrývající se s prvním!

5
Většinou geny s podobnou funkcí (pro stejnou metabolickou dráhu).
cistron – část DNA nebo RNA kódující jeden protein či jednu nekódující RNA operon – u prokaryot, několik genů v těsné blízkosti přepisující se do jedné společné mRNA, která pak ale kóduje více proteinů a je tedy polycistronní. Většinou geny s podobnou funkcí (pro stejnou metabolickou dráhu). EUKARYOTA PROKARYOTA OPERON transkripce Cistron kodujici protein = ORF. Trypanosoma je eukaryot, ale ma jako jedina polycistronni RNA. translace MONOCISTRONNÍ RNA POLYCISTRONNÍ RNA
. EUKARYOTA. PROKARYOTA. OPERON. transkripce. Cistron kodujici protein = ORF. Trypanosoma je eukaryot, ale ma jako jedina polycistronni RNA. translace. MONOCISTRONNÍ RNA. POLYCISTRONNÍ RNA.")
6
Na eukaryotní RNA probíhá po transkripci mnoho úprav, než je ji možno přepsat do proteinu!
úpravy export z jádra Figure Molecular Biology of the Cell (© Garland Science 2008)
")
7
Ne celá mRNA kóduje protein!
Struktura genu Ne celá mRNA kóduje protein! ORF Regulační elementy v UTRs… Enhancery u eukaryot… promotor – část DNA před genem, na kterou se váže RNA polymeráza a další proteiny umožňující začátek transkripce 5’UTR (5’untranslated region) – oblast mRNA na jejím 5’konci, která není translatována 3’UTR (3’untranslated region) – oblast mRNA na jejím 3’konci, která není translatována, za stop kodonem ORF (open reading frame) – otevřený čtecí rámec, souvislý sled bazí DNA(RNA) kódujcí protein
 – oblast mRNA na jejím 5’konci, která není translatována. 3’UTR (3’untranslated region) – oblast mRNA na jejím 3’konci, která není translatována, za stop kodonem. ORF (open reading frame) – otevřený čtecí rámec, souvislý sled bazí DNA(RNA) kódujcí protein.")
8
TRANSKRIPCE U PROKARYOT
Proces samotné syntézy RNA je vpodstatě totožný u prokaryot a eukaryot, ale liší se regulační mechanismy.

9
Jak RNA polymeráza rozpozná správné místo na DNA, kde začít transkripci?
dvojvláknová DNA komplex RNA polymerázy jednovláknová mRNA

10
RNA polymeráza Molekulární interakce se ‘sigma () faktorem’ umožní RNA polymeráze vázat DNA na specifických úsecích DNA nazývaných promotory -35 -10 +1 Samotné ‘core’ RNA polymerázového komplexu je schopné syntézy RNA, ale není schopné rozpoznat a vázat DNA Ze sigma podjednotky specificke, ty reguluji do určite miry transkripci… Figure 6-11 (part 1 of 7) Molecular Biology of the Cell (© Garland Science 2008)
 Molecular Biology of the Cell (© Garland Science 2008)")
11
Rozpoznání promotoru -35 -10 -10 a -35 sekvence
(vzhledem k počátku transkripce) sekvence rozeznávané sigma podjednotkou, umožňují vazbu core, rozpletení DNA a počátek transkripce. Sigma faktor se na DNA nemůže vázat sám, ale vždy v komplexu se zbytkem polymerázy. Sigma pokrývá 80 bp! Core = tělo podle Vlasaka holoenzym RNA polymerázy = core ze 4 podjednotek + sigma podjednotka (= 5 celkem)
 sekvence rozeznávané sigma podjednotkou, umožňují vazbu core, rozpletení DNA a počátek transkripce. Sigma faktor se na DNA nemůže vázat sám, ale vždy v komplexu se zbytkem polymerázy. Sigma pokrývá 80 bp! Core = tělo podle Vlasaka. holoenzym RNA polymerázy. = core ze 4 podjednotek + sigma podjednotka (= 5 celkem)")
12
úsek bohatý na TA, snadněji se rozvolní (taje)
-10 a -35 sekvence E.coli -35 -10 ‘konsensus‘ sekvence úsek bohatý na TA, snadněji se rozvolní (taje) Síla promotoru je dána podobností -10 a -35 sekvence ke konsensu (čím podobnější, tím častější nasedání polymerázy a tím silnější promotor) Promotorové sekvence se liší mezi jednotlivými geny i mezi jednotlivými druhy prokaryot.
 Síla promotoru je dána podobností -10 a -35 sekvence ke konsensu (čím podobnější, tím častější nasedání polymerázy a tím silnější promotor) Promotorové sekvence se liší mezi jednotlivými geny i mezi jednotlivými druhy prokaryot.")
13
Průběh transkripce RNA polymeráza
trankripční bublina žlábek uvnitř polymerázy pojme 16bp DNA (u eukaryot 25) antisense vlákno rostoucí řetězec mRNA RNA polymeráza sense vlákno směr syntézy Po navázání sigma faktoru se vytvoří transkripční bublina na -10 oblasti (TA rich), díky tomu otevření DNA a umožnění syntézy prvních 8-9 párů bazí mRNA. Bublina pokračuje spolu s polymerázou po celé délce genu. První transkribovaná báze je většinou A obklopená dvěma pyrimidiny (CAT sekvence), někdy G. Není potřeba primer jako u DNA replikace.
 antisense vlákno. rostoucí řetězec mRNA. RNA. polymeráza. sense vlákno. směr syntézy. Po navázání sigma faktoru se vytvoří transkripční bublina na -10 oblasti (TA rich), díky tomu otevření DNA a umožnění syntézy prvních 8-9 párů bazí mRNA. Bublina pokračuje spolu s polymerázou po celé délce genu. První transkribovaná báze je většinou A obklopená dvěma pyrimidiny (CAT sekvence), někdy G. Není potřeba primer jako u DNA replikace.")
14
iniciační fáze: RNA polymeráza se naváže a zůstává sedět na promotoru až do syntézy prvních 8-9 bazí RNA elongační fáze: sigma faktor odpadne, core polymerázy se uvolní z promotoru a syntetizuje zbytek mRNA rychlostí 40 nukleotidů za sekundu (replikace 1000 bp/s, translace 15 AA/s) kontrola správnosti inkorporovaných bazí je malá. okamžité pokrytí mRNA ribozomy, mRNA před translací nepodléhá dalším úpravám gyráza přidává negativní superotáčky před polymerázou, topoizomeráza I odebírá negativní superotáčky za polymerázou terminační fáze: ukončení transkripce odpojením RNA od polymerázy na specifických místech
 kontrola správnosti inkorporovaných bazí je malá. okamžité pokrytí mRNA ribozomy, mRNA před translací nepodléhá dalším úpravám. gyráza přidává negativní superotáčky před polymerázou, topoizomeráza I odebírá negativní superotáčky za polymerázou. terminační fáze: ukončení transkripce odpojením RNA od polymerázy na specifických. místech.")
15
Jak RNA polymeráza rozpozná správné místo, kde skončit transkripci?
dvojvláknová DNA komplex RNA polymerázy jednovláknová mRNA

16
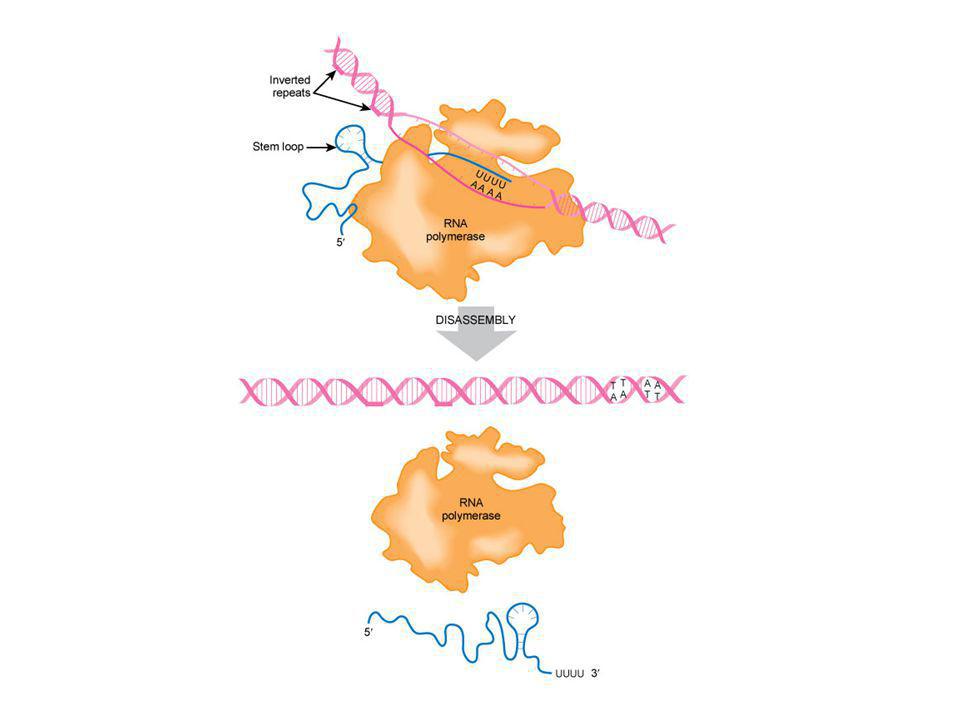
TERMINACE TRANSKRIPCE
terminátor - invertovaná repetice, za ní několik A, tvorba vlásenky mRNA na svém 3’konci vytvoří vlásenku, což způsobí zastavení polymerázy (asi 60 sekund) slabé vazby mezi úsekem UUU v mRNA a AAA v DNA způsobí samovolné odpojení mRNA odpadne i polymeráza
 slabé vazby mezi úsekem UUU v mRNA a AAA v DNA způsobí samovolné odpojení mRNA. odpadne i polymeráza.")
18
Terminace – Rho independentní (intrinsic termination)
– Rho dependentní Rho faktor ATP dependentní helikáza, odděluje řetězce DNA hexamer 6 stejných podjednotek cestuje od začátku spolu s polymerázou na mRNA se váže nukleotidů před terminátorem poté, co RNA polymeráza nasyntetizuje jeho vazebné místo (G-rich, C-poor)
")
19
Transkripční cyklus u prokaryot - video
Taky video Lac operonu…

20
Jak buňka ví, které geny zapnout?
Kontitutivní geny – zajišťují základní buněčné funkce, nutné pořád. Mají -10 a -35 sekvence blízké konsensu, sigma faktor a polymeráza nasedají samovolně, i když s různou frekvencí podle síly promotoru. Geny nutné pouze za určitých podmínek (inducibilní) Rozpoznávány specifickými sigma faktory Například sigma32 se váže pouze na geny tepelného šoku (nejsou rozpoznávány normálním sigma faktorem, jiná rozpoznávací sekvence ), umožňuje bakterii přežití vysokých teplot. Za normálních podmínek je sigma32 inaktivován vazbou na jiný protein, po odeznění tepelného šoku se opět inaktivuje. Sekvence -10 a -35 je tak málo podobná konsensu, že sigma podjednotka úsek sama nerozezná a je potřeba aktivační protein (různé pro různé skupiny genů)
 Rozpoznávány specifickými sigma faktory. Například sigma32 se váže pouze na geny tepelného šoku (nejsou rozpoznávány. normálním sigma faktorem, jiná rozpoznávací sekvence ), umožňuje bakterii přežití. vysokých teplot. Za normálních podmínek je sigma32 inaktivován vazbou na jiný. protein, po odeznění tepelného šoku se opět inaktivuje. Sekvence -10 a -35 je tak málo podobná konsensu, že sigma podjednotka úsek sama nerozezná a je potřeba aktivační protein (různé pro různé skupiny genů)")
21
Aktivační protein (aktivátor) – váže se na sekvenci v promotoru a usnadňuje nasednutí sigma faktoru, obdoba transkripčních faktorů u eukaryot. Proč se aktivační proteiny na DNA nevážou pořád?

22
Aktivační proteiny a represory mění svou konformaci
po vazbě signálních molekul vazebné místo pro signální molekulu signální molekula (například některá z živin nutných pro růst buňky) forma neschopná vazby DNA DNA vazebné místo Alostericky… U prokaryot jsou ziviny dostupne primo trasnkripcnim faktorum, u eukaryot je jadro a ziviny se tedy vazou na proteiny signalnich kaskad… forma schopná vazby DNA alosterická změna DNA vazebné místo
 forma neschopná vazby DNA. DNA vazebné místo. Alostericky… U prokaryot jsou ziviny dostupne primo trasnkripcnim faktorum, u eukaryot je jadro a ziviny se tedy vazou na proteiny signalnich kaskad… forma schopná vazby DNA. alosterická změna. DNA vazebné místo.")
23
Regulační proteiny často vážou invertované repetice na DNA
‘head-to-head’ orientace Díky 3D struktuře DNA jsou vazebná místa na stejné straně helixu.

24
Maltóza působí jako induktor
MalT protein aktivuje expresi genů zpracovávajících maltózu pokud není maltóza v buňce přítomna, MalT se není schopen vázat na DNA a transkripce genů se nekoná maltóza působí jako induktor, váže se na MalT a způsobí jeho konformační změnu. Odkryje se DNA vazebná doména, MalT se může vázat na DNA a spustí transkripci genů zpracovávající maltózu Jen příklad, totožné se slidem předtím podobné mechanismy u ostatních živin

25
Represory negativně regulují transkripci
Lac I protein blokuje expresi genů zpracovávajících laktózu váže se na operátor, sekvenci překrývající se s promotorem, čímž brání vazbě RNA polymerázy operátor promotor laktóza způsobí alosterickou konformační změnu Lac I vedoucí k odpojení od DNA, transkripce může začít induktor Může se vázat více downstream a blokovat elongaci nebo se vazou spolecne na promotor, ale polymeráza nemůže oddělit vlákna a zacit transkripci… Neplest si operator a operon induktor promotor

26
Transkripce rRNA a tRNA
rrnB operon 90% veškeré RNA v bakteriální buňce představuje rRNA: 16S, 23S a 5S rRNA jsou syntetizovány na stejném primárním transkriptu, v intronech navíc některé tRNA (ostatní tRNA samostatně nebo spolu s jinými geny). Vyštěpení RNAsami. Schematic drawing of the rrnB operon. (A) Nucleolytic processing of the rrnB primary transcript. The rRNA and tRNA species, promoters P1 and P2, and terminators T1 and T2 are indicated, as well as the processing sites of RNase III (III), RNase G (G), RNase E (E), RNase P (P), RNase T (T), and the unknown RNases (?). (B) Promoter region of the rrnB operon. Locations of FIS- and H-NS-binding sites and the UP, discriminator, and nut sequences are marked. Arrows show the start sites of transcription. Regulace pri nedostatku nabitych tRNA… Modifikace RNA bazi, CCA addition to tRNAs… jediný ‘sestřih’ probíhá u rRNA a tRNA, ne u mRNA Kaczanowska M , and Rydén-Aulin M Microbiol. Mol. Biol. Rev. 2007;71:
. Vyštěpení RNAsami. Schematic drawing of the rrnB operon. (A) Nucleolytic processing of the rrnB primary transcript. The rRNA and tRNA species, promoters P1 and P2, and terminators T1 and T2 are indicated, as well as the processing sites of RNase III (III), RNase G (G), RNase E (E), RNase P (P), RNase T (T), and the unknown RNases ( ). (B) Promoter region of the rrnB operon. Locations of FIS- and H-NS-binding sites and the UP, discriminator, and nut sequences are marked. Arrows show the start sites of transcription. Regulace pri nedostatku nabitych tRNA… Modifikace RNA bazi, CCA addition to tRNAs… jediný ‘sestřih’ probíhá u rRNA a tRNA, ne u mRNA. Kaczanowska M , and Rydén-Aulin M Microbiol. Mol. Biol. Rev. 2007;71:")
27
TRANSKRIPCE U EUKARYOT

28
mnohem složitější regulace exprese
Eukaryota mají 10x více genů než prokaryota, navíc je DNA uzavřená v jádře mnohem složitější regulace exprese transkripce probíhá v jádře, ne v cytoplazmě 3 RNA polymerázy, k tomu zvláštní RNA polymeráza v mitochondriích a chloroplastech RNA polymeráza I ribozomální RNA (5.8S, 18S, 28S), ½ až ¾ celkové RNA v buňce! RNA polymeráza III geny pro tRNA, 5S ribozomální RNA, některé snRNA a small RNA většinou konstitutivní exprese, ribozomy a tRNA nutná stále Poměry se liší podle typu buňky… hnRNA = před sestřihem, proto je jí mnohem víc. Navíc velká část nově vznikající hnRNA je degradována ještě před dokončením (než ji ochrání cap a polyA) Spis je to hnRNP – obalene proteiny… RNA polymeráza II geny kódující proteiny, snoRNA, miRNA, siRNA, snRNA potřebuje obecné (TFI, TFII, TFIII) a specifické transkripční faktory iniciační komplex se sestavuje až na promotoru (prokaryota maji sigma faktor asociovaný už před vazbou na DNA)
, ½ až ¾ celkové RNA v buňce! RNA polymeráza III. geny pro tRNA, 5S ribozomální RNA, některé snRNA a small RNA. většinou konstitutivní exprese, ribozomy a tRNA nutná stále. Poměry se liší podle typu buňky… hnRNA = před sestřihem, proto je jí mnohem víc. Navíc velká část nově vznikající hnRNA je degradována ještě před dokončením (než ji ochrání cap a polyA) Spis je to hnRNP – obalene proteiny… RNA polymeráza II. geny kódující proteiny, snoRNA, miRNA, siRNA, snRNA. potřebuje obecné (TFI, TFII, TFIII) a specifické transkripční faktory. iniciační komplex se sestavuje až na promotoru (prokaryota maji sigma faktor asociovaný už před vazbou na DNA)")
29
Transkripce rRNA pomocí PolI probíhá v genových klastrech
netranskribované oblasti V bakteriich je rRNA genu take vice, ale nejsou v clusterech Asi 200 genu 5.8S 5.8S 5.8S Klastry (shluky) mnoha tandemových repetic rRNA genů na pěti rozdílných chromozomech RNA polymeráza I transkribuje 45S RNA, která je dále štěpena na 18S, 5.8 a 28S 5S je transkribována PolIII a musí být dopravena do jadérka Ribozomání proteiny trankribovány PolII a také musejí být dopraveny do jadérka
 mnoha tandemových repetic rRNA genů na pěti rozdílných chromozomech. RNA polymeráza I transkribuje 45S RNA, která je dále štěpena na 18S, 5.8 a 28S. 5S je transkribována PolIII a musí být dopravena do jadérka. Ribozomání proteiny trankribovány PolII a také musejí být dopraveny do jadérka.")
30
Většina rRNA se transkribuje a upravuje v jadérku
fibrilární centrum obsahující RNA PolI a nově transkribované rRNA granulární oblast obsahující rRNA+ribozomální proteiny ‘vánoční stromeček’

31
RNA polymeráza I UBF1 – upstream binding factor I
Váže se na ‘core promotor’ a ‘upstream control element’, obojí GC bohaté a téměř totožné SL1 komplex obsahuje 4 proteiny včetně TBP (TATA binding protein) Promotor prekryvajici se s pocatkem transkripce Rozvolní DNA, umožní navázání RNA Pol I za pomoci TIF1A a dalších transkripčních faktorů
 Promotor prekryvajici se s pocatkem transkripce. Rozvolní DNA, umožní navázání RNA Pol I za pomoci TIF1A a dalších transkripčních faktorů.")
32
RNA polymeráza III promotor uvnitř genu
TFIIIC nebo TFIIIA se váže 50bp downstream, rozvolní chromatin umožní vazbu TFIIIB komplexu obsahující TBP a následně PolIII SL1 a TFIIIB – podobná úloha jako sigma faktory u prokaryot, správné umístění polymerázy.

33
RNA polymeráza II clamp doména
10-14 podjednotek The clamp would adopt the open conforma- tion detected in one of the 3D crystal forms (2) to allow straight double- stranded promoter DNA to lie along the bottom of the active site cleft, clear the wall, and reach the RNAPII active site. In a subsequent step of the initiation mechanism, the clamp would switch to a closed state and effectively ‘‘clamp’’ the DNA in place, thereby explaining the processivity of the polymerase. karboxy terminální doména asi 50 repetic 7 aminokyselin (Tyr Ser Pro Thr Ser Pro Ser) fosforylace na Ser a Thr umožňuje elongaci polymerázy clamp doména Otevřená umožňuje vazbu DNA, pak se uzavře a obejme DNA, umožňuje její procesivitu (elongaci bez odpadnutí Pol)
 to allow straight double- stranded promoter DNA to lie along the bottom of the active site cleft, clear the wall, and reach the RNAPII active site. In a subsequent step of the initiation mechanism, the clamp would switch to a closed state and effectively ‘‘clamp’’ the DNA in place, thereby explaining the processivity of the polymerase. karboxy terminální doména. asi 50 repetic 7 aminokyselin. (Tyr Ser Pro Thr Ser Pro Ser) fosforylace na Ser a Thr umožňuje elongaci polymerázy. clamp doména. Otevřená umožňuje vazbu DNA, pak se uzavře a obejme DNA, umožňuje její procesivitu (elongaci bez odpadnutí Pol)")
34
Regulační oblasti protein kódujících genů
specifické transkripční faktory (tkáňově nebo vývojově) obecné (bazální) transkripční faktory (TFIID,A,B,F,E,H,J) některé specifické TF ENHANCER (nebo silencer) PROMOTOR Sekvence upstream nebo downstream od promotoru vážou specifické TF. Kombinace vazebných míst pro různé TF je zodpovědná za obrovskou rozmanitost exprese jednotlivých genů, rozdílnou expresi genů v závislosti na typu tkáně, signálům z vnějšku atd.
 obecné (bazální) transkripční faktory. (TFIID,A,B,F,E,H,J) některé specifické TF. ENHANCER. (nebo silencer) PROMOTOR. Sekvence upstream nebo downstream od promotoru vážou specifické TF. Kombinace vazebných míst pro různé TF je zodpovědná za obrovskou rozmanitost exprese. jednotlivých genů, rozdílnou expresi genů v závislosti na typu tkáně, signálům z vnějšku atd.")
35
Iniciátor – sekvence DNA těsně před počátkem transkripce
PROMOTOR Iniciátor – sekvence DNA těsně před počátkem transkripce – první transkribovaná báze A jako u prokaryot TATA box - 25bp upstream, rozpoznáván TBP proteinem (TATA binding protein), TA bohata oblast uprostřed GC bohate oblasti Upstream elementy polymeráza může začít transkripci pouze za přítomnosti TATA boxu a iniciátoru, ale to je málo účinný proces TBP vaze maly zlabek… upstream elementy zvyšují účinnost transkripce po vazbě transkripčních faktorů GC box – GGGCGG, váže SP1 transkripční faktor CAAT box – GGCCAATCT, váže CTF/NF1 oktamer element – ATTTGCAT, váže Oct1
, TA. bohata oblast uprostřed GC bohate oblasti. Upstream elementy. polymeráza může začít transkripci pouze za přítomnosti TATA boxu a iniciátoru, ale to je málo účinný proces. TBP vaze maly zlabek… upstream elementy zvyšují účinnost transkripce po vazbě transkripčních faktorů. GC box – GGGCGG, váže SP1 transkripční faktor. CAAT box – GGCCAATCT, váže CTF/NF1. oktamer element – ATTTGCAT, váže Oct1.")
36
PRŮBĚH TRANSKRIPCE RNA POLYMERÁZOU II
Iniciace TFIID obsahující TBP se váže na TATA box Stejně jako u PolI a PolIIi je vždy promotor rozeznáván napřed DNA vázajícím proteinem, který teprve umožní vazbu samotné polymerázy. TFIIA, TFIIB Szntetizuje prvních pár bp TFIIB na iniciator polymeráza + TFIIH Polymeráza schopná syntézy, ale nemůže se odpoutat do promotoru – ‘připravená polymeráza’ (poised polymerase). Mnoho genů je takto zastaveno po velmi dlouhou dobu, připraveny k rychlému použití.
. Mnoho genů je takto zastaveno po velmi dlouhou dobu, připraveny k rychlému použití.")
37
Uvolnění polymerázy a elongace
TFIIF, TFIIE, TFIIJ TFIIH (CDK7) fosforyluje Ser5 v CTD doméně PolII Polymeráza se odpoutá a pokračuje elongací, všechny TFII odpadnou kromě TFIIH
 fosforyluje. Ser5 v CTD doméně PolII. Polymeráza se odpoutá a pokračuje elongací, všechny TFII odpadnou kromě TFIIH.")
38
NELF (negative elongation factor)
DSIF (DRB-sensitivity inducing factor) Vážou se na PolII a zastaví transkripci po krátkém úseku elongace – ‘pauzující polymeráza’ (paused polymerase), pokud nejsou fosforylovány spolu s CTD (Ser2), a to pomocí některých specifických transkripčních faktorů rekrutujících P-TEFB (positive transcription elongation factor B) PTEF-B NELF se dopojí Poised – na promotoru Paused – kousek po elongaci K efektivní elongaci je tedy nutné fosforylovat CTD, NELF a DSIF. Několikeré jištění, aby transkripce běžela pouze tehdy, když je to potřeba. Fosforylovaná CTD na sebe váže další faktory modifikující chromatin a usnadňující elongaci
 Vážou se na PolII a zastaví transkripci po krátkém úseku elongace – ‘pauzující polymeráza’ (paused polymerase), pokud nejsou fosforylovány spolu s CTD (Ser2), a to pomocí některých specifických transkripčních faktorů rekrutujících. P-TEFB (positive transcription elongation factor B) PTEF-B. NELF se dopojí. Poised – na promotoru. Paused – kousek po elongaci. K efektivní elongaci je tedy nutné fosforylovat CTD, NELF a DSIF. Několikeré jištění, aby transkripce běžela pouze tehdy, když je to potřeba. Fosforylovaná CTD na sebe váže další faktory modifikující chromatin a usnadňující elongaci.")
39
RNA polymeráza má 10-12 podjednotek, z toho 3 společné s PolI a PolIII
RNA polymeráza má podjednotek, z toho 3 společné s PolI a PolIII. TFII faktory často obsahují také několik podjednotek, celý iniciační komplex obsahuje mnoho různých proteinů.

40
Specifické transkripční faktory váží upstream control elementy promotoru a zároveň TFIID nebo TFIIB a TFIIA (nikdy ne přímo polymerázu). Tím napomáhají sestavení iniciačního komplexu zvyšují frekvenci transkripce.

41
ENHANCERY (a SILENCERY)
sekvence upstream nebo downstream od promotoru (tedy i uvnitř genů), často velmi daleko fungují v jakékoliv orientaci (‘5-3’nebo 3’-5’) obsahují shluky vazebných míst pro různé specifické transkripční faktory (aktivátory nebo represory) specifické transkripční faktory s sebou přinášejí proteiny pozitivně nebo negativně modifikující strukturu chromatinu a průběh transkripce (koaktivátory a korepresory) zesilují nebo zeslabují transkripci díky kontaktu s transkripčním aparátem (často přes tzv. mediátorový komplex) ohyb DNA a vytvoření smyčky (looping) Mediátor je vlastne taky koaktivátor…
, často velmi daleko. fungují v jakékoliv orientaci (‘5-3’nebo 3’-5’) obsahují shluky vazebných míst pro různé specifické transkripční faktory (aktivátory nebo represory) specifické transkripční faktory s sebou přinášejí proteiny pozitivně nebo negativně modifikující strukturu chromatinu a průběh transkripce (koaktivátory a korepresory) zesilují nebo zeslabují transkripci díky kontaktu s transkripčním aparátem (často přes tzv. mediátorový komplex) ohyb DNA a vytvoření smyčky (looping) Mediátor je vlastne taky koaktivátor…")
42
proteiny remodelující chromatin
mediátor proteiny modifikující histony

43
Iniciace transkripce u eukaryot - video

44
Terminace transkripce u Eukaryot
PolII PolII přepíše vysoce konzervovanou AAUAAA sekvenci na 3’ konci genu Na AAUAAA sekvenci se váže proteinový komplex obsahující endonukleázu a také polyadenylázu FIGURE Addition of the poly(A) tail to the primary RNA transcript of eukaryotes. Pol II synthesizes RNA beyond the segment of the transcript containing the cleavage signal sequences, including the highly conserved upstream sequence (5′)AAUAAA. 1 The cleavage signal sequence is bound by an enzyme complex that includes an endonuclease, a polyadenylate polymerase, and several other multisubunit proteins involved in sequence recognition, stimulation of cleavage, and regulation of the length of the poly(A) tail. 2 The RNA is cleaved by the endonuclease at a point 10 to 30 nucleotides 3′ to (downstream of) the sequence AAUAAA. 3 The polyadenylate polymerase synthesizes a poly(A) tail 80 to 250 nucleotides long, beginning at the cleavage site. RNA je štěpena nukleotidů za AAUAAA, polyadenyláza syntetizuje poly-A ocas. RNA polymeráza odpadne.
 tail to the primary RNA transcript of eukaryotes. Pol II synthesizes RNA beyond the segment of the transcript containing the cleavage signal sequences, including the highly conserved upstream sequence (5′)AAUAAA. 1 The cleavage signal sequence is bound by an enzyme complex that includes an endonuclease, a polyadenylate polymerase, and several other multisubunit proteins involved in sequence recognition, stimulation of cleavage, and regulation of the length of the poly(A) tail. 2 The RNA is cleaved by the endonuclease at a point 10 to 30 nucleotides 3′ to (downstream of) the sequence AAUAAA. 3 The polyadenylate polymerase synthesizes a poly(A) tail 80 to 250 nucleotides long, beginning at the cleavage site. RNA je štěpena nukleotidů za AAUAAA, polyadenyláza syntetizuje poly-A ocas. RNA polymeráza odpadne.")
45
Terminace transkripce u Eukaryot
prokaryota PolI - několik A na 3’konci genu, za nimi Sal box sekvence rozpoznávaná terminačním faktorem. Uvolnění RNA po jejím štěpení. PolII Transcription termination by nuclear RNA polymerases Patricia Richard and James L. Manleym , Genes and Dev , 2009 PolI PolIII - podobný prokaryotické terminaci, v RNA je GC bohatá část a za ní 4xU, někdy bez vlásenky. K terminaci dochází tedy samovolně, není potřeba dalších proteinů. PolIII

46
Transkripční továrny (transcription factories)
Taky specles. For splicing etc… Proximity of active genes in a shared transcription factory. (a) Co-regulated genes cluster in a specialized transcription factory. Transcription factors (yellow, red, and blue) bind their target genes while probing their nuclear environment. Upon relocation to a transcription factory, potentiated genes initiate transcription (nascent transcripts depicted in yellow and red). Dynamically bound transcription factors may dissociate from their target genes, freeing transcription factors for use by other co-regulated genes in close proximity. Thus, genes in a factory with other co-regulated genes may have a higher probability of re-initiation in that factory through dynamic exchange of transcription factors, stabilizing their presence there. By contrast, genes transcribing in the absence of other network partners (genes regulated by red and blue factors) may be more likely to dissociate from the factory after an initial burst of transcription. Repetition of factor dissociation and binding cycles would result in a transcription site highly enriched in specific binding sites and factors, seemingly specialized to preferentially transcribe a subset of co-regulated genes. (b) Close proximity between transcripts generated in a transcription factory may allow specific exons to be joined by trans-splicing. (c) Juxtaposition of active genes in a shared transcription factory may also increase the probability of translocations between loci. Current Opinion in Genetics & Development 2010, 20:127–133
 Co-regulated genes cluster in a specialized transcription factory. Transcription factors. (yellow, red, and blue) bind their target genes while probing their nuclear environment. Upon relocation to a transcription factory, potentiated genes. initiate transcription (nascent transcripts depicted in yellow and red). Dynamically bound transcription factors may dissociate from their target genes, freeing transcription factors for use by other co-regulated genes in close proximity. Thus, genes in a factory with other co-regulated genes may have a. higher probability of re-initiation in that factory through dynamic exchange of transcription factors, stabilizing their presence there. By contrast, genes. transcribing in the absence of other network partners (genes regulated by red and blue factors) may be more likely to dissociate from the factory after. an initial burst of transcription. Repetition of factor dissociation and binding cycles would result in a transcription site highly enriched in specific binding. sites and factors, seemingly specialized to preferentially transcribe a subset of co-regulated genes. (b) Close proximity between transcripts generated. in a transcription factory may allow specific exons to be joined by trans-splicing. (c) Juxtaposition of active genes in a shared transcription factory may. also increase the probability of translocations between loci. Current Opinion in Genetics & Development 2010, 20:127–133.")
47
Metody studia promotorů a DNA-proteinových interakcí
(str , kapitola 17)
")
48
Deleční analýza promotorů
Kde jsou v promotoru lokalizované regulační elementy? Klonování promotorových úseků před reportérový gen a měření transkripce podle síly exprese reportérového genu. studovaný gen gen pro luciferázu, b-galaktosidázu atd., jehož produkt lze snadno kvantifikovat reportérový gen hybridní gen neboli genová fůze

49
Deleční analýza promotorů
enhancer silencer Vážou dané úseky DNA proteny? DNAse footprint, gel shift assay, ChIP…

50
Elektroforéza nukleových kyselin horizontální elektroforéza
Separace DNA nebo RNA molekul v agarózovém gelu na základě jejich rozdílné velikosti. agarózový gel Koncentrace agarózy horizontální elektroforéza

51
polyamidakrylový gel vertikální elektroforéza

52
Váže daný úsek DNA protein(y)?
In vitro testy: DNAseI footprint koncetrace proteinového extraktu Váže daný úsek DNA protein(y)? Inkubace značené DNA próby s nukleárním extraktem a její štěpení DNAsouI. Úseky DNA pokryté proteinem nebudou přístupné štěpení. In vitro techniques used to detect DNA–protein interactions. (A) Electromobility shift assays can be used to determine direct binding between a specific sequence of radioactively labeled DNA and a purified protein. Unbound DNA, termed free probe, migrates at a relatively low molecular weight in the agarose gel. Binding of protein to this sequence results in the DNA band shifting to a high molecular weight region. Addition of an antibody that recognizes the bound protein causes an even greater shift in mobility, called supershifting. This assay can also be used with protein complexes to detect indirect protein–DNA interactions. (B) DNase footprinting assays allow identification of regions of DNA bound by proteins. A DNA oligomer is radioactively labeled on one end and mixed with the protein of interest. The DNA is then digested by a DNA endonuclease (DNase). The regions of DNA that are bound by proteins are protected from digestion. When the DNA is run out on a gel, the protected region shows up as a break in the laddering produced by DNase digestion. Vinckevicius A , and Chakravarti D J Mol Endocrinol 2012;49:R113-R123
 Inkubace značené DNA próby s nukleárním extraktem a její štěpení DNAsouI. Úseky DNA pokryté proteinem nebudou přístupné štěpení. In vitro techniques used to detect DNA–protein interactions. (A) Electromobility shift assays can be used to determine direct binding between a specific sequence of radioactively labeled DNA and a purified protein. Unbound DNA, termed free probe, migrates at a relatively low molecular weight in the agarose gel. Binding of protein to this sequence results in the DNA band shifting to a high molecular weight region. Addition of an antibody that recognizes the bound protein causes an even greater shift in mobility, called supershifting. This assay can also be used with protein complexes to detect indirect protein–DNA interactions. (B) DNase footprinting assays allow identification of regions of DNA bound by proteins. A DNA oligomer is radioactively labeled on one end and mixed with the protein of interest. The DNA is then digested by a DNA endonuclease (DNase). The regions of DNA that are bound by proteins are protected from digestion. When the DNA is run out on a gel, the protected region shows up as a break in the laddering produced by DNase digestion. Vinckevicius A , and Chakravarti D J Mol Endocrinol 2012;49:R113-R123.")
53
(Gel retardation assay)
In vitro testy: Gel shift assay (Gel retardation assay) a Váže se daný protein na určitý úsek DNA? Stanovení migrace gelem. Volná DNA gelem migruje rychleji než DNA s navázaným proteinem. In vitro techniques used to detect DNA–protein interactions. (A) Electromobility shift assays can be used to determine direct binding between a specific sequence of radioactively labeled DNA and a purified protein. Unbound DNA, termed free probe, migrates at a relatively low molecular weight in the agarose gel. Binding of protein to this sequence results in the DNA band shifting to a high molecular weight region. Addition of an antibody that recognizes the bound protein causes an even greater shift in mobility, called supershifting. This assay can also be used with protein complexes to detect indirect protein–DNA interactions. (B) DNase footprinting assays allow identification of regions of DNA bound by proteins. A DNA oligomer is radioactively labeled on one end and mixed with the protein of interest. The DNA is then digested by a DNA endonuclease (DNase). The regions of DNA that are bound by proteins are protected from digestion. When the DNA is run out on a gel, the protected region shows up as a break in the laddering produced by DNase digestion. Vinckevicius A , and Chakravarti D J Mol Endocrinol 2012;49:R113-R123
 a. Váže se daný protein na určitý úsek DNA Stanovení migrace gelem. Volná DNA gelem migruje rychleji než DNA s navázaným proteinem. In vitro techniques used to detect DNA–protein interactions. (A) Electromobility shift assays can be used to determine direct binding between a specific sequence of radioactively labeled DNA and a purified protein. Unbound DNA, termed free probe, migrates at a relatively low molecular weight in the agarose gel. Binding of protein to this sequence results in the DNA band shifting to a high molecular weight region. Addition of an antibody that recognizes the bound protein causes an even greater shift in mobility, called supershifting. This assay can also be used with protein complexes to detect indirect protein–DNA interactions. (B) DNase footprinting assays allow identification of regions of DNA bound by proteins. A DNA oligomer is radioactively labeled on one end and mixed with the protein of interest. The DNA is then digested by a DNA endonuclease (DNase). The regions of DNA that are bound by proteins are protected from digestion. When the DNA is run out on a gel, the protected region shows up as a break in the laddering produced by DNase digestion. Vinckevicius A , and Chakravarti D J Mol Endocrinol 2012;49:R113-R123.")
54
DNA afinitní chromatografie
Který protein se váže na danou DNA sekvenci? buněčný lyzát kolonka s navázanou DNA různých sekvencích odmytí proteinů, které nevážou DNA eluce proteinů vázajících DNA DNA vazebné proteiny z kroku 1 kolonka s DNA pouze se sekvencí odmytí proteinů, které nevážou Izolace proteinů vážících specifickou DNA sekvenci Izolace všech DNA vazebných proteinů

55
Jakou sekvenci váže daný protein?
Selekce oligonuklotidů Jakou sekvenci váže daný protein?

56
In vivo testy: Chromatinová precipitace Váže se daný protein na DNA přímo v jádře dané buňky? Kroslinkování všech proteinů v jádře k jejich DNA vazebným místům, lýze buňky, fragmentace DNA. Immunoprecipitace DNA-proteinových komplexů za pomoci specifické protilátky rozeznávající daný protein. Odmytí nenavázaných komplexů. Dekroslinkování a analýza izolované DNA.

57
Jaká je 3D struktura DNA-proteinováho komplexu v jádře dané buňky?
In vivo testy: ChIA-pet (chromatin interaction analysis paired end-tag sequencing) Jaká je 3D struktura DNA-proteinováho komplexu v jádře dané buňky? 1. ChIP, izolace DNA-proteinových komplexů. 2. Ligovaní dvou typů linkerů na konce fragmentů a jejich spojení ligací. Koncentrace DNAsy taková, aby se preferenčně ligovaly úseky v ramci jednotlivého komplexu. Vyštěpení ligovaných konců a jejich sekvenování. Lze vidět, které části DNA se v dané chvíli nacházely blízko daného proteinu. Koncentrace DNAsy takova, aby se preferencne ligovaly úseky v ramci jednotliveho komplexu. Figure 19.14 ChIA-PET Procedure Inside the nucleus, DNA protein interactions are 3D and involve DNA loops. After crosslinking, different regions of a chromosome are often associated with a single protein complex. After ChIP, each of these DNA sequences can be determined with paired end-tag sequencing. First, the immunoprecipitated DNA:protein complex is divided into two samples and each DNA end is connected to a different linker DNA. Then the two samples are recombined and mixed with very dilute ligase. The linkers anneal preferentially within the same complex, but occasionally there are interligations of paired tags. The ligated tags have a restriction enzyme site for MmeI, which recognizes its sequence in the tag, but cuts 20 nucleotides away in the DNA sequence. These small pieces of DNA are then sequenced using paired end-sequencing technology.
 Jaká je 3D struktura DNA-proteinováho komplexu v jádře dané buňky 1. ChIP, izolace DNA-proteinových komplexů. 2. Ligovaní dvou typů linkerů na konce fragmentů a jejich spojení ligací. Koncentrace DNAsy taková, aby se preferenčně ligovaly úseky v ramci jednotlivého komplexu. Vyštěpení ligovaných konců a jejich sekvenování. Lze vidět, které části DNA se v dané chvíli nacházely blízko daného proteinu. Koncentrace DNAsy takova, aby se preferencne ligovaly úseky v ramci jednotliveho komplexu. Figure ChIA-PET Procedure. Inside the nucleus, DNA protein interactions are 3D and involve DNA loops. After crosslinking, different regions of a chromosome are often associated with a single protein complex. After ChIP, each of these DNA sequences can be determined with paired end-tag sequencing. First, the immunoprecipitated DNA:protein complex is divided into two samples and each DNA end is connected to a different linker DNA. Then the two samples are recombined and mixed with very dilute ligase. The linkers anneal preferentially within the same complex, but occasionally there are interligations of paired tags. The ligated tags have a restriction enzyme site for MmeI, which recognizes its sequence in the tag, but cuts 20 nucleotides away in the DNA sequence. These small pieces of DNA are then sequenced using paired end-sequencing technology.")
59
Conceptual questions




>")




>")

>")










