Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Předmět: KBB/BB1P; KBB/BUBIO

2
Cytoskelet, komunikace mezi buňkami, tkáně Cytoskelet, komunikace mezi buňkami, tkáně Cíl přednášky: seznámit posluchače se strukturou a funkcí cytoskeletu a se základy buněčné signalizace a buněčné struktury tkání Klíčová slova: cytoskelet, percepce a transdukce signálu, signální kaskády, první a druhý posel, tkáň, vývoj tkání

3
Molecular Biology of the Cell Fifth Edition Molecular Biology of the Cell Fifth Edition Alberts Johnson Lewis Raff Roberts Walter Obrázky v následující prezentaci jsou převzaty z níže uvedené knihy výlučně k výukovým účelům. The illustrations in following lecture are taken from Alberts et al. Molecular Biology of the Cell, 5th Edition (Garland Science 2008) only and exclusively for the educational purposes. Copyright © Garland Science 2008
 only and exclusively for the educational purposes. Copyright © Garland Science")
4
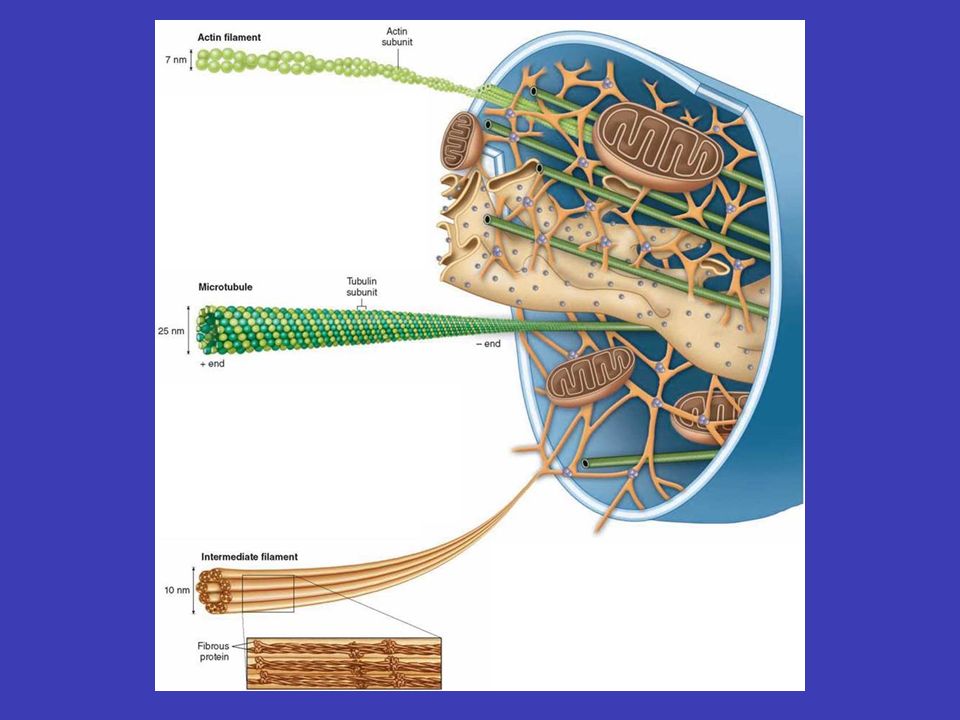
Cytoskelet cytoskelet „oporně-pohybovou“ eukaryotní buňky obsahují v cytoplazmě složitou síť proteinových vláken (cytoskelet), která jim umožňuje měnit tvar, vykonávat pohyby, zabezpečovat vezikulární transport atd., můžeme ho považovat za „oporně-pohybovou“ soustavou buňky. Cytoskelet se skládá ze tří typů vláken: střední (intermediární) filamenta střední (intermediární) filamenta (mají velkou pevnost v tahu a umožňují buňce zvládat mechanický stres, nacházejí se i v jádře) mikrotubuly mikrotubuly (jejich síť určuje pozici organel a řídí transport uvnitř buňky) aktinová vlákna aktinová vlákna (jsou důležitá pro buněčný pohyb, zejména pohybuje-li se buňka po nějakém povrchu)
 filamenta střední (intermediární) filamenta (mají velkou pevnost v tahu a umožňují buňce zvládat mechanický stres, nacházejí se i v jádře) mikrotubuly mikrotubuly (jejich síť určuje pozici organel a řídí transport uvnitř buňky) aktinová vlákna aktinová vlákna (jsou důležitá pro buněčný pohyb, zejména pohybuje-li se buňka po nějakém povrchu).")
6
Cytoskelet je vysoce dynamická struktura

7
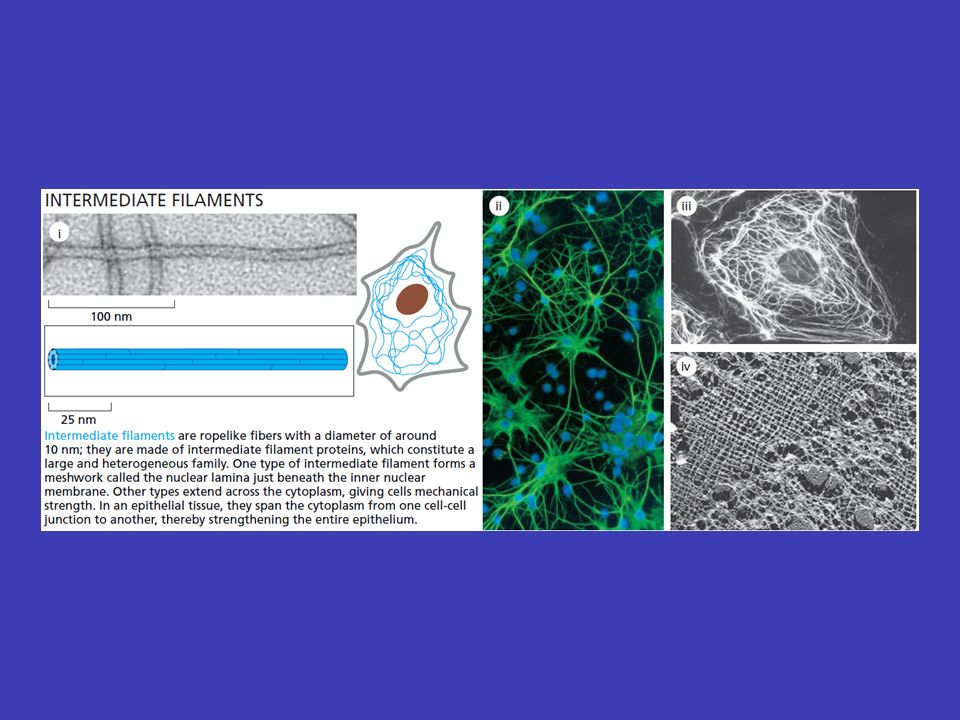
Střední filamenta pevnost v tahu mají velkou pevnost v tahu a jejich hlavní funkcí je umožnit buňkám vydržet mechanický stres cytoplazmatickou síť nacházejí se u většiny buněk kde tvoří cytoplazmatickou síť, která odklopuje jádro a odtud dosahuje až k periferii buněk desmozomů na periferii buněk je často ukotvena do plazmatické membrány v místech mezibuněčných spojů – desmozomů jadernou laminu střední filamenta se nacházejí také v buněčném jádře, kde pod jadernou membránou vytváří síťovitou strukturu – jadernou laminu, která zesiluje jadernou membránu všech eukaryontních buněk

9
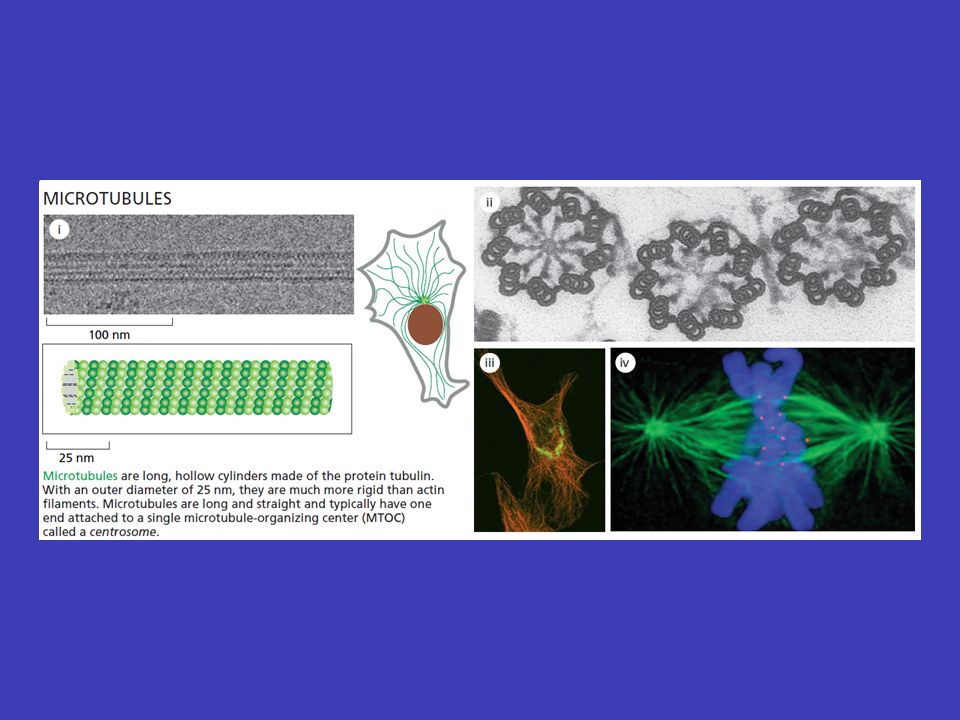
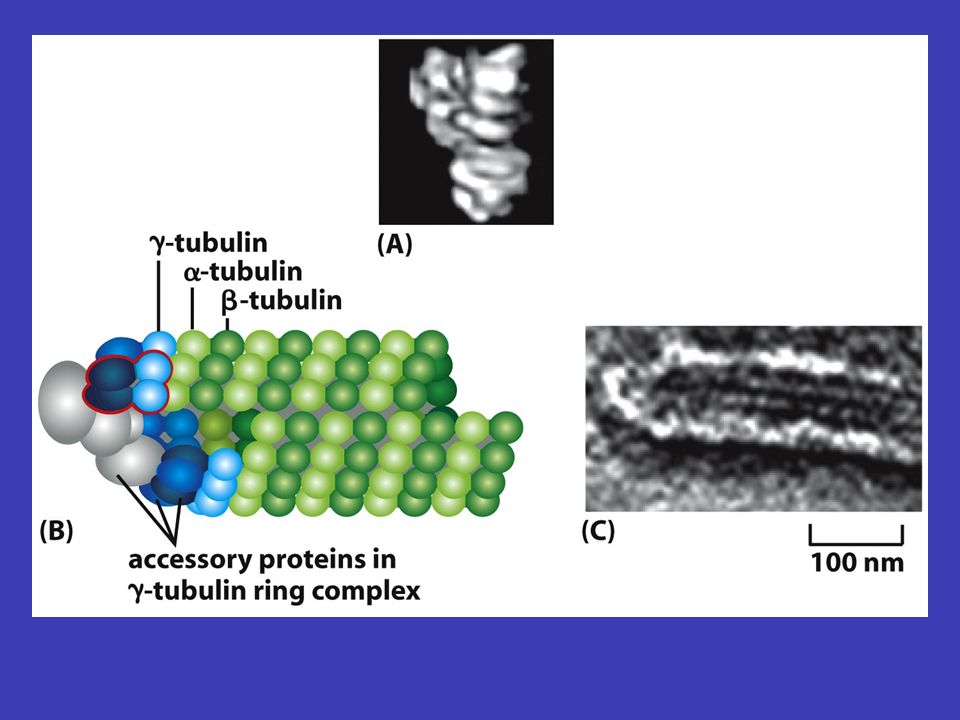
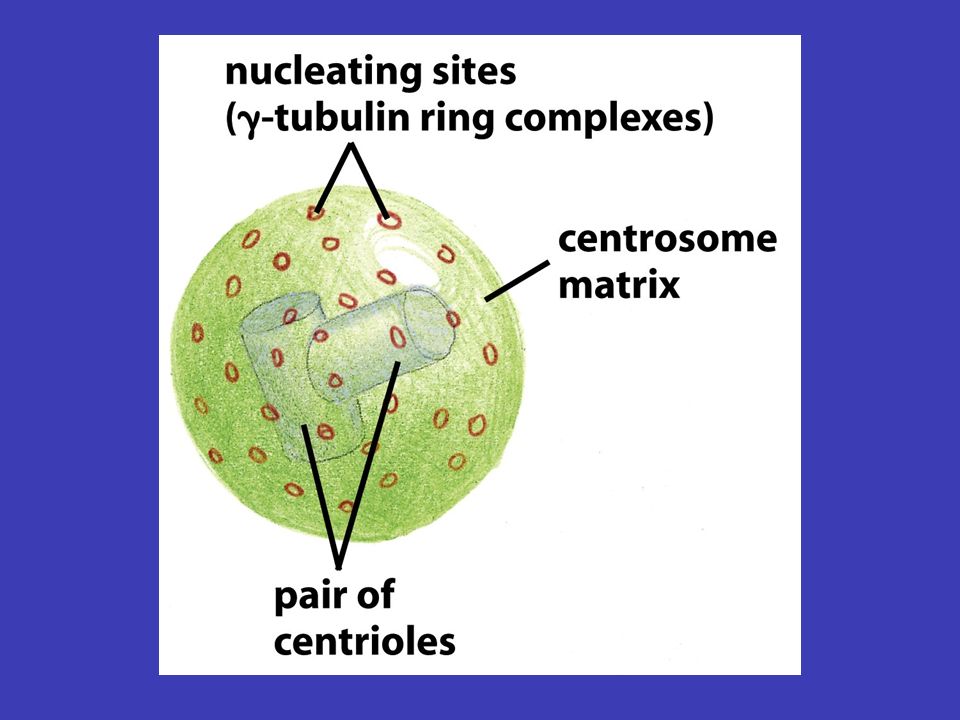
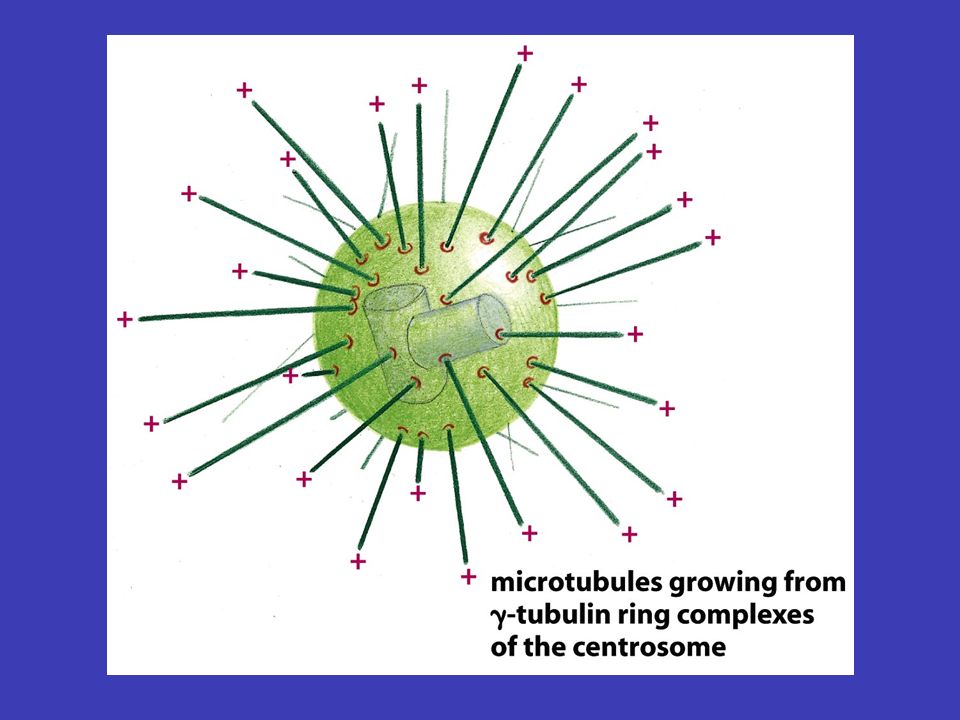
Mikrotubuly centrozomu v typické živočišné buňce mikrotubuly vyrůstají z malé struktury ve středu buňky, tzv. centrozomu – odtud sahají k periferii buňky, čili vytvářejí systém drah, podél nichž se transportují váčky, organely atd. tubulinu depolymerizacepolymerizace měnit prostorovou orientaci mikrotubuly tvoří duté vlákna skládající se z tubulinu a jsou neustále rozkládány na jednotky tubulinu a zase skládány zpátky (depolymerizace a polymerizace mikrotubulů), což jim umožňuje měnit prostorovou orientaci mikrotubul nejprve roste z centrozomu k plasmatická membráně (i několik minut), a pak začne „couvat“ (tj. je zpět odbouráván na tubuliny) buď úplně až k centrozomu nebo jen částečně – po „couvání“ ale zase roste
, což jim umožňuje měnit prostorovou orientaci mikrotubul nejprve roste z centrozomu k plasmatická membráně (i několik minut), a pak začne „couvat (tj. je zpět odbouráván na tubuliny) buď úplně až k centrozomu nebo jen částečně – po „couvání ale zase roste.")
14
Mikrotubuly mohou být stabilizovány stabilizaci pomocí tzv. čepičky jestliže neustále rostoucí a couvající mikrotubuly narazí tam, kde dorostou, na nějakou situaci, která je potřebuje, dojde k jejich stabilizaci pomocí tzv. čepičky takové mikrotubuly jsou pak využívány po žádoucí dobu a potom je z nich čepička opět odňata náhodně z centrozomu rostoucí mikrotubuly tedy náhodně rostou do různých směrů a jaksi naslepo zkoušejí najít místo, kde je jich zapotřebí (buňka nemůže „vědět“, kde právě potřebuje jejich přítomnost, proto se v evoluci prosadilo toto „slepé“ řešení)
.")
16
molekulové motory dopravu „nákladu“ (např. organel, vezikul) podél mikrotubulů zajišťují tzv. molekulové motory

17
Úloha mikrotubulů v buněčném dělení

18
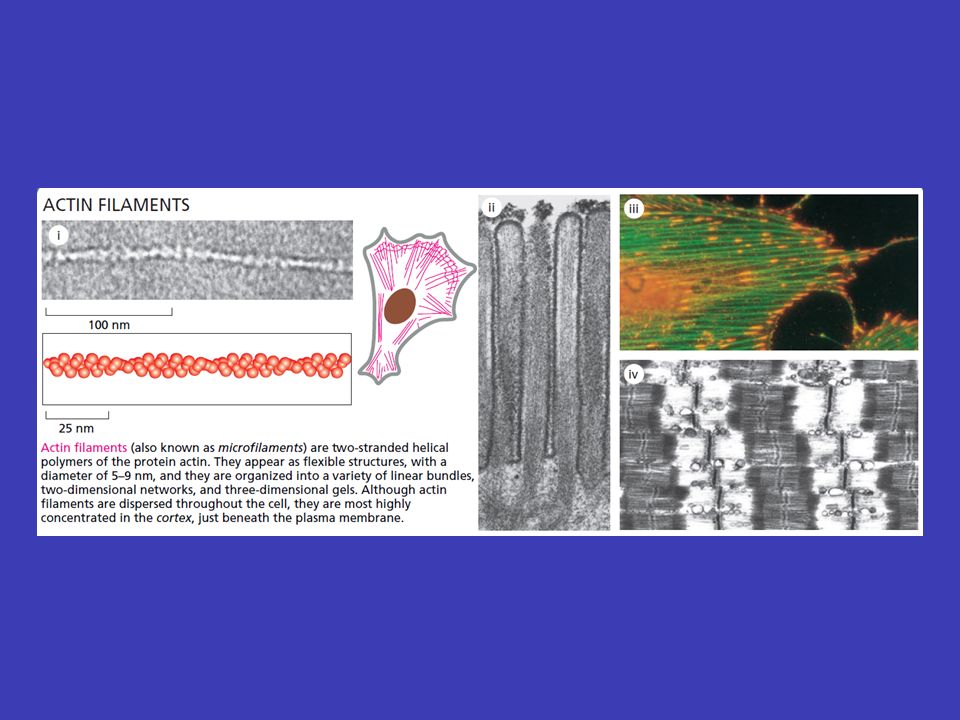
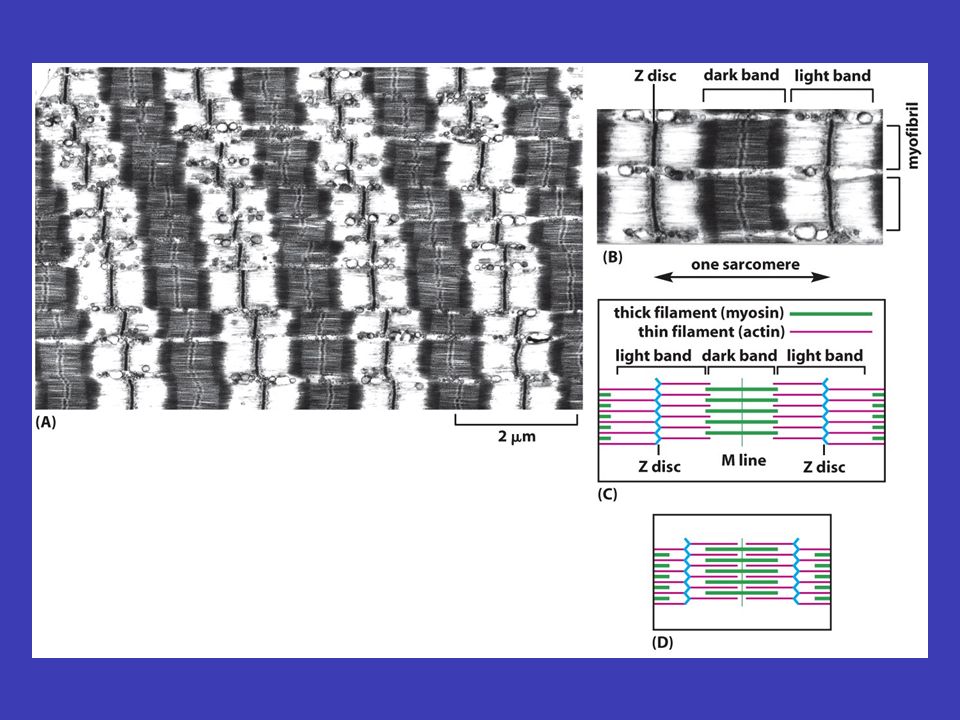
Aktinová vlákna aktinu buněčný pohyb aktinová vlákna jsou tenká a pružná, vznikají polymerizací základní jednotky aktinu a jsou zásadně důležitá pro buněčný pohyb, zejména když je uskutečňován prostřednictvím buněčného povrchu nestabilní podobně jako mikrotubuly, většina aktinových vláken je nestabilní aktin-vázajících proteinů aktinová vlákna se spojují s velkým počtem aktin-vázajících proteinů, co vede k možnosti vykonávat různé buněčné funkce a také k stabilizaci aktinových vláken ve většině buněk je aktin uložen těsně pod plasmatickou membránou v buněčném kortexu myosinem spojení aktinu s myosinem tvoří podstatu svalové práce

21
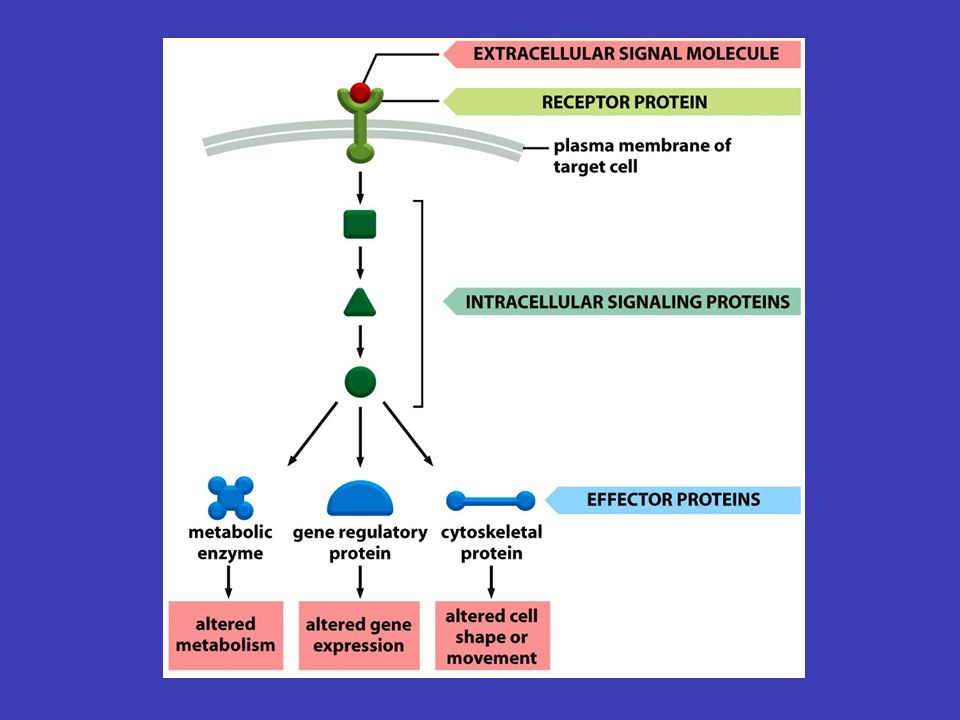
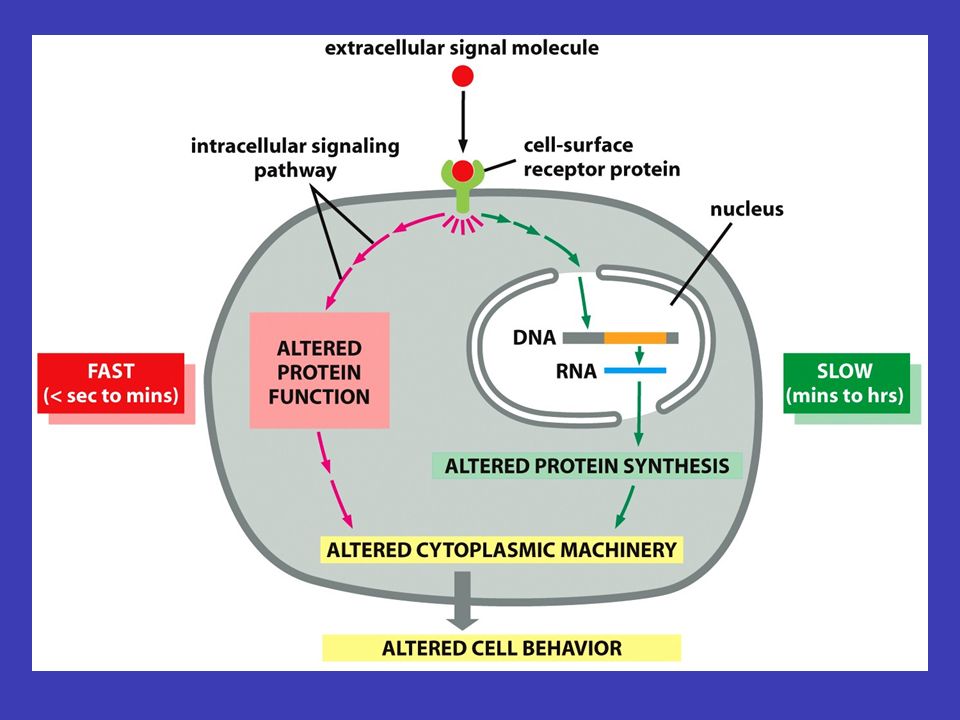
Obecné principy buněčné komunikace forem signálu percepce transdukce signálu k přenosu informace (např. aby se buňka v lidském těle začala dělit) dochází pomocí různých forem signálu – příjem signálu se nazývá percepce, přeměna jedné formy signálu v jinou formu signálu se nazývá transdukce signálu receptorového proteinu signalizující buňka produkuje specifický typ molekul, jež jsou předávány cílové buňce prostřednictvím receptorového proteinu, který rozpoznává signální molekulu a specificky na ni odpovídá buněčné signalizace v cílové buňce je přicházející extracelulární signál převáděn na intracelulární signály, jež řídí chování buňky. Přijímání a transdukce signálu spadají do svébytného oboru v rámci buněčné biologie, do tzv. buněčné signalizace
 dochází pomocí různých forem signálu – příjem signálu se nazývá percepce, přeměna jedné formy signálu v jinou formu signálu se nazývá transdukce signálu receptorového proteinu signalizující buňka produkuje specifický typ molekul, jež jsou předávány cílové buňce prostřednictvím receptorového proteinu, který rozpoznává signální molekulu a specificky na ni odpovídá buněčné signalizace v cílové buňce je přicházející extracelulární signál převáděn na intracelulární signály, jež řídí chování buňky. Přijímání a transdukce signálu spadají do svébytného oboru v rámci buněčné biologie, do tzv. buněčné signalizace.")
24
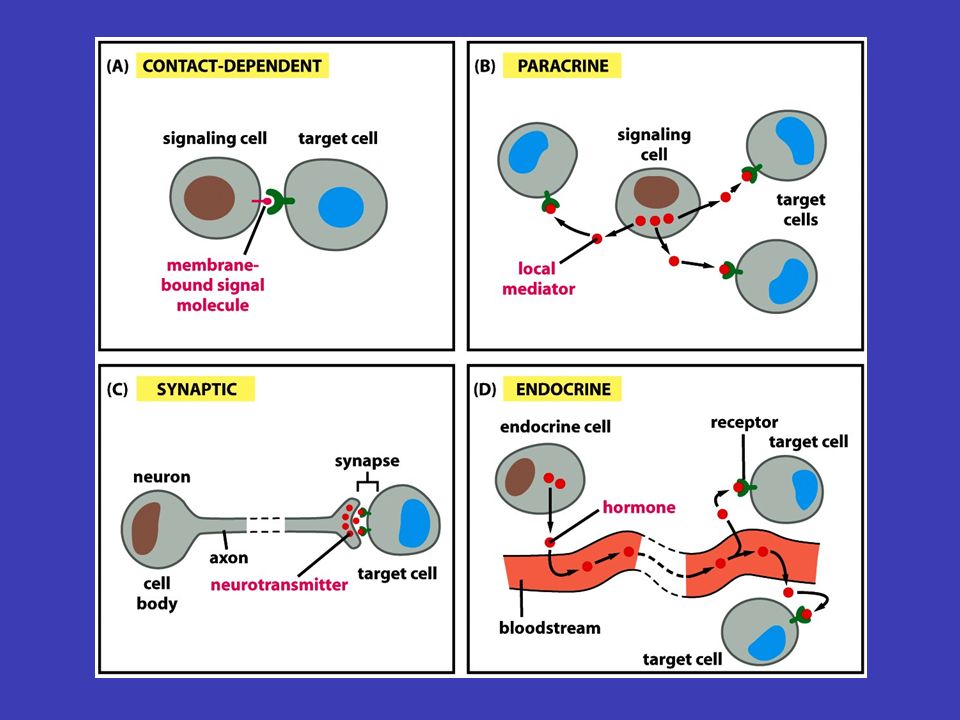
Čtyři typy signalizace hormony endokrinních buňkáchendokrinní/hormonální signalizace vyslání signálu pro celé tělo tak, že signální molekuly (hormony) se uvolní do krve živočichů nebo do mízy rostlin, tyto molekuly jsou u živočichů produkovány v endokrinních buňkách – endokrinní/hormonální signalizace lokální mediátoryparakrinní signalizace lokální difúze signálních molekul extracelulárním médiem – signály zůstávají v sousedství buněk, které je vylučují a fungují jako lokální mediátory – parakrinní signalizace neuronová signalizace nervového mediátoru acetylcholin neuronová signalizace – signály jsou předávány na velké vzdálenosti sítí vzájemně propojených neuronů prostřednictvím elektrických impulsů, jakmile dospěje impuls na konec axonu je elektrický signál transformován na extracelulární chemickou formu – vyloučení nervového mediátoru (např. acetylcholin) přímo mezi sousedními buňkami poslední typ signalizace je přímo mezi sousedními buňkami
 přímo mezi sousedními buňkami poslední typ signalizace je přímo mezi sousedními buňkami.")
26
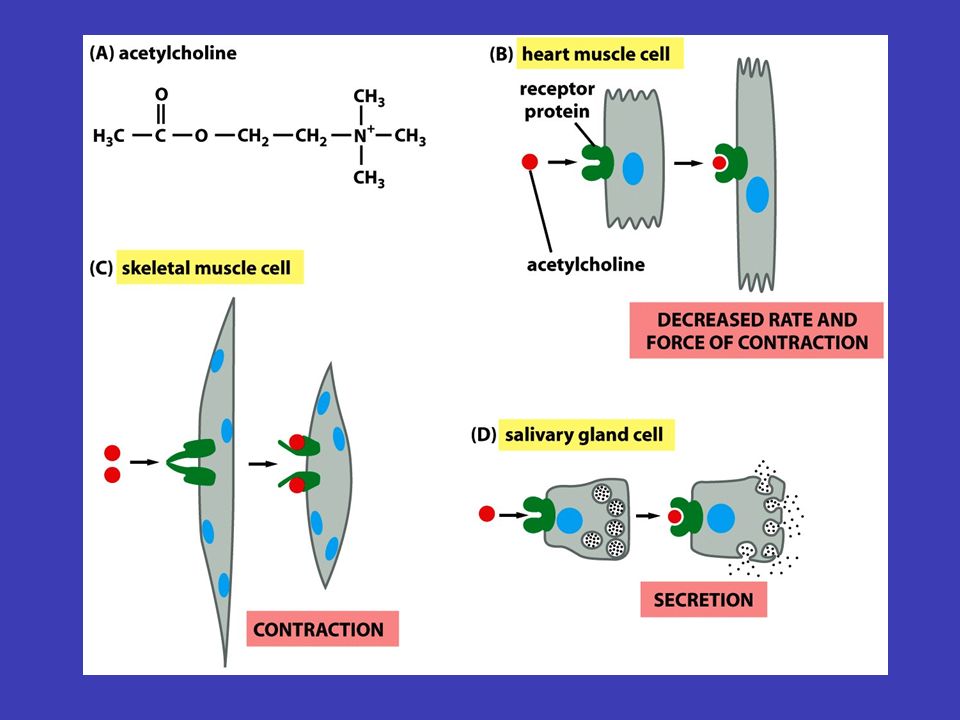
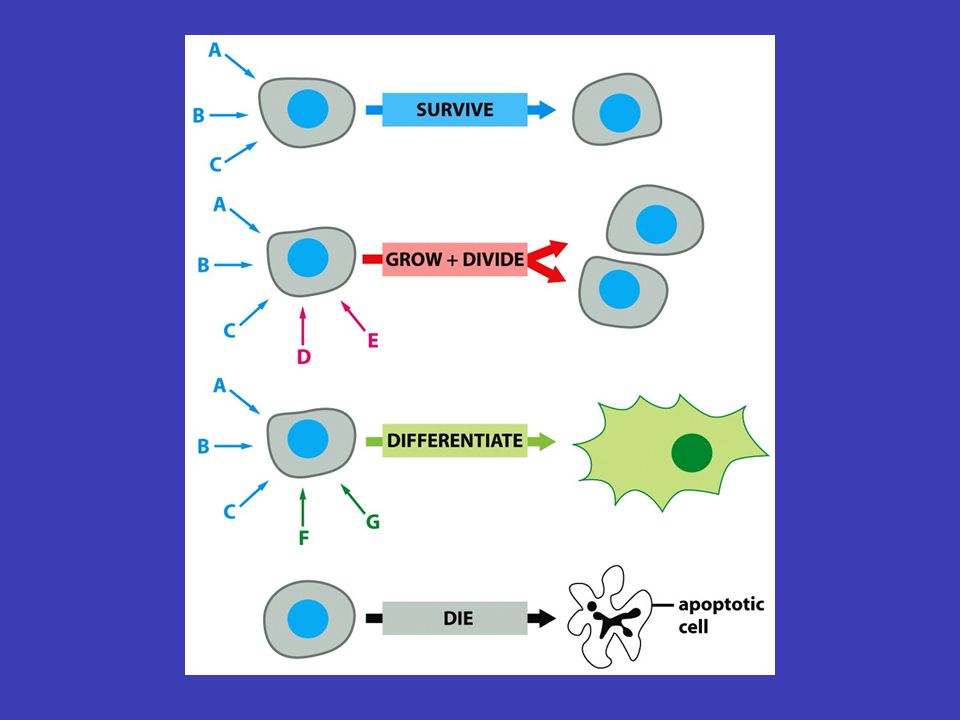
Buňky odpovídají jen na některé signály většině ale „nevěnuje pozornost“ pouze na specifické typická buňka v mnohobuněčném organismu je vystavena velkému množství signálních molekul v okolí – většině ale „nevěnuje pozornost“, neboť je zařízena tak, aby reagovala pouze na specifické (pro ni určené) signály (závisí to zejména na tom, zda má na své plazmatické membráně receptory pro daný signál) i malé množství přijímaných signálů umožňuje ale velmi složitou regulaci chování buňky v různých buňkách mít různé následky tentýž signál může navíc v různých buňkách mít různé následky: např. na acetylcholin reagují buňky srdečního svalu snížením frekvence stahů, kdežto buňky slinné žlázy vylučují složky slin

28
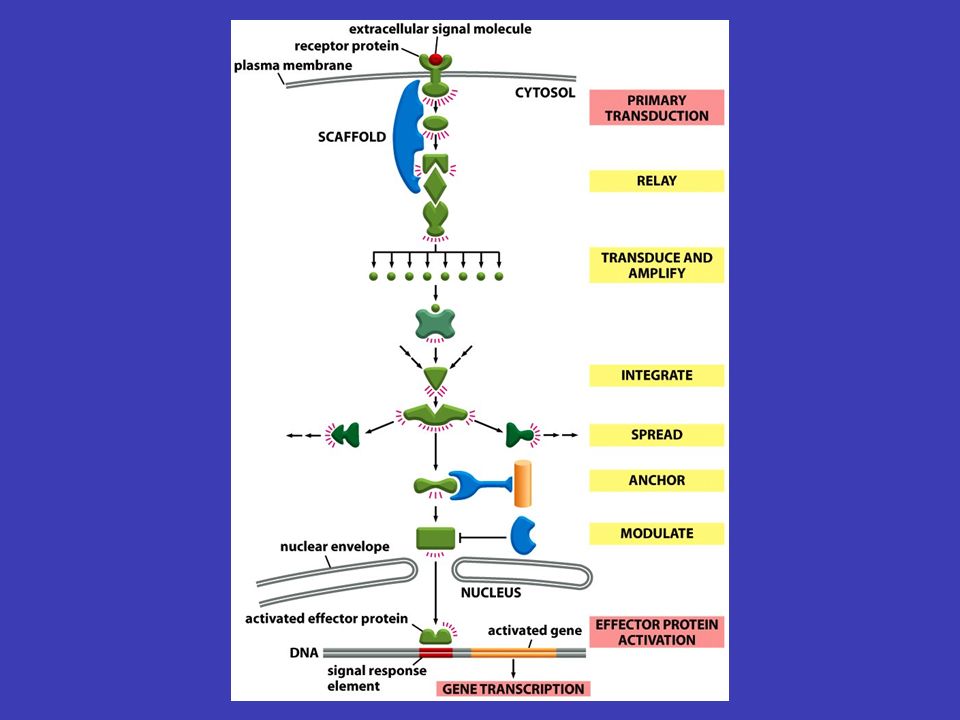
současně a různý „mix“ signálů vyvolá různé reakce signály mohou působit současně a různý „mix“ signálů vyvolá různé reakce informace zachycená zvnějšku na povrchu buňky se přenáší uvnitř od jednoho souboru intracelulárních signálních molekul k druhému – konečným výsledkem bývá zpravidla exprese konkrétních genů, aktivaci enzymů nebo aktivování cytoskeletu signální kaskády odněkud někamnějakým způsobemrůzně pozměňován štafetové řetězce přenosu signálu uvnitř buňky se nazývají signální kaskády a mají několik základních funkcí (je třeba si představit tento děj v reálné buňce, kde putuje odněkud někam, putuje nějakým způsobem a může být různě pozměňován) Signální kaskády
 Signální kaskády")
30
Funkce signálních kaskád fyzický přenos zabezpečují fyzický přenos signálu od místa, kde byl přijat, k místu jeho určení transformuje v signálních kaskádách signál probíhá samozřejmě ve formě molekul a chemických reakcí – jedna část kaskády tedy signál transformuje do chemické podoby další části kaskády amplifikace signál se během kaskády může měnit, často se např. zesiluje (tzv. amplifikace signálu) rozdělit a ovlivnit více míst signální kaskáda se také může v různých bodech rozdělit a ovlivnit více míst v buňce naráz, vznikají komplexní odpovědi modulován podle podmínek, které uvnitř i vně buňky převládají, může být signál během signální kaskády různě modulován (čili pozměněn v souvislosti s jinými signály)
 rozdělit a ovlivnit více míst signální kaskáda se také může v různých bodech rozdělit a ovlivnit více míst v buňce naráz, vznikají komplexní odpovědi modulován podle podmínek, které uvnitř i vně buňky převládají, může být signál během signální kaskády různě modulován (čili pozměněn v souvislosti s jinými signály).")
32
Extra/intracelulární receptory pro většinu signálních molekul platí, že se do buňky nemohou dostat volnou difúzí a jejich receptory tedy leží vně buňky – existují však takové signální molekuly, které snadno projdou buněčnou membránou a mají své receptory uvnitř buňky hydrofobní steroidykortisol, estradiol, testosteron thyroidní hormonythyroxin mezi hormony s intracelulárními receptory patří hydrofobní steroidy (kortisol, estradiol, testosteron) a thyroidní hormony (thyroxin) – např. když chybí specifický receptor pro testosteron při vývoji lidského zárodku, narodí se žena, která je ovšem geneticky mužem oxid dusnatý významným příkladem jiné molekuly s intracelulárním receptorem je oxid dusnatý (roztahuje cévy)
.")
33
většina povrchových receptorů spadá do jedné ze tří hlavních skupin: receptory spojené s iontovými kanály receptory spojené s iontovými kanály (ty převádějí chemické signály na elektrické tím, že otvírají a zavírají iontové kanály, čímž mění membránový potenciál) Tři hlavní třídy povrchových receptorů
 Tři hlavní třídy povrchových receptorů")
34
receptory spojené s G-proteiny receptory spojené s G-proteiny (jde o proteiny, sedmkrát prostupující plasmatickou membránu tam a sem – patří sem receptory důležité pro čich a zrak)
")
35
receptory spojené s enzymy tyrosinkinázy receptory spojené s enzymy (mají transmembránové podjednotky – patří sem např. receptorové tyrosinkinázy, které po přijetí signálu fosforylují molekulu tyrosinu v buňce)
.")
36
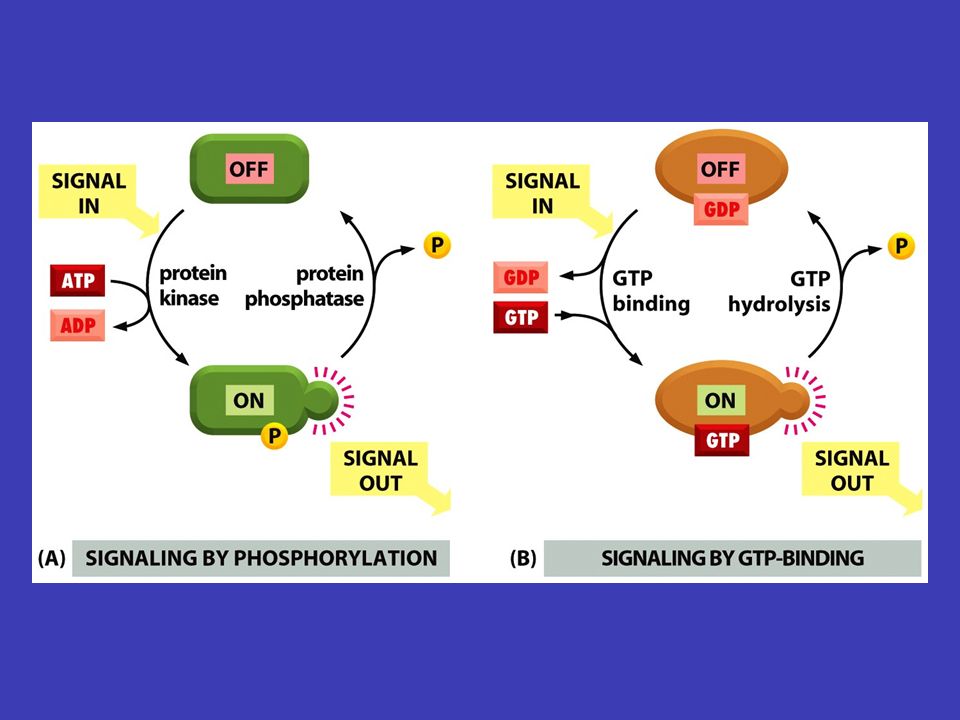
Molekulové přepínače je zapne je vypne většina proteinů ze signálních kaskád se chová jako molekulové přepínače: přijetí signálu (např. fosforylace) je zapne a přenos signálu dále (např. v důsledku toho, že takto fosforylována molekula fosforyluje jinou molekulu) je vypne vypnutí bývá stejně důležité jako zapnutí vypnutí bývá stejně důležité jako zapnutí a nedojde-li k němu v pravý čas, důsledky mohou být pro buňku či organismus fatální fosforylační kaskády velmi důležitou součástí signálních kaskád jsou tzv. fosforylační kaskády, v nichž se jednotlivé molekuly kaskády navzájem fosforylují (proto jde často o kinázy)
 je zapne a přenos signálu dále (např. v důsledku toho, že takto fosforylována molekula fosforyluje jinou molekulu) je vypne vypnutí bývá stejně důležité jako zapnutí vypnutí bývá stejně důležité jako zapnutí a nedojde-li k němu v pravý čas, důsledky mohou být pro buňku či organismus fatální fosforylační kaskády velmi důležitou součástí signálních kaskád jsou tzv. fosforylační kaskády, v nichž se jednotlivé molekuly kaskády navzájem fosforylují (proto jde často o kinázy).")
38
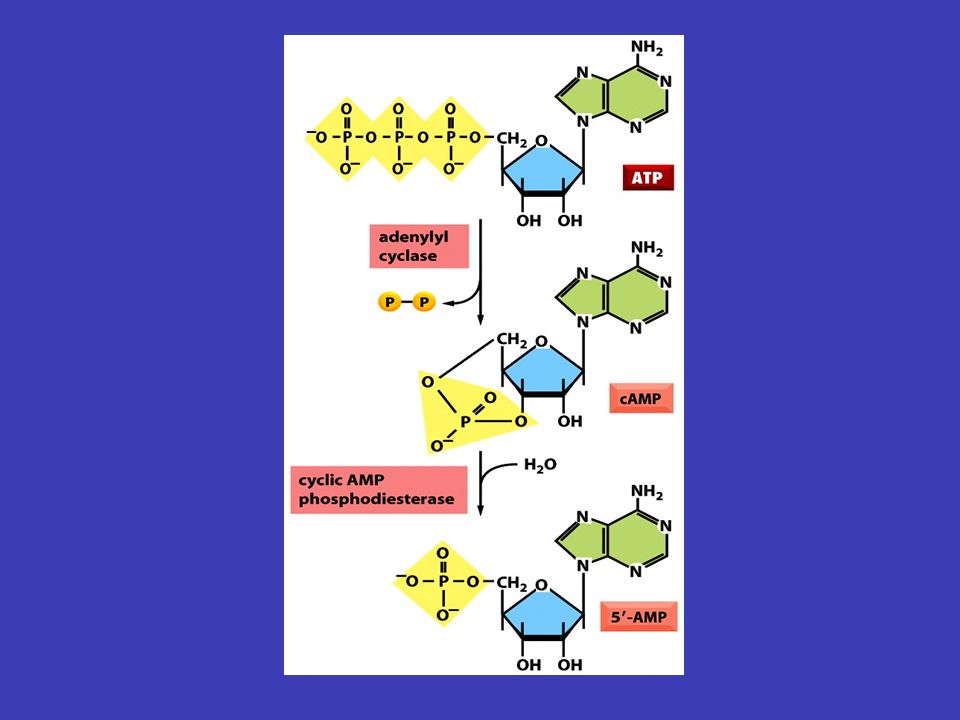
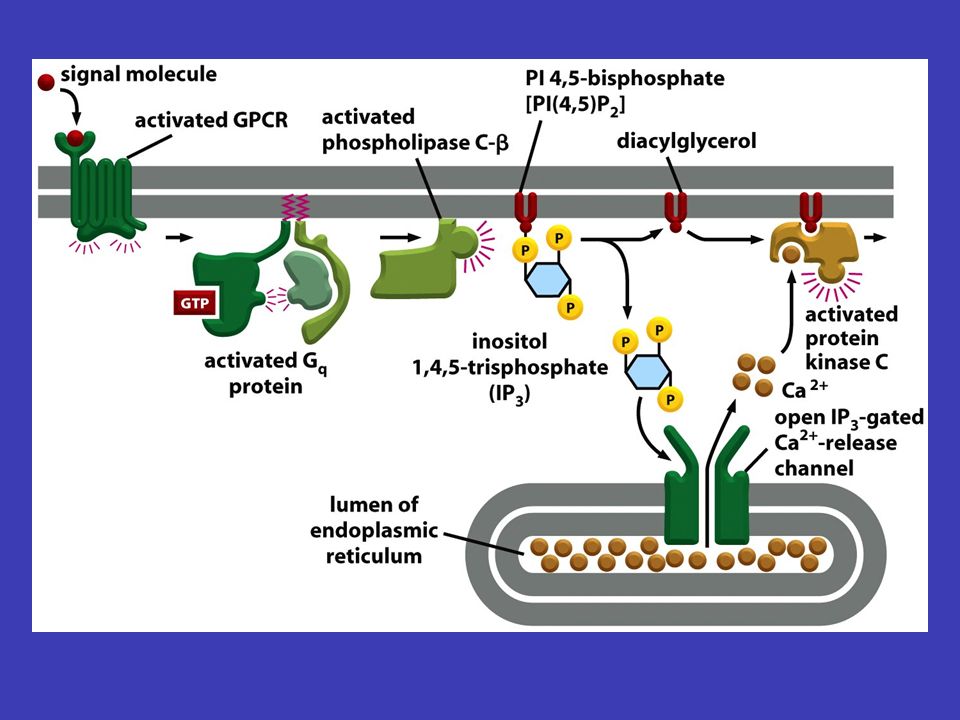
první posel jako první posel se označuje látka (např. hormon), která přináší signál zvnějšku a váže se na receptor druhý posel allostericky aktivuje nebo inhibuje vápenatý ion Ca 2+ cyklický adenosin monofosfátcAMP naopak druhý posel je látka, která v důsledku vazby prvního posla na receptor přenáší signál dále, a to tak, že allostericky aktivuje nebo inhibuje proteiny uvnitř buňky – příkladem druhého posla je např. vápenatý ion (Ca 2+ ) nebo cyklický adenosin monofosfát (cAMP) příkladem signální kaskády může být: hormon se naváže na G-proteinový receptor – aktivuje se G-protein – ten aktivuje adenylátcyklázu – ta produkuje cAMP – aktivuje proteinkinázu A – ta aktivuje v jádře daný transkripční faktor – a ten spouští expresi konkrétních genů První a druhý posel
, která přináší signál zvnějšku a váže se na receptor druhý posel allostericky aktivuje nebo inhibuje vápenatý ion Ca 2+ cyklický adenosin monofosfátcAMP naopak druhý posel je látka, která v důsledku vazby prvního posla na receptor přenáší signál dále, a to tak, že allostericky aktivuje nebo inhibuje proteiny uvnitř buňky – příkladem druhého posla je např. vápenatý ion (Ca 2+ ) nebo cyklický adenosin monofosfát (cAMP) příkladem signální kaskády může být: hormon se naváže na G-proteinový receptor – aktivuje se G-protein – ten aktivuje adenylátcyklázu – ta produkuje cAMP – aktivuje proteinkinázu A – ta aktivuje v jádře daný transkripční faktor – a ten spouští expresi konkrétních genů První a druhý posel.")
39
první posel druhý posel receptor spojený s G proteinem adenylátcykláza aktivovaná proteinkináza A

42
Tkáně tkáně pletiva většina buněk mnohobuněčných organismů je uspořádána do spolupracujících uskupení, nazývajících se tkáně (živočichové) nebo pletiva (rostliny) extracelulární matrix tkáně ovšem nejsou tvořeny jenom buňkami, nýbrž i extracelulární matrix, kterou buňky tvoří vylučováním a jež je zodpovědná za pevnost podpůrných tkání komplexní struktury tkáně obsahují nervová zakončení, cévy atd., čili vytvářejí komplexní struktury, které se neustále mění, odumírají a obnovují se pevnost rostlinných pletiv je dána buněčnými stěnami a vnitřní komplexnost pletiv je menší ve srovnání s tkáněmi – my naši pozornost upřeme ke tkáním
 nebo pletiva (rostliny) extracelulární matrix tkáně ovšem nejsou tvořeny jenom buňkami, nýbrž i extracelulární matrix, kterou buňky tvoří vylučováním a jež je zodpovědná za pevnost podpůrných tkání komplexní struktury tkáně obsahují nervová zakončení, cévy atd., čili vytvářejí komplexní struktury, které se neustále mění, odumírají a obnovují se pevnost rostlinných pletiv je dána buněčnými stěnami a vnitřní komplexnost pletiv je menší ve srovnání s tkáněmi – my naši pozornost upřeme ke tkáním")
43
Typy tkání pojivovou tkáňepitelynervovousvalovou rozlišujeme čtyři hlavní typy živočišných tkání: pojivovou tkáň, epitely, nervovou a svalovou tkáň kolagenem glukosaminoglykany pro pojivové tkáně (šlachy, škára kůže, rosol uvnitř oka, kosti, chrupavky) je typické, že obsahují velké množství extracelulární matrix, v níž jsou umístněné buňky - pevnost je přitom zajištěna nikoli celulózou jako u rostlin, ale fibrilárním proteinem kolagenem, naopak „řídkou“ složkou pojivových tkání jsou tzv. glukosaminoglykany nervovou a svalovou tkání se blíže zabývat nebudeme, zmíníme se blíže jen o epitelech

44
Fibroblast v kolagenu.

45
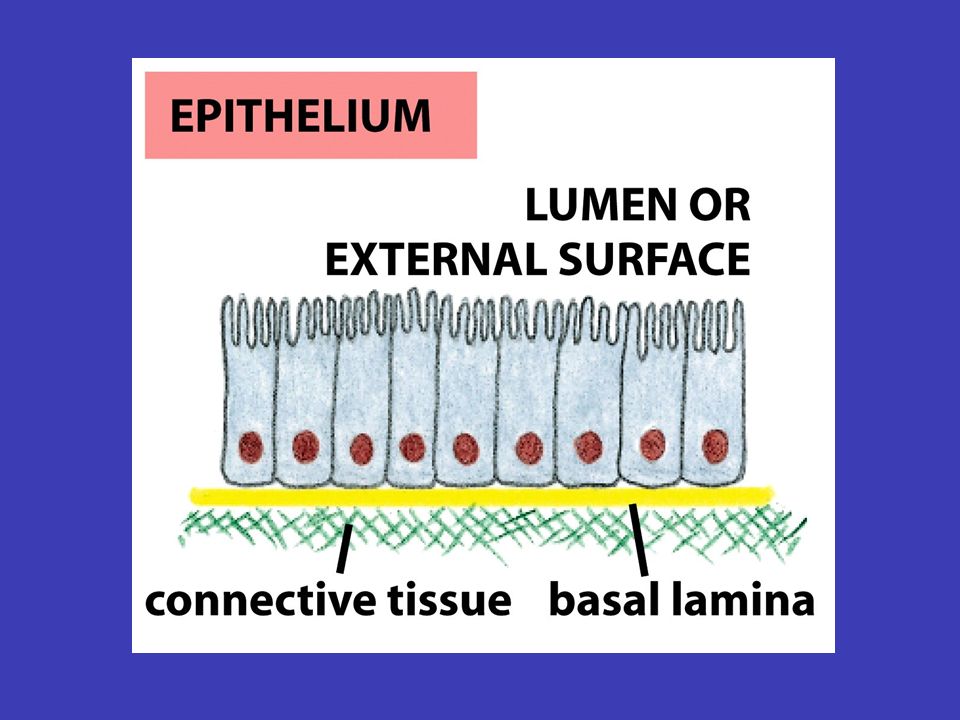
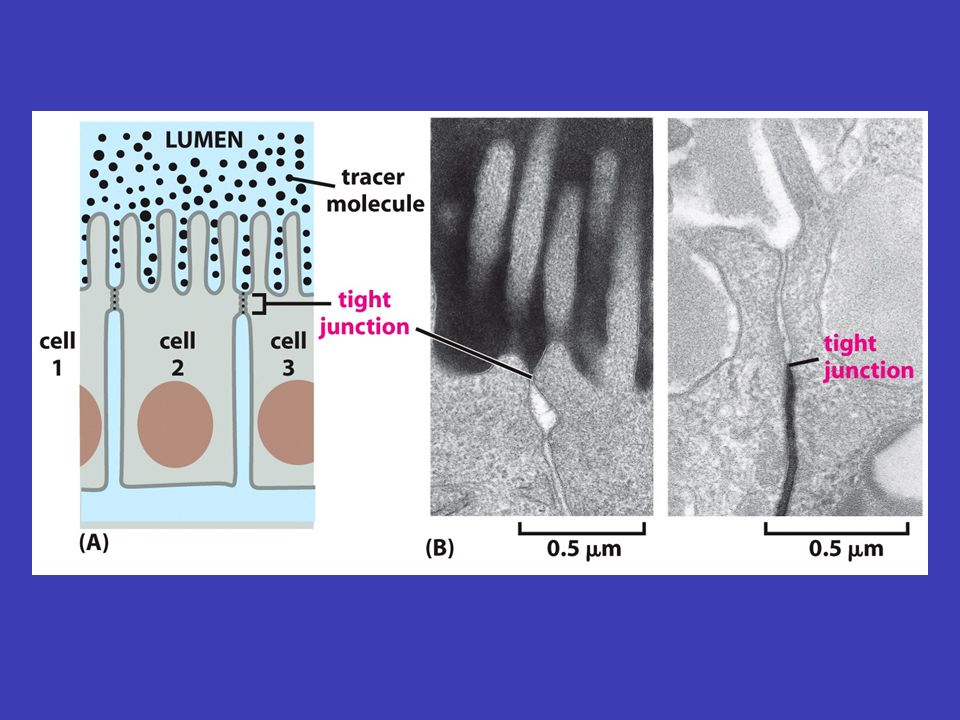
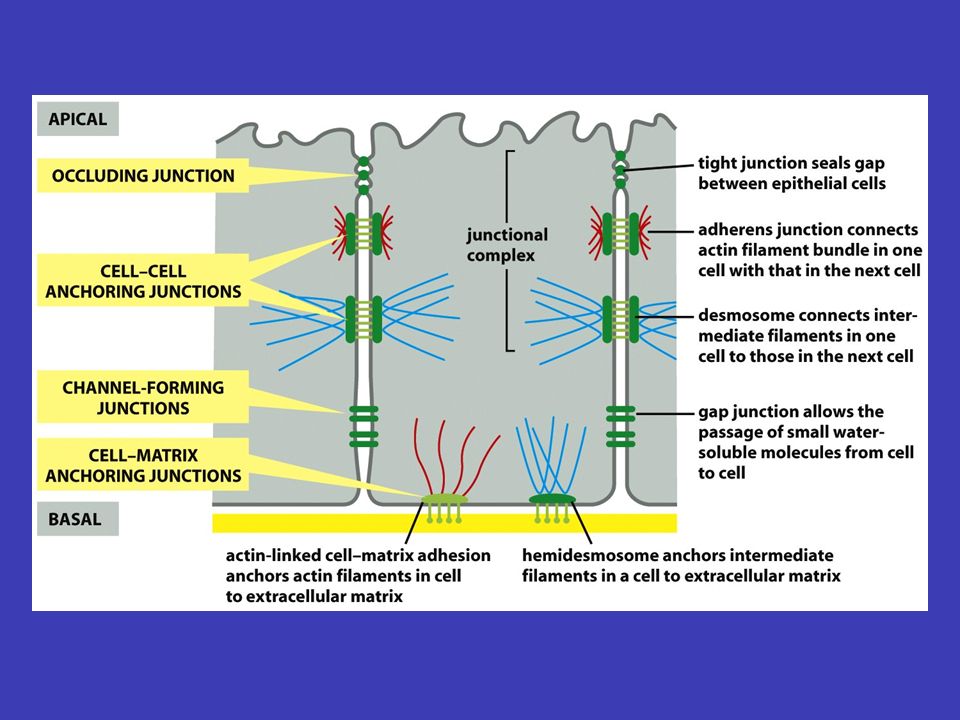
Epitely a mezibuněčné spoje mnohobuněčné vrstvy v epitelech jsou spolu buňky spojeny buněčnou membránou tak, že vytvářejí mnohobuněčné vrstvy (někdy se epitel skládá z více vrstev, např. kůže, jindy z jedné, např. výstelka střeva) apikální bazální bazální membrána/lamina epitel má dvě strany: apikální (povrch je volný a je vystavený vnějšímu okolí tkáně) a bazální (přiléhá k povrchu jiné tkáně, obvykle pojivové) – pod bazálním povrchem epitelu leží tuhá vrstva extracelulární matrix, nazývaná bazální membrána/lamina, která dává epitelu oporu základní funkcí epitelu je oddělovat bazální a apikální stranu a umožňovat mezi nimi kontrolovanou výměnu různých látek (např. vstřebávání složek potravy ve střevě přes střevní epitel)
 apikální bazální bazální membrána/lamina epitel má dvě strany: apikální (povrch je volný a je vystavený vnějšímu okolí tkáně) a bazální (přiléhá k povrchu jiné tkáně, obvykle pojivové) – pod bazálním povrchem epitelu leží tuhá vrstva extracelulární matrix, nazývaná bazální membrána/lamina, která dává epitelu oporu základní funkcí epitelu je oddělovat bazální a apikální stranu a umožňovat mezi nimi kontrolovanou výměnu různých látek (např. vstřebávání složek potravy ve střevě přes střevní epitel).")
49
Obnova tkání buněčné komunikaciselektivní mezibuněčné adhezi „buněčné paměti“ buňky většiny tkání průběžně umírají a jsou zase nahrazovány jinými – organizace tkáně zůstává ovšem zachována díky buněčné komunikaci, selektivní mezibuněčné adhezi a „buněčné paměti“ terminálně diferencované kmenových buněk některé buňky (např. červené krvinky) jsou tzv. terminálně diferencované (nemohou se dělit), a proto vznikají z tzv. kmenových buněk, které nejsou plně diferencovány a mohou se neomezeně dělit z jedné kmenové buňky může vzniknout více buněčných typů (např. lymfocyt, červená krvinka, neutrofil, buňky tvořící krevní destičky atd.)
 jsou tzv. terminálně diferencované (nemohou se dělit), a proto vznikají z tzv. kmenových buněk, které nejsou plně diferencovány a mohou se neomezeně dělit z jedné kmenové buňky může vzniknout více buněčných typů (např. lymfocyt, červená krvinka, neutrofil, buňky tvořící krevní destičky atd.).")
50
Vývin organismu při vývoji dospělého organismu z vajíčka hraje klíčovou roli právě buněčná signalizace a vytváření tkání formování tělního plánu daná buňka byla celou dobu buňkou té či oné tkáně buňky v procesu formování tělního plánu zapínají odlišné sady genů podle své polohy a historie (buňky si „pamatují“, že byly vystaveny daným signálům, protože jejich geny zůstávají zapnuty i po odeznění signálu – a to tak, aby daná buňka byla celou dobu buňkou té či oné tkáně) díky „buněčné paměti“ stačí k vývoji od oplodněného vajíčka k dospělému jedinci jen několik základních signálních mechanismů
 díky „buněčné paměti stačí k vývoji od oplodněného vajíčka k dospělému jedinci jen několik základních signálních mechanismů")
51
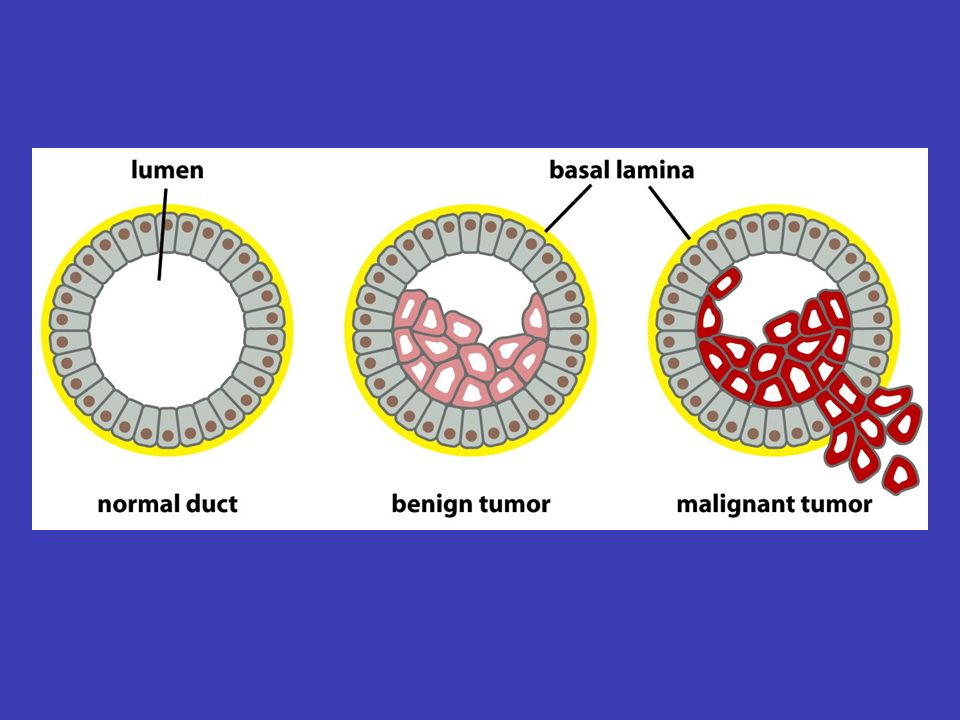
Rakovina onkogeny když dojde k určité souhře mutací (vznikají a zapínají se tzv. onkogeny) v dělící se buňce mnohobuněčného organismu, buňka se zvrhne a získá schopnost donekonečna se dělit a okupovat místa v těle, určená pro jiné buňky – jestliže organismus nezvládne toto množení zastavit (třeba imunitním systémem), vzniká onkologické onemocnění (rakovina) malignímbenigní nádor, vytvořený nekontrolovatelně se množícími buňkami, se stává maligním (opak je benigní) právě svou schopností metastázovat do jiných částí těla a tam úspěšně růst obvykle je zapotřebí 5 až 6 nezávislých mutací, aby mohlo začít nadměrné bujení – a mnoho dalších mutací a okolností je potřeba k tomu, aby se vzniklý nádor stal maligním
 v dělící se buňce mnohobuněčného organismu, buňka se zvrhne a získá schopnost donekonečna se dělit a okupovat místa v těle, určená pro jiné buňky – jestliže organismus nezvládne toto množení zastavit (třeba imunitním systémem), vzniká onkologické onemocnění (rakovina) malignímbenigní nádor, vytvořený nekontrolovatelně se množícími buňkami, se stává maligním (opak je benigní) právě svou schopností metastázovat do jiných částí těla a tam úspěšně růst obvykle je zapotřebí 5 až 6 nezávislých mutací, aby mohlo začít nadměrné bujení – a mnoho dalších mutací a okolností je potřeba k tomu, aby se vzniklý nádor stal maligním.")
53
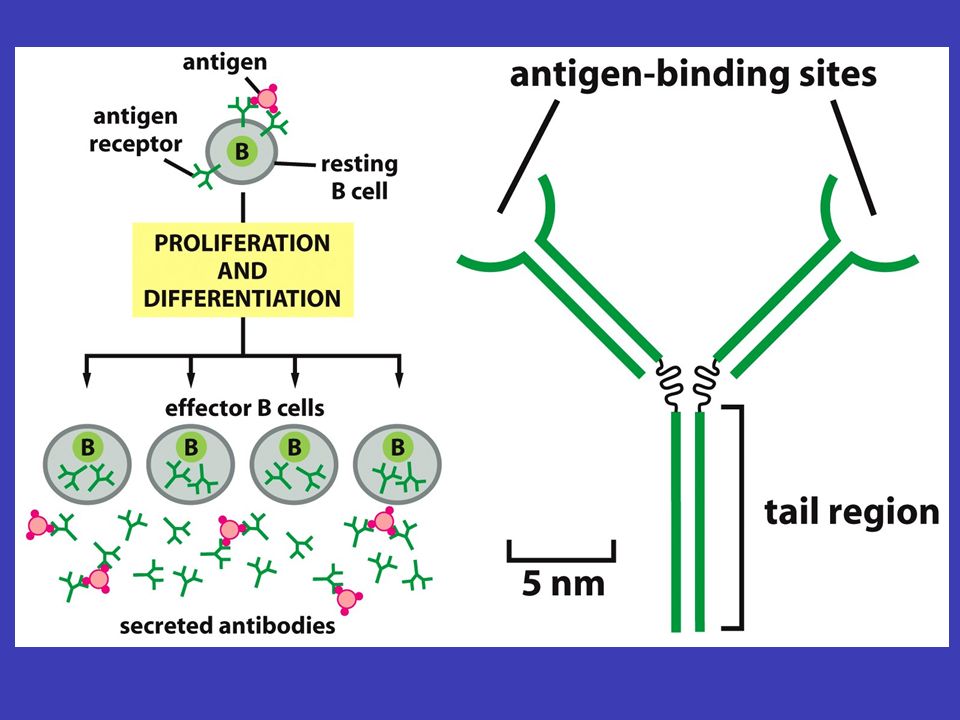
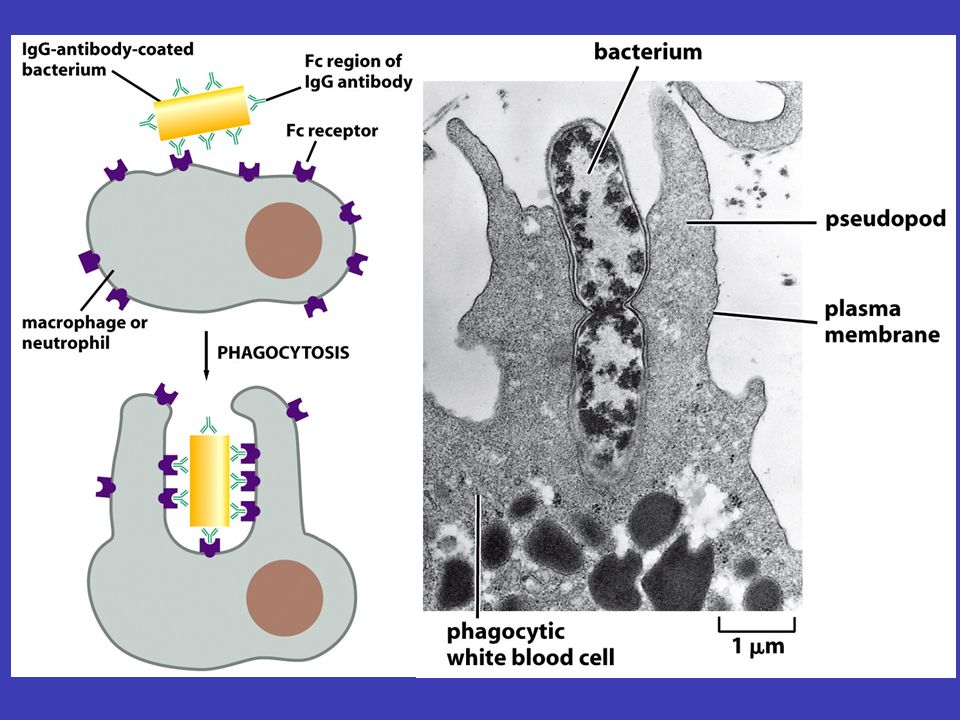
Protilátky a imunita imunita autoimunitní onemocnění jednou z důležitých vlastností organismu je imunita, která brání organismus proti onemocněním, infekcím, ale např. i proti rakovině – imunita se ovšem může vymknout kontrole: autoimunitní onemocnění (např. diabetes 1. typu) protilátkami antigeny zde se z celé složité imunitní odpovědi organismu budeme zabývat pouze protilátkami, jež se vysoce specificky vážou, a tak rozpoznávají, určité molekuly, zvané antigeny. Touto vazbou protilátky buď antigen přímo zneškodní nebo jej určí k likvidaci jinými buňkami B-lymfocyty protilátky jsou v těle vyráběny B-lymfocyty: tyto buňky mají protilátky na svém povrchu a jakmile se na ně naváže daný antigen, začínají vyrábět a vypouštět příslušnou protilátku
 protilátkami antigeny zde se z celé složité imunitní odpovědi organismu budeme zabývat pouze protilátkami, jež se vysoce specificky vážou, a tak rozpoznávají, určité molekuly, zvané antigeny. Touto vazbou protilátky buď antigen přímo zneškodní nebo jej určí k likvidaci jinými buňkami B-lymfocyty protilátky jsou v těle vyráběny B-lymfocyty: tyto buňky mají protilátky na svém povrchu a jakmile se na ně naváže daný antigen, začínají vyrábět a vypouštět příslušnou protilátku.")
Podobné prezentace