Proč cyklové hodiny tikají? A co to vypovídá o evoluci?? (Teoretické přístupy ke studiu BC
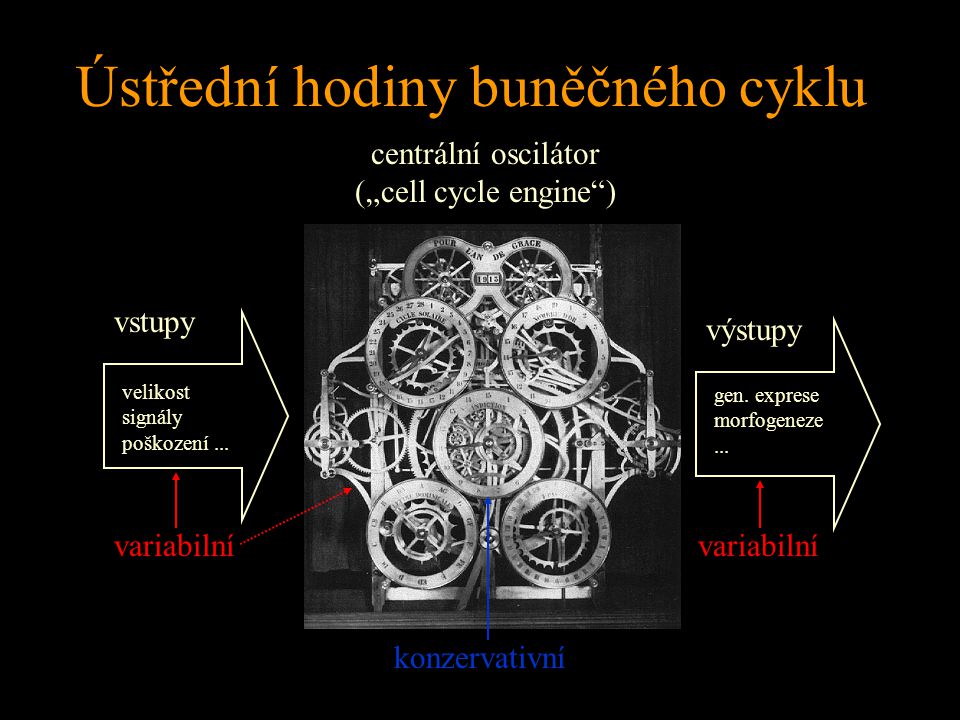
Ústřední hodiny buněčného cyklu vstupy výstupy centrální oscilátor („cell cycle engine“) velikost signály poškození... gen. exprese morfogeneze... konzervativní variabilní
O čem to bude? „Systémová biologie“ (systems biology) Samouspořádání (self-organization) v čase Modely BC
Karl Ludwig von Bertalanffy ,1940 – Theoretische Biologie 1952 – Problems of Life Otec obecné teorie systémů (General Systems Theory)
A System is an assembly of parts or modules with an underlying hidden complexity that governs its structure and the dynamics of its modules.
Systém definovaný soubor (množina) prvků resp. podsystémů a vztahů mezi nimi soubor prvků, který se ve stanovených vztazích projevuje jako celek a může vykazovat zajímavé vlastnosti, např. –stabilitu v širokém rozmezí podmínek vstupů a výstupů –robustnost –...
Příklady vztahů mezi prvky systému Hierarchie Zpětná vazba Redundance...
Biological System “The large numbers of functionally diverse, and frequently multifunctional, sets of components interact selectively and nonlinearly to produce coherent rather than complex behavior.” -- Hiroaki Kitano, “Computational Systems Biology”
(Ivan M. Havel)
Lee Hartwell
Moduly a protokoly – příklad I: Modul: kámen ze stavebnice Protokol: zasunovací spojení
Moduly a protokoly – příklad II: Modul: MAPkinázová kaskáda Protokol: fosforylace cílových proteinů
(Univ. Gent) „produce, manage, mine and model large volumes of (functional genomics) data, and to be able to derive new hypotheses about the function of genes and networks“
Samoorganizace (a periodicita v prostoru a čase)
Jak vůbec lze zajistit periodicitu (v čase nebo prostoru?) Alan TURING Reakčně-difusní model
reakčně-difusní model (
Minimální oscilátor (jeden z mnoha) Je lepší než jiné?? Problém robustnosti! (Ingolia a Murray, Curr. Biol. 2004)
Oscilátor v kontextu tradičního pohledu Tyson and Novak, J. Theor. Biol. 210: , 2001
Dva stavy „cyklových hodin“ StavG1S/M CDK APC ORCprereplik.postreplik. Start Finish (Novak et al., Phil.Trans.R.Soc.Lond.B 353: , 1998) Růst Nedoreplikovaná DNA Neúplná metafáze
Chen et al., Mol. Biol. Cell. 15: , 2004 Východiska matematického modelu
Model minimálního cyklu Jádro: CDK/cykliny + APC Start regulován růstem Finiš regulován dokončením replikace + vřeténka prostřednictvím „aktivátoru“ APC (ACT) Osciluje v širokém rozmezí parametrů! (Novak et al., Phil.Trans.R.Soc.Lond.B 353: , 1998)
Matematický model BC podle Tysona et al.
(demo)
Regulace – jemné ladění Růst působí prostřednictvím akumulace specif. nestabilních molekul z cytoplasmy v jádře – paralogy CDK/cyklinů (Cdk1/Cln) CDK inhibitory (Sic1) a regulační fosforylace Cdc25 „přídavkem“ k zajištění kontroly (checkpoints) Aktivátory APC: alternativní podjednotky – Cdc20 a CDK regulovaná Cdh1
Pokus o rekonstrukci evoluce BC Původní „hodiny“ byly jednodušší Co je nutné a co ne? Pomohou matematické modely??
Evoluční distribuce tradičních „centrálních regulátorů cyklu“ RegulátorA. thalianaS. cerevisiaeH. sapiens CDK1 CDK2 CDKA CDKB CDK4/6 CDKC CDK7 CDKD CDK8 CDKE Cyclin CKI Cdc25 (typic.) Wee1 Rb, E2F atd. ?
Současný pohled na fylogenezi eukaryot: rostliny „mají nárok“ být jiné? (Simpson and Roger, Curr. Biol. 14:R693, 2005)
Ale: co jsou „centrální regulátory“?? Tradiční pojetí: „core cell cycle genes“ – geny hojně studované a asi důležité CDK, cykliny, CKI kinázy a fosfatázy působící na CDK (Cdc25, Wee1) transkripční regulátory (pRB, E2F...) – klavně pokud jsou též onkogeny, případně kontrolují expresi cyklinů a CKI
Co můžeme čekat ve světle modelu?? CDK/cykliny S/M třídy Jádro APC Regulační podjednotky APC (Cdc20/Cdh1) Další podtřídy CDK/cyklinů CDK inhibitory Fosforylace/defosforylace CDK (Cdc25/Wee1) Transkripční regulátory kontrolující „klíčové“ geny (pRB atd.) „Výstupy“ „core cell cycle genes“ e.g. Vandepoele et al., Plant Cell 14: , 2002 Míra konzervace
RegulátorA. thalianaS. cerevisiaeH. sapiens CDK1 CDK2 CDKA CDKB CDK4/6 CDKC CDK7 CDKD CDK8 CDKE RegulátorA. thalianaS. cerevisiaeH. sapiens CDK1/2/4/5/6/A/B CDK7/D/F CDK8/E CDK9/C CDK10/11 Co nacházíme? CDK a cykliny jsou konzervativní! (viz 3. přednáška)
Ostatní regulátory a „regulátory“: APC, Cdc20, Cdh1: „první pohled“ – konzervativní (Capron et al., TiPS 8:83-89, 2003) CDK inhibitory –i u savců VELMI rozmanité (fylogeneze dosud není) Wee1 u rostlin je, Cdc25 není (aspoň ne u Arabidopsis a ne klasická – jiná doménová struktura; Ostreococcus má) mnoho údajů o Rb a trans. faktorech (E2F) – dobře konzervovány kromě kvasinek
Evoluční distribuce centrálních regulátorů BC (bez uvozovek) RegulátorO. tauriA. thalianaS. cerevisiaeH. sapiens CDK (1)6 Cyclin (9 „cyklových“) min.22 APC1-6 každá 1x APC3 2x, ost. 1x každá 1x APC7 11?1 APC8/Cdc APC9 ??1? APC10/Doc1p 1111 APC Cdc Cdh CKI 1?716 Cdc25 (typic.) 1atyp.13 Wee Rb, E2F atd. každá 1xkaždá 1x-3x0každá 1x-6x
Evoluční distribuce centrálních regulátorů buněčného cyklu dává smysl (jen?) ve světle CDK-APC modelu!
Jak tedy asi vypadal ancestrální cyklus? Minimum: CDK + cyklin + APC + Cdc20/Cdh1 zřejmě již od počátku spřaženo s transkripcí