Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Entomologie pro pokročilé 13. Symbionti a patogeny hmyzu
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/ Entomologie pro pokročilé 13. Symbionti a patogeny hmyzu Igor Malenovský Ústav botaniky a zoologie, Přírodovědecká fakulta MU, Brno Entomologické oddělení, Moravské zemské muzeum, Brno

2
Symbionti hmyzu symbióza: vztah hostitele a symbionta
kontinuum od mutualismu, přes komenzalismus po parazitismus vnitrobuněční/mimobuněční viry, bakterie, houby, protista minimálně 9 řádů hmyzu často více druhů symbiontů u jednoho druhu hmyzu – seriální endosymbiotická teorie mnoho symbiontů nedokáže přežívat samostatně mimo hostitele hostitelé zbavení symbiontů často mají sníženou vitalitu, produkují méně potomstva apod.

3
Funkce symbiontů obligátní/fakultativní
často schopnost metabolizovat látky, které hmyz nedokáže strávit (např. rostlinné polymery)/vyrobit (např. vitamíny, steroly, aminokyseliny, toxiny, feromony) většinou hostiteli prospěšní/neutrální v různých tkáních/orgánech (např. specializované mycetomy/bakteriomy, bakteriocyty v tukovém tělese) vliv na rozmnožování (wolbachie) imunosuprese hostitele u parazitoidů (poly-DNA viry a VLP u Braconidae a Ichneumonidae)
/vyrobit (např. vitamíny, steroly, aminokyseliny, toxiny, feromony) většinou hostiteli prospěšní/neutrální. v různých tkáních/orgánech (např. specializované mycetomy/bakteriomy, bakteriocyty v tukovém tělese) vliv na rozmnožování (wolbachie) imunosuprese hostitele u parazitoidů (poly-DNA viry a VLP u Braconidae a Ichneumonidae)")
4
Symbionti v trávícím traktu
zejména u xylofágních druhů hmyzu (živících se dřevem) problematické trávení celulózy: obrovské molekuly složené z mnoha tisíc glukózových jednotek, seřazené do mikrofibril, řetězce blízko sebe, ve vodě nerozpustné – energeticky náročná hydrolýza
 problematické trávení celulózy: obrovské molekuly složené z mnoha tisíc glukózových jednotek, seřazené do mikrofibril, řetězce blízko sebe, ve vodě nerozpustné – energeticky náročná hydrolýza.")
5
Struktura buněčné stěny rostlin
kombinace s rozvětvenými řetězci hemicelulózy

6
Struktura dřeva lignifikace: řetězce celulózy a hemicelulózy uložené v amorfní pryskyřici z fenolických polymerů: ligninu s extrémně stabilními vazbami

7
Trávení dřeva a rostlinných buněčných stěn
nutná synergická aktivita několika typů enzymů: endoglukanázy – štěpí řetězce celulózy na náhodných místech uprostřed exoglukanázy – štěpí volné konce celulózových řetězců enzymy štěpící hemicelulózu enzymy rozbíjející ligno-celulózový komplex enzymy přítomny zj. u některých bakterií (aktinomycety), prvoků a hub nízká nutriční hodnota dřeva (vysoký poměr C/N), nedostatek důležitých živin (aminokyseliny, vitamíny, steroly)
, prvoků a hub. nízká nutriční hodnota dřeva (vysoký poměr C/N), nedostatek důležitých živin (aminokyseliny, vitamíny, steroly)")
8
Trávení lignocelulózy u termitů
komplexní proces zahrnující aktivitu termitů a pestré mikroflóry zadní části jejich střeva rozmělnění dřeva kusadly a proventrikulem na mikroskopické části – zvětšení reakčního povrchu endoglukanázy v sekretu slinných žláz a epitelu mezodea extrémně zvětšené proctodeum – potrava setrvává déle a v anaerobním prostředí (fermentační komora), obrovský povrch Buchner 1928 Thoracotermes macrothorax Potosia cuprea Tipula flaveolineata
, obrovský povrch. Buchner Thoracotermes. macrothorax. Potosia. cuprea. Tipula. flaveolineata.")
9
Fylogeneze termitů Aanen & Boomsma 2005

10
Trávení lignocelulózy u termitů
„nižší termiti“: fagocytóza natrávených dřevních částeček symbiotickými bičíkovci (Trychonympha agilis, Dinenympha) vyplňujícími značnou část střeva, na povrchu a uvnitř prvoků je pestrá mikroflóra symbiotických bakterií Brune 2009
 vyplňujícími značnou část střeva, na povrchu a uvnitř prvoků je pestrá mikroflóra symbiotických bakterií. Brune")
11
Trávení lignocelulózy u termitů
Termitidae (s výjimkou Macrotermitinae – houbové „zahrádky“): celulolýza symbiotickými bakteriemi na povrchu epitelu proctodea (Fibrobacteres, Spirochetes) trávení zdravého/rozloženého dřeva, půdy, výkalů Termitinae: extrémně alkalické prostředí v trávícím traktu (pH=12) Brune 2009
: celulolýza symbiotickými bakteriemi na povrchu epitelu proctodea (Fibrobacteres, Spirochetes) trávení zdravého/rozloženého dřeva, půdy, výkalů. Termitinae: extrémně alkalické prostředí v trávícím traktu (pH=12) Brune")
12
metanogenní archea redukují CO2 na metan
bičíkovci depolymerizují celulózu na jednodušší cukry, které jsou bakteriemi fermentovány na organické kyseliny (resorbované epitelem střeva), vodík a CO2 metanogenní archea redukují CO2 na metan homoacetogenní bakterie (spirochety) redukují CO2 na acetát nutné anaerobní /mikroaerobní prostředí vs. gradienty kyslíku ve střevě Brune 2009
, vodík a CO2. metanogenní archea redukují CO2 na metan. homoacetogenní bakterie (spirochety) redukují CO2 na acetát. nutné anaerobní /mikroaerobní prostředí vs. gradienty kyslíku ve střevě. Brune")
13
Trávení lignocelulózy u termitů
adaptace na nízký obsah N ve dřevě: recyklace dusíku odpadní produkty Malpighiho žláz (močovina a kys. močová) nejsou vylučovány ven z těla, ale mineralizovány střevní mikroflórou na amonné ionty, které jsou začleněny do biomasy mikroorganismů mikroorganismy jsou částečně termity tráveny: proktodeální trofalaxe – krmení jiných jedinců v kolonii výkaly konzumace exuvií a mrtvých jedinců 30-60% dusíku v těle termitů zřejmě pochází z fixace vzdušného dusíku některými bakteriemi podobně Blattodea: Cryptocercidae
 nejsou vylučovány ven z těla, ale mineralizovány střevní mikroflórou na amonné ionty, které jsou začleněny do biomasy mikroorganismů. mikroorganismy jsou částečně termity tráveny: proktodeální trofalaxe – krmení jiných jedinců v kolonii výkaly. konzumace exuvií a mrtvých jedinců % dusíku v těle termitů zřejmě pochází z fixace vzdušného dusíku některými bakteriemi. podobně Blattodea: Cryptocercidae.")
14
Trávení lignocelulózy u termitů
Brune 2009

15
Endosymbionti v trávícím traktu jiného hmyzu
specializovaná mikroflóra zj. u xylofágních, detritofágních a humivorních taxonů, např. Coleoptera: Scarabaeidae (štětičkovité chitinózní výrůstky ve střevě), Diptera: Tipulidae, Blattodea, Orthoptera: Gryllidae celulolytické a hemicelulolytické bakterie, metanogenní archea, anaerobní nálevníci s metanogenními endosymbionty mikroflóru larvy hmyzu získávají de novo po vylíhnutí z vajíčka i každém svlékání (požírání exuvií, u termitů přenos mezi generacemi koprofágií a proktodeální trofalaxí)
, Diptera: Tipulidae, Blattodea, Orthoptera: Gryllidae. celulolytické a hemicelulolytické bakterie, metanogenní archea, anaerobní nálevníci s metanogenními endosymbionty. mikroflóru larvy hmyzu získávají de novo po vylíhnutí z vajíčka i každém svlékání (požírání exuvií, u termitů přenos mezi generacemi koprofágií a proktodeální trofalaxí)")
16
Endosymbionti u krevsajícího hmyzu
dodávají živiny, které chybí v potravě (např. vitamíny B-komplex) vliv na fertilitu a přenos patogenů např. Glossinidae – různé druhy proteobakterií ve střevě, hemolymfě, tukovém tělese a vaječnících: Wigglesworthia glossinidia – primární intracelulární symbiont v bakteriomu tvaru U ve stomodeu Sodalis glossinidis – sekundární symbiont v epiteliálních buňkách mezodea Wolbachia – pohlavní orgány
 vliv na fertilitu a přenos patogenů. např. Glossinidae – různé druhy proteobakterií ve střevě, hemolymfě, tukovém tělese a vaječnících: Wigglesworthia glossinidia – primární intracelulární symbiont v bakteriomu tvaru U ve stomodeu. Sodalis glossinidis – sekundární symbiont v epiteliálních buňkách mezodea. Wolbachia – pohlavní orgány.")
17
Endosymbionti vrtulí: Enterobacteriaceae
Diptera: Tephritidae (např. Rhagoletis pomonella, Ceratitis capitata, Anastrepha ludens) fakultativně jako biofilm na provrchu trávící trubice Enterobacter agglomerans, Klebsiella oxytoca přirozený výskyt v prostředí a na živných rostlinách degradace purinů: přísun dusíku ve využitelné formě Hoy 2010
 fakultativně jako biofilm na provrchu trávící trubice. Enterobacter agglomerans, Klebsiella oxytoca. přirozený výskyt v prostředí a na živných rostlinách. degradace purinů: přísun dusíku ve využitelné formě. Hoy")
18
Endosymbionti mšic: Buchnera aphidicola
proteobakterie (Enterobacteriaceae), intracelulárně v bakteriocytech většiny mšic (1 mšice: až 6 mil. bakterií) asociace milionu let, vertikální přenos z matky na potomstvo, koevoluce dodává esenciální aminokyseliny (tryptofan, leucin – geny na plasmidech), nepřítomnost symbionta způsobuje sterilitu, vliv na přenos patogenů (symbionin chránící virové kapsidy) mnohonásobně (až 200x) amplifikovaný, ale malý genom (600 genů, 650 kb): neesenciální aminokyseliny (glutamin), lipopolysacharidy a další látky dodává mšice bakterii – obligátní vztah (analogie organel)
, intracelulárně v bakteriocytech většiny mšic (1 mšice: až 6 mil. bakterií) asociace milionu let, vertikální přenos z matky na potomstvo, koevoluce. dodává esenciální aminokyseliny (tryptofan, leucin – geny na plasmidech), nepřítomnost symbionta způsobuje sterilitu, vliv na přenos patogenů (symbionin chránící virové kapsidy) mnohonásobně (až 200x) amplifikovaný, ale malý genom (600 genů, 650 kb): neesenciální aminokyseliny (glutamin), lipopolysacharidy a další látky dodává mšice bakterii – obligátní vztah (analogie organel)")
19
Endosymbionti mšic: Buchnera aphidicola
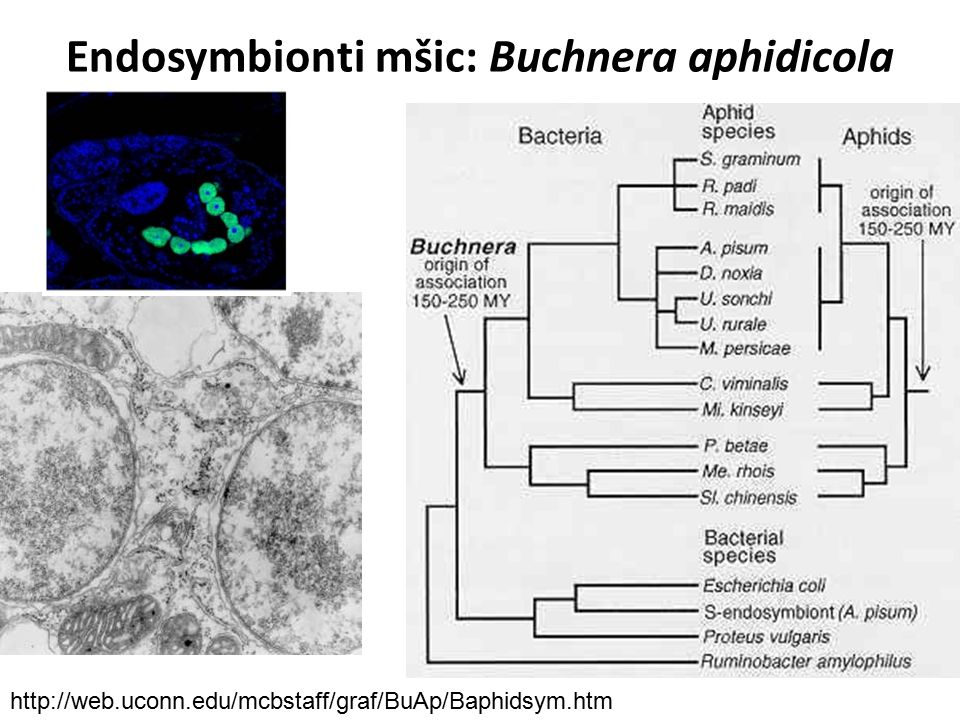
20
Endosymbionti mer: Carsonella rudii
proteobakterie, intracelulárně v bakteriocytech všech mer (1 zisk, koevoluce) nejmenší zatím známý bakteriální genom (160 kb), nejvíce A-T párů (83.4 %) další sekundární symbionti v bakteriocytech, získaní později (Enterobacteriaceae) – zřejmě komplementární metabolická funkce Moran & Sloan 2012
 nejmenší zatím známý bakteriální genom (160 kb), nejvíce A-T párů (83.4 %) další sekundární symbionti v bakteriocytech, získaní později (Enterobacteriaceae) – zřejmě komplementární metabolická funkce. Moran & Sloan")
21
Dvojí endosymbionti křísů
2 druhy endosymbiontů v bakteriomech u taxonů živících se sáním xylému (Cicadellidae: Cicadellinae, Cicadidae): Sulcia muelleri (Bacteroidetes): produkce esenciálních aminokyselin Baumannia cicadellinicola (Proteobact. : produkce vitamínu B,…) Moran 2007
: Sulcia muelleri (Bacteroidetes): produkce esenciálních aminokyselin. Baumannia cicadellinicola (Proteobact. : produkce vitamínu B,…) Moran")
22
Dvojí endosymbionti křísů
Moran 2007

23
Červci – endosymbionti v endosymbiontech
Pseudococcidae (Planococcus citri) γ-proteobakterie (Moraniella) uvnitř β-proteobakterií (Tremblaya) v bakteriocytech von Dohlen et al. 2001
 γ-proteobakterie (Moraniella) uvnitř β-proteobakterií (Tremblaya) v bakteriocytech. von Dohlen et al")
24
„Ultrasobecké“ bakterie
nejčastěji Wolbachia: α-proteobakterie, Gram-negativní, vláknovitý tvar objeveno 1924 v buňkách Culex pipiens, popsáno jako Wolbachia pipientis Hertig, 1936 (Rickettsiales) vnitrobuněční symbionti potvrzení asi u 20-75% druhů členovců a hlístů (PCR), často jen v pohlavních orgánech (vaječníky, testes), u některých druhů ale i v jiných tkáních společný předek wolbachií: mil. let (společný předek členovců 200 mil. let), fylogeneze obou skupin si neodpovídají – horizontální přenos
 vnitrobuněční symbionti potvrzení asi u 20-75% druhů členovců a hlístů (PCR), často jen v pohlavních orgánech (vaječníky, testes), u některých druhů ale i v jiných tkáních. společný předek wolbachií: mil. let (společný předek členovců 200 mil. let), fylogeneze obou skupin si neodpovídají – horizontální přenos.")
25
Ultrasobecké bakterie
přenos z infikované matky do vajíčka (samci nepřenášejí) experimentální přenos injikováním cytoplasmy infikovaného vajíčka do zdravého vajíčka původce cytoplasmatické inkompatibility pohlavních buněk (Yen & Barr 1971) změna poměru pohlaví: indukce partenogeneze (thelytokie), feminizace, male killing (Stouthamer 1990) vajíčko chalcidky Trichogramma kaykai s wolbachiemi (Stouthamer 2009)
 experimentální přenos injikováním cytoplasmy infikovaného vajíčka do zdravého vajíčka. původce cytoplasmatické inkompatibility pohlavních buněk (Yen & Barr 1971) změna poměru pohlaví: indukce partenogeneze (thelytokie), feminizace, male killing (Stouthamer 1990) vajíčko chalcidky Trichogramma kaykai. s wolbachiemi (Stouthamer 2009)")
26
Cytoplasmatická inkompatibilita
neslučitelnost spermií a vajíčka, typicky v případě infikovaný samec + neinfikovaná samice/nesoucí jiný kmen wolbachie – smrt zygoty mechanismus – modifikace samčích chromozomů, narušení synchronie mitózy zygoty již v raném stádiu mj. u komárů, octomilek, zavíječů, brouků (Tribolium confusum, Hypera postica), chalcidek (Nasonia), křísů (Laodelphax striatella), roztočů (svilušek) a dalších Wolbachia pipientis, Cardinium (u chalcidek rodu Encarsia) úplná, většinou ale částečná ( %, rozmnožování některých jedinců je úspěšné kvůli nedokonalému přenosu Wolbachie do pohlavních buněk nebo antibiotikám/teplotě >30°C v prostředí) jednosměrná/obousměrná, superinfekce wolbachie se v populaci rychle šíří – infikované samičky převládnou teoretické modely: vertikální přenos z matky na potomstvo, žádný negativní vliv wolbachie na nositele možný vliv na speciaci hmyzu – podpora reprodukční izolace (sterilita hybridů, inkompatibilita linií se dvěma různými kmeny wolbachií, thelytokie)
, chalcidek (Nasonia), křísů (Laodelphax striatella), roztočů (svilušek) a dalších. Wolbachia pipientis, Cardinium (u chalcidek rodu Encarsia) úplná, většinou ale částečná ( %, rozmnožování některých jedinců je úspěšné kvůli nedokonalému přenosu Wolbachie do pohlavních buněk nebo antibiotikám/teplotě >30°C v prostředí) jednosměrná/obousměrná, superinfekce. wolbachie se v populaci rychle šíří – infikované samičky převládnou. teoretické modely: vertikální přenos z matky na potomstvo, žádný negativní vliv wolbachie na nositele. možný vliv na speciaci hmyzu – podpora reprodukční izolace (sterilita hybridů, inkompatibilita linií se dvěma různými kmeny wolbachií, thelytokie)")
27
Určení pohlaví u hmyzu velmi rozmanité v rámci řádů až druhů (Musca domestica) chromozomální určení pohlaví: XY (samci), XX (samice): většina skupin XO (samci), XX (samice): řada skupin ZZ (samci), ZW (samice): Lepidoptera, Trichoptera časté translokace pohlavních chromozomů (neo-Y) a duplikace (např. blecha Xenopsylla cheopis: X1X2Y (samec), X1X1X2X2 (samice)) některé druhy nemají pohlavní chromozomy, jen dominantní geny pro samce: M/+ (samci), +/+ (samice) nebo epigenetické ovlivnění pohlaví genem matky nezávisle na genotypu otce haplodipoidní určení pohlaví (arrhenotokie): samice z oplozených vajíček (2n), samici z neoplozených (1n): Hymenoptera, Thysanoptera, některá Hemiptera (Aleurodidae, Margarodidae) a Coleoptera (Micromalthus debilis) vzácně vliv environmentálních faktorů (např. teplota: Drosophila) extrémně vzácně hermafroditismus (3 druhy červců z rodu Icerya, rozmnožování samooplozením)
, XX (samice): většina skupin. XO (samci), XX (samice): řada skupin. ZZ (samci), ZW (samice): Lepidoptera, Trichoptera. časté translokace pohlavních chromozomů (neo-Y) a duplikace (např. blecha Xenopsylla cheopis: X1X2Y (samec), X1X1X2X2 (samice)) některé druhy nemají pohlavní chromozomy, jen dominantní geny pro samce: M/+ (samci), +/+ (samice) nebo epigenetické ovlivnění pohlaví genem matky nezávisle na genotypu otce. haplodipoidní určení pohlaví (arrhenotokie): samice z oplozených vajíček (2n), samici z neoplozených (1n): Hymenoptera, Thysanoptera, některá Hemiptera (Aleurodidae, Margarodidae) a Coleoptera (Micromalthus debilis) vzácně vliv environmentálních faktorů (např. teplota: Drosophila) extrémně vzácně hermafroditismus (3 druhy červců z rodu Icerya, rozmnožování samooplozením)")
28
Ovlivnění poměru pohlaví bakteriemi - feminizace
změna pohlaví geneticky determinovaných samců na samice nebo neplodné pseudo-samice (Wolbachia, Cardinium) jen u skupin, kde samice jsou heterogametické pohlaví (např. motýli, stínky: samice ZW, samci ZZ) zavíječ Ostrinia furnacalis (Crambidae) – 100% samičí linie, po aplikaci antibiotik se objevili samci mechanismus: vliv aktivity 1 genu, který zablokuje expresi jiných genů, zodpovědných za vývoj androgenní žlázy
 jen u skupin, kde samice jsou heterogametické pohlaví (např. motýli, stínky: samice ZW, samci ZZ) zavíječ Ostrinia furnacalis (Crambidae) – 100% samičí linie, po aplikaci antibiotik se objevili samci. mechanismus: vliv aktivity 1 genu, který zablokuje expresi jiných genů, zodpovědných za vývoj androgenní žlázy.")
29
Partenogeneze u hmyzu partenogeneze: rozmnožování neoplodněnými vajíčky: arrhenotokie – haplodiploidní určení pohlaví, samice se líhnou z oplozených, samci z neoplozených vajíček deuterotokie – obě pohlaví se líhnou z neoplozených vajíček pseudogamie (gynogeneze)– vznikají pouze samičky z oplozených vajíček, předávají se ale jen samičí geny (spermie slouží jen k aktivaci) – u různých skupin, ale jen vzácně pseudogamie (androgeneze) – v části oplodněných vajíček se uplatňuje jen samčí genom (mravenec Wasmannia auropunctata, v kombinaci s gynogenezí) hybridogeneze – páření samic s jiným druhem (Phasmatodea: Bacillus) thelytokie – vznikají pouze samičky z neoplozených vajíček
– vznikají pouze samičky z oplozených vajíček, předávají se ale jen samičí geny (spermie slouží jen k aktivaci) – u různých skupin, ale jen vzácně. pseudogamie (androgeneze) – v části oplodněných vajíček se uplatňuje jen samčí genom (mravenec Wasmannia auropunctata, v kombinaci s gynogenezí) hybridogeneze – páření samic s jiným druhem (Phasmatodea: Bacillus) thelytokie – vznikají pouze samičky z neoplozených vajíček.")
30
Thelytokie důsledky thelytokie:
samice předávají úspěšný genotyp 100 % potomkům produkovány pouze samice (maximalizace nárůstu populace) ušetření energie a času na hledání samce a páření výskyt thelytokie u Hexapoda: relativně vzácně, ale přesto u mnoha skupin (80 čeledí) zejména časté u Thysanoptera, Psocodea, Hemiptera: Sternorrhyncha, Phasmatodea méně časté u Coleoptera (Curculionidae), Lepidoptera (Psychidae), Diptera (Chironomidae) mechanismy: apomixe (chybí meióza) automixe (meióza proběhne, diploidní stav je obnoven duplikací sady chromosomů nebo splynutím 2 buněk
 ušetření energie a času na hledání samce a páření. výskyt thelytokie u Hexapoda: relativně vzácně, ale přesto u mnoha skupin (80 čeledí) zejména časté u Thysanoptera, Psocodea, Hemiptera: Sternorrhyncha, Phasmatodea. méně časté u Coleoptera (Curculionidae), Lepidoptera (Psychidae), Diptera (Chironomidae) mechanismy: apomixe (chybí meióza) automixe (meióza proběhne, diploidní stav je obnoven duplikací sady chromosomů nebo splynutím 2 buněk.")
31
Thelytokie fakultativní:
tychopartenogeneze: malý počet (4-8 %) neoplodněných vajíček se po nějaké době vyvine v samice: u jinak pohlavních druhů Ephemeroptera, Polyneoptera, Psocodea, Lepidoptera, Diptera) geografická (pouze některé populace v horách, na okraji areálu apod.) cyklická (Hemiptera: Aphidoidea, Hymenoptera: Cynipoidea) obligátní vznik thelytokie: hybridizací nezávislý na hybridizaci indukce endosymbiotickými bakteriemi
 neoplodněných vajíček se po nějaké době vyvine v samice: u jinak pohlavních druhů Ephemeroptera, Polyneoptera, Psocodea, Lepidoptera, Diptera) geografická (pouze některé populace v horách, na okraji areálu apod.) cyklická (Hemiptera: Aphidoidea, Hymenoptera: Cynipoidea) obligátní. vznik thelytokie: hybridizací. nezávislý na hybridizaci. indukce endosymbiotickými bakteriemi.")
32
Indukce partenogeneze bakteriemi
zj. u parazitoidů (Hymenoptera, Diptera: Tachinidae), Thysanoptera (Franklinothrips vespiformis), chvostoskoků (Folsomia candida) a roztočů s původně haplodiploidním určením pohlaví mechanismus – wolbachie zdvojnásobí sadu chromosomů u haploidních vajíček při první mitóze kmeny infikované bakteriemi (zj. Wolbachia, ale i Cardinium, Rickettsia) jsou thelytokní, po aplikaci antibiotik nebo vysokých teplot se rodí i samci vznik několikrát nezávisle v různých liniích, horizontální přenos mezi liniemi v populaci monomorfní thelytokie nebo dimorfní thelytokie a arrhenotokie (Trichogramma spp.)
, Thysanoptera (Franklinothrips vespiformis), chvostoskoků (Folsomia candida) a roztočů s původně haplodiploidním určením pohlaví. mechanismus – wolbachie zdvojnásobí sadu chromosomů u haploidních vajíček při první mitóze. kmeny infikované bakteriemi (zj. Wolbachia, ale i Cardinium, Rickettsia) jsou thelytokní, po aplikaci antibiotik nebo vysokých teplot se rodí i samci. vznik několikrát nezávisle v různých liniích, horizontální přenos mezi liniemi. v populaci monomorfní thelytokie nebo dimorfní thelytokie a arrhenotokie (Trichogramma spp.)")
33
Zabíjení samců (male-killing)
známo u 5 řádů hmyzu (Hemiptera: Lygaeidae, Coleoptera: Coccinellidae, Lepidoptera: Nymphalidae, Hymenoptera, Diptera) a 2 druhů roztočů rozmanití původci: α- a γ-proteobakterie, mykoplasmy, flavobakterie, microsporidie – vznik mnohokrát nezávisle 2 strategie: samci umírají v posledním larválním stádiu (symbionti se v těle mohou namnožit, horizontální přenos): mikrosporidie u komárů samci jsou ničeni během embryogeneze (není horizontální přenos, ale zvýšená fitness samice): bakterie výhody pro hmyz: snížení inbreedingu (páření s „bratry“) a více zdrojů pro samice
 a 2 druhů roztočů. rozmanití původci: α- a γ-proteobakterie, mykoplasmy, flavobakterie, microsporidie – vznik mnohokrát nezávisle. 2 strategie: samci umírají v posledním larválním stádiu (symbionti se v těle mohou namnožit, horizontální přenos): mikrosporidie u komárů. samci jsou ničeni během embryogeneze (není horizontální přenos, ale zvýšená fitness samice): bakterie. výhody pro hmyz: snížení inbreedingu (páření s „bratry ) a více zdrojů pro samice.")
34
Zabíjení samců u slunéček (např. Adalia bipunctata, Harmonia axyridis): poměr pohlaví při oplodnění 1:1, ale mortalita (samčích) vajíček 50 % - mrtvá vajíčka jsou zkonzumována mladými samičími larvami – výhoda oproti neinfikovaným samicím (přežijí déle než najdou první efemerní potravu - mšice) poměr pohlaví 1:1 až 1:2 ve prospěch samic v závislosti na prevalenci bakterií (Spiroplasma): 5-50 % slabý pohlavní výběr, nízká investice samců do rozmnožování (max. 3 spermatofory) Majerus 2009
: poměr pohlaví při oplodnění 1:1, ale mortalita (samčích) vajíček 50 % - mrtvá vajíčka jsou zkonzumována mladými samičími larvami – výhoda oproti neinfikovaným samicím (přežijí déle než najdou první efemerní potravu - mšice) poměr pohlaví 1:1 až 1:2 ve prospěch samic v závislosti na prevalenci bakterií (Spiroplasma): 5-50 % slabý pohlavní výběr, nízká investice samců do rozmnožování (max. 3 spermatofory) Majerus")
35
Zabíjení samců babočky Acraea encedon, Acraea encedana, Hypolimnas bolina: až 95 % samic v populaci samci jsou limitující pro rozmnožování, mnoho samic umírá bez páření reverze pohlavních strategií – panenské samice tvoří roje a soupeří o samce Acraea encedon

36
Další vlivy ultrasobeckých bakterií
snížení genetické diverzity organel (např. mitochondrií) přenášených v cytoplasmě z matky na vajíčko vliv na molekulárně fylogenetické analýzy založené na mt-genech větší rezistence hostitele vůči některým virům využití v praxi (biologická ochrana): parazitoidi, slunéčka – zvýšení fitness samic, redukce nákladů cytoplasmatická inkompatibilita – vypouštění nekompatibilních samců do přírody přenos určitých genů v populaci (stačí nízký počet jedinců): inhibice přenosun malárie, dengue
 přenášených v cytoplasmě z matky na vajíčko. vliv na molekulárně fylogenetické analýzy založené na mt-genech. větší rezistence hostitele vůči některým virům. využití v praxi (biologická ochrana): parazitoidi, slunéčka – zvýšení fitness samic, redukce nákladů. cytoplasmatická inkompatibilita – vypouštění nekompatibilních samců do přírody. přenos určitých genů v populaci (stačí nízký počet jedinců): inhibice přenosun malárie, dengue.")
37
Patogeny hmyzu viry a mikroorganismy (bakterie, houby, protista)
původce chronických/akutních/fatálních chorob často relativně úzké hostitelské spektrum

38
Iridoviry (Iridoviridae)
lineární 2S-DNA, obalené, velké ikozahedrální kapsidy ( nm) světlo se od napadeného hostitele odráží v barevných odlescích daných interferencí od parakrystalů viru 30 typů ve vlhkém prostředí: v larvách Diptera, Coleoptera, Lepidoptera; Isopoda, žáby a ryby prevalence a mortalita v přírodě <1 %, špatné šíření
 světlo se od napadeného hostitele odráží v barevných odlescích daných interferencí od parakrystalů viru. 30 typů ve vlhkém prostředí: v larvách Diptera, Coleoptera, Lepidoptera; Isopoda, žáby a ryby. prevalence a mortalita v přírodě <1 %, špatné šíření.")
39
Cytoplasmic Polyhedrosis Viruses (Reoviridae)
CPV, Cypovirus: lineární 2S-RNA (9 segmentů) v trávícím traktu hmyzu, tvoří velká kulovitá okluzní tělíska v cytoplasmě epiteliálních buněk mezenteronu (jeví se bíle) chronická infekce, zpomalení vývoje larev až smrt často u Lepidoptera a Diptera: Nematocera přenos potravou, široký okruh hostitelů
 v trávícím traktu hmyzu, tvoří velká kulovitá okluzní tělíska v cytoplasmě epiteliálních buněk mezenteronu (jeví se bíle) chronická infekce, zpomalení vývoje larev až smrt. často u Lepidoptera a Diptera: Nematocera. přenos potravou, široký okruh hostitelů.")
40
Entomopoxviry (Poxviridae)
lineární 2S-DNA, obalené, velké viriony (150x300 nm), příbuzné viru neštovic v cytoplasmě různých tkání Coleoptera, Lepidoptera, Diptera, Orthoptera (30 izolovaných typů) akutní, smrtelná choroba
, příbuzné viru neštovic. v cytoplasmě různých tkání Coleoptera, Lepidoptera, Diptera, Orthoptera (30 izolovaných typů) akutní, smrtelná choroba.")
41
Askoviry (Ascoviridae)
kruhová 2S-DNA, velké viriony (130x400 nm) ledvinovitého až tyčinkovitého tvaru pouze u Lepidoptera: Noctuidae, přenášeno parazitickými Hymenoptera chronická, smrtelná choroba viriony se hromadí ve váčcích hemolymfě, která se mění z průhledně zelené na mléčně bílou
 ledvinovitého až tyčinkovitého tvaru. pouze u Lepidoptera: Noctuidae, přenášeno parazitickými Hymenoptera. chronická, smrtelná choroba. viriony se hromadí ve váčcích hemolymfě, která se mění z průhledně zelené na mléčně bílou.")
42
Bakuloviry (Baculoviridae)
kruhová 2S-DNA, obalené, velké viriony (80-200x280 nm) vysoce infekční, přijímané s potravou, epidemie s až 90% mortalitou v populacích housenek , +- specifické nuclear polyhedrosis viruses (NPV) – hlavně Lepidoptera (přes 500 izolátů), ale i Symphyta, replikace v jádrech, různé tkáně, často mezenteron granulosis viruses (GV) – pouze Lepidoptera, různé tkáně, často v tukovém tělese
 vysoce infekční, přijímané s potravou, epidemie s až 90% mortalitou v populacích housenek , +- specifické. nuclear polyhedrosis viruses (NPV) – hlavně Lepidoptera (přes 500 izolátů), ale i Symphyta, replikace v jádrech, různé tkáně, často mezenteron. granulosis viruses (GV) – pouze Lepidoptera, různé tkáně, často v tukovém tělese.")
43
Bakuloviry (Baculoviridae)
kruhová 2S-DNA, obalené, velké viriony (80-200x280 nm) vysoce infekční, přijímané s potravou, epidemie s až 90% mortalitou v populacích housenek , +- specifické nuclear polyhedrosis viruses (NPV) – hlavně Lepidoptera (přes 500 izolátů), ale i Symphyta, replikace v jádrech, různé tkáně, často mezenteron granulosis viruses (GV) – pouze Lepidoptera, různé tkáně, často v tukovém tělese
 vysoce infekční, přijímané s potravou, epidemie s až 90% mortalitou v populacích housenek , +- specifické. nuclear polyhedrosis viruses (NPV) – hlavně Lepidoptera (přes 500 izolátů), ale i Symphyta, replikace v jádrech, různé tkáně, často mezenteron. granulosis viruses (GV) – pouze Lepidoptera, různé tkáně, často v tukovém tělese.")
44
Využití virů biologická ochrana – většinou však časově omezená – nutná opakovaná aplikace (mikrobiální insekticidy) nejvhodnější jsou bakuloviry (NPV a GV) – hojné a lehce izolovatelné z hostitelů, levná produkce, bezproblémová aplikace (jako postřiky insekticidů), na druhou stranu napadají jen úzký okruh hostitelů a nejdou produkovat masově biotechnologie: AcMNPV k produkci proteinů in vitro – např. modifikované kapsidy virů používané jako vakcíny pro obratlovce Gilpinia hercyniae
 – hojné a lehce izolovatelné z hostitelů, levná produkce, bezproblémová aplikace (jako postřiky insekticidů), na druhou stranu napadají jen úzký okruh hostitelů a nejdou produkovat masově. biotechnologie: AcMNPV k produkci proteinů in vitro – např. modifikované kapsidy virů používané jako vakcíny pro obratlovce. Gilpinia hercyniae.")
45
Entomopatogenní bakterie
lehce kultivovatelné na médiích Bacillus thuringiensis (Bt) – komplex různých typů, běžně v půdě a na povrchu rostlin, ve výkalech hmyzu a součást mikroflóry mezenteronu produkce spor s krystaly bílkovinných endotoxinů, vysoce insekticidní pro některé druhy hmyzu – po sežrání a natrávení obalu rychle (1-2 dny) likvidují buňky mezenteronu součást mnoha komerčních insekticidů: B.t. subsp. kurstaki (Btk): polní a lesní škůdci Lepidoptera B. t. subsp. Israelensis (Bti): larvy komárů a muchniček B.t. subsp. morrisoni (pathovar tenebrionis): Coleoptera transgenní rostliny: Bt-kukuřice a Bt-bavlník Federici 2009
 – komplex různých typů, běžně v půdě a na povrchu rostlin, ve výkalech hmyzu a součást mikroflóry mezenteronu. produkce spor s krystaly bílkovinných endotoxinů, vysoce insekticidní pro některé druhy hmyzu – po sežrání a natrávení obalu rychle (1-2 dny) likvidují buňky mezenteronu. součást mnoha komerčních insekticidů: B.t. subsp. kurstaki (Btk): polní a lesní škůdci Lepidoptera. B. t. subsp. Israelensis (Bti): larvy komárů a muchniček. B.t. subsp. morrisoni (pathovar tenebrionis): Coleoptera. transgenní rostliny: Bt-kukuřice a Bt-bavlník. Federici")
46
Entomopatogenní bakterie
Bacillus sphaericus (Bs) – larvy komárů Paenibacillus popilliae – „milky disease“ larev chroustů Serratia entomophila –proti chroustovi Costelystra zealandica („amber disease“)
 – larvy komárů. Paenibacillus popilliae – „milky disease larev chroustů. Serratia entomophila –proti chroustovi Costelystra zealandica („amber disease )")
47
Entomopatogenní houby
často ve vlhkém prostředí tvoří mycelia, infekce sporami, sporangiemi nebo konidiemi hmyz napadají aktivně růstem skrz kutikulu do tělní dutiny, kolonizace trvá ca dní, končí úmrtím hmyzu některé houby produkují toxiny – smrt do 48 hodin při vlhkosti 90% vytvoří reprodukční struktury a spory významný faktor mortality hmyzu v přírodě, komerčně ale využitelný jen obtížně některé skupiny spíše parazitické - Laboulbeniales

48
Vodní entomopatogenní houby
Coelonomyces (Chytridiomycetes, 80 spp.) – obligátně parazitické houby, pohlavní fáze napadá drobné korýše (Copepoda), nepohlavní larvy komárů (včetně Anopheles gambiae), obrvené spory, celosvětové rozšíření Lagenidium giganteum (Oomycetes) – jednoduchá kultura, napadá larvy komárů jako jediné hostitele, mobilní zoospory, kolonizace 2-3 dny
 – obligátně parazitické houby, pohlavní fáze napadá drobné korýše (Copepoda), nepohlavní larvy komárů (včetně Anopheles gambiae), obrvené spory, celosvětové rozšíření. Lagenidium giganteum (Oomycetes) – jednoduchá kultura, napadá larvy komárů jako jediné hostitele, mobilní zoospory, kolonizace 2-3 dny.")
49
Terestrické entomopatogenní houby
Entomophthorales (Zygomycetes) – mnoho druhů na členovcích, často křísi a ploštice, ale i mouchy, brouci, orthoptera a housenky většinou hostitelsky specifické Conidiobolus, Erynia, Entomophthora (mšice), Zoophthora (mšice, housenky, brouci), Entomophaga (saranče, housenky) obtížně kultivovatelné a komerčně využitelné
 – mnoho druhů na členovcích, často křísi a ploštice, ale i mouchy, brouci, orthoptera a housenky. většinou hostitelsky specifické. Conidiobolus, Erynia, Entomophthora (mšice), Zoophthora (mšice, housenky, brouci), Entomophaga (saranče, housenky) obtížně kultivovatelné a komerčně využitelné.")
50
Terestrické entomopatogenní houby
Hyphomycetes (Deuteromycotina) – bez známé pohlavní fáze, infekčním stádiem jsou konidia většina nyní zařazena do Ascomycota široké spektrum hostitelů, relativně snadná kultivace např. Metarhizium/Metacordyceps anisopliae, Beauveria/Cordyceps bassiana, Paecilomyces fumoso-rosea, Verticillium lecanii k boji proti larvám brouků v půdě, ostruhovníkům na rýži a molicím ve sklenících Federici 2009
 – bez známé pohlavní fáze, infekčním stádiem jsou konidia. většina nyní zařazena do Ascomycota. široké spektrum hostitelů, relativně snadná kultivace. např. Metarhizium/Metacordyceps anisopliae, Beauveria/Cordyceps bassiana, Paecilomyces fumoso-rosea, Verticillium lecanii. k boji proti larvám brouků v půdě, ostruhovníkům na rýži a molicím ve sklenících. Federici")
51
Terestrické entomopatogenní houby
Metarhizium/Metacordyceps anisopliae „green muscardine disease“ Beauveria/Cordyceps bassiana „white muscardine disease“

52
Mikrosporidie (hmyzomorky)
více než 1000 druhů, většina napadá hmyz, zj. Coleoptera, Lepidoptera, Diptera, Orthoptera obligátní vnitrobuněční parazité, nemají mitochondrie (nelze kultivovat in vitro), produkují spory s polárním filamentem infekce pozřením spor/transovariálně, uvnitř cytoplasmy se ze sporoplasmy vytváří plasmodium, posléze velké množství spor onemocnění trvá několik týdnů, napadení populace může být masivní, je ale těžko předvídatelné nelze využít k cílenému biologickému boji
, produkují spory s polárním filamentem. infekce pozřením spor/transovariálně, uvnitř cytoplasmy se ze sporoplasmy vytváří plasmodium, posléze velké množství spor. onemocnění trvá několik týdnů, napadení populace může být masivní, je ale těžko předvídatelné. nelze využít k cílenému biologickému boji.")
53
Mikrosporidie (hmyzomorky)
")
54
Literatura Capinera J.L. (ed.) 2008: Encyclopedia of Entomology. Springer. Hesla: Symbionts of Insects, Symbiosis Between Planthoppers and Microorganisms, Symbiotic Viruses of Parasitic Wasps, Pathogens of Whiteflies Resh V. & Cardé R.T. (ed.) 2009: Encyclopedia of Insects. Academic Press. Hesla: Symbionts aiding digestion, Symbionts-Bacterial, Wolbachia, Sex determination, Parthenogenesis in Insects and Mites, Pathogens of Insects Bourtzis K. & Miller T.A. (eds.) 2003: Insect Symbiosis, Vol : Vol : Vol. 3. CRC Press, Boca Raton. Heming B. S. 2003: Insect Development and Evolution. Cornell University Press. Vega F.E. & Blackwell M. 2005: Insect-Fungal Associations. Ecology and Evolution. Oxford University Press.
 2008: Encyclopedia of Entomology. Springer. Hesla: Symbionts of Insects, Symbiosis Between Planthoppers and Microorganisms, Symbiotic Viruses of Parasitic Wasps, Pathogens of Whiteflies. Resh V. & Cardé R.T. (ed.) 2009: Encyclopedia of Insects. Academic Press. Hesla: Symbionts aiding digestion, Symbionts-Bacterial, Wolbachia, Sex determination, Parthenogenesis in Insects and Mites, Pathogens of Insects. Bourtzis K. & Miller T.A. (eds.) 2003: Insect Symbiosis, Vol : Vol : Vol. 3. CRC Press, Boca Raton. Heming B. S. 2003: Insect Development and Evolution. Cornell University Press. Vega F.E. & Blackwell M. 2005: Insect-Fungal Associations. Ecology and Evolution. Oxford University Press.")
Podobné prezentace












 - Jsou to nejrozšířenější organické látky, tvoří největší podíl organické hmoty na Zemi. Funkce: zásobní látky v organismu.>")






