Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Bioelektrické jevy a jejich měření 5. přednáška

2
Obecná elektrofyziologie tkání Tkáň = soubor buněk stejného embryonálního původu sdružené k tomu, aby plnily nějakou funkci; tkáň může obsahovat všechny buňky typově stejné, nebo i více různých typů. Nositelem specializované funkce bývá jeden typ buněk, další typy (často i morfologicky a funkčně poněkud odlišné) tuto funkci podporují. A) Tkáň tvořená apolárními buňkami umístěním obvodu nenaměříme žádný potenciálový rozdíl, dokonce ani tehdy, jsou-li buňky náhodně polarizované a oscilující (vektorový součin je = 0) B) Tkáň složená z buněk tvořících dipóly umístěním obvodu naměříme v různých místech určitý potenciálový rozdíl, ať už tkáň tvořená dipóly umístěnými v sérii nebo paralelně + - + - + - + - + - + - 1 dipól + - + - + - + - suma dipólů Typická polarizovaná tkáň = žabí kůže. Žába má schopnost aktivně transportovat Na + : je to hlavní extracelulární kation a ve sladké vodě je ho málo, žába by o něj přicházela. Kolik Na + (gramionů iontů) projde přes membránu za nějaký čas (napěťový zámek a měření toků iontů!): Na net = I. t F
 tuto funkci podporují. A) Tkáň tvořená apolárními buňkami umístěním obvodu nenaměříme žádný potenciálový rozdíl, dokonce ani tehdy, jsou-li buňky náhodně polarizované a oscilující (vektorový součin je = 0) B) Tkáň složená z buněk tvořících dipóly umístěním obvodu naměříme v různých místech určitý potenciálový rozdíl, ať už tkáň tvořená dipóly umístěnými v sérii nebo paralelně dipól suma dipólů Typická polarizovaná tkáň = žabí kůže. Žába má schopnost aktivně transportovat Na + : je to hlavní extracelulární kation a ve sladké vodě je ho málo, žába by o něj přicházela. Kolik Na + (gramionů iontů) projde přes membránu za nějaký čas (napěťový zámek a měření toků iontů!): Na net = I. t F.")
3
Elektrický potenciálový rozdíl lze zaznamenat i v apolární tkáni, pokud je část buněk poškozena. Mezi poškozenou a intaktní částí tkáně je potenciálový rozdíl díky difusním potenciálům. Z poškozené buňky/buněk se uvolňují ionty, mění se jejich extracelulární koncentrace, difundují extracelulárním prostředím... + - + - Polarizace tkání a schopnost buněk aktivně transportovat ionty např. z apikální strany buňky na bazální má značný fyziologický význam. Klasika = střevo... ale např. pro normální vývoj plic: buňky výstelek dýchacích cest plodu secernují chloridy, s nimiž je „tažena“ do dýchacích cest i voda. Několik dní po narození tento transport vody do cest dýchacích výrazně klesá, už před porodem a těsně po něm plicní epitel naopak tekutinu absorbuje, aby mohlo docházet k výměně dýchacích plynů. Za tuto změnu je velkou měrou odpovědný aktivní transport Na + přes plicní epitel. Roste hladina mRNA pro epiteliální Na + IKs a pro Na + /K + ATPAsu aj. apikální strana bazolaterální strana -agonista Všechny měkké tkáně obsahují dlouhé řetězce proteinů jako kolagen, elastin či keratin – molekuly s pravidelnou strukturou opakujících se podjednotek. Tkáně jako ligamenta, fascie, šlachy obsahují denzní plošné ( -sheets) sítě těchto proteinů, zejména kolagenu. Chrupavky se skládají z kolagenu a proteoglykanů, kost je v podstatě kalcifikovaná kolagenní struktura. Všechny tyto útvary mají společné to, že pokud jsou vystaveny mechanickému stresu (komprese, natažení, zkroucení aj.), vznikají na jejich vnitřních i vnějších površích piezoelektrické náboje a potenciály. Např. na kosti vystavená torzi naměříme povrchový potenciál 10-150 mV (podle míry deformace). Takto vyvolaný povrchový náboj trvá po celou dobu podnětu a může se synchronně měnit s kolísáním daného podnětu.
 sítě těchto proteinů, zejména kolagenu. Chrupavky se skládají z kolagenu a proteoglykanů, kost je v podstatě kalcifikovaná kolagenní struktura. Všechny tyto útvary mají společné to, že pokud jsou vystaveny mechanickému stresu (komprese, natažení, zkroucení aj.), vznikají na jejich vnitřních i vnějších površích piezoelektrické náboje a potenciály. Např. na kosti vystavená torzi naměříme povrchový potenciál mV (podle míry deformace). Takto vyvolaný povrchový náboj trvá po celou dobu podnětu a může se synchronně měnit s kolísáním daného podnětu..")
4
Polarizované mohou být i tkáně vůči sobě navzájem. Jak zjistit, že se v ptačím vejci vyvíjí embryo? Embryo má vůči žloutku a bílku pozitivní náboj, a protože je lehčí než jeho okolní prostředí, plave při povrchu skořápky. Lze naměřit indukovaný náboj i přes skořápku, která má vysoký odpor: následným otočením skořápky o 180° lze získat superponovanou hodnotu tohoto potenciálu, který se průběhu vývoje zárodku mění. + + + + + + - - - Př. torze kosti: Při působení síly na kost lze měřit v různých místech na jejím povrchu potenciálové rozdíly, navozené změnou v distribuci jejího povrchového náboje. Torzí vyvolaný povrchový náboj trvá po celou dobu podnětu a může se synchronně měnit s kolísáním daného podnětu. Každá jednotlivá ohýbaná molekula přispívá k celkovému potenciálovému rozdílu v různých místech (příspěvky povrchových nábojů molekul se sčítají). Stlačené konkávní povrchy generují záporné potenciálové rozdíly, stlačené konvexní povrchy potenciálové rozdíly kladné. Změny povrchového náboje se měří nejčastěji v pC/cm 2. Ve šlachách vyvolá tah navozený svalovou kontrakcí a zkrácením svalu mezi jeho šlachou a úponem do fascie nebo na kost paralelní (iso)plochy povrchového náboje, orientované v podélné ose šlachy. Podobně lze měřit změny povrchového náboje ve fasciích aj. potenciál (mV) čas (s) Relaxace tkáně po mechanickém stresu. Stres se projevil změnou elektrického potenciálu (měřeno z vnitřku) a změnou elektrochemockého potenciálu celé tkáně. Distribuce elektrického potenciálu uvnitř tkáně v různých časech od aplikovaného tlaku 0,3 MPa. Tkáň byla stlačována v komůrce směrem seshora.
. Stlačené konkávní povrchy generují záporné potenciálové rozdíly, stlačené konvexní povrchy potenciálové rozdíly kladné. Změny povrchového náboje se měří nejčastěji v pC/cm 2. Ve šlachách vyvolá tah navozený svalovou kontrakcí a zkrácením svalu mezi jeho šlachou a úponem do fascie nebo na kost paralelní (iso)plochy povrchového náboje, orientované v podélné ose šlachy. Podobně lze měřit změny povrchového náboje ve fasciích aj. potenciál (mV) čas (s) Relaxace tkáně po mechanickém stresu. Stres se projevil změnou elektrického potenciálu (měřeno z vnitřku) a změnou elektrochemockého potenciálu celé tkáně. Distribuce elektrického potenciálu uvnitř tkáně v různých časech od aplikovaného tlaku 0,3 MPa. Tkáň byla stlačována v komůrce směrem seshora..")
5
Tkáně jako vodiče mají oblasti s různou vodivostí. Distribuci elektrického náboje není snadné vypočítat. Podélný řez lidským stehnem, na němž jsou umístěny dvě povrchové elektrody (A,K). Q,H a G, svalová tkáň; F, tuk; H, kyčel; B, stehenní kost; K, koleno. Ekvipotenciály jsou od sebe vzdáleny 4 V. Šíření akčního potenciálu v m. rectus femoris stimulovaného bifázickými pulsy (30, 80V, 25 Hz).
. Q,H a G, svalová tkáň; F, tuk; H, kyčel; B, stehenní kost; K, koleno. Ekvipotenciály jsou od sebe vzdáleny 4 V. Šíření akčního potenciálu v m. rectus femoris stimulovaného bifázickými pulsy (30, 80V, 25 Hz)..")
6
II V tok proudu nízké i vysoké frekvence většinou vysoké frekvence Lze měřit nejen povrchový náboj, potenciály, proudy a jejich změny jako primární výstupní data, častá jsou v případě tkání tzv. impedanční měření, tedy měření dielektrických vlastností dané tkáně. Provádějí se zpravidla tak, že do tkáně je zaváděn slabý proud o různých frekvencích. Nejlepší výsledky se samozřejmě dosahují přímým kontaktem s tkání, ale i tehdy je třeba brát v potaz vliv okolních tkání, které mohou značně přispívat „parazitickou“ informací. U impedančních měření je tedy velmi přesně polohovat elektrody a brát v potaz velikost a tvar těla nebo jeho relevantní části. Proud si vždy „vybírá“ cestu nejmenšího odporu. Máme-li dobré vodivé médium, vedou i tkáně dobře tkáně s vysokým obsahem vody jsou nejlepší vodiče. Tkáň odpovídá na průchod elektrického proudu různými typy odpovědí. Mezi nejdůležitější patří odpovědi teplotní, chemické a systémové. Teplotní odpovědi zahrnují vzrůst teploty v proud vedoucích tkáních a následnou disipaci tepla z tkáně vyšší elektrický odpor tkáně způsobený vzrůstem teploty proudy stimulující nervy a svaly mají malý teplotní efekt, neboť průměrný tok proudu je velmi malý ohřev tkání je vyšší při vyšších frekvencích proudů Systémové odpovědi zahrnují kontrakce svalů změny ve vnímání bolestivých podnětů Chemické odpovědi zahrnují na plus pólu: acidické reakce způsobující koagulaci proteinů a tvrdnutí tkáně na minus pólu: alkalické reakce způsobují zkapalňování proteinů a měknutí tkání

7
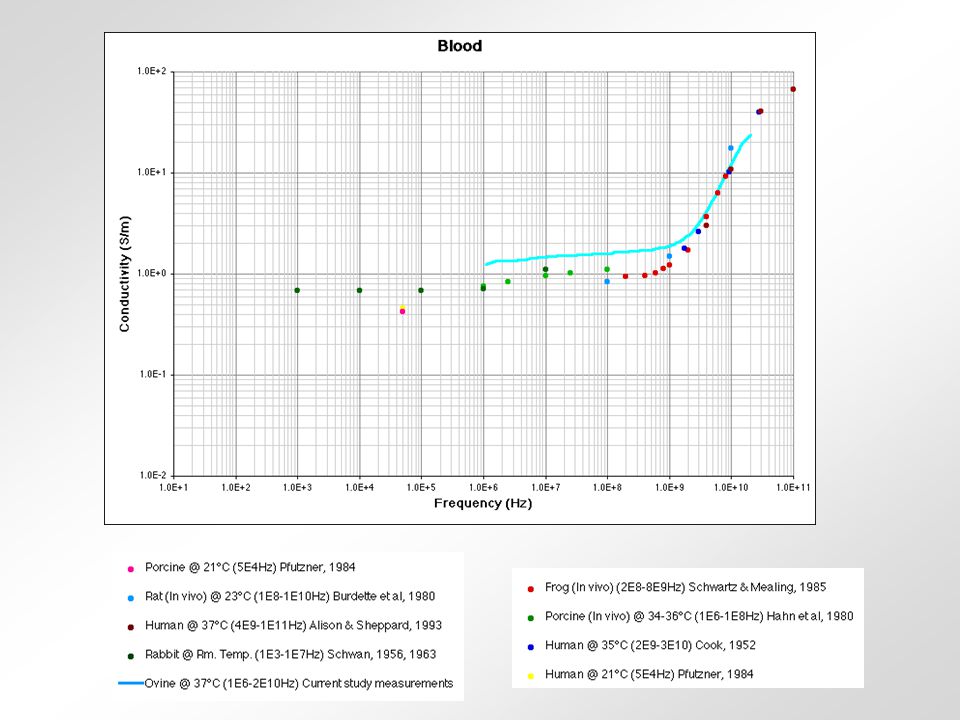
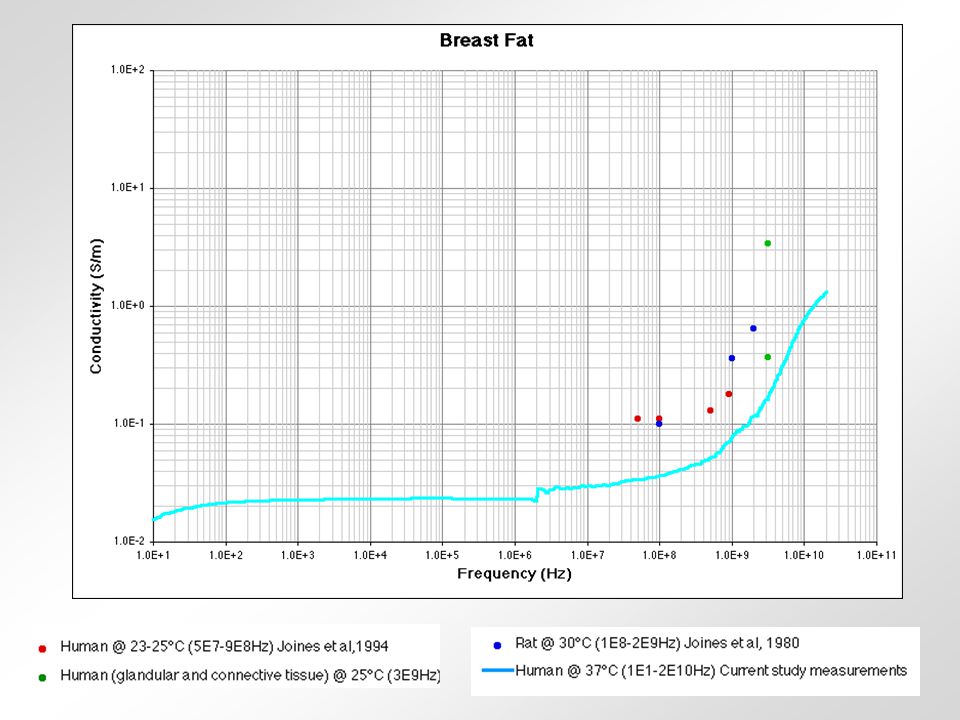
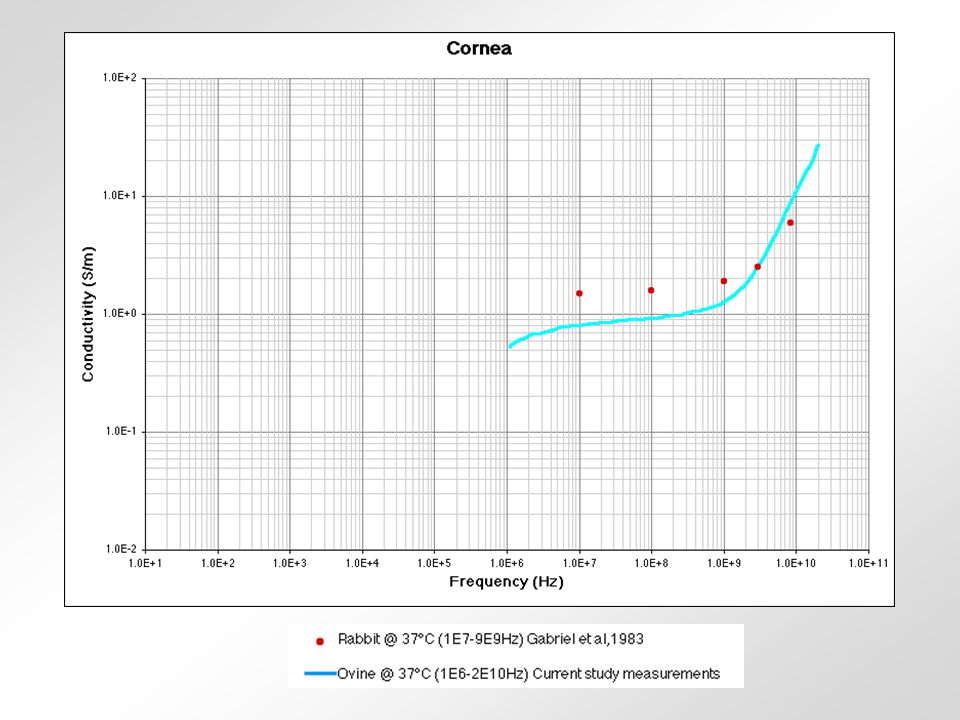
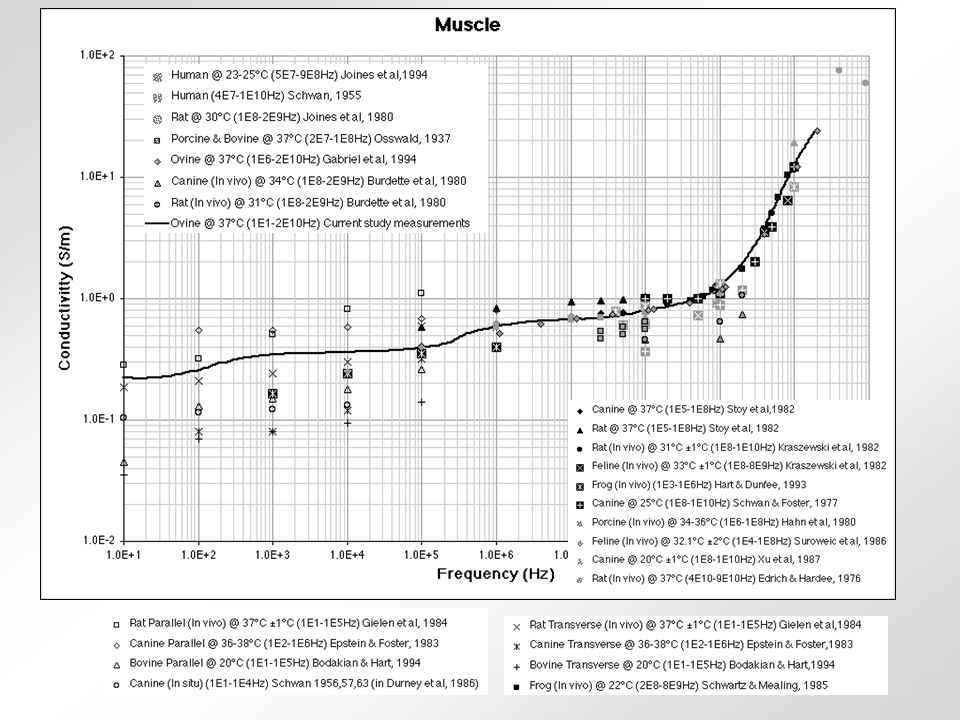
relativní permitivita sval vodivost tuk frekvence (Hz) Krev - obsahuje značné množství vody a iontů - jeden z nejlepších vodičů z biologických tkání Sval (kosterní) - obsahuje 75% vody - jeho vodivost závisí na tocích iontů při kontrakci - nejlepší vodivost má v longitudinálním směru Tuk - obsahuje 14% vody - velmi špatný vodič Kůže - různý obsah vody - u savců považována za izolant - příprava kůže před aplikací stimulu Svalová šlacha - obsahuje málo vody - velmi špatný vodič Periferní nervy - vodivost 6x vyšší než u svalu - myelinové pochvy jsou velmi špatný vodič Kost - obsahuje 5% vody - nejhorší vodič z biologických tkání Srovnání některých tkání podle jejich vodivosti 0,2430,239-0,3320,3000,288100 kHz 0,2220,238-0,2560,2850,27610 kHz 0,1960,1950,2300,2540,21650 Hz KrkNohaRukaTrupHlavaCelé těloFrekvence Vodivost v S/m celého těla a některých jeho částí.
 Krev - obsahuje značné množství vody a iontů - jeden z nejlepších vodičů z biologických tkání Sval (kosterní) - obsahuje 75% vody - jeho vodivost závisí na tocích iontů při kontrakci - nejlepší vodivost má v longitudinálním směru Tuk - obsahuje 14% vody - velmi špatný vodič Kůže - různý obsah vody - u savců považována za izolant - příprava kůže před aplikací stimulu Svalová šlacha - obsahuje málo vody - velmi špatný vodič Periferní nervy - vodivost 6x vyšší než u svalu - myelinové pochvy jsou velmi špatný vodič Kost - obsahuje 5% vody - nejhorší vodič z biologických tkání Srovnání některých tkání podle jejich vodivosti 0,2430,239-0,3320,3000, kHz 0,2220,238-0,2560,2850,27610 kHz 0,1960,1950,2300,2540,21650 Hz KrkNohaRukaTrupHlavaCelé těloFrekvence Vodivost v S/m celého těla a některých jeho částí.")
13
rohovka prsní tuk krev kost Srovnání některých tkání podle jejich vodivosti

14
Rostlinné buňky vnímají svou polohu v tkáni a jsou schopny na základě vnějších podnětů (externích gradientů, zejména teploty, světelné intenzity, pH a gradientu elektrického potenciálu) tuto polohu vyhodnotit a případně změnit – např. přesměrovat růst jiným směrem. Model: buňky hnědých řas (Fucus sp., Pelvetia sp.) uvolňujících gamety bez buněčné stěny do moře. Polarizace zygoty hnědé řasy (Fucus sp.) Nepolarizovaná zygota světlo zastíněná půlka plasmatická membrána jádro polarizace ztráta polarizace zafixování polarizace vtok Ca 2+ via Ca 2+ IKs výtok Ca 2+ tok iontů pučící rhizoid aktinová filamenta budování buněčné stěny dělení buňky sekrece látek pro stavbu buněčné stěny Labilní osa polarizacePolarizovaná zygotaEmbryo - gamety zprvu apolární, osa polarizace se formuje během několika hodin od oplodnění vlivem unilaterálního světelného podnětu - změnou směru osvětlení se může měnit i osa polarizace, fixuje se 10-14 hodin po oplodněn - vzniká polarizovaná jednobuněčná zygota s rhizoidním a stélkovým pólem - zygota se dále dělí a vznikají polarizované buňky: rhizoidní a stélková Po osvitu zygoty se začnou na povrchu buňky redistribuovat Ca 2+ IKs a buňkou začnou protékat transcelulární vápníkové proudy v řádu asi 100 pA. Tyto proudy mění i membránový potenciál a ovlivňují např. distribuci membránových proteinů či polarizaci aktinového cytoskeletu. V rostoucích rhizoidech Fucusů vzniká Ca 2+ gradient v rozmezí 100 nM Ca 2+ na bázi až 450 nM na vrcholku rhizodu.
 uvolňujících gamety bez buněčné stěny do moře. Polarizace zygoty hnědé řasy (Fucus sp.) Nepolarizovaná zygota světlo zastíněná půlka plasmatická membrána jádro polarizace ztráta polarizace zafixování polarizace vtok Ca 2+ via Ca 2+ IKs výtok Ca 2+ tok iontů pučící rhizoid aktinová filamenta budování buněčné stěny dělení buňky sekrece látek pro stavbu buněčné stěny Labilní osa polarizacePolarizovaná zygotaEmbryo - gamety zprvu apolární, osa polarizace se formuje během několika hodin od oplodnění vlivem unilaterálního světelného podnětu - změnou směru osvětlení se může měnit i osa polarizace, fixuje se hodin po oplodněn - vzniká polarizovaná jednobuněčná zygota s rhizoidním a stélkovým pólem - zygota se dále dělí a vznikají polarizované buňky: rhizoidní a stélková Po osvitu zygoty se začnou na povrchu buňky redistribuovat Ca 2+ IKs a buňkou začnou protékat transcelulární vápníkové proudy v řádu asi 100 pA. Tyto proudy mění i membránový potenciál a ovlivňují např. distribuci membránových proteinů či polarizaci aktinového cytoskeletu. V rostoucích rhizoidech Fucusů vzniká Ca 2+ gradient v rozmezí 100 nM Ca 2+ na bázi až 450 nM na vrcholku rhizodu..")
15
Tento vápníkový gradient může být velmi důležitý pro skládání aktinových vláken nebo pro uvolňování transportních váčků vypučených z Golgiho aparátu. Transportní váčky vypučené z Golgiho aparátu přenášejí součásti budoucí/tvořené buněčné stěny. Proto je třeba je zanést tam, kde tkáň roste. Směr transportu je určen gradientem vápenatých iontů. Vlevo: pučící pylová zrnka různých rostlin vykazují gradient Ca 2+, s nejvyšší koncentrací (asi 1 M) na růstovém vrcholku. 20 m nižšívyšší Ca 2+ koncentraceLilium longiflorum Nicotiana sylvestris Tradescantia virginiana Pučící pylová zrnka různých rostlin vykazují gradient Ca 2+, s nejvyšší koncentrací (asi 1 M) na růstovém vrcholku (Fura 2). Pokud zablokujeme tvorbu nových proteinů cykloheximidem, oddálíme vývoj rhizoidu, ale ne vznik a fixaci osy polarity embrya. Vznik polarity embrya není závislý na nových proteinech, ale na přeskupení proteinů už existujících. (a) (b) Transportní váčky oddělěné od Golgiho aparátu se hromadí na rhizoidním pólu zygoty (a) a v rhizoidní buňce embrya (b), ve směru růstu buňky (toluidinová modř). Aktinová filamenta jsou velmi dynamická a jejich tvorba z aktinových monomerů je vysoce regulovaná. Účastní se mj. transportu sekrečních váčků. Pokud zablokujeme tvorbu filament cytochalazinem B, nevznikne polarizace embrya a ustanou i transcelulární proudy.
 na růstovém vrcholku. 20 m nižšívyšší Ca 2+ koncentraceLilium longiflorum Nicotiana sylvestris Tradescantia virginiana Pučící pylová zrnka různých rostlin vykazují gradient Ca 2+, s nejvyšší koncentrací (asi 1 M) na růstovém vrcholku (Fura 2). Pokud zablokujeme tvorbu nových proteinů cykloheximidem, oddálíme vývoj rhizoidu, ale ne vznik a fixaci osy polarity embrya. Vznik polarity embrya není závislý na nových proteinech, ale na přeskupení proteinů už existujících. (a) (b) Transportní váčky oddělěné od Golgiho aparátu se hromadí na rhizoidním pólu zygoty (a) a v rhizoidní buňce embrya (b), ve směru růstu buňky (toluidinová modř). Aktinová filamenta jsou velmi dynamická a jejich tvorba z aktinových monomerů je vysoce regulovaná. Účastní se mj. transportu sekrečních váčků. Pokud zablokujeme tvorbu filament cytochalazinem B, nevznikne polarizace embrya a ustanou i transcelulární proudy..")
16
Detekce nádorů v prsu pomocí mikrovlnného impedančního měření Detekce nádorů v prsu pomocí mikrovlnných imepdančních měření se používá jako doplňková metoda k ultrazvuku, rentgenu (mammografii) či magnetické resonanci (MRI, magnetic resonance imaging). Mikrovlnné frekvence – 300 MHz až 30 GHz. Potenciálně dobře detekuje malé pevné nádory, je lacinější než např. MRI, detekuje jen nádory, ne benigní cysty a je citlivá. Při mikrovlnných frekvencích lze interakci biologické tkáně s vybuzeným elektrickým (elektromagnetickým) polem popsat její celkovou permitivitou . Ta se skládá z dielektrické konstanty (“reálná“ složka) ´ a z tzv. faktoru ztrát ´´. Dielektrická konstanta určuje schopnost tkáně udržet energii elektrického pole, faktor ztrát nám říká, kolik z této energie se při expozici tkáně poli přemění v teplo a disipuje. prsní tkáň vysílač přijímače prsní tkáň nádor vysílač přijímače Zdravá tkáň propouští mikrovlny úměrně své dielektrické konstantě. Ty jsou pak zaznamenány přijímači. Nádorová tkáň má jiné elektrické charakteristiky. Zatímco prsní tkáň propouští mikrovlny úměrně své dielektrické konstantě, od buněk nádoru se vlny částečně odrážejí a jsou zachyceny anténou vysílače.
 polem popsat její celkovou permitivitou . Ta se skládá z dielektrické konstanty ( reálná složka) ´ a z tzv. faktoru ztrát ´´. Dielektrická konstanta určuje schopnost tkáně udržet energii elektrického pole, faktor ztrát nám říká, kolik z této energie se při expozici tkáně poli přemění v teplo a disipuje. prsní tkáň vysílač přijímače prsní tkáň nádor vysílač přijímače Zdravá tkáň propouští mikrovlny úměrně své dielektrické konstantě. Ty jsou pak zaznamenány přijímači. Nádorová tkáň má jiné elektrické charakteristiky. Zatímco prsní tkáň propouští mikrovlny úměrně své dielektrické konstantě, od buněk nádoru se vlny částečně odrážejí a jsou zachyceny anténou vysílače..")
17
Modře, zeleně, bíle, fialově a žlutě jsou různé žlázy. 6 mm nádor je červená kulička. Zbytek prostoru je vyplněn prsním tukem. Vodivost tumoru byla 4 S/m, jeho dielektrická konstanta =50. Vodivost prsní tkáně byla 0,4 S/m, dielektrická konstanta =9. Tloušťka kůže byla 2mm, její vodivost 4 S/m a dielektrická konstanta =36. Hemisférický model prsu o průměru 14 cm. bradavka kůže s nádorem bez nádoru čas (s) Normalizované napětí Rozdílné odpovědi tkáně s nádorem a bez nádoru. Praxe: žena leží a má prsa ponořena v roztoku glycerinu ve vodě (tato směs má velmi malý faktor ztrát). Do nádrže s roztokem je umístěno 16 vysílačů s anténami, signály jsou měřeny 15 přijímači na 12 různých frekvencích celého užívaného pásma. Doba záznamu je asi 14 minut.
 Normalizované napětí Rozdílné odpovědi tkáně s nádorem a bez nádoru. Praxe: žena leží a má prsa ponořena v roztoku glycerinu ve vodě (tato směs má velmi malý faktor ztrát). Do nádrže s roztokem je umístěno 16 vysílačů s anténami, signály jsou měřeny 15 přijímači na 12 různých frekvencích celého užívaného pásma. Doba záznamu je asi 14 minut..")
18
Elektrické děje během dozrávání gamet a oplodnění Gamety jsou buňky poměrně hojně vybavené iontovými kanály, schopné odpovídat na elektrické stimuly. Jako jeden z prvních studoval roli iontů ve fyziologii oocytu Edward Chambers (1946, role výměny Na + a K + během oplození ježovky). Na hvězdicích, ježovkách a žahavcích se také studovaly elektrofyziologické vlastnosti oocytární membrány. Nezralé oocyty hvězdice exprimují 3 typy napěťově ovládaných IKs: dovnitř usměrňující Ca 2+ IKs, rychlé přechodné K + IKs a dovnitř usměrňující K + IKs. Během dozrávání se mění amplitudy všech těchto proudů: vápníkový proud roste, oba proudy draslíkové se snižují. Klesá také vodivost membrány oocytu a dochází k lehké depolarizaci klidového membránového potenciálu (díky změnám ve vodivosti pro K + a Na + ). Dozrávávní oocytární membrány Výbava iontovými kanály v membráně oocytu se liší druhově: Boltenia vilosa je „osázena“ dovnitř usměrňujícími přechodnými Na + IKs, dovnitř usměrňujícími přechodnými Ca 2+ IKs a dovnitř usměrňující K + IKs. U žab dominují u nezralých oocytů K + a Cl - proudy, během dozrávání jsou nahrazeny Na + proudy. U savců není měření oocytárních proudů nezralého oocytu snadné, nebot oocyt je obalen buňkami tvořícími tzv. cumullus oophorus. Tyto dvě „funkční jednotky“ se spontánně oddělí po dozrání oocytu. Umělé odstraněné cumulus oophorus může vést k různým artefaktům. Buňky cumullu a oocytu mají různý membránový potenciál, ačkoliv je cumullus s oocytem spojen vodivě pomocí gap junctions. Takto je asi oocyt udržován v rané fázi meiosy – jeho membrána je lehce hyperpolarizovaná (oproti cumullu). Působení gonadotropinů na buňky cumullu navodí depolarizaci membrány oocytu a jeho dozrání.
. Na hvězdicích, ježovkách a žahavcích se také studovaly elektrofyziologické vlastnosti oocytární membrány. Nezralé oocyty hvězdice exprimují 3 typy napěťově ovládaných IKs: dovnitř usměrňující Ca 2+ IKs, rychlé přechodné K + IKs a dovnitř usměrňující K + IKs. Během dozrávání se mění amplitudy všech těchto proudů: vápníkový proud roste, oba proudy draslíkové se snižují. Klesá také vodivost membrány oocytu a dochází k lehké depolarizaci klidového membránového potenciálu (díky změnám ve vodivosti pro K + a Na + ). Dozrávávní oocytární membrány Výbava iontovými kanály v membráně oocytu se liší druhově: Boltenia vilosa je „osázena dovnitř usměrňujícími přechodnými Na + IKs, dovnitř usměrňujícími přechodnými Ca 2+ IKs a dovnitř usměrňující K + IKs. U žab dominují u nezralých oocytů K + a Cl - proudy, během dozrávání jsou nahrazeny Na + proudy. U savců není měření oocytárních proudů nezralého oocytu snadné, nebot oocyt je obalen buňkami tvořícími tzv. cumullus oophorus. Tyto dvě „funkční jednotky se spontánně oddělí po dozrání oocytu. Umělé odstraněné cumulus oophorus může vést k různým artefaktům. Buňky cumullu a oocytu mají různý membránový potenciál, ačkoliv je cumullus s oocytem spojen vodivě pomocí gap junctions. Takto je asi oocyt udržován v rané fázi meiosy – jeho membrána je lehce hyperpolarizovaná (oproti cumullu). Působení gonadotropinů na buňky cumullu navodí depolarizaci membrány oocytu a jeho dozrání..")
19
L-typ Ca 2+ IKs K + IKs Ca 2+ - aktivované K + IKs Cl - IKs out in dozráváníoplození chemotaxe akrosomální reakce Ca 2+ Během rané fáze meiosy dominuje vliv K + a Ca 2+ IKs L-typu, který klesá během metafáze II. Po oplození spermie navodí uvolnění Ca 2+ z intracelulárních rezervoárů, což aktivuje K + IKs. Otevírají se i vápníkové kanály, vápník vtéká do buňky, membrána se depolarizuje. Plasmatická membrána bovinního oocytu během dozrávání a oplození. I membrána spermií doznává během chemotaktického pohybu a akrosomální reakce změn. Akrosomální reakce (AR) je děj regulovaný iontovými kanály: vtok Ca 2+ je zásadním požadavkem pro AR u všech živočišných druhů. Zvyšuje se také pH (výtok H + a výměna H + /Na + ). Na hlavičce spermie jsou také 4 typy Cl - IKs s různou vodivostí; eflux chloridů je zřejmě navozen progesteronem.
 je děj regulovaný iontovými kanály: vtok Ca 2+ je zásadním požadavkem pro AR u všech živočišných druhů. Zvyšuje se také pH (výtok H + a výměna H + /Na + ). Na hlavičce spermie jsou také 4 typy Cl - IKs s různou vodivostí; eflux chloridů je zřejmě navozen progesteronem..")
20
Prvním (a velmi univerzálním) krokem je spermií navozené uvolnění vápníkových zásob v oocytu (možná zprostředkované aktivací fosfolipasy C při kontaktu oocyt-spermie). Následných procesů vápníkové signalizace v oocytu se účastní řada dalších receptorů (RyR aj.). Spermie navodí na membráně oocytu krátkodobou potenciálovou změnu nazývanou fertilizační potenciál. Proud zodpovědný za tento potenciál je u savců veden iontově-nespecifickými kanály o vodivosti cca 70pS, dále Na + a Ca + IKs. Depolarizační krok (šipka) v momentu oplození ježovčích oocytů, následovaný fertilizačním potenciálem. Dale et al., 1978) Typy IKs zapojených do aktivace spermie a oocytu a změn membránového potenciálu. Všimněte se, že u obratlovců jde výhradně o hyperpolarizační projevy. RP = klidový membránový potenciál. Oplození je vysoce specifický proces zahrnující rozpoznání gamet navzájem, jejich navázání a fúzi. Klíčovým momentem je reciproká aktivace těchto gamet.
. Spermie navodí na membráně oocytu krátkodobou potenciálovou změnu nazývanou fertilizační potenciál. Proud zodpovědný za tento potenciál je u savců veden iontově-nespecifickými kanály o vodivosti cca 70pS, dále Na + a Ca + IKs. Depolarizační krok (šipka) v momentu oplození ježovčích oocytů, následovaný fertilizačním potenciálem. Dale et al., 1978) Typy IKs zapojených do aktivace spermie a oocytu a změn membránového potenciálu. Všimněte se, že u obratlovců jde výhradně o hyperpolarizační projevy. RP = klidový membránový potenciál. Oplození je vysoce specifický proces zahrnující rozpoznání gamet navzájem, jejich navázání a fúzi. Klíčovým momentem je reciproká aktivace těchto gamet..")
21
Elektrické orgány ryb - elektroplaky Vznikly z modifikovaných svalových vláken. Jsou inervovány cholinergně. Jejich nervosvalová spojení jsou mnohem větší a početnější než u vláken kosterní svaloviny. Postsynaptické membrány jsou hustě osázeny acetylcholinovými Rs nikotinického typu (z jednoho elektroplaku parejnoka rodu Torpedo lze „vytěžit“ až miligramy purifikovaného receptoru!). E-list postsynaptikcé membrány Torpeda připravený lámáním za mrazu. nAChRs jsou v řádcích. Úsečka = 0,1 m. motorický neuron elektroplak -90 mV 0 mV Jedna stran ploché buňky plaku (= elektrocytu) je inervovaná, má vysoký elektrický odpor a značné množství nAChRs, Na + a K + IKs. Druhá strana takto vybavená není a odpor má nízký. Buněk může být v jednom plaku až 5000. Navzájem jsou propojeny elektrickými synapsemi (synchronizace!) Na obou stranách plaku v klidu je na membráně potenciál asi – 90 mV. Jednotlivé „články“ jsou spojeny do série. Vzpomeňte si na Voltův sloup!
. E-list postsynaptikcé membrány Torpeda připravený lámáním za mrazu. nAChRs jsou v řádcích. Úsečka = 0,1 m. motorický neuron elektroplak -90 mV 0 mV Jedna stran ploché buňky plaku (= elektrocytu) je inervovaná, má vysoký elektrický odpor a značné množství nAChRs, Na + a K + IKs. Druhá strana takto vybavená není a odpor má nízký. Buněk může být v jednom plaku až Navzájem jsou propojeny elektrickými synapsemi (synchronizace!) Na obou stranách plaku v klidu je na membráně potenciál asi – 90 mV. Jednotlivé „články jsou spojeny do série. Vzpomeňte si na Voltův sloup!.")
22
motorický neuron elektroplak -90 mV 0 mV motorický neuron elektroplak +40 mV -90 mV 130 mV Na nervový podnět dojde k výlevu ACh a masivní depolarizaci vzrušivé strany plaku. Vznikne akční potenciál, dojde ke zvratu polarity a na inervované straně plaku je potenciálový rozdíl mezi vnějškem a vnitřkem membrány +40 mV. Druhá strana plaku je nevzrušivá a zachovává si potenciál –90 mV. Celý plak tedy vytvoří potenciálový rozdíl 130 mV. Pokud je 5 000 elektroplaků spojeno do série a na každém je potenciálový rozdíl 130 mV, výsledný rozdíl napětí na pólech sloupce bude 5 000 x 0,130 = 650 V Sladkovodní úhoř Electrophorus electricus může vyvinout až 800 V (i 650 V stačí na usmrcení člověka).
..")
23
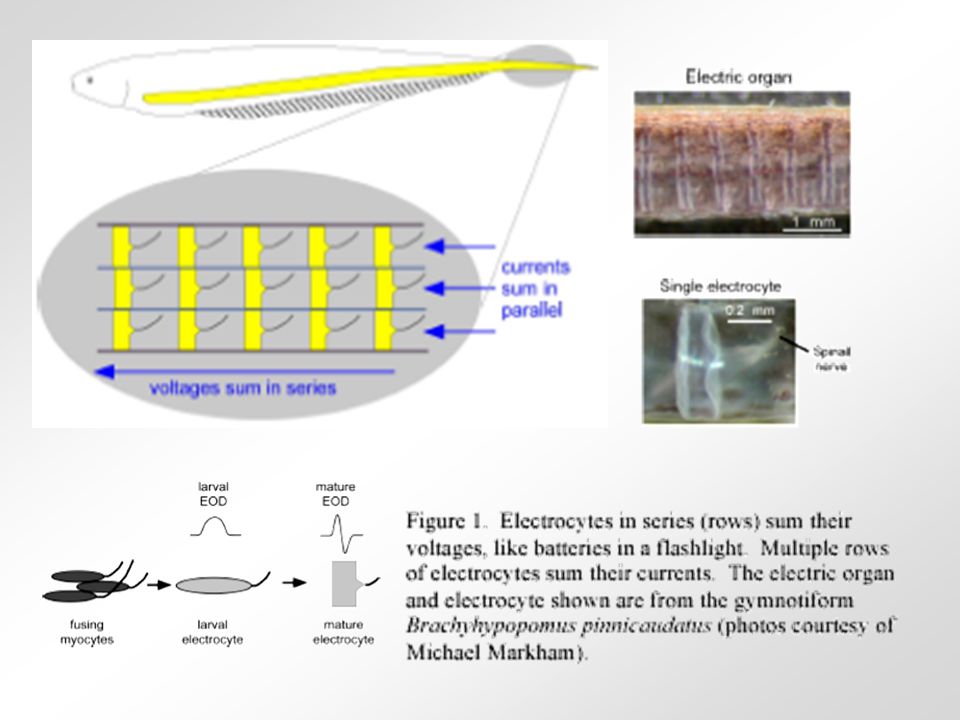
Elektrické orgány se vyvinuly nezávisle přinejmenším v 6 různých skupinách ryb (rejnoci, parejnoci, Gymnotiformes, Mormyriformes aj.). U některých generují silné elektrické výboje (Torpedo sp., Gymnotus sp., Raja sp.), u jiných slabé. Pulsy slouží k např. elektrolokaci, rozpoznávání hrubého tvaru, vodivosti a lokace blízkých objektů, k rozpoznání jedinců téhož druhu, lákání partnera, obraně či lovu kořisti. Morfologie elektrocytů se liší mezidruhově a souvisí s typem výbojů, který ryba generuje. Slabě elektrické ryby generují výboje o síle stovek milivoltů až jednotek voltů, silně elektrické ryby generují až stovky voltů. Slabě elektrické ryby generují výboje buď v nepravidelně se objevujících pulsech, nebo v pravidelných sinusových vlnách o frekvenci 50-1000 Hz. Elektrocyty zástupce rodu Gymnotus mají asi 1 mm v průměru a jsou cca 300 m tlusté. Produkují jen zlomky voltů, což stačí k elektrolokaci. Inervovány jsou podobně jako u Torpeda.
, u jiných slabé. Pulsy slouží k např. elektrolokaci, rozpoznávání hrubého tvaru, vodivosti a lokace blízkých objektů, k rozpoznání jedinců téhož druhu, lákání partnera, obraně či lovu kořisti. Morfologie elektrocytů se liší mezidruhově a souvisí s typem výbojů, který ryba generuje. Slabě elektrické ryby generují výboje o síle stovek milivoltů až jednotek voltů, silně elektrické ryby generují až stovky voltů. Slabě elektrické ryby generují výboje buď v nepravidelně se objevujících pulsech, nebo v pravidelných sinusových vlnách o frekvenci Hz. Elektrocyty zástupce rodu Gymnotus mají asi 1 mm v průměru a jsou cca 300 m tlusté. Produkují jen zlomky voltů, což stačí k elektrolokaci. Inervovány jsou podobně jako u Torpeda..")
24
Elektrocyty (EL) zástupce rodu Eigenmannia sp. D, dermis A, ocasní ploutev Šipkou je označena inervace motorickými nervy

26
Sternopygus macurus, 50-200 Hz Slabě elektrické ryby se zřejmě v evoluci vydělily dvakrát. Jedna skupina žije v Africe (Mormyriformes), druhá v Jižní Americe (Gymnotiformes). Žijí v bahnitých vodách, kde jim elektrolokace nahrazuje do značné míry zrak. S výjimkou jediného zástupce samičky těchto druhů vydávají výboje o vyšší frekvenci než samci. Zdá se, že hormony ovlivňují také amplitudu výboje. Eigenmannia virescens, 250-600 Hz Apteronotus leptorhynchus, 650-1100 Hz výboj mozek elektrický orgán receptorvýboj
, druhá v Jižní Americe (Gymnotiformes). Žijí v bahnitých vodách, kde jim elektrolokace nahrazuje do značné míry zrak. S výjimkou jediného zástupce samičky těchto druhů vydávají výboje o vyšší frekvenci než samci. Zdá se, že hormony ovlivňují také amplitudu výboje. Eigenmannia virescens, Hz Apteronotus leptorhynchus, Hz výboj mozek elektrický orgán receptorvýboj.")
27
Sexuální dimorfismus výboje u Hyppomus occidentalis. Samci (A) jsou větší a mají delší tenčí ocas. Jejich výboj je asymetrický, neboť druhý výboj bifázického pulsu je delší než první. Samice (B) jsou kratší a jejich puls je kratší, symetrický.
 jsou větší a mají delší tenčí ocas. Jejich výboj je asymetrický, neboť druhý výboj bifázického pulsu je delší než první. Samice (B) jsou kratší a jejich puls je kratší, symetrický..")
28
Vnímání elektrického pole není doménou ryb; jsou jej schopny paryby (žraloci, rejnoci), některé kostnaté ryby (sumci), mloci, axolotli nebo např. i ptakopysci. Vnímají i velmi slabé elektrické pole: rejnok Raja clavata je schopen detekce 0,01-0,02 V/cm (hodnota produkovaná napětím monočlánku na vzdálenost asi 1 000 km). K vnímání intensity elektrického pole slouží ampulární orgány. Ampulární orgány jsou složeny z kanálků o průměru asi 1 mm a délce 15-20 cm. Jsou distribuovány po těle živočicha a otevírají se přímo do (mořské) vody. kinocilie tight junction mikroklky receptor podpůrná buňka synaptický hřebínek bazální membrána kanál alveoly vnitřek alveolu nervová vlákna kanálky otvory oko kapsula břišní strana
. K vnímání intensity elektrického pole slouží ampulární orgány. Ampulární orgány jsou složeny z kanálků o průměru asi 1 mm a délce cm. Jsou distribuovány po těle živočicha a otevírají se přímo do (mořské) vody. kinocilie tight junction mikroklky receptor podpůrná buňka synaptický hřebínek bazální membrána kanál alveoly vnitřek alveolu nervová vlákna kanálky otvory oko kapsula břišní strana.")
29
Žraloci a spol. žijící ve mořské vodě s vysokou koncentrací iontů mají nízký odpor kůže. Elektrické pole ve vodě tedy není příliš rušeno tělem živočicha. Stěny kanálků ampulárních orgánů mají odpor vyšší, uniká přes ně jen málo náboje. Pokud jsou kanál a vnější elektrické pole orientovány stejným směrem, lze intenzitu elektrického pole (V/cm) vypočítat jako součin délky kanálku a síly pole. Při konstantní intenzitě pole bude napěťový rozdíl dráždící receptor tím větší, čím delší bude kanál. Díky různé délce kanálků mohou např. rejnoci vnímat různé amplitudy v různých směrech a místech. Tyto receptory se pravděpodobně vyvinuly z vláskových buněk postranní čáry. Jejich excitačním neuropřenašečem je zřejmě glutamát. Struktura receptorů vnímajících změny v intenzitě elektrického pole je u sladkovodních ryb jednoduchá. Jde o prostý kanál otevřený do vody, na jehož „dně“ jsou receptorové buňky. Pokud je napětí vně ryby kladnější než uvnitř, tekou proudy dovnitř receptorových buněk přes luminální membránu a pak ven na membránu bazální. lumen kanálu receptorové buňky luminální membrána bazální membrána Ta se depolarizuje, zvyšuje se pravděpodobnost otevření Ca 2+ IKs, do přilehlé nervové buňky vstupuje vápník, depolarizuje ji a navodí výlev neuropřenašeče (receptor je sekundární). Ten depolarizuje postsynaptické nervové vlákno a zvyšuje frekvenci vzniku APs.
 vypočítat jako součin délky kanálku a síly pole. Při konstantní intenzitě pole bude napěťový rozdíl dráždící receptor tím větší, čím delší bude kanál. Díky různé délce kanálků mohou např. rejnoci vnímat různé amplitudy v různých směrech a místech. Tyto receptory se pravděpodobně vyvinuly z vláskových buněk postranní čáry. Jejich excitačním neuropřenašečem je zřejmě glutamát. Struktura receptorů vnímajících změny v intenzitě elektrického pole je u sladkovodních ryb jednoduchá. Jde o prostý kanál otevřený do vody, na jehož „dně jsou receptorové buňky. Pokud je napětí vně ryby kladnější než uvnitř, tekou proudy dovnitř receptorových buněk přes luminální membránu a pak ven na membránu bazální. lumen kanálu receptorové buňky luminální membrána bazální membrána Ta se depolarizuje, zvyšuje se pravděpodobnost otevření Ca 2+ IKs, do přilehlé nervové buňky vstupuje vápník, depolarizuje ji a navodí výlev neuropřenašeče (receptor je sekundární). Ten depolarizuje postsynaptické nervové vlákno a zvyšuje frekvenci vzniku APs..")
30
Ampulární receptory jsou tonické – pomalu se adaptují a reagují na relativně stálé změny v intenzitě okolního pole. Jiný typ receptorů slouží k elektrolokaci: tuberózními receptory (?, tuberous receptors. Tyto receptory reagují na pulsy vysílané rybou (generované elektrocyty). mozek elektrický orgán receptorvýboj Tyto receptory nejsou spojeny kanálem s vnějším prostředím, ale jsou uzavřeny a odděleny vrstvou epiteliálních buněk. Mají různý počet receptorových buněk, jejichž membrána často ční do kanálku. U bází jsou spojeny těsnými spoji (tight junction). Nemají cílie, ale často jsou pokryty mikroklky. Jejich bazální membrána také obsahuje synaptické hřebínky a zpravidla k ní přiléhá jen jeden aferentní nerv. Elektroloka(liza)ci popsal prvně H.W. Lissmann. epidermis epitelové buňky lumen kanálku receptorové buňky nerv
. mozek elektrický orgán receptorvýboj Tyto receptory nejsou spojeny kanálem s vnějším prostředím, ale jsou uzavřeny a odděleny vrstvou epiteliálních buněk. Mají různý počet receptorových buněk, jejichž membrána často ční do kanálku. U bází jsou spojeny těsnými spoji (tight junction). Nemají cílie, ale často jsou pokryty mikroklky. Jejich bazální membrána také obsahuje synaptické hřebínky a zpravidla k ní přiléhá jen jeden aferentní nerv. Elektroloka(liza)ci popsal prvně H.W. Lissmann. epidermis epitelové buňky lumen kanálku receptorové buňky nerv.")
31
Ampulární receptory jsou tonické – pomalu se adaptují a reagují na relativně stálé změny v intenzitě okolního pole. Tuberózní receptory slouží k elektrolokaci, receptory reagují na pulsy vysílané rybou (generované elektrocyty). Rozsah sensitivit obou typů receptorů se liší. Obecně jsou citlivější ampulární receptory, rozkódují i nižší frekvence.
. Rozsah sensitivit obou typů receptorů se liší. Obecně jsou citlivější ampulární receptory, rozkódují i nižší frekvence..")
32
Někteří zástupci mohou mít i dva či tři elektrické orgány, jako např. Electrophorus electricus. Hlavní orgán a přední část Hunterova orgánu produkují vysokonapěťové výboje, zatímco Sachsův orgán a zbytek orgánu Hunterova výboje nízkonapěťové. Hlavní orgán Sachsův orgán Hunterův orgán

33
Elektrický orgán generuje výboj (podle orientace sloupce elektroplaku + nebo -). Proud opustí plak na jeho anteriorním konci a na konci posteriorním se vrací. To vytvoří kolem ryby 3D gradient napětí, který zvíře detekuje.

34
Napětí rychle klesá se vzdáleností a slouží k získání informace do vzdálenosti max. 1-2 délky těla ryby. Pokud je v blízkosti ryby izolant (kamínek aj.) nebo vodič (červ k snědku, rostlina aj.), dojde k narušení ekvipotenciálních ploch. Aby ryby nerušily toto pole vlastním pohybem, dochází u nich jen k vlnivým pohybům hřbetní ploutve. izolant vodič
 nebo vodič (červ k snědku, rostlina aj.), dojde k narušení ekvipotenciálních ploch. Aby ryby nerušily toto pole vlastním pohybem, dochází u nich jen k vlnivým pohybům hřbetní ploutve. izolant vodič.")
35
Co si pamatovat z dnešní přednášky - co je to tkáň - apolární a polární buňky - potenciály v poškozené a mechanicky namáhané tkáni - odpověď tkáně na průchod proudu - impedanční a vodivostní měření: srovnání tkání, detekce nádorů v tkáních - vývoj polarity rostlinných buněk (tkání) - elektrické jevy při dozrávání zárodečných buněk - ryby a jejich elektrické orgány (elektroplaky a receptory)
 - elektrické jevy při dozrávání zárodečných buněk - ryby a jejich elektrické orgány (elektroplaky a receptory)")
Podobné prezentace