Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
OBECNá protistologie - MB160P63
Úvod, genomy protist (Hampl) Symbiózy protist (Čepička) Semiautonomní organely (Čepička) Horizontální genový přenos u protist (Zubáčová) Způsoby získávání potravy u protist (Hampl) Metabolické zvláštnosti protist (Hampl) Mnohobuněčnost (Čepička) Buněčný cyklus, pohlavní rozmnožování a druhovost (Čepička) Pohyb bičíky a fyzikální zvláštnosti života v mikrosvětě (Hampl) Další způsoby pohybu (Hampl) Každý musí mít vybráno téma eseje Diverzita protist na různých úrovních (kryptické druhy, objevy nových skupin, flagship species apod.) (Čepička) Odevzdání esejů Prezentace studentských esejů ESEJ – Volné pojednání založené na oborném článku/článcích. Téma si můžete zvolit sami, ale my jej musíme schválit. Téma se nesmí týkat diplomové nebo dizertační práce. Rozsah minimálně 2 strany A4, písmo 12, řádkování 1. PREZENTACE – 15 minut + diskuze. PÍSEMKA – 10 otázek (20 bodů, 18-20=1, 15-17=2, 11-14=3) Celková známka = (esej + prezentace + písemka) /3 Materiály na Moodle klíč:protista jsou nejlepsi
 Symbiózy protist (Čepička) Semiautonomní organely (Čepička) Horizontální genový přenos u protist (Zubáčová) Způsoby získávání potravy u protist (Hampl) Metabolické zvláštnosti protist (Hampl) Mnohobuněčnost (Čepička) Buněčný cyklus, pohlavní rozmnožování a druhovost (Čepička) Pohyb bičíky a fyzikální zvláštnosti života v mikrosvětě (Hampl) Další způsoby pohybu (Hampl) Každý musí mít vybráno téma eseje Diverzita protist na různých úrovních (kryptické druhy, objevy nových skupin, flagship species apod.) (Čepička) Odevzdání esejů Prezentace studentských esejů. ESEJ – Volné pojednání založené na oborném článku/článcích. Téma si můžete zvolit sami, ale my jej musíme schválit. Téma se nesmí týkat diplomové nebo dizertační práce. Rozsah minimálně 2 strany A4, písmo 12, řádkování 1. PREZENTACE – 15 minut + diskuze. PÍSEMKA – 10 otázek (20 bodů, 18-20=1, 15-17=2, 11-14=3) Celková známka = (esej + prezentace + písemka) /3. Materiály na Moodle klíč:protista jsou nejlepsi.")
3
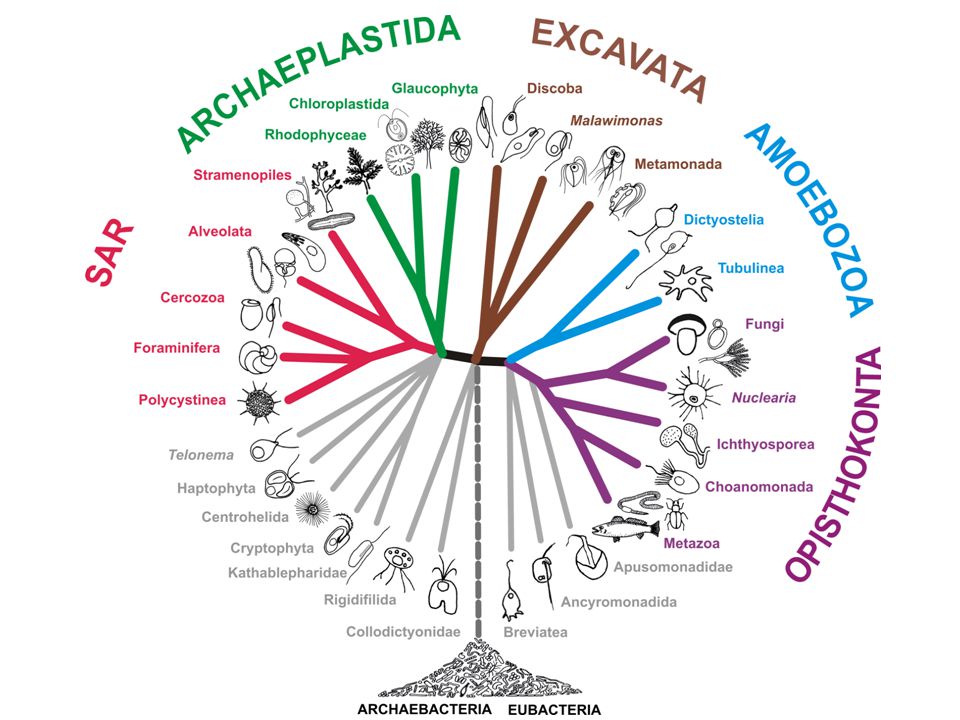
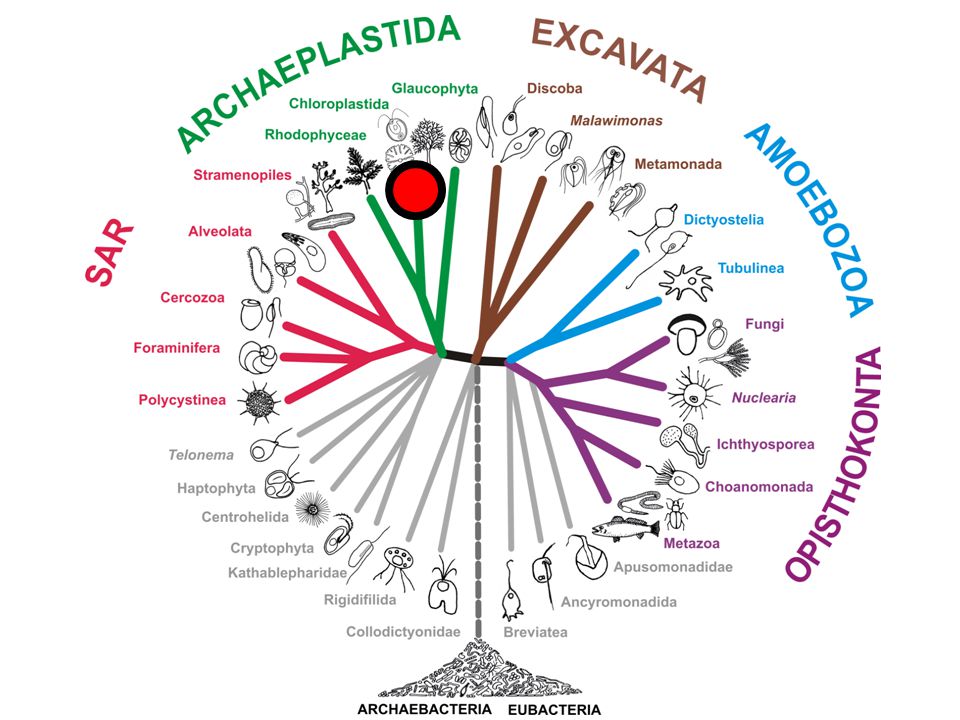
CORTICATA DIAPHORETICKES AMORPHEA (UNIKONTA) Primitivní mitochondrie
Nepřítomnost TOM40, syntéza cytochromu c Primitivní mitochondrie Vzácné genomické změny Analýza genů původem z bakterií Analýza mitochondriálních genů AMORPHEA (UNIKONTA)
")
4
Jaderné genomy prvoků

5
Základní pojmy Hodnota C – množství DNA v jádře (pg) Velikost genomu
– počet párů bazí (Mbp) Euchromatin - Volněji sbalená DNA Heterochromatin Těsněji sbalená neexprimovaná DNA Nekódující DNA - Přítomná v hetero i euchromatinu – mezigenové oblasti, introny, repetitivní sekvence Genová densita genomu - Počet genů na 1 bp (1 kbp)
 Euchromatin. - Volněji sbalená DNA. Heterochromatin. Těsněji sbalená neexprimovaná DNA. Nekódující DNA. - Přítomná v hetero i euchromatinu – mezigenové oblasti, introny, repetitivní sekvence. Genová densita genomu. - Počet genů na 1 bp (1 kbp)")
6
Rozsah velikostí genomů skupin organizmů
Velikost genomů eukaryot je obecně vyšší než u prokaryot Varíruje x Za tento rozsah mohou především prvoci 1pg = asi 1 GB

7
Keeling 2005

8
Redukované genomy

10
Ostreococcus tauri Buňka má µm v průměru a je µm tlustá. Nemá bičíky. 12.6 Mbp (7 735genů) 71 kbp
 71 kbp.")
11
Ostreococcus – důsledky malého a hustého genomu
Zkracování mezigenových oblastí (průměr 197 nt) Snižování velikosti genových rodin (2.5–3.8% genů jsou transkripční faktory, u eukaryot to je obvykle 12-15%) Menší počet kopií genů pro fotosyntézu Fůze genů (přes 300 fúzovaných genů) Selenocysteinové proteiny – mají vyšší aktivitu, stačí jich méně na buňku, tj. buňka může zmenšovat svůj objem V buňce není místo pro klasické dělící vřeténko a není jasné, jak se buňka vlastně dělí
 Snižování velikosti genových rodin (2.5–3.8% genů jsou transkripční faktory, u eukaryot to je obvykle 12-15%) Menší počet kopií genů pro fotosyntézu. Fůze genů (přes 300 fúzovaných genů) Selenocysteinové proteiny – mají vyšší aktivitu, stačí jich méně na buňku, tj. buňka může zmenšovat svůj objem. V buňce není místo pro klasické dělící vřeténko a není jasné, jak se buňka vlastně dělí.")
12
Gan a kol. 2011

13
Chromozom 2: Obsahuje mnoho intronů, transpozonů a má nízký obsah GC
20 chromozomů (12.6 Mbp, 7 735genů) Chromozom 2: Obsahuje mnoho intronů, transpozonů a má nízký obsah GC Chromozom 19: Obsahuje mnoho intronů, transpozonů a genů jiného původu než ze zelených řas General characteristics of the 20 O. tauri chromosomes. TEs, transposon frequency. Size is indicated to the left of each chromosome (Mb). Colored bars indicate the percentage G+C content (upper bar) and of transposons (lower bar). Derelle E. et.al. PNAS 2006;103: ©2006 by National Academy of Sciences
 Chromozom 2: Obsahuje mnoho intronů, transpozonů a má nízký obsah GC. Chromozom 19: Obsahuje mnoho intronů, transpozonů a genů jiného původu než ze zelených řas. General characteristics of the 20 O. tauri chromosomes. TEs, transposon frequency. Size is indicated to the left of each chromosome (Mb). Colored bars indicate the percentage G+C content (upper bar) and of transposons (lower bar). Derelle E. et.al. PNAS 2006;103: ©2006 by National Academy of Sciences.")
14
Syntenie mezi chromozomy O. tauri (Ot) and O. lucimarinus (Ol)
Synteny between the chromosomes of O. tauri (Ot) and O. lucimarinus (Ol). Depicted areas in red show collinear regions (conserved gene order and content) as described in Methods. Blocks of different colors denote different sorts of duplications: blue, an internally duplicated segment; green, a duplicated segment that is collinear with a segment on a different chromosome in both Ot and Ol; yellow, a duplicated segment that is collinear with a segment on a different chromosome in Ol; orange, a duplicated segment that is collinear with a segment on a different chromosome in Ot. Palenik B. et.al. PNAS 2007;104: ©2007 by National Academy of Sciences
 and O. lucimarinus (Ol). Depicted areas in red show collinear regions (conserved gene order and content) as described in Methods. Blocks of different colors denote different sorts of duplications: blue, an internally duplicated segment; green, a duplicated segment that is collinear with a segment on a different chromosome in both Ot and Ol; yellow, a duplicated segment that is collinear with a segment on a different chromosome in Ol; orange, a duplicated segment that is collinear with a segment on a different chromosome in Ot. Palenik B. et.al. PNAS 2007;104: ©2007 by National Academy of Sciences.")
15
Micromonas obsahuje také dva Chromosomy s nižším GC obsahem
(21,9 MB, genů) (20,9 MB, genů) Worden a kol. 2009
 (20,9 MB, genů) Worden a kol")
16
MIKROSPORIDIE

17
Srovnání velikosti s jinými organizmy
Kromě parazitického intracelulárního způsobu života může na zmenšování genomu tlačit skutečnost, že jádro prochází velmi tenkou trubicí (na obrázku). Peyretaillade a kol. 2011
. Peyretaillade a kol")
18
Genomy mikrosporidií Corradi a kol. 2010

19
Genomy mikrosporidií Skoro stejně genů, ale menší hustota a mnoho transpozonů Corradi a kol. 2009

20
Genomy mikrosporidií

21
Encephalitozoon intestinalis (2,3 MB) – redukce hlavně na koncích chromozomů a ztráty ORF s neznámou funkcí Corradi a kol. 2011

22
Genomy mikrosporidií

23
Enterocytozoon bieneusi (6 MB) postrádá enzymy pro metabolizmus cukrů
Core carbon metabolic pathways believed to form the backbone of energy metabolism in microsporidia. Enzymes are in red and metabolites in black. All the enzymes shown are present in the genomes of Encephalitozoon cuniculi, Antonospora locustae, Nosema ceranae, and Octosporea bayeri, with the single exception of lactonase (in gray) which has not been identified. In contrast, only two enzymes in any of these pathways are present in Enterocytozoon bieneusi (shown in red on black). Keeling a kol. 2010 Je to také jediný eukaryotický genom, ve kterém nebyly nalezeny introny ani geny pro sestřih
 which has not been identified. In contrast, only two enzymes in any of these. pathways are present in Enterocytozoon bieneusi (shown in red on black). Keeling a kol Je to také jediný eukaryotický genom, ve kterém nebyly nalezeny introny ani geny pro sestřih.")
24
Dochází k překryvu transkriptů
11% transkriptů u mikrosporidie Antonospora locustae obsahuje více než jeden ORF (otevřený čtecí rámec). U Encephalitozoon cuniculi to je až 80 % zkoumaných transkriptů. Nejedná se o polycystronickou transkripci jako u bakterií nebo trypanosom. Exprimován je v tomto případě zřejmě vždy jen jeden gen. Často jsou v transkriptu jen necelé kusy genů před nebo za exprimovaným genem. Často jsou geny kódovány v opačných směrech, takže nemohou být oba translatovány. K překryvu transkriptů došlo zřejmě proto, že v krátkých mezigenových oblastech nebylo dost místa na regulační sekvence. K překryvu transkriptů dochází častěji ve sporách Williams a kol, 2005, Corradi a kol. 2008, 2010
. U Encephalitozoon cuniculi to je až 80 % zkoumaných transkriptů. Nejedná se o polycystronickou transkripci jako u bakterií nebo trypanosom. Exprimován je v tomto případě zřejmě vždy jen jeden gen. Často jsou v transkriptu jen necelé kusy genů před nebo za exprimovaným genem. Často jsou geny kódovány v opačných směrech, takže nemohou být oba translatovány. K překryvu transkriptů došlo zřejmě proto, že v krátkých mezigenových oblastech nebylo dost místa na regulační sekvence. K překryvu transkriptů dochází častěji ve sporách. Williams a kol, 2005, Corradi a kol. 2008,")
25
Genomy mikrosporidií mají poměrně konzervované pořadí genů
Pombert a kol. 2011

26
Genomy mikrosporidií mají konzervovanější mezigenové sekvence než sekvence genů
Corradi a kol. 2011

27
Genetická variabilita v rámci druhu
Počet jednonukleotidových záměn SNP na 1000 bazí: Encephalitozool cuniculli SNPs/kb Plasmodium vivax SNPs/kb Plasmodium falciparum SNPs/kb Cryptosporidium parvum SNPs/kb Entamoeba histolytica SNPs/kb Saccharomyces cerevisiae volně žijící, a 6.1 SNPs/kb patogenní kmeny Člověk – cca 1 SNPs/kb

28
Zopakujme si, čím to je, že jsou genomy mikrosporidií tak malé?
Úbytek genů (pod 3000), možná i z důvodu parazitického způsobu života Miniaturizace mezigenových oblastí (100ky nt) a intronů (10ky nt). V jednom případě introny eliminovány zcela. Zkracování samotných proteinů Možná v důsledku miniaturizace genomu dochází k překryvu transkriptů, vysoké konzervovanosti mezigenových oblastí a malé frekvenci přestaveb genomu.
, možná i z důvodu parazitického způsobu života. Miniaturizace mezigenových oblastí (100ky nt) a intronů (10ky nt). V jednom případě introny eliminovány zcela. Zkracování samotných proteinů. Možná v důsledku miniaturizace genomu dochází k překryvu transkriptů, vysoké konzervovanosti mezigenových oblastí a malé frekvenci přestaveb genomu.")
29
Expandované genomy

30
OBRNĚNKY

31
Dinokaryon Typický dinokaryon u Gymnodinium sp.

32
Jsou neustále spiralizované
Během mitózy zůstává zachovaná jaderná membrána a mikrotubuly vřetýnka se dostávají ke chromozomům skrze kanálky. Obrázky v rámečku. Main features of dinoflagellate chromosomes revealed by TEM. (a) Thin section of Prorocentrum micans (phase contrast image in inlet) showing nuclei with permanently condensed chromosomes. (b) Chromosome detail showing fibers organized in stacked series of nested arches. (c, d)Whole mounted chromosomes showing the screw-like arrangement. (e) Phosphotungstic acid (PTA) positive stained, and (f) PTA negative stained whole mounted chromosomes. (g) Schematic representation of an entirely mounted chromosome showing two differentiated coiling regions. (h) Thin section of the dividing region comprised of fibers occasionally packed in a triangular configuration. (i) PTA-positive strain of a whole mounted chromosome shows that the sequential division starts by the duplication of the upper ends. (j) Y configuration of a whole mounted dividing chromosome. (k) Detail of division region in a whole mounted PTA-positive stained chromosome. (l) Schematic representation of the possible spatial kinetics of chromosome segregation inside the three-dimensional volume of the nucleus, in which to allow chromatid growth, the still undivided chromosome body should rotate around its own axis in a left-handed direction inside the nucleoplasm. Obrázek vpravo dole P. micans nucleus showing chromosomes (Chr) with different condensation states corresponding to different cell Costas a kol. 2005 Alverca a kol. 2007
 Thin section of Prorocentrum micans (phase contrast image in inlet) showing nuclei with permanently condensed chromosomes. (b) Chromosome. detail showing fibers organized in stacked series of nested arches. (c, d)Whole mounted chromosomes showing the screw-like arrangement. (e) Phosphotungstic acid (PTA) positive stained, and (f) PTA negative. stained whole mounted chromosomes. (g) Schematic representation of an. entirely mounted chromosome showing two differentiated coiling regions. (h) Thin section of the dividing region comprised of fibers occasionally. packed in a triangular configuration. (i) PTA-positive strain of a whole. mounted chromosome shows that the sequential division starts by the duplication. of the upper ends. (j) Y configuration of a whole mounted dividing. chromosome. (k) Detail of division region in a whole mounted PTA-positive. stained chromosome. (l) Schematic representation of the possible spatial. kinetics of chromosome segregation inside the three-dimensional volume of. the nucleus, in which to allow chromatid growth, the still undivided chromosome. body should rotate around its own axis in a left-handed direction inside. the nucleoplasm. Obrázek vpravo dole. P. micans nucleus showing chromosomes (Chr) with different condensation states corresponding to different cell. Costas a kol Alverca a kol")
33
Model uspořádání DNA v chromozomech
Chromozomy nejsou stabilizovány histony (i když v genomu jsou), ale histon-like proteiny (HLP, proteiny:DNA = 1:10) a pravděpodobně také RNA a netranskribovanou DNA. HLP Model chromozomů dinoflagelátů. (a) Schematické zobrazení chromozomů jako sloupec disků, ve kterých je DNA uložena v paralelních vláknech. Sousední disky jsou vůči sobě mírně pootočeny. (b) Průběh vláken v jednotlivých discích varíruje, nejsou úplně rovná lehce se odchylují od průměrnéhé směru. 1. Kostka ukazuje distribuci DNA vláken ve třech přilehlých vrstvách. 2. Průměrný směr vláken v jednotlivých vrstvách (oboustranné šipky). Kužely okazují směrodatnou odchylku. 3. Průměrný směr vláken je naznačen čarami, patrné je mírné pootočení směru v přilehlých vrstvách. (c) DNA může vytvářet kličky vycházející do stran z jednotlivých disků. Na těchto kličkách pravděpodobně probíhá transkripce. Další poznámka: Experiments by Liu et al. (2000) explored this topic from another point of view. These authors isolated dinoflagellate chromosomes, which were incubated with extracts of Xenopus laevis. After 20 min the rod-like chromosomes elongate and swell, losing their general fibrillar structure, which 60 min later is completely decondensed, to the point that only some chromatin fibers are still discernible. Afterwards, (90–120 min of incubation) chromatin reorganizes into tightly packed spheres, which are finally surrounded by a nuclear envelope with nuclear pores. This leads to a cell nucleus with uniformly dispersed chromatin, which is undistinguishable from that of common eukaryotes Na kličkách se odehrává transkripce Wisecaver a kol. 2011
, ale histon-like proteiny (HLP, proteiny:DNA = 1:10) a pravděpodobně také RNA a netranskribovanou DNA. HLP. Model chromozomů dinoflagelátů. (a) Schematické zobrazení chromozomů jako sloupec disků, ve kterých je DNA uložena v paralelních vláknech. Sousední disky jsou vůči sobě mírně pootočeny. (b) Průběh vláken v jednotlivých discích varíruje, nejsou úplně rovná lehce se odchylují od průměrnéhé směru. 1. Kostka ukazuje distribuci DNA vláken ve třech přilehlých vrstvách. 2. Průměrný směr vláken v jednotlivých vrstvách (oboustranné šipky). Kužely okazují směrodatnou odchylku. 3. Průměrný směr vláken je naznačen čarami, patrné je mírné pootočení směru v přilehlých vrstvách. (c) DNA může vytvářet kličky vycházející do stran z jednotlivých disků. Na těchto kličkách pravděpodobně probíhá transkripce. Další poznámka: Experiments. by Liu et al. (2000) explored this topic from another. point of view. These authors isolated dinoflagellate chromosomes, which were incubated with extracts of Xenopus laevis. After 20 min the rod-like chromosomes elongate and swell, losing. their general fibrillar structure, which 60 min later is completely. decondensed, to the point that only some chromatin. fibers are still discernible. Afterwards, (90–120 min of incubation) chromatin reorganizes into tightly packed spheres, which. are finally surrounded by a nuclear envelope with nuclear. pores. This leads to a cell nucleus with uniformly dispersed. chromatin, which is undistinguishable from that of common. eukaryotes. Na kličkách se odehrává transkripce. Wisecaver a kol")
34
Jaderný protein DVNP Gronik a kol 2013
Dostal se do dinoflagelátů horizontálním genovým přenosem z virů skupiny phycodnaviridae.

35
Evoluce podivného genomu obrněnek
HLP DVNP (HLP, protein:DNA = 1:10) (DVNP, prot.n:DNA = 1:2) (histony, prot.:DNA = 1:1)
 (DVNP, prot.n:DNA = 1:2) (histony, prot.:DNA = 1:1)")
36
Mají telomery (zeleně), takže jsou lineární
Fig. 2. In situ hybridization of the digoxigenin-labelled pAtT4 (to je próba na telomery) to P. micans cells. ( A – C ) Interphase nucleus. ( A ) DAPI staining for DNA (blue) displaying the long and numerous chromosomes densely packed inside the typical heart-shaped nucleus of this species. ( B ) pAtT4 probe detected with an FITC conjugate. ( C ) Merged image of A and B , showing the localization of telomeres at chromosome ends in close association with the nuclear envelope. ( D–H ) Isolated chromosomes of P. micans with different condensation stages show the distal localization of the single telomere signals in the haploid chromosomes. ( D , G ) DAPI. ( E ) pAtT4 signals. ( F , H ) Merged images showing the terminal position of the hybridization sites in chromosomes. Scale bar represents 10 m. Alverca a kol. 2007
 to P. micans cells. ( A – C ) Interphase nucleus. ( A ) DAPI staining for DNA (blue) displaying the long and numerous chromosomes densely packed inside the typical heart-shaped. nucleus of this species. ( B ) pAtT4 probe detected with an FITC conjugate. ( C ) Merged image of A and B , showing the. localization of telomeres at chromosome ends in close association with the nuclear envelope. ( D–H ) Isolated chromosomes. of P. micans with different condensation stages show the distal localization of the single telomere signals in the. haploid chromosomes. ( D , G ) DAPI. ( E ) pAtT4 signals. ( F , H ) Merged images showing the terminal position of the hybridization. sites in chromosomes. Scale bar represents 10 m. Alverca a kol")
37
Jsou velmi konzervované
Sekvence telomer Jsou velmi konzervované

38
Množství DNA v haploidním genomu různých obrněnek
Počty chromozomů jsou velmi různé a pohybují se od 4 (Syndinium borgeti) do 325 (Endodinium chattoni). 1pg = asi 1 GB
 do 325 (Endodinium chattoni). 1pg = asi 1 GB.")
39
Alexandrium ostenfeldii (6,2 MB GSS)
Odhadovaná velikost genomu 115 GB (16 genů) Jaeckisch a kol. 2011
 Jaeckisch a kol")
40
Některé geny se nacházejí v mnoha kopiích
Lin 2011

41
Symbiodinium minutum (1,5 Gb)
Shoguchi a kol. 2013

42
Symbiodinium minutum (1,5 Gb)
Asi genů (42% proteomu) vzniklo pravděpodobně genovými duplikacemi. Přítomny eukaryotické histony (kromě H1). 19 genů homologických s DVNPs a 15 „bakteriálních“ histone-like proteinů. Nalezli expandovanou rodinu genů RCC1 (189 kopií). Produkty těchto genů se váží na chromatin chromatin a hrají roli v genové expresi. Možná se podílejí na zvláštním uspořádání chromosomů. Mnoho těchto genů je původem z bakterií (na stromu níže jsou vyznačeny červeně). Shoguchi a kol. 2013
 vzniklo pravděpodobně genovými duplikacemi. Přítomny eukaryotické histony (kromě H1). 19 genů homologických s DVNPs a 15 „bakteriálních histone-like proteinů. Nalezli expandovanou rodinu genů RCC1 (189 kopií). Produkty těchto genů se váží na chromatin chromatin a hrají roli v genové expresi. Možná se podílejí na zvláštním uspořádání chromosomů. Mnoho těchto genů je původem z bakterií (na stromu níže jsou vyznačeny červeně). Shoguchi a kol")
43
Symbiodinium minutum Spliceosomálních intronů je velké množství a mají nekanonické hranice. Geny jsou na chromosomech uspořádány ve skupinách v jednom směru a jsou přepisovány po větších skupinách. Shoguchi a kol. 2013

44
Trans-splicing mRNA Conservation of SL RNAs (SL donor) in dinoflagellates. (A) Alignment of SL RNA and genomic sequences. Nucleotides identical in least two species are shaded, and those identical in all species are denoted by asterisks. Exon (SL) is shown in uppercase, and intron is shown in lowercase. D = T, A, or G; r = a or g; v = a, c or g; m = a or c; k = t or g. Kmi, K. micrum; Ppi, P. piscicida; Pmi, Pr. minimum; g, gene sequence; c, cDNA, with the number immediately after it indicating different forms. Numbers in parentheses are the number of identical sequences retrieved. For cDNAs of P. piscicida SL RNA, only the longest two clones are shown. (B) Schematic diagram illustrating how SL (exon) in the SL RNA is trans-spliced to pre-mRNA to yield mature mRNA with SL at the 5′ end. A putative 3′ acceptor site (i.e., SL splice site), ag, is underlined. Zhang H. et.al. PNAS 2007;104: ©2007 by National Academy of Sciences
 in dinoflagellates. (A) Alignment of SL RNA and genomic sequences. Nucleotides identical in least two species are shaded, and those identical in all species are denoted by asterisks. Exon (SL) is shown in uppercase, and intron is shown in lowercase. D = T, A, or G; r = a or g; v = a, c or g; m = a or c; k = t or g. Kmi, K. micrum; Ppi, P. piscicida; Pmi, Pr. minimum; g, gene sequence; c, cDNA, with the number immediately after it indicating different forms. Numbers in parentheses are the number of identical sequences retrieved. For cDNAs of P. piscicida SL RNA, only the longest two clones are shown. (B) Schematic diagram illustrating how SL (exon) in the SL RNA is trans-spliced to pre-mRNA to yield mature mRNA with SL at the 5′ end. A putative 3′ acceptor site (i.e., SL splice site), ag, is underlined. Zhang H. et.al. PNAS 2007;104: ©2007 by National Academy of Sciences.")
45
Zopakujme si, čím to je, že je genom obrněnek tak velký?
Většina DNA je nekódující a pravděpodobně repetitivní Kóduje velký počet unikátních genů Karenia brevis (podle transkritpomu) Alexandrium spp (podle transkritpomu) Symbiodinium minutum (podle genomu) Lingulodinium polyedrum (podle transkritpomu) Obsahuje velké genové rodiny a některé geny jsou namnoženy do tisíců kopií. Geny jsou dlouhé, protože obsahují velké množství intronů.
 Alexandrium spp (podle transkritpomu) Symbiodinium minutum (podle genomu) Lingulodinium polyedrum (podle transkritpomu) Obsahuje velké genové rodiny a některé geny jsou namnoženy do tisíců kopií. Geny jsou dlouhé, protože obsahují velké množství intronů.")
46
Další eukaryota s velkými genomy - améby
Amoeba dubia 670 GB Amoeba proteus 290 Gb Původní článek: The biochemical composition of the free-living amoebae Chaos chaos, Amoeba dubia and Amoeba proteus.

47
Další eukaryota s velkými genomy - Euglenida
1.5 GB Velké jádro s permanentně kondenzovanými chromozomy

48
Korelace mezi velikostí genomu a objemem buňky (Cavalier-Smith 2005)
Eukaryota s heterochromatickými jádry Jednobuněčná eukaryota s malým množstvím heterochromatinu Eukaryota se směsí eu- a heterochromatinu FI G . 1. Three contrasting scaling laws for genome size evolution. Law 1. Bacteria have single replicons per chromosome, so are under constant selective pressure to limit the accumulation of non-genic DNA that would lengthen replication times and slow reproductive rates. Thus their genome size (open circles) is related simply to gene numbers, which increase in proportion to metabolic and structural complexity—on average somewhat greater in larger cells (slope of 0_28). Law 2. Nuclei have numerous replicons origins per chromosome, which can be replicated simultaneously—so replication time is mechanistically independent of genome size and can be far less than in bacteria. When the nucleus evolved, DNA acquired a new function—to nucleate assembly of the nuclear envelope; in eukaryotes genome size scales proportionally to cell volume (slope 1_03, not significantly different from 1) because of this function and because the karyoplasmic volume ratio is essentially invariant with cell volume—probably because of a functionally essential quantitative balance between rates of nuclear RNA and cytoplasmic protein synthesis. The relationship is complicated by the presence of heterochromatin. It is simplest in unicellular eukaryotes with negligible amounts of heterochromatin (filled circles and line U: mostly green algae, diatoms and dinoflagellates, plus a few yeasts with tiny cells and amoebae with giant ones: Shuter et al., 1983), where the DNA/cell volume is minimal. In salamander red blood cells (S) the whole nucleus is heterochromatic and shrunken, so the DNA/cell volume ratio is about 50-fold greater. Cells with a mixture of compact heterochromatin and swollen euchromatin have intermediate DNA/cell volume ratios irrespective of whether they are unicellular (like cryptomonads: crosses) or multicellular (angiosperm meristem cells: dashed line). Law 3. Nucleomorph genome size is essentially invariant with cell volume because, unlike bacteria, their gene content is virtually constant and, unlike ordinary nuclei, their transcriptional/RNA processing needs do not increase significantly with cell volume (squares if volume measured by Beaton and Cavalier-Smith [1999] and stars if estimated from the literature); selection prevents non-coding DNA accumulating—because of its metabolic cost and perhaps also a constraint on chromosome arm length arising from the loss of higher-order chromatin folding (Cavalier- Smith, 1985b). Figure based on Shuter et al. (1983) and Cavalier-Smith and Beaton (1999). Cavalier-Smith 2005 Objem buňky (fL)
 is related simply to gene numbers, which increase in proportion to metabolic and structural complexity—on. average somewhat greater in larger cells (slope of 0_28). Law 2. Nuclei have. numerous replicons origins per chromosome, which can be replicated. simultaneously—so replication time is mechanistically independent of. genome size and can be far less than in bacteria. When the nucleus. evolved, DNA acquired a new function—to nucleate assembly of the. nuclear envelope; in eukaryotes genome size scales proportionally to cell. volume (slope 1_03, not significantly different from 1) because of this. function and because the karyoplasmic volume ratio is essentially. invariant with cell volume—probably because of a functionally essential. quantitative balance between rates of nuclear RNA and cytoplasmic. protein synthesis. The relationship is complicated by the presence of. heterochromatin. It is simplest in unicellular eukaryotes with negligible. amounts of heterochromatin (filled circles and line U: mostly green. algae, diatoms and dinoflagellates, plus a few yeasts with tiny cells and. amoebae with giant ones: Shuter et al., 1983), where the DNA/cell volume. is minimal. In salamander red blood cells (S) the whole nucleus. is heterochromatic and shrunken, so the DNA/cell volume ratio is about. 50-fold greater. Cells with a mixture of compact heterochromatin and. swollen euchromatin have intermediate DNA/cell volume ratios. irrespective of whether they are unicellular (like cryptomonads: crosses) or multicellular (angiosperm meristem cells: dashed line). Law 3. Nucleomorph genome size is essentially invariant with cell volume. because, unlike bacteria, their gene content is virtually constant and, unlike ordinary nuclei, their transcriptional/RNA processing needs do not. increase significantly with cell volume (squares if volume measured by. Beaton and Cavalier-Smith [1999] and stars if estimated from the. literature); selection prevents non-coding DNA accumulating—because. of its metabolic cost and perhaps also a constraint on chromosome arm. length arising from the loss of higher-order chromatin folding (Cavalier- Smith, 1985b). Figure based on Shuter et al. (1983) and Cavalier-Smith and. Beaton (1999). Cavalier-Smith Objem buňky (fL)")
49
Genomy trichomonád – korelace mezi velikostí genomu a objemem buňky a jádra (Zubáčová a kol. 2008)
")
50
Teorie skeletální DNA V jádře probíhá syntéza a zrání mRNA.
Větší buňky potřebují k růstu více proteinů a tedy i více mRNA. Potřeba optimalizovat (= udržovat) poměr objemu jádra a cytoplasmy. DNA slouží také jako strukturní molekula podepírající jadernou membránu a určující velikost jádra. Aby zvětšila jádro, zvětšuje buňka velikost genomu nebo mění jeho konformaci.
 poměr objemu jádra a cytoplasmy. DNA slouží také jako strukturní molekula podepírající jadernou membránu a určující velikost jádra. Aby zvětšila jádro, zvětšuje buňka velikost genomu nebo mění jeho konformaci.")
51
Mnohojadernost Snyderella
(Parabasala) Mnohojadernost Řešení jak zajistit dostatečný objem jaderného kompartmentu pro potřeby velkých buněk Opalina Acantharea Physarum polycephalum Baroella (Oxymonády)
 Mnohojadernost. Řešení jak zajistit dostatečný objem jaderného kompartmentu pro potřeby velkých buněk. Opalina. Acantharea. Physarum polycephalum. Baroella (Oxymonády)")
52
Životní cyklus dírkonošce Allogromia laticollaris
Polyploidizace Životní cyklus dírkonošce Allogromia laticollaris Téma na esej: Genomika dírkonošců Parfrey a kol. 2010, 2012 Habura a kol 2011 300±105N některé části genomu jsou zřejmě namnoženy ještě víc FIG. 1.—Nuclear dynamics during the life cycle of Allogromia laticollaris CSH. DAPI-stained individuals representing all life cycle stages observed. (A) Agamont I, (B) Juvenile Agamont II within parental test, (C) Agamont II, (D) Many juvenile Agamont I cells within parental test, (E) cell filled with gametic nuclei, and (F) Cell filled with zygotic nuclei. All scale bars 50 lm. Redrawn from McEnery and Lee (1976). 83±25MB 100±35N
 Agamont I, (B) Juvenile Agamont II within parental test, (C) Agamont II, (D) Many juvenile Agamont I cells within parental test, (E) cell filled with. gametic nuclei, and (F) Cell filled with zygotic nuclei. All scale bars 50 lm. Redrawn from McEnery and Lee (1976). 83±25MB. 100±35N.")
53
Podivné genomy

54
NÁLEVNÍCI

55
Obecný životní cyklus nálevníků

56
Transformace mikro- na makronukleus
Vystřižení internal eliminated segments (IESs) – modré oblasti. Nalámání mikronukleárních chromozomů v poměrně přesně určených bodech zlomu – červené oblasti. Vystřižení většiny repetitivní DNA. Dosyntetizování telomer na makronukleární chromozomy – telomery jsou označeny jako šipky. Namnožení makronukleárních chromozomů. Eisen a kol., 2006
 – modré oblasti. Nalámání mikronukleárních chromozomů v poměrně přesně určených bodech zlomu – červené oblasti. Vystřižení většiny repetitivní DNA. Dosyntetizování telomer na makronukleární chromozomy – telomery jsou označeny jako šipky. Namnožení makronukleárních chromozomů. Eisen a kol.,")
57
Tetrahymena thermophila
Mikronukleus – diploidní, 5 chromozomových párů Makronukleus – 250 – 300 typů chromozomů, cca 104 Mbp, více než předpovězených proteinů (20 – 25 tis. v genomu člověka). Asi 6000 IES je odstraněno při tvorbě MACu, haploidní genom MAC je tedy asi o 10-20% menší než MIC. rDNA chromozom obsahující geny pro 5.8S, 17S, and 26S rRNA se v MAC vyskytuje asi v 9000 kopiích. Šest dalších sledovaných. chromozomů se vyskytuje průměrně v 45 kopiích. MAC chromozomy nemají centromery Velikostní distribuce makronukleárních scaffoldů s telomerami na obou stranách – tj. chromozomů Eisen a kol., 2006
. Asi 6000 IES je odstraněno při tvorbě MACu, haploidní genom MAC je tedy asi o 10-20% menší než MIC. rDNA chromozom obsahující geny pro 5.8S, 17S, and 26S rRNA se v MAC vyskytuje asi v 9000 kopiích. Šest dalších sledovaných. chromozomů se vyskytuje průměrně v 45 kopiích. MAC chromozomy nemají centromery. Velikostní distribuce makronukleárních scaffoldů s telomerami na obou stranách – tj. chromozomů. Eisen a kol.,")
58
Tetrahymena thermophila
Dělení makronukleu „Amitózou“. Před dělením makronukleu se haploidní genom sdruží do „kuličky“ – Macronuclear genome unit. Macronuclear genome unit ~ haploidní genom makronukleu Endo a Sugai 2011

59
Makronukleární genomy jiných nálevníků
Swart a kol. 2013

60
Tvorba makronukleu u spirotrichních nálevníků
Téma na esej: Tvorba makronukleu Swart a kol. 2013, Heyse a kol. 2010, Orias a kol. 2011, Goldman a Landweber 2012, Bracht a kol, 2013

61
ZÁVĚR Zástupci protist se velmi liší velikostí genomu 2,3MB-600GB(?)
Malé genomy se vyznačují větší hustotou genů, menším počtem genů, kratšími proteiny, překryvem transkriptů, menším počtem (v jednom případě i ztrátě) intronů nebo zmenšováním jejich velikosti (až na 18 nt). Velké genomy se vyznačují velkým množství nekódující DNA a zmnožením genů. Velké genomy jsou málo prozkoumané a odhad jejich velikosti může být velmi nepřesný. Velikost genomu souvisí s velikostí buňky a jádra, protože DNA má také strukturální funkci. V životním cyklu nálevníků dochází k přestavbě mikronukleu na makronukleus, při které dochází k fragmentaci chromosomů, eliminaci některých úseků, přestavbě genů a navýšení ploidie.
 intronů nebo zmenšováním jejich velikosti (až na 18 nt). Velké genomy se vyznačují velkým množství nekódující DNA a zmnožením genů. Velké genomy jsou málo prozkoumané a odhad jejich velikosti může být velmi nepřesný. Velikost genomu souvisí s velikostí buňky a jádra, protože DNA má také strukturální funkci. V životním cyklu nálevníků dochází k přestavbě mikronukleu na makronukleus, při které dochází k fragmentaci chromosomů, eliminaci některých úseků, přestavbě genů a navýšení ploidie.")
62
KONEC

Podobné prezentace

>")









>")
>")






