Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
OBECNÁ MYKOLOGIE (místy se zvláštním zřetelem k makromycetům)
MODULARIZACE VÝUKY EVOLUČNÍ A EKOLOGICKÉ BIOLOGIE CZ.1.07/2.2.00/ OBECNÁ MYKOLOGIE (místy se zvláštním zřetelem k makromycetům) Vymezení pojmů „houby“ a „mykologie“ • Historický výskyt a teorie o původu hub • Stavba houbové buňky (cytoplazma, organely, jádro a bun. cyklus, bun. stěna) • Výživa a obsahové látky hub • Vegetativní stélka hub (nemyceliální houby, hyfy, hyfové útvary, pletivné útvary, stélka lišejníků, růst houbové stélky) • Rozmnožování hub (vegetativní, nepohlavní, pohlavní) • Genetika hub • Plodnice hub (sporokarpy, askokarpy, bazidiokarpy, anatomie plodnic, hymenofor, hymeniální elementy) • Spory hub (typy a stavba, šíření a klíčení) • Nomenklatura hub • Sběr, určování a konzervace hub mykolog
 Vymezení pojmů „houby a „mykologie • Historický výskyt a teorie o původu hub. • Stavba houbové buňky (cytoplazma, organely, jádro a bun. cyklus, bun. stěna) • Výživa a obsahové látky hub • Vegetativní stélka hub (nemyceliální houby, hyfy, hyfové útvary, pletivné útvary, stélka lišejníků, růst houbové stélky) • Rozmnožování hub (vegetativní, nepohlavní, pohlavní) • Genetika hub. • Plodnice hub (sporokarpy, askokarpy, bazidiokarpy, anatomie plodnic, hymenofor, hymeniální elementy) • Spory hub (typy a stavba, šíření a klíčení) • Nomenklatura hub • Sběr, určování a konzervace hub. mykolog.")
2
SPORY HUB Jako spory (česky výtrusy) jsou označovány buňky sloužící k reprodukci (případně též k přežívání určitého období), z nichž může po vyklíčení vzniknout nový jedinec (na rozdíl od semen vyšších rostlin nejsou tvořeny specializovanými strukturami a jsou jednobuněčné – sporou může být nazvána každá buňka, odpovídající uvedené charakteristice); charakteristickými znaky těchto buněk (ve srovnání s vegetativními) bývá silnější stěna, hustší cytoplazma a pomalejší metabolismus. Podle způsobu vzniku lze rozlišit dva základní typy – mitospory (zoospory, sporangiospory, konidie, viz kapitolu rozmnožování hub) a meiospory (zygo-, asko-, bazidiospory). Rozměry spor jsou mikroskopické, v rozsahu zhruba od 2 do 200 mikrometrů. Tvar a ornamentika výtrusů a barva stěny (určující pak barvu výtrusného prachu) jsou druhově stálými znaky, sloužícími jako dobrá pomůcka pro určování. V přírodě hrají spory hub úlohu při šíření druhu a obsazování nových stanovišť => přínosem může být genetická výměna s jinými jedinci tam rostoucími, případně velký "nálet" spor může být základem k prosazení v kompetičním "boji" s jinými organismy.
 jsou označovány buňky sloužící k reprodukci (případně též k přežívání určitého období), z nichž může po vyklíčení vzniknout nový jedinec (na rozdíl od semen vyšších rostlin nejsou tvořeny specializovanými strukturami a jsou jednobuněčné – sporou může být nazvána každá buňka, odpovídající uvedené charakteristice); charakteristickými znaky těchto buněk (ve srovnání s vegetativními) bývá silnější stěna, hustší cytoplazma a pomalejší metabolismus. Podle způsobu vzniku lze rozlišit dva základní typy – mitospory (zoospory, sporangiospory, konidie, viz kapitolu rozmnožování hub) a meiospory (zygo-, asko-, bazidiospory). Rozměry spor jsou mikroskopické, v rozsahu zhruba od 2 do 200 mikrometrů. Tvar a ornamentika výtrusů a barva stěny (určující pak barvu výtrusného prachu) jsou druhově stálými znaky, sloužícími jako dobrá pomůcka pro určování. V přírodě hrají spory hub úlohu při šíření druhu a obsazování nových stanovišť => přínosem může být genetická výměna s jinými jedinci tam rostoucími, případně velký nálet spor může být základem k prosazení v kompetičním boji s jinými organismy.")
3
• Oomycota vytvářejí akrokontní (s bičíky na přední straně buňky)
TYPY A STAVBA SPOR Jednoduché organismy se rozmnožují a šíří nejčastěji nepohlavně vznikajícími zoosporami: • Oomycota vytvářejí akrokontní (s bičíky na přední straně buňky) primární zoospory, ty se encystují a z cyst následně vyrejdí sekundární zoospory, které jsou pleurokontní (s bičíky vycházejícími na boku); vzhledem k příbuznosti s heterokontními řasami mají i Oomycota dva různé bičíky, delší péřitý (se dvěma řadami tubulárních mastigonemat) a kratší hladký (obdobný pár bičíků mají zoospory labyrintul). Tato dvě bičíkatá stadia v životním cyklu mají druhy diplanetické (běžné mezi Saprolegniales), redukce primárních zoospor vede k monoplanetismu a nakonec u suchozemských aplanetických druhů žádné zoospory nevyrejdí a sporangia klíčí přímo hyfou; šíření Peronosporales zajišťují odlomená sporangia, typicky monosporická – vzhledem k podobě a klíčení hyfou označovaná mylně za konidie (pravá konidie však má jednu buněčnou stěnu, zatímco zde jsou dvě: vnější vrstva patří sporangiu, vnitřní pak obsažené spoře). • Dva různé bičíky mají i zoospory nádorovek (Plasmodiophoromycota). T. Kalina et J. Váňa: Sinice, řasy, houby, mechorosty a podobné organismy v současné biologii. Karolinum, Praha, (Překresleno z: X. Llimona et al.: Fongos i líquens. Barcelona, 1991)
 primární zoospory, ty se encystují. a z cyst následně vyrejdí sekundární. zoospory, které jsou pleurokontní. (s bičíky vycházejícími na boku); vzhledem k příbuznosti s heterokontními řasami mají i Oomycota dva různé bičíky, delší péřitý (se dvěma řadami tubulárních mastigonemat) a kratší hladký (obdobný pár bičíků mají zoospory labyrintul). Tato dvě bičíkatá stadia v životním cyklu mají druhy diplanetické (běžné mezi Saprolegniales), redukce primárních zoospor vede k monoplanetismu a nakonec u suchozemských aplanetických druhů žádné zoospory nevyrejdí a sporangia klíčí přímo hyfou; šíření Peronosporales zajišťují odlomená sporangia, typicky monosporická – vzhledem k podobě a klíčení hyfou označovaná mylně za konidie (pravá konidie však má jednu buněčnou stěnu, zatímco zde jsou dvě: vnější vrstva patří sporangiu, vnitřní pak obsažené spoře). • Dva různé bičíky mají i zoospory nádorovek (Plasmodiophoromycota). T. Kalina et J. Váňa: Sinice, řasy, houby, mechorosty a podobné organismy v současné biologii. Karolinum, Praha, (Překresleno z: X. Llimona et al.: Fongos i líquens. Barcelona, 1991)")
4
• Naproti tomu na zoosporách odd
• Naproti tomu na zoosporách odd. Chytridiomycota (v pomalu, ale jistě opouštěném pojetí Chytridiomycota sensu lato, tedy včetně Blastocladi-ales a Neocallimastigales, hodno-cených dnes na úrovni samostat-ných oddělení) lze demonstrovat příbuznost hub a živočichů – právě u těchto organismů se totiž tvoří pohyblivá stadia s jedním opisto-kontním (na zadní straně buňky umístěným) neboli tlačným bičíkem (výjimkou jsou Neocallimastigales s více bičíky). V poslední době bylo uznáváno pět řádů a u každého lze najít specifickou ultrastrukturu zoo-spor; u řádů Chytridiales a Mono-blepharidales byl na zoosporách pozorován tzv. rumposom, propo-jený mikrotubuly s kinetosomem a považovaný za fotoreceptor. T. Kalina et J. Váňa: Sinice, řasy, houby, mechorosty ..., 2005.
 lze demonstrovat příbuznost hub a živočichů – právě u těchto organismů se totiž tvoří pohyblivá stadia s jedním opisto-kontním (na zadní straně buňky umístěným) neboli tlačným bičíkem (výjimkou jsou Neocallimastigales s více bičíky). V poslední době bylo uznáváno pět řádů a u každého lze najít specifickou ultrastrukturu zoo-spor; u řádů Chytridiales a Mono-blepharidales byl na zoosporách pozorován tzv. rumposom, propo-jený mikrotubuly s kinetosomem a považovaný za fotoreceptor. T. Kalina et J. Váňa: Sinice, řasy, houby, mechorosty ...,")
5
K aktivnímu uvolňování a šíření může dojít v případě
Pohlavně vzniklé silnostěnné útvary (cysty nádorovek, oospory u oddělení Oomycota, trvalá sporangia chytridií) slouží hlavně k přečkání určitého období v prostředí (v našich podmínkách obvykle přezimování) před vyklíčením hyfy nebo vyrejděním zoospor. Hlenky vytvářejí silnostěnné spory, které se pasivně uvolňují ze sporokarpů a následně z nich vyjdou myxomonády. Zygospory spájivých hub (principiálně se jedná o zygosporangium obsahující jednu zygosporu, ale běžně jsou tyto pojmy slučovány) se uvolňují pasivně a jejich velikost a silná stěna je předurčuje k přežití delšího období, ne však k šíření na velké vzdálenosti. K aktivnímu uvolňování a šíření může dojít v případě nepohlavních spor vznikajících ve sporangiích. Příkladem je Pilobolus (česky měchomršť), jenž má pod sporangiem ztlustlinu (subsporangiální vak), působící jako čočka, která soustřeďuje světlo => natočení sporangia za světlem; při dostatku vody (srážející se kapky, typicky rosa v přirozeném pro- středí houby) ve ztlustině vzrůstá turgor, až nakonec dojde k odtržení a odmrštění celého sporangia (vlastní odmrštění asi přímo nezávisí na světle – právě tato houba v případě "zatemnění" uplatní své "biologické hodiny", viz dále).
 slouží hlavně k přečkání určitého období v prostředí (v našich podmínkách obvykle přezimování) před vyklíčením hyfy nebo vyrejděním zoospor. Hlenky vytvářejí silnostěnné spory, které se pasivně uvolňují ze sporokarpů a následně z nich vyjdou myxomonády. Zygospory spájivých hub (principiálně se jedná o zygosporangium obsahující jednu zygosporu, ale běžně jsou tyto pojmy slučovány) se uvolňují pasivně a jejich velikost a silná stěna je předurčuje k přežití delšího období, ne však k šíření na velké vzdálenosti. K aktivnímu uvolňování a šíření může dojít v případě. nepohlavních spor vznikajících ve sporangiích. Příkladem je Pilobolus (česky měchomršť), jenž má. pod sporangiem ztlustlinu (subsporangiální vak), působící jako čočka, která soustřeďuje světlo => natočení sporangia za světlem; při dostatku vody. (srážející se kapky, typicky rosa v přirozeném pro- středí houby) ve ztlustině vzrůstá turgor, až nakonec. dojde k odtržení a odmrštění celého sporangia. (vlastní odmrštění asi přímo nezávisí na světle – právě tato houba. v případě zatemnění uplatní své biologické hodiny , viz dále).")
6
Askospory vřeckatých hub (poznámka: ve striktním pojetí by se neměly označovat jako meiospory, protože při typickém počtu 8 jejich vytvoření přímo předchází "dodatečná" mitóza a nevznikají tedy přímo meioticky – nicmémě běžně se o nich mluví jako o meiosporách) se tvoří ve vřeckách, z nichž vycházejí ven různými způsoby: • pasivní uvolňování u primitivních skupin – rozpad stěn prototunikátních vřecek; • naproti tomu u odvozenějších typů je uvolňování aktivní (vřecka unitunikátní a bitunikátní): s dozráváním stoupá turgor cytoplazmy uvnitř vřecka => na vrcholu praská stěna nebo se odtrhne víčko, stěna vřecka se smrští => spory jsou vytlačeny ven; protože spory vřeckatých hub při svém uvolnění směrem nahoru musejí překonávat odpor vzduchu i gravitaci, je potřeba značné síly na jejich vystřelení (také mají značný "dostřel" – v extrému až 3 metry) a jsou obvykle větší a těžší než spory hub stopkovýtrusných. Podzemní plodnice (např. Tuberaceae) vydávají intenzivní vůni, jež láká živočichy, kteří pak mechanicky rozruší pevný obal plodnice (tu obvykle sežerou a spory šíří ve svých útrobách, viz dále u endozoochorie). Půdní houby jako např. Fusarium nebo Trichoderma jsou příkladem hub tvořících slizké spory (resp. konidie), které pak na sebe mohou nalepit různé organismy.
: s dozráváním stoupá turgor cytoplazmy uvnitř vřecka => na vrcholu praská stěna nebo se odtrhne víčko, stěna vřecka se smrští => spory jsou vytlačeny ven; protože spory vřeckatých hub při svém uvolnění směrem nahoru musejí překonávat odpor vzduchu i gravitaci, je potřeba značné síly na jejich vystřelení (také mají značný dostřel – v extrému až 3 metry) a jsou obvykle větší a těžší než spory hub stopkovýtrusných. Podzemní plodnice (např. Tuberaceae) vydávají intenzivní vůni, jež láká živočichy, kteří pak mechanicky rozruší pevný obal plodnice (tu obvykle sežerou a spory šíří ve svých útrobách, viz dále u endozoochorie). Půdní houby jako např. Fusarium nebo Trichoderma jsou příkladem hub tvořících slizké spory (resp. konidie), které pak na sebe mohou nalepit různé organismy.")
7
U některých hub dochází k přerůstání zralých vřecek
Spory jsou běžně jednobuněčné, není však vzácností tvorba mnoho-buněčných spor – např. u jazourků (čeleď Geoglossaceae) se tvoří mnohobuněčné jehlicovité spory až 200 µm dlouhé (každá buňka může vyklíčit vlastní hyfou). Richard T. Hanlin: Illustrated Genera of Ascomycetes, vol. 2. APS Press, St. Paul, 1997. U některých hub dochází k přerůstání zralých vřecek nad úroveň thecia (= hymenia, u plodnic typu apothecium) nebo k prorůstání vřecek ven z ostiola (v případě perithecií). /Konidie a artrospory (nepohlavní spory vřeckatých a vzácněji i stopkovýtrusných hub) a jejich vznik byly popsány v textu o nepohlavním a vegetativním rozmnožování./
 se tvoří mnohobuněčné jehlicovité spory až 200 µm dlouhé (každá buňka může vyklíčit vlastní hyfou). entry=Trichoglossum%20hirsutum. Richard T. Hanlin: Illustrated Genera of Ascomycetes, vol. 2. APS Press, St. Paul, U některých hub dochází k přerůstání zralých vřecek. nad úroveň thecia (= hymenia, u plodnic typu apothecium) nebo k prorůstání vřecek ven z ostiola (v případě perithecií). /Konidie a artrospory (nepohlavní spory vřeckatých a vzácněji i stopkovýtrusných hub) a jejich vznik byly popsány v textu o nepohlavním a vegetativním rozmnožování./")
8
Pleurotus ostreatus Bazidiospory stopkovýtrusných hub (ani zde to není s termínem meiospory jednoznačné, protože buňkami vzniklými přímo meiozou vlastně nejsou spory, ale haplocyty) jsou typicky jednobuněčné a základ-ní tvar je eliptický, obvykle drobně prohloubený (supra-apikulární deprese) nad apikulem ("stopečka" v místě, kde spora vyrůstá ze sterigmatu), kde po odlomení zůstává "jizva" - hilum (mnohdy je jako hilum označován apikulus). Rozměry jsou drobné (řádově několik mikrometrů), vzhledem k vypadávání z hymenoforu nebo vyfoukávání z teřichu nepotřebují být "dělové koule" jako u vřeckatých hub a naopak je výhodou být malé a lehké pro snadný přenos. Clémençon: Cytology and Plectology of the Hymenomycetes, 2004. Foto Merton F. Brown et Harold G. Brotzman,
 jsou typicky jednobuněčné a základ-ní tvar je eliptický, obvykle drobně prohloubený (supra-apikulární deprese) nad apikulem ( stopečka v místě, kde spora vyrůstá ze sterigmatu), kde po odlomení zůstává jizva - hilum (mnohdy je jako hilum označován apikulus). Rozměry jsou drobné (řádově několik mikrometrů), vzhledem k vypadávání z hymenoforu nebo vyfoukávání z teřichu nepotřebují být dělové koule jako u vřeckatých hub a naopak je výhodou být malé a lehké pro snadný přenos. Clémençon: Cytology and Plectology of the Hymenomycetes, Foto Merton F. Brown et Harold G. Brotzman,")
9
Nejpůvodnější(?) způsob uvolňování je repetitivní "pučení spor" u "stopkovýtrus-ných kvasinek" (Sporobolomyces), kde se vytvoří sterigma => spora a po jejím odlomení se může vytvořit z téže buňky zas další sterigma => spora ... Tvorbě spor u typických zástupců oddělení Basidiomycota předchází meiotické dělení a formování sterigmat, do kterých vstupují vzniklá haploidní jádra => na vrcholcích sterigmat se vytváří apofýzy (protržení stěny bazidie => na povrchu apofýzy se vytváří nová stěna, odlišná od stěn hyf a bazidií), asymetricky narůstají (na vrcholcích sterigmat, ale do strany) a po vstupu jader se z nich formují spory. Heinz Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica, vol J. Cramer, Berlin-Stuttgart, 2004.
, asymetricky narůstají (na vrcholcích sterigmat, ale do strany) a po vstupu jader se z nich formují spory. Heinz Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica, vol J. Cramer, Berlin-Stuttgart,")
10
Heinz Clémençon: Cytology and Plectology of the Hymenomycetes
Heinz Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica, vol. 199. Berlin-Stuttgart, 2004. Zároveň u báze bazidie narůstají vakuoly (nebo jedna velká vakuola), která vtlačí cytoplazmu skrz sterig-mata do tvořících se spor; ty se tvoří typicky v počtu čtyř na jedné bazidii, ale u různých druhů se mohou tvořit různé počty od 2 do 8. U většiny druhů totiž dochází po meioze ještě k mitóze ("třetí jaderné dělení"), ale obvykle čtyři ze vzniklých jader zůstanou v bazidii a abortují.
, která vtlačí cytoplazmu skrz sterig-mata do tvořících se spor; ty se tvoří typicky v počtu čtyř na jedné bazidii, ale u různých druhů se mohou tvořit různé počty od 2 do 8. U většiny druhů totiž dochází po meioze ještě k mitóze ( třetí jaderné dělení ), ale obvykle čtyři ze vzniklých jader zůstanou v bazidii a abortují.")
11
Ke vzniku bisporické bazidie může dojít několika cestami: některá jádra zůstanou v bazidii a abortují nebo se vytvoří čtyři jádra, ale jen dvě sterigmata => vzniknou dvojjaderné spory, které mohou být i heterokary-otické (případ sekundárního homo-thalismu jako u Agaricus bisporus, viz obr. vlevo), nebo nedochází ke karyo gamii a meioze, ale jen k mitóze. Poznámka: jen vzácně vznikají spory stopkovýtrusných hub partenogeneticky, tedy jen v haploidním cyklu bez plazmogamie, karyogamie a meiozy – jak patrno výše, vznik bisporických bazidií zdaleka nemusí být důsledkem partenogeneze. Zvláštností některých tropických druhů je dimorfismus spor – tvorba dvou typů spor výrazně jiné velikosti na téže plodnici. Heinz Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica, vol J. Cramer, Berlin-Stuttgart, 2004.

12
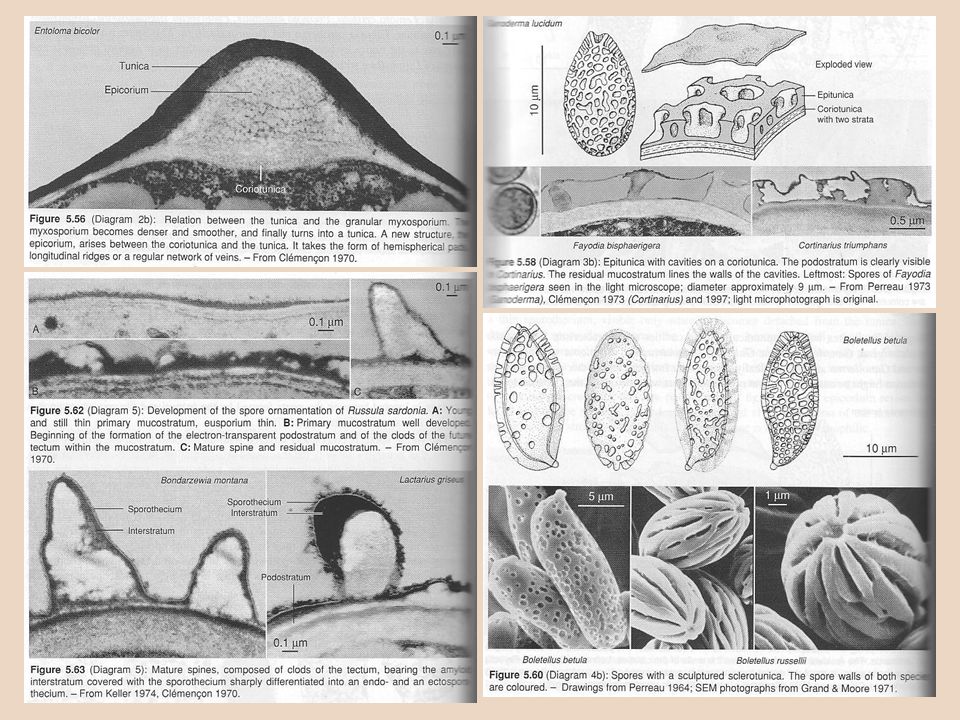
Stěna výtrusů je u různých druhů hladká nebo ornamentovaná, bezbarvá nebo zbarvená; skládá se z několika vrstev: • vnitřní eusporium (vzniká z vnitřní vrstvy stěny apofýzy) je obvykle bezbarvé, dělí se na: – endosporium – zcela vnitřní vrstva stěny; – episporium – základ stěny spory, pevná a bezbarvá vrstva. Jiné dělení eusporia (na základě elektronové mikroskopie): uvnitř průsvitné corium, vně tmavší coriotunika (u rodu Entoloma rozlišeno též epicorium – vnější vrstva pod myxosporiem, která se podílí na formování ornamentiky; obr. viz dále spolu s tunikou). H. Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica 199. J. Cramer, Berlin-Stuttgart, 2004.
: uvnitř průsvitné corium, vně tmavší coriotunika (u rodu Entoloma rozlišeno též epicorium – vnější vrstva pod myxosporiem, která se podílí na formování ornamentiky; obr. viz dále spolu s tunikou). H. Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica 199. J. Cramer, Berlin-Stuttgart,")
13
Schizophyllum nebo Mycena tvoří tenkostěnné, hladké a bezbarvé spory bez eusporia – stěnou spory zůstává stěna apofýzy (protože ta je odlišná od stěn hyf, projeví se odlišnost ve stěně při klíčení hyfy, viz dále). Spory některých Cortinariaceae mají na vrcholu papilu – oblast ztlustlého eusporia (coriotuniky). Clémençon: Cytology and Plectology ..., 2004 Tlustostěnné spory řady druhů mají na vrcholku založen klíční pór – "otvor" v episporiu, vyplněný strukturálně i chemicky odlišnou hmotou z endosporia tvořící "dřeň", "víčko" (z čel. Strophariaceae, Coprinaceae, Bolbitiaceae) nebo "zátku" (bedly aj. Agaricaceae). Malé klíční póry vznikají lokálním ztenčením eusporia.
. Clémençon: Cytology and Plectology ..., Tlustostěnné spory řady druhů mají na vrcholku založen klíční pór – otvor v episporiu, vyplněný strukturálně i chemicky odlišnou hmotou z endosporia tvořící dřeň , víčko (z čel. Strophariaceae, Coprinaceae, Bolbitiaceae) nebo zátku (bedly aj. Agaricaceae). Malé klíční póry vznikají lokálním ztenčením eusporia.")
14
víčko struktura běžná u tmavovýtrusých hub
Clémençon: Cytology and Plectology ..., 2004. zátka malé klíční póry dřeň víčko struktura běžná u tmavovýtrusých hub coriotunika corium Pozn.: silná obou-stranně vypouklá víčka jsou označo-vána jako „čočky“.

15
za zralosti slizovatí a mizí.
• vnější myxosporium (vzniká z vnější vrstvy stěny apofýzy) – zde se v průběhu dozrávání tvoří mechanicky odolné struktury + případná ornamentika; dělí se na: – exosporium – zprvu gelatinózní, posléze chrupavčitá a dosti odolná vrstva; zde se formuje ornamentika a zbarvení stěny; – perisporium – vnější vrstva zralých spor, gelatinózní a bezbarvá, často rozpraskaná; – ectosporium – zcela vnější tenká vrstva, za zralosti slizovatí a mizí. Clémençon: Cytology and Plectology ..., 2004 Myxosporium se vytváří jako tzv. primární mukostratum => může zůstat jako měkká struktura (reziduální mukostratum), ale obvykle struktura tvrdne => mění se na (podle stoupající tvrdosti) tuniku, epituniku, sklerotuniku nebo tectum; případně báze myxosporia ztvrdne v podostratum, z nějž se vytváří ornamentika.
 – zde se v průběhu dozrávání tvoří mechanicky odolné struktury + případná ornamentika; dělí se na: – exosporium – zprvu gelatinózní, posléze chrupavčitá a dosti odolná vrstva; zde se formuje ornamentika a zbarvení stěny; – perisporium – vnější vrstva zralých spor, gelatinózní a bezbarvá, často rozpraskaná; – ectosporium – zcela vnější tenká vrstva, za zralosti slizovatí a mizí. Clémençon: Cytology and Plectology ..., Myxosporium se vytváří jako tzv. primární mukostratum => může zůstat jako měkká struktura (reziduální mukostratum), ale obvykle struktura tvrdne => mění se na (podle stoupající tvrdosti) tuniku, epituniku, sklerotuniku nebo tectum; případně báze myxosporia ztvrdne v podostratum, z nějž se vytváří ornamentika.")
17
Zvláštními modifikacemi myxosporia jsou expandující myxosporium (odděluje se od eusporia, vzniká "perisporiální vak") nebo "pláž" (oblast bez ornamentiky v supra-apikulární oblasti). Heinz Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica, vol J. Cramer, Berlin-Stuttgart, 2004.

18
Typické pro rouškaté houby jsou aktivně odmršťované balistospory.
Na stěně spory (v oblasti supra-apikulární deprese) se vytváří plochá kapka a zároveň zformovaná spora vylučuje kapku v místě, kde přirůstá na sterigma (je známa pod názvem hilární nebo Bullerova kapka – nejedná se o čistou vodu, ale hygroskopický roztok cukrů a proteinů) ... Heinz Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica, vol. 199. J. Cramer, Berlin-Stuttgart, 2004. převzato z: McLaughlin et al.: Ultrastructure and evolution of ballistosporic basidiospores. Bot. J. Linnean Soc. 91: , 1985.
 se vytváří plochá kapka a zároveň zformovaná spora vylučuje kapku v místě, kde přirůstá na sterigma (je známa pod názvem hilární nebo Bullerova kapka – nejedná se o čistou vodu, ale hygroskopický roztok cukrů a proteinů) ... Heinz Clémençon: Cytology and Plectology of the Hymenomycetes. Bibliotheca Mycologica, vol J. Cramer, Berlin-Stuttgart, převzato z: McLaughlin et al.: Ultrastructure and evolution of ballistosporic basidiospores. Bot. J. Linnean Soc. 91: ,")
19
=> současně v oblasti apikulu centripetálně vroste přepážka mezi sporu a sterigma a v témže místě se zeslabí stěna (u tenkostěnných spor se stěna prostě zbortí, u tlustostěnných vzniká pod vlastní přepážkou "zátka", případně ještě bazální čepička) => Bullerova kapka postupně narůstá, ale ve finále rychle nasaje vlhkost z prostředí (příklad: pětinásobné zvětšení během 5–30 sekund) => zvětšená kapka dosáhne kontaktu s plochou kapkou na stěně spory a dojde k jejich rychlému splynutí => prudká změna těžiště odtrhne a "odstřelí" sporu od sterigmatu v místě přepážky, kde zbyde "jizva" – hilum (energii dodá též uvolnění povrchového napětí při splynutí kapek) => osiřelá bazidie se následně "bortí". Clémençon: Cytology and Plectology of the Hymenomycetes.
 => zvětšená kapka dosáhne kontaktu s plochou kapkou na stěně spory a dojde k jejich rychlému splynutí. => prudká změna těžiště odtrhne a odstřelí sporu od sterigmatu v místě přepážky, kde zbyde jizva – hilum. (energii dodá též uvolnění povrchového napětí při splynutí kapek) => osiřelá bazidie se následně bortí . Clémençon: Cytology and Plectology of the Hymenomycetes.")
20
Balistická dráha vystřelené spory je označována termínem sporabola; na rozdíl od "extrémních výkonů" spor vřeckatých hub jsou zde spory vystřelovány na vzdálenost v řádu desetin milimetru (max. 1,5 mm) a "dostřel" u různých druhů obvykle je v přímé úměře ke vzdálenosti sousedních lupenů (nebo ostnů, průměru pórů aj.) – jde o to, aby spory úspěšně vypadly z hymeno-foru, málokdy se uchytí na protějším lupenu (ostnu, stěně póru aj.). Podobné vystřelování je známo i u některých konidií (například u Tremellales) – tzv. balistokonidie. Heinz Clémençon: Cytology and Plectology of the Hymenomycetes, 2004. Dozrávání spor probíhá u lupenatých hub víceméně současně (izotropické hymenium) nebo postupně – na jednom lupenu mohou být spory v různých fázích vývoje (formující se bazidie, zralé spory i zborcené bazidie po oddělení spor – anizotropické hymenium, viz kapitolu Plodnice hub). Faktory, které ovlivňují uvolňování výtrusů z plodnice, jsou na jedné straně genetické faktory a biologický rytmus houby a na straně druhé vliv klimatických podmínek (vlhkost, teplota, vítr).
 – tzv. balistokonidie. Heinz Clémençon: Cytology and Plectology of the Hymenomycetes, Dozrávání spor probíhá u lupenatých hub víceméně současně (izotropické hymenium) nebo postupně – na jednom lupenu mohou být spory v různých fázích vývoje (formující se bazidie, zralé spory i zborcené bazidie po oddělení spor – anizotropické hymenium, viz kapitolu Plodnice hub). Faktory, které ovlivňují uvolňování výtrusů z plodnice, jsou na jedné straně genetické faktory a biologický rytmus houby a na straně druhé vliv klimatických podmínek (vlhkost, teplota, vítr).")
21
Číšenky a pohárovky mají pohárkovité plodnice, kryté
Břichatky nemají balistospory; spory jsou uzavřeny v plodnici a uvolňují se za zralosti otvorem – peristomem. Jejich uvolňování může napomáhat déšť nebo nárazy vátého písku na plodnici => dojde ke stlačení plodnice a vyfouknutí spor. Botany. Wm.C. Brown Publishers, 1995. K.R. Stern, D. Vodopich: R. Moore, W.D. Clark, Foto Paľo °; /obrazok_detail.php?obrazok_id=143712 peridiola funikulus Crucibulum laeve /obrazok_detail.php?obrazok_id=143694 Foto: Paľo °; řez peridiolou bazidiospory Číšenky a pohárovky mají pohárkovité plodnice, kryté v mládí tenkou blankou – epifragmou (v průběhu dozrávání se roztrhá podobně jako plachetky rouškatých hub). Uvnitř nalezneme drobné peridioly ("plodničky" obsahující spory), přichycené na dně pohárku vláknem – funikulem => dopadem dešťové kapky je peridiola vymrštěna ven a funikulem se uchytí na trávě => čeká až ji sežere býložravec => teprve po průchodu jeho trávicím traktem spory klíčí.
. Uvnitř nalezneme drobné peridioly ( plodničky obsahující spory), přichycené na dně pohárku vláknem – funikulem => dopadem dešťové kapky je peridiola vymrštěna ven a funikulem se uchytí na trávě => čeká až ji sežere býložravec => teprve po průchodu jeho trávicím traktem spory klíčí.")
22
Speciální vymršťování má hrachovec
(Sphaerobolus): peridiola je uložena v "kalíšku", jehož vrstvy tvoří vnitřní a vnější okrovka – v buňkách jedné z vrstev vnější okrovky probíhá hydrolýza glykogenu na glukózu => vysoká koncentrace glukózy => způsobí osmotický tlak => pro jeho vyrovnání buňky prudce nasávají vodu => vrstvička nabobtná a vytlačí vzhůru vnitřní okrovku, která tak "vystřelí" peridiolu (až na vzdálenost 4 metrů!). Uvolňování spor napomáhá i když některé části plodnic hygroskopicky nabobtnají – např. pohyby vláken (elater) v okrovce Battarraea – nebo když dojde za sucha ke svinutí cípů vnější okrovky => vytlačení spor u hvězdáku (Astraeus). Sphaerobolus stellatus Kresba: /fungi/birds-nest-cannonball.html Foto: L. Hagara, V. Antonín, J. Baier: Houby. Aventinum, Praha, 1999. Battarraea stevenii Foto Pavol Kešeľák, /obrazok_detail.php?obrazok_id=102115 Foto Jiří Polčák, /obrazok_detail.php?obrazok_id=95937 Množství spor vyprodukovaných jednou plodnicí je různé od řádu miliónů u malých plodnic po 1015 (7 x 1015 spor v teřichu Langermannia gigantea, srovnatelné množství je 5,5 x 1015 z rouškaté plodnice Ganoderma lipsiense během 6 měsíců). Langermannia gigantea
: peridiola je uložena. v kalíšku , jehož vrstvy tvoří vnitřní. a vnější okrovka – v buňkách jedné. z vrstev vnější okrovky probíhá. hydrolýza glykogenu na glukózu. => vysoká koncentrace glukózy. => způsobí osmotický tlak => pro jeho vyrovnání buňky prudce nasávají vodu => vrstvička nabobtná a vytlačí vzhůru vnitřní okrovku, která tak vystřelí peridiolu (až na vzdálenost 4 metrů!). Uvolňování spor napomáhá i když některé části plodnic. hygroskopicky nabobtnají – např. pohyby vláken (elater) v okrovce Battarraea – nebo když dojde za sucha ke svinutí cípů vnější okrovky => vytlačení spor u hvězdáku (Astraeus). Sphaerobolus stellatus. Kresba: /fungi/birds-nest-cannonball.html. Foto: L. Hagara, V. Antonín, J. Baier: Houby. Aventinum, Praha, Battarraea stevenii. Foto Pavol Kešeľák, /obrazok_detail.php obrazok_id= Foto Jiří Polčák, /obrazok_detail.php obrazok_id= Množství spor vyprodukovaných jednou plodnicí je různé od řádu miliónů u malých plodnic po 1015 (7 x 1015 spor v teřichu Langermannia gigantea, srovnatelné množství je 5,5 x 1015 z rouškaté plodnice Ganoderma lipsiense během 6 měsíců). Langermannia gigantea.")
Podobné prezentace