Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Poruchy hospodaření s vodou a elektrolyty
Přednáška pro zubní lékaře

3
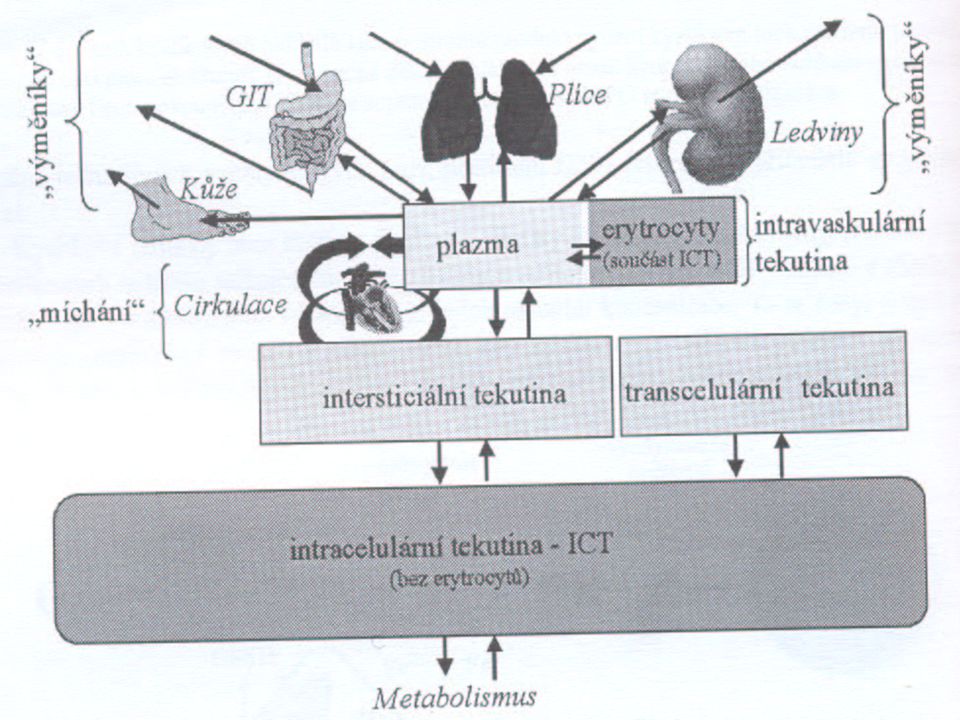
Tělesné kompartmenty tekutin
Voda je v organismu kompartmentalizovaná do několika oddílů. Intracelulární tekutina (ICF) zahrnuje 2/3 celkové vody. Primárně se jedná o roztok K+ a organických aniontů, proteinů etc. Regulace: buněčné membrány + buněčný metabolismus.
 zahrnuje 2/3 celkové vody. Primárně se jedná o roztok K+ a organických aniontů, proteinů etc. Regulace: buněčné membrány + buněčný metabolismus.")
4
Tělesné kompartmenty tekutin
Extracelulární tekutina (ECF) zahrnuje zbývající 1/3 tělesné vody. ECF je primárně roztok NaCl a NaHCO3. ECF se dále dělí na 3 subkompartmenty: Intersticiální tekutina (ISF) obklopuje buňky, ale necirkuluje. Zahrnuje asi 3/4 ECF. Plasma cirkuluje jako extracelulární komponenta krve. Je to 1/4 ECF. Transcelulární tekutina je tekutina mimo tyto kompartmenty (1-2 litry- cerebrospinální tekutina, trávicí šťávy, hlen etc.).
 zahrnuje zbývající 1/3 tělesné vody. ECF je primárně roztok NaCl a NaHCO3. ECF se dále dělí na 3 subkompartmenty: Intersticiální tekutina (ISF) obklopuje buňky, ale necirkuluje. Zahrnuje asi 3/4 ECF. Plasma cirkuluje jako extracelulární komponenta krve. Je to 1/4 ECF. Transcelulární tekutina je tekutina mimo tyto kompartmenty (1-2 litry- cerebrospinální tekutina, trávicí šťávy, hlen etc.).")
6
Speciální poznámky: Všechny kompartmenty jsou v osmotické rovnováze (s výjimkou přechodných změn) Ionty a malé molekuly roztoků, které tvoří ECF, jsou v rovnováze, při podobných koncentracích v každém subkompartmentu Objem ECF je proporcionální celkovému obsahu Na+

7
Iontové složení tělesných tekutin
Plasma obsahuje cca 7 volumových % proteinů a lipidů Aktivita iontů je limitovaná obsahem vody v roztoku Některé ionty se vážou na proteiny nebo jiné ionty

8
Elektrolyty Plasma, (mEq/L) [molarita] Plasmatická voda (mEq/L) [molalita] Intersticiální tekutina (mEq/L) Intracelulární tekutina (mEq/L) Kationty: Na+ 142 153 145 10 K+ 4 4.3 160 Ca++ 5 5.4 2 Mg++ 2.2 26 Kationty celkově: 165 156 198 Anionty“ Chloridy 101 108.5 114 3 Bikarbonáty 27 29 31 Fosfáty 100 Sulfáty 1 20 Organické kyseliny 6 6.5 7 Proteiny 16 17 65 Anionty celkově:
![Elektrolyty Plasma, (mEq/L) [molarita] Plasmatická voda (mEq/L) [molalita] Intersticiální tekutina (mEq/L)](http://slideplayer.cz/slide/5761837/18/images/8/Elektrolyty+Plasma%2C+%28mEq%2FL%29+%5Bmolarita%5D+Plasmatick%C3%A1+voda+%28mEq%2FL%29+%5Bmolalita%5D+Interstici%C3%A1ln%C3%AD+tekutina+%28mEq%2FL%29.jpg "Intracelulární tekutina (mEq/L) Kationty: Na K Ca Mg Kationty celkově: Anionty Chloridy Bikarbonáty Fosfáty Sulfáty Organické kyseliny Proteiny Anionty celkově:")
9
Výměna mezi intracelulárními a extracelulárními kompartmenty:
ICF a ECF jsou odděleny membránami buněk. Bílkovinné komponenty buněčných membrán zaručují podstatnou část permeability membrán pro vodu při pozorném řízení selektivní permeability pro ionty. Buněčné membrány jsou flexibilní. Jestliže voda teče dovnitř (ven) do (z) buněk, roztahují se (kontrahují). Hydrostatické tlaky proto nehrají signifikantní roli a osmóza vede spíše k tokům než ke změnám tlaku. Osmóza se objevuje, pokud vzniká gradient nepropustného solutu přes membránu permeabilní pro vodu.
 do (z) buněk, roztahují se (kontrahují). Hydrostatické tlaky proto nehrají signifikantní roli a osmóza vede spíše k tokům než ke změnám tlaku. Osmóza se objevuje, pokud vzniká gradient nepropustného solutu přes membránu permeabilní pro vodu.")
10
Výměna mezi intracelulárními a extracelulárními kompartmenty:
V buňkách se objevují osmotické toky, pokud vzniká osmotický gradient mezi intracelulárními a extracelulárnímu tekutinami V celém těle jsou tyto kompartmenty vždy v osmotické rovnováze přesto, že složení tekutin v těchto kompartmentech může být velmi odlišné Přidání nebo odebrání vody nebo solutů jednomu nebo několika tělesným kompartmentům povede k výměně vody mezi ICF a ECF, pokud došlo k narušení osmotické rovnováhy

11
Výměna tekutin mezi plasmou a intersticiální tekutinou
Vodné roztoky plasmy a intersticiální tekutiny se vyměňují přes stěny kapilár. Primární síly, které řídí tuto výměnu, jsou: * Hydrostatický tlak * Osmóza

12
Membránové transportní mechanismy
Póry v buněčné membráně dané strukturou transmembranózních bílkovin umožňují pasáž malých iontů (H+, K+, Na+) přes membránu pasivní difuzí přes tuto bílkovinu (= iontový kanál) Nebo transmembránový protein může investovat energii obyčejně z ATP do aktivace přesunu iontů přes buněčnou membránu. V tom případě se chová jako jako iontová pumpa
 přes membránu pasivní difuzí přes tuto bílkovinu (= iontový kanál) Nebo transmembránový protein může investovat energii obyčejně z ATP do aktivace přesunu iontů přes buněčnou membránu. V tom případě se chová jako jako iontová pumpa.")
13
Membránové transporty jsou řízeny silami,
které působí na úrovní membrány Na molekulu může působit více sil najednou (např. koncentrační a napěťový gradient) Pohyb vody je řízen obvykle jak tlakovým, tak osmotických gradientem Množství vody, které se přesunuje během osmózy je veliké, což vede ke změnám objemu buňky
 Pohyb vody je řízen obvykle jak tlakovým, tak osmotických gradientem. Množství vody, které se přesunuje během osmózy je veliké, což vede ke změnám objemu buňky.")
14
Transmembránový protein
Transmembránový protein je integrální mebránový protein. Hydrofobní doména zasahuje do lipidového vnitřku membrány, zatímco hydrofilní domény zasahují do vodního prostředí uvnitř nebo vně buňky nebo kompartmentu. N-terminální část bílkoviny je obvykle orientována extracelulárně a C-terminální část do cytoplasmatické oblasti. Mnohé transmembránové proteiny mají alfa helix, který mnohokrát prochází membránou, čímž bílkovinu kotví v membráně.

15
Typy transmembranózních proteinů
Dva základní typy: Prochází jen jednou Prochází mnohokrát a má k tomu mnohočetné topogenní sekvence

16
Integrální membránový protein transmembránového typu

17
Funkce integrálních membránových proteinů (IMP)
Transportér různých molekul Identifikace buňky pro rozpoznání jinými buňkami nebo okolím buňky iniciace intracelulární odpovědi na vnější molekuly Integrální membránové proteiny mohou být receptory kanály enzymy

18
Typy IMP Integrin Cadherin Insulin receptor
NCAM (neural cell adhesion molecule) Selectin Buněčné adhezivní proteiny Receptorové proteiny Glykoforin
 Selectin. Buněčné adhezivní proteiny. Receptorové proteiny. Glykoforin.")
19
Buněčné adhezivní molekuly
Jsou často transmembránovými receptory. Extracelulární doménou se často váže na jiný protein: Na povrchu sousedních buněk (adheze buňka-buňka) Na složku extracelulární matrix (adheze buňka-extracelulární matrix). Molekula, na kterou se váže adhezivní protein se nazývá ligand.
 Na složku extracelulární matrix (adheze buňka-extracelulární matrix). Molekula, na kterou se váže adhezivní protein se nazývá ligand.")
20
Transport makromolekul přes buněčné membrány
Se děje pomocí endocytózy: buněčná membrána invaginuje do buňky a tvoří vezikulu (endosom), který obsahuje extracelulární tekutina a další látky Umožňuje transport makromolekul, které se jinak nedostanou přes buněčnou membránu Některé endosomy jsou pokryty receptory, které selektivně vážou molekuly (LDL receptor) Makromolekuly jsou obvykle zlikvidovány lysozomy
, který obsahuje extracelulární tekutina a další látky. Umožňuje transport makromolekul, které se jinak nedostanou přes buněčnou membránu. Některé endosomy jsou pokryty receptory, které selektivně vážou molekuly (LDL receptor) Makromolekuly jsou obvykle zlikvidovány lysozomy.")
21
Transportní mechanismy
Pohyb tekutin je umožněn primárně působením tlaku Hydrostatický tlak způsobuje pohyb tekutin a látek v nich rozpuštěných přes póry v plasmatické membráně Tento pohyb je úměrný tlakovému gradientu, ploše a propustnosti této bariéry

22
Membránové transporty jsou řízeny silami,
které působí na úrovní membrány Typ transportu Rozdíly v Síla Difúze koncentraci Koncentrační gradient Elektrický proud napětí (voltage) “Voltage“ gradient Objem (objemový tok) tlaku Tlakový gradient Osmóza (objemový tok) osmotickém tlaku Osmotický gradient
 Voltage gradient. Objem (objemový tok) tlaku. Tlakový gradient. Osmóza (objemový tok) osmotickém tlaku. Osmotický gradient.")
23
Difúze Náhodný pohyb molekul z místa s vyšší koncentrací do místa s nižší koncentrací Difúze různých látek spolu neinterferují Látky mohou překročit membrány difúzí, pouze pokud jsou hydrofobní nebo přestupují přes tight junctions Difúze vody podle koncentračního gradientu se nazývá osmóza

24
Difúze Prostá difúze přes membrány se označuje jako permeabilita
Nevyžaduje speciální dodávku ATP Je úměrná koncentračnímu gradientu Nedochází k saturaci Tok = (konstanta permeability) x (koncentrační rozdíl) Konstanta permeability je poměr difúzní konstanty a tloušťky membrány; rychle pronikající látky mají vysokou permeabilitní konstantu (hydrofobní látky mají vyšší permeabilitu přes lipidovou dvojvrstvu membrán) Tok (flux) se děje vždy z místa vyšší koncentrace do místa s nižší koncentrací
 x (koncentrační rozdíl) Konstanta permeability je poměr difúzní konstanty a tloušťky membrány; rychle pronikající látky mají vysokou permeabilitní konstantu (hydrofobní látky mají vyšší permeabilitu přes lipidovou dvojvrstvu membrán) Tok (flux) se děje vždy z místa vyšší koncentrace do místa s nižší koncentrací.")
25
Usnadněná difúze Proteiny fungují jako nosiče nebo póry, které dovolují průnik látek, které nemohou projít přímo, bez pomoci. Tento pohyb je ještě pasivní, z místa vyšší koncentrace do místy nižší koncentrace difundující látky Funguje jen přes buněčné membrány. Příbuzné látky mohou soutěžit o stejného nosiče nebo póry. Maximální transport je charakterizován jako Tm (transportní maximum).
.")
26
Usnadněná difúze Některé transportní proteiny tvoří kanály s vrátky ("gates„); vrátka jsou normálně uzavřena a otevírají se na elektrické nebo chemické stimuly Některé trasnportéry přenášejí více než jeden typ molekuly (spřažený transport)
; vrátka jsou normálně uzavřena a otevírají se na elektrické nebo chemické stimuly. Některé trasnportéry přenášejí více než jeden typ molekuly (spřažený transport)")
27
Usnadněná difúze Usnadněná difúze využívá membránových proteinových kanálů k tomu, aby molekuly s nábojem (které jinak nemohou proniknout přes membránu) volně difundovaly dovnitř a ven z buňky. Časté jsou zejména kanály pro transport K+, Na+ a Cl- Rychlost usnadněné difúze je limitována počtem dostupných molekul kanálů na rozdíl od difúze, jejíž rychlost je závislá pouze na koncentračním gradientu
 volně difundovaly dovnitř a ven z buňky. Časté jsou zejména kanály pro transport K+, Na+ a Cl- Rychlost usnadněné difúze je limitována počtem dostupných molekul kanálů na rozdíl od difúze, jejíž rychlost je závislá pouze na koncentračním gradientu.")
28
Rozdíly v transportní rychlosti mezi pasivním transportem, usnadněnou difúzí a aktivním transportem

29
Osmóza: Osmóza je difúze vody podél jejího koncentračního gradientu
Čistá voda má molekulární váhu 18 g/M, takže její koncentrace je asi 55 M!

30
Osmóza Roztoky zabírají místo, které by za jiných okolnosti vyplňovala voda v roztoku, a jsou často doprovázeny molekulami vody, což snižuje jejich aktivitu (efektivní koncentrace) Buněčné membrány jsou pro vodu propustné, ale nepropouštějí všechny nebo žádné rozpuštěné látky Za těchto podmínek znamená gradient této látky také gradient v koncentraci volné vody
 Buněčné membrány jsou pro vodu propustné, ale nepropouštějí všechny nebo žádné rozpuštěné látky. Za těchto podmínek znamená gradient této látky také gradient v koncentraci volné vody.")
31
Osmolalita versus osmolarita
Osmolalita vyjadřuje osmotický tlak v jednom kilogramu rozpouštědla a je přímo úměrná počtu rozpuštěných částic v roztoku dané hmotnosti (vyjadřuje se tedy v jednotkách mmol/kg) Osmolarita se vztahuje k objemu roztoku (a vyjadřuje se v jednotkách mmol/l)
 Osmolarita se vztahuje k objemu roztoku (a vyjadřuje se v jednotkách mmol/l)")
32
Hrubý odhad osmolarity séra
Osmolarita séra = 2 x (konc. Na+ + konc. K+) + 5 Osmolarita séra = 2 x (konc. Na+ + konc. glu + konc. urey)
 + 5. Osmolarita séra = 2 x (konc. Na+ + konc. glu + konc. urey)")
33
Osmolarita versus tonicita
Osmolarita měří efektivní gradient pro vodu za předpokladu, že osmoticky účinné látky nikam neprostupují. Je to jednoduše počet rozpuštěných částic. Proto 300 mM roztok glukózy, 300 mM roztok urey a 150 mM roztok NaCl mají stejnou osmolaritu Buňka se v těchto roztocích chová různě: Ve 150 mM roztoku NaCl budou stejné osmotické síly na obou stranách buněčné mebrány (NaCl neprochází přes buněčnou membránu) a buňka si podrží svůj objem Urea velmi dobře prochází přes buněčné membrány. Proto buňka v 300 mM roztoku urey rychle oteče, protože urea i voda rychle do buňky vstoupí
 a buňka si podrží svůj objem. Urea velmi dobře prochází přes buněčné membrány. Proto buňka v 300 mM roztoku urey rychle oteče, protože urea i voda rychle do buňky vstoupí.")
34
Osmolarita versus tonicita
Tonicita je funkční termín, který popisuje tendenci roztoku odolat expanzi extracelulárního objemu Dva roztoky jsou izoosmotické, pokud obsahují stejný počet rozpuštěných částic bez ohledu na to, jak mnoho vody proteče přes danou membránu Dva roztoky jsou izotonické, pokud nedojde k žádnému pohybu vody přes membránovou bariéru bez ohledu na to, jak mnoho částic je v nich rozpuštěných 150 mM roztok NaCl bude izoosmotický a zároveň i izotonický- buňka v něm nebude otékat ani se nebude svrašťovat. V izoosmotickém roztoku 300 mM urey dojde k otoku buňky až k jejímu prasknutí, protože tento roztok urey se bude chovat jako hypotonický

35
Fyziologie membránového transportu
Typy transportu 1

36
2

37
Leaky a tight epitely a sekreční a resorbční funkce
20

38
„Leaky“ a „tight“ epitely
Rozdělení podle proporce para- a transcelulárních toků Leaky epitely: podstatné objemy transportu vody a solutů, většina resorbčních epitelů. 19

39
Rozložení typů transportu ve vztahu k membránám
Aktivní transport se uskutečňuje pouze přes buněčné membrány (pumpy a symporty) Paracelulárně se realizují pouze pasivní transporty (cestou tight junctions v epitelech) Protože v leaky epitelech je paracelulární tok snadno dosažitelný, může docházet snadno k paralyzaci aktivních toků. V tomto případě není možno dosáhnout vysokého koncentračního gradientu (částice se mohou paracelárně vracet) ani vysokého elektrického gradientu
 Paracelulárně se realizují pouze pasivní transporty (cestou tight junctions v epitelech) Protože v leaky epitelech je paracelulární tok snadno dosažitelný, může docházet snadno k paralyzaci aktivních toků. V tomto případě není možno dosáhnout vysokého koncentračního gradientu (částice se mohou paracelárně vracet) ani vysokého elektrického gradientu.")
40
Aktivní transport Aktivní transport jako jediný umožňuje přesun po i proti koncentračnímu gradientu Je limitován počtem přítomných molekul transportérů Primární-membránový protein sám spotřebovává energii (obvykle získanou hydrolýzou ATP) ke konformační změně, která umožňuje transport molekuly přes tento protein ( Na+-K+ pumpu). Sekundární
 ke konformační změně, která umožňuje transport molekuly přes tento protein ( Na+-K+ pumpu). Sekundární.")
41
Primární aktivní transport

42
Na/K pumpa Nalezena ve všech typech buněk
U člověka zahrnuje 30% bazálního metabolismu Pumpuje 3 ionty Na+ z buňky a 2 K+ ionty do buňky 3 Na+ se vážou na místa uvnitř buňky ATP se po vazbě hydrolyzuje na ADP a uvolňuje fosfát, který se váže na pumpu Pumpa změní konformaci a exponuje místa na povrchu buňky 3 Na+ opouštějí buňku a 2 K+ se vážou na různá místa v buňce Fosfát se uvolňuje z pumpy Dojde opět ke změně konformace, což vede k expozici vazných míst uvnitř buňky 2K+ se uvolní dovnitř buňky

43
Sekundární aktivní transport

44
Iontové kanály dosud popsáno více než 100 typů kanálů Funkce:
Elektrická excitabilita svalových buněk Elektrická signalizace v nervovém systému (jednotlivý neuron obsahuje 10 a více iontových kanálů, umístěných v různých doménách plasmatické membrány). Přítomné také v buňkách rostlin a v mikroorganismech.
. Přítomné také v buňkách rostlin a v mikroorganismech.")
45
Iontové kanály Od prostých pórů pro vodu je odlišují tyto vlastnosti:
Iontová selektivita Póry musí být dostatečně úzké v místě působení sil tak, aby prošly pouze ionty vybrané velikosti a náboje. Má se za to, že pronikající ionty se musí zbavit většiny doprovázející vody, aby prošly přes nejužší část kanálu: to limituje rychlost jejich průchodu

46
Iontové kanály Iontové kanály nejsou stále otevřené, nýbrž jsou vrátkované Otevírají se na specifické podněty, zejména na změnu napětí na membráně (voltage-gated channels), mechanický stres (mechanically gated channels) vazbu ligandu (ligand-gated channels). Ligandem může být extracelulární mediátor, např. neurotransmiter (transmitter-gated channels) intracelulární mediátor, např. iont (ion-gated channels), nebo nukleotid (nucleotide-gated channels) Aktivita mnohých iontových kanálů je dále modulována jejich fosforylací nebo defosforylací
, mechanický stres (mechanically gated channels) vazbu ligandu (ligand-gated channels). Ligandem může být. extracelulární mediátor, např. neurotransmiter (transmitter-gated channels) intracelulární mediátor, např. iont (ion-gated channels), nebo nukleotid (nucleotide-gated channels) Aktivita mnohých iontových kanálů je dále modulována jejich fosforylací nebo defosforylací.")
47
Typický iontový kanál, který mění konformaci.
Transmembránový proteinový komplex vytváří v lipidové dvojvrstvě hydrofilní póry jen tehdy, pokud jsou vrátka otevřena. Polární postranní řetezce aminokyselin vytvářejí stěnu póru, hydrofobní postranní řetězce vstupují do interakce s lipidovou dvojvrstvou.

48
Regulace volumu a tonicity
Voda: asi l,5 l se vyměňuje denně nezbytně (z toho moče musí být min. 0,5 l), zbytek obratu je fakultativní. Reguluje se příjmem (žízní) a vylučováním moče (adiuretinem). Tonicita se reguluje vodou, cirkulující objem v poslední instanci sodíkem
, zbytek obratu je fakultativní. Reguluje se příjmem (žízní) a vylučováním moče (adiuretinem). Tonicita se reguluje vodou, cirkulující objem v poslední instanci sodíkem.")
49
Poruchy bilance Poruchy distribuce

50
Příjem vody Složka potravy Produkt metabolizmu
Pití, regulačně významná cesta příjmu vody - žízeň

51
Výdej vody Kůží (perspiratio insensibilis, pot)
Dýchací systém (perspiratio insensibilis) Stolice Moč, regulačně významná cesta výdeje vody - ADH
 Stolice. Moč, regulačně významná cesta výdeje vody - ADH.")
52
Denní bilance vody

53
Regulace volumu a tonicity
Regulace tonicity – osmoreceptory v hypothalamu → zadní lalok hypofýzy, žízeň → ADH → resorpce vody v ledvině Regulace volumu – baroreceptory, uplatňují se za extrémních podmínek, odpověď pomalejší než z osmoreceptorů Objem překoná tonicitu

54
ADH

55
Stimulace sekrece ADH Vzestup osmolarity plazmy (zmenšení objemu buněk v osmoreceptorech) Pokles efektivního cirkulačního objemu Zvýšená hladina Ang II CNS: stres, bolest, strach, sexuální vzrušení dopamin, nikotin, hypoxie, hyperglykémie, některé léky Tlumení sekrece ADH Hypervolémie, hypoosmolarita, ADH (zpětnovazebně) Enkefaliny, glukokortikoidy, alkohol
 Enkefaliny, glukokortikoidy, alkohol.")
56
Místem působení ADH Aquaporiny řízený transport vody v renálním sběrném kanálku. Stimulace receptoru 2 pro vasopresin způsobuje inzerci aquaporinu (prostřednictvím cAMP) do apikální membrány, což umožní transport vody podél osmotického gradientu
 do. apikální membrány, což umožní transport vody podél osmotického gradientu.")
57
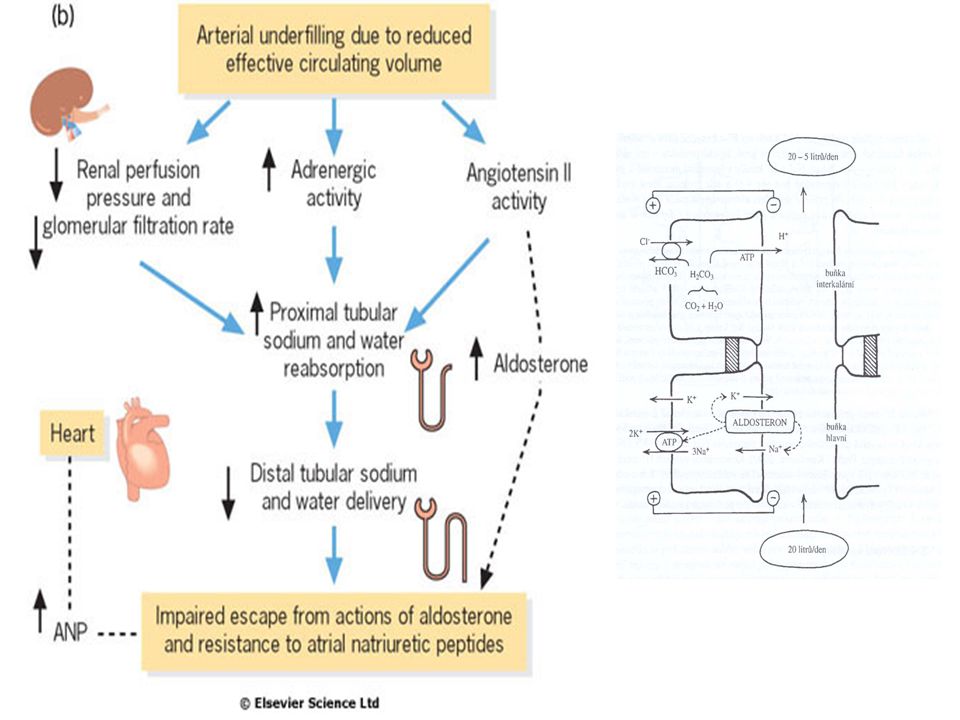
Stimulace RAS Snížená perfúze ledvin (baroreceptory)
Snížení dodávky NaCl (macula densa)
")
59
Regulace volumu a tonicity
5

60
Vliv renálních patofyziologických stavů na volum a osmolalitu
H2O Akutní nemoci glomerulů Retence Stenóza art. renalis, velmi vysoký TK, tlaková diuréza Exkrece Prerenální azotémie účelné k úpravě tlaku nebo volumu

61
Vliv renálních patofyziologických stavů na volum a osmolalitu
H2O Akutní selhání ledvin, inic. fáze Retence ASL-fáze restituce (polyurická)- ledvina ztrácející sůl Exkrece Chronické renální selhání (až do pokročilého stupně) GFR 10-20mL/min Bez poruch Tubulointersticiální nemoci, adrenální insuficience, diuretika, některé nefropatie
- ledvina ztrácející sůl. Exkrece. Chronické renální selhání. (až do pokročilého stupně) GFR 10-20mL/min. Bez poruch. Tubulointersticiální nemoci, adrenální insuficience, diuretika, některé nefropatie.")
62
Edematózní stavy * s výjimkou primární renální retence 17

63
Struktura kapilárního systému
arterie se větví 6-8 krát, terminální větvě průměr pod 20 μm – arterioly arterioly se větví 2-5 krát do průměru 5-9 μm – kapiláry venuly vény

64
Uspořádání mikrocirkulace

65
Kapilární stěna 0,5 μm silná
jedna vrstva buněk, vnější obal tvoří bazální membrána

66
Funkční anatomie CNS plíce kůže kosterní sval střevo exokrinní žlázy
ledvinné glomeruly chorioidální plexus játra slezina kostní dřeň Kontinuální Fenestrované Diskontinuální

67
Průtok krve kapilárou nebývá kontinuální, v závislosti na prekapilárním sfinkteru krev teče nebo neteče (změny v sekundách až minutách) hlavním regulačním faktorem množství kyslíku v tkáních hovoří-li se o průtoku, tlaku, atd. má se tedy na mysli průměrná hodnota všech kapilár v dané tkáni

68
Transendoteliální transport
je ovlivněn: chemickým gradientem, gradientem hydrostatického a onkotického tlaku permeabilitou kapilár silové gradienty přes membránu hydrostatika: DP a Dp netgradienty onkotického a hydrostatického tlaku chemicky: DC koncentrační gradient glucose CO2 Pc Pif pc pif

69
Síly určující pohyb přes kapilární membránu
kapilární tlak: působí směrem ven z kapiláry tlak intersticiální tekutiny: dovnitř nebo ven, podle toho je-li pozitivní nebo negativní koloidně-osmotický tlak plazmy: dovnitř koloidně-osmotický tlak IST: ven

70
1. Kapilární tlak (Pc) 30-40 mm Hg na arteriálním konci
10-15 mm na venózním konci uprostřed kolem 25 mm Hg

71
2. Tlak IST (Pif) úzké spojení s lymfatickým systémem
většinou se pohybuje kolem –3 mm Hg

72
3. Koloidně osmotický tlak plazmy (Πp)
28 mm Hg 19 mm Hg způsobeno proteiny 9 mm Hg kationty, které drží v plazmě díky proteinům (Donnanův efekt)
")
73
4. Koloidně osmotický tlak IST (Πif)
v intersticiu je asi 30 g/l proteinů (40% hodnot plazmy) jejich onkotický tlak činí asi 8 mm Hg
 jejich onkotický tlak činí asi 8 mm Hg.")
74
Arterielní konec kapiláry
síly směřující ven z kapiláry: kapilární tlak tlak IST onkotický tlak IST 8,0 41 síly směřující do kapiláry: onkotický tlak plazmy celkem 41-28=13 mmHg směrem ven z kapiláry (0,5 % plazmy)
")
75
Venózní konec kapiláry
síly směřující do kapiláry: onkotický tlak plazmy 28 síly směřující ven z kapiláry: kapilární tlak 10 tlak IST onkotický tlak IST 8,0 21 celkem 28-21=7 mmHg směrem do kapiláry (0,5 % plazmy)
")
76
Funkční důsledek tlak nasávající tekutinu na venózním konci je výrazně nižší ale: venózní konec má vyšší permeabilitu a proto: 90 % tekutiny, která vyteče na arteriolárním konci se na venózním vrátí

77
Starlingova rovnováha
průměrný kapilární tlak Pc je výpočtem 17,3 mm Hg síly působící ven jsou pak Pc + Pif + Πif = 17, = 28,3 mm Hg směrem do kapiláry působí jen Πp = 28 celkem tedy ven 0,3 mm Hg (2 ml/min, dorovnání lymfatickým systémem)
")
78
Starlingovy síly

79
Porušení rovnováhy vzestup Pc o 20 mmHg (ze 17) zvýší filtrační tlak na 20,3 mmHg, tj. 68x zvýšené množstí tekutiny v ISP nestačí lymfatický systém pojmout edémy naopak, při poklesu Pc objem IST klesá

80
Patogeneze edémů

81
Edémy při zvýšení hydrostatického tlaku

82
Edémy při snížení onkotického tlaku

83
Edémy při zánětu

84
Objemová a osmotická bilance
Poruchy objemu a osmolarity jsou v klinických podmínkách úzce spojeny Bilance mezi příjmem a výdejem vody určí, zda se normální objem (normovolémie, euvolémie) - zvětší (hypervolémie) - sníží (hypovolémie) Relativní poměr mezi příjmem či ztrátami solutů na jedné straně a vody na straně druhé určí hodnotu osmolarity - při větším příjmu vody než solutů nebo po větších ztrátách solutů než vody se isoosmolarita změní na hypoosmolaritu - při větší retenci solutů než vody nebo po větších ztrátách vody než solutů se vyvine hyperosmolarita
 - zvětší (hypervolémie) - sníží (hypovolémie) Relativní poměr mezi příjmem či ztrátami solutů na jedné straně a vody na straně druhé určí hodnotu osmolarity. - při větším příjmu vody než solutů nebo po větších ztrátách. solutů než vody se isoosmolarita změní na hypoosmolaritu. - při větší retenci solutů než vody nebo po větších ztrátách. vody než solutů se vyvine hyperosmolarita.")
85
Kombinace poruch volumu a tonicity v extracelulárním prostoru (9 teoreticky možných kombinací)
6

86
Poruchy tonicity poruchy vody: stavy 1, 4, 6, 9
Poruchy volumu poruchy sodíku: stavy 2, 3, 8, 7

87
Hypervolemické stavy Kapacita ledvin pro vylučování vody je natolik velká, že ani extrémní zátěž vodou při zdravých ledvinách nevede k retenci tekutin v extracelulárním prostoru Ledviny dokážou vyloučit i velká kvanta sodíku (kapacita však může být překonána v extrémních případech) Retence vody může být způsobena: Přesunem tekutiny z intravazálního prostoru do intersticia a následnou signalizací sníženého efektivního objemu 2) Retence sodíku a vody ledvinami (primární nemoci ledvin nebo zvýšené hladiny faktorů regulujících vylučování sodíku a vody v ledvinách)
 Retence vody může být způsobena: Přesunem tekutiny z intravazálního prostoru do intersticia a následnou signalizací sníženého efektivního objemu. 2) Retence sodíku a vody ledvinami (primární nemoci ledvin nebo zvýšené hladiny faktorů regulujících vylučování sodíku a vody v ledvinách)")
88
Přesun tekutiny z intravazálního prostoru do intersticia

89
STAV 1 hyperhydratace hypoosmolální
Tělo dostává (zadržuje) převážně vodu Příčiny: infuze glukózových roztoků, nefrotický sy cirhóza psychogenní polydipsie renální oligo/anurie při zvýš. tubul. rezorpci vody, SIADH, chlorpropamid selhání srdce renální oligo/anurie sníž. GFR, zvl. je-li podána voda nebo glukózový roztok
 převážně vodu. Příčiny: infuze glukózových roztoků, nefrotický sy. cirhóza. psychogenní polydipsie. renální oligo/anurie při zvýš. tubul. rezorpci vody, SIADH, chlorpropamid. selhání srdce. renální oligo/anurie sníž. GFR, zvl. je-li podána voda. nebo glukózový roztok.")
90
STAV 2 hyperhydratace izoosmolální
Tělo dostává (zadržuje) vodu izoosmoticky Příčiny: iv. infuze izoosmotických tekutin nefrotický syndrom cirhóza selhání srdce nesteroidní antiflogistika selhávající ledvina (GFR) akutně i chronicky, zvl. jsou- li podávány izoosmotické roztoky STAV 2 hyperhydratace izoosmolální
 vodu izoosmoticky. Příčiny: iv. infuze izoosmotických tekutin. nefrotický syndrom. cirhóza. selhání srdce. nesteroidní antiflogistika. selhávající ledvina (GFR) akutně i chronicky, zvl. jsou- li podávány izoosmotické roztoky. STAV 2 hyperhydratace izoosmolální.")
92
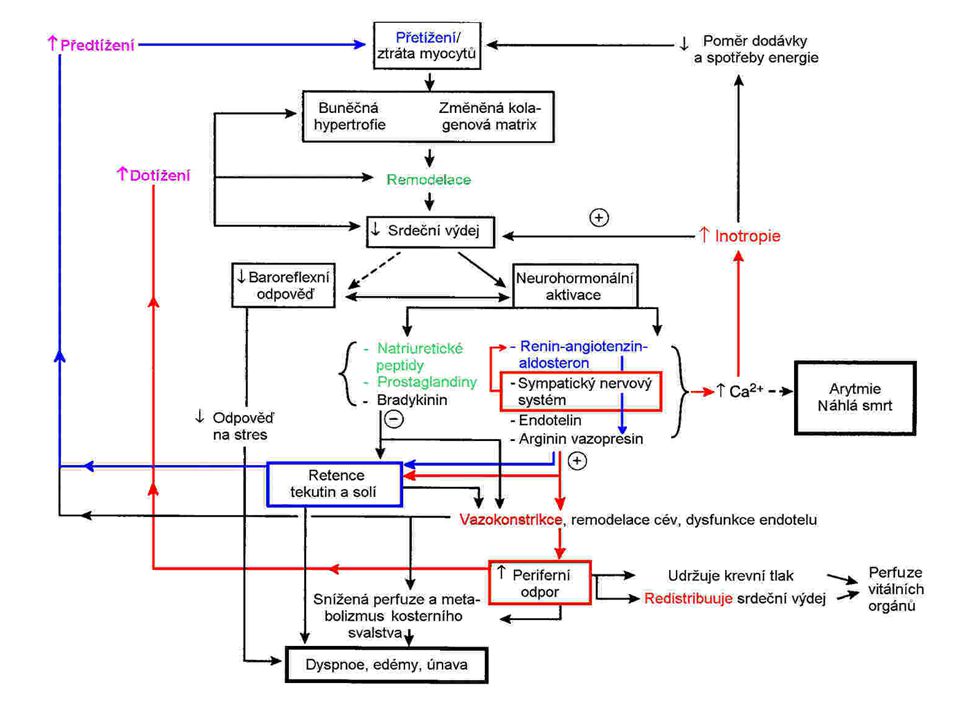
Důsledky hypervolémie
Hypervolémie zvýšené předtížení levé komory zvýšený srdeční výdej srdeční výdej * nezměněný perif. odpor = = arteriální tlak arteriální tlak hydrostatický kapilární tlak filtrace do IC prostoru edém

93
STAV 3 hyperhydratace hyperosmolální
Tělo dostává (zadržuje) převážně Na+ Příčiny: masivní příjem Na+ (per os, mořská voda, i.v.) primární nadbytek mineralokortikoidů akutní nemoci glomerulů oboustranná parench. onem. ledvin s chronickou ledv. nedostatečností (GFR < 10 mL/min)
 převážně Na+ Příčiny: masivní příjem Na+ (per os, mořská voda, i.v.) primární nadbytek mineralokortikoidů. akutní nemoci glomerulů. oboustranná parench. onem. ledvin s chronickou ledv. nedostatečností (GFR < 10 mL/min)")
94
Hypovolemické stavy Příčinou je negativní bilance vody
Ta je ale prakticky vždy spojena s bilancí sodíku - Snížení objemu vody v extracelulární tekutině je zpravidla spojeno i s poklesem celkového množství sodíku

95
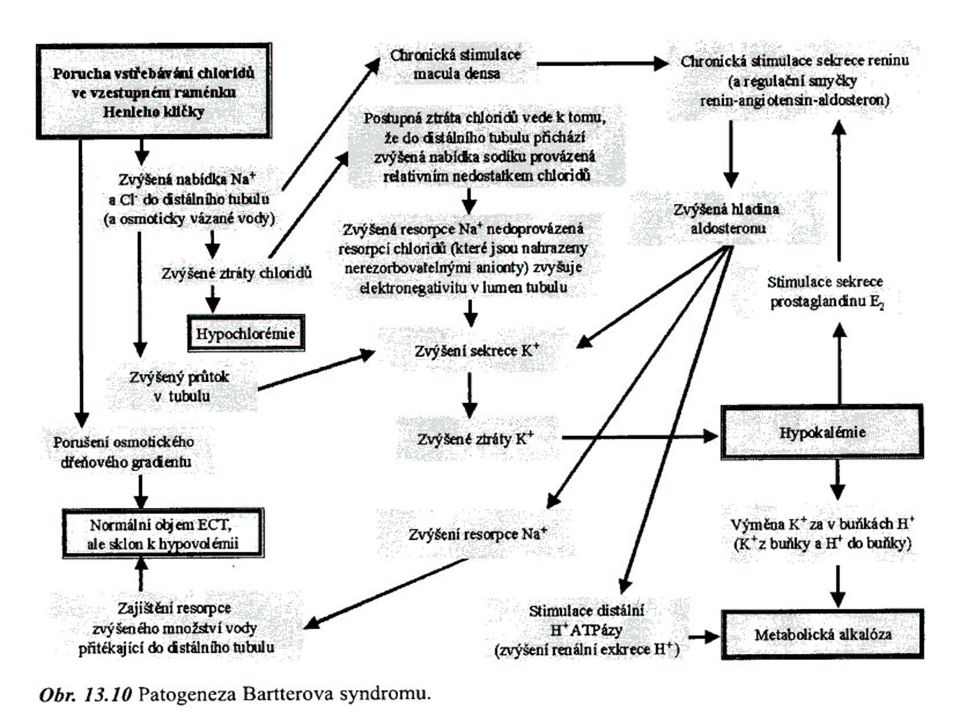
STAV 7 dehydratace hypoosmolální
Tělo ztrácí převážně Na+ Příčiny: aliment. nedost. soli v kombinaci se ztrátami primární nedostatek mineralokortikoidů renální ztráty soli: polyurie při akutním sehání ledvin ztráta hypotonických tekutin osmotická diuréza tlaková diuréza u extrémně TK BARTTERŮV sy abusus diuretik

97
STAV 8 dehydratace izoosmolální
Tělo propouští vodu izoosmoticky Příčiny: ztráta krve nebo plazmy, popáleniny punkce ascitu těžký průjem (jinak hyperosmolární dehydratace), žlučový drén, píštěle únik do intersticia nebo 3. prostoru rozdrcení tkání střevní obstrukce pankreatitis
, žlučový drén, píštěle. únik do intersticia nebo 3. prostoru. rozdrcení tkání. střevní obstrukce. pankreatitis.")
98
Tělo nedostává (propouští) převážně vodu
STAV 9 dehydratace hyperosmolální Tělo nedostává (propouští) převážně vodu Příčiny: zvracení průjem pocení insenzibilní ztráty hyperventilace horečka, horko hyperglykémie u diabetes mellitus mannitol
 převážně vodu. Příčiny: zvracení. průjem. pocení. insenzibilní ztráty. hyperventilace. horečka, horko. hyperglykémie u diabetes mellitus. mannitol.")
99
STAV 9 dehydratace hyperosmolální
snížená žízeň novorozenc bezvědomí diabetes insipidus (centrální) osmot. diuréza u diabetes mellitus diabetes insipidus (nefrogenní) polyurie při akutním selhání ledvin
 osmot. diuréza u diabetes mellitus. diabetes insipidus (nefrogenní) polyurie při akutním selhání ledvin.")
100
Přehled poruch volumu a tonicity včetně příčin
8

101
Vysvětlivky k obrázku:
a – přehnaná kompenzace hyperosmolality (stav 9) vodou b – kompromis pomocí ADH: hypervolemie nestoupá při značném vzestupu NaEC tak výrazně, aby se udržela izoosmolalita c – pokles efektivního krevního volumu d – tři faktory retence Na (GFR, aldosteron, 3. faktor) e – pomocí ADH f – nesteroidní antiflogistika (acetylosalicylová kyselina, salicylát sodný, fenacetin, paracetamol) tlumí ochranné prostaglandiny v ledvině pokles GFR g – SIADH (Inappropriate secretion of antidiuretic hormone) je klinicky euvolemický, subklinicky hypervolemický h – pomocí žízně a ADH, předpokládá se ovšem i jistá ztráta soli
 vodou. b – kompromis pomocí ADH: hypervolemie nestoupá při značném vzestupu NaEC tak výrazně, aby se udržela izoosmolalita. c – pokles efektivního krevního volumu. d – tři faktory retence Na (GFR, aldosteron, 3. faktor) e – pomocí ADH. f – nesteroidní antiflogistika (acetylosalicylová kyselina, salicylát. sodný, fenacetin, paracetamol) tlumí ochranné prostaglandiny v ledvině pokles GFR. g – SIADH (Inappropriate secretion of antidiuretic hormone) je klinicky euvolemický, subklinicky hypervolemický. h – pomocí žízně a ADH, předpokládá se ovšem i jistá ztráta soli.")
102
Vysvětlivky k obrázku i – ačkoliv může být dehydratace těla při ztrátě hypotonických tekutin značná, pokles cirkulujího volumu bývá při ní zanedbatelný (čistá ztráta vody hrazena z 90% nikoliv z cirkulujícího objemu) j – je-li ztráta vody o dost vyšší než ztráta soli, může být snížení NaEC provázeno zvýšením PNa k – organizmus masivně ztratil sůl i vodu, rychlou zpětnou vazbou přes žízeň a ADH se však v této extrémní situaci snaží zachovat spíš objem, což se mu zdaří jen zčásti, a ještě za cenu hypotonie (opět kompromis); ztráty soli jsou zde hrazeny pouze pitím l – Na v moči < 10 mmol/L m – Na v moči > 20 mmol/L – příčinou ztráty Na je moč sama n – při malém objemu moče Na v moči > 600 mmol/L
 j – je-li ztráta vody o dost vyšší než ztráta soli, může být snížení. NaEC provázeno zvýšením PNa. k – organizmus masivně ztratil sůl i vodu, rychlou zpětnou vazbou přes žízeň a ADH se však v této extrémní situaci snaží zachovat spíš objem, což se mu zdaří jen zčásti, a ještě za cenu hypotonie (opět kompromis); ztráty soli jsou zde hrazeny pouze pitím. l – Na v moči < 10 mmol/L. m – Na v moči > 20 mmol/L – příčinou ztráty Na je moč sama. n – při malém objemu moče Na v moči > 600 mmol/L.")
Podobné prezentace









INFUZI 1 LITRU IZOTONICKÉ TEKUTINY b)POŽITÍ 2 LITRŮ VODY.>")










