Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
第十章 脂类代谢 脂类概述 脂肪的分解代谢 脂肪的生物合成

2
一、脂类概述 1. 概念 脂类是脂肪和类脂的总称,它是有脂肪酸与醇作用生成的酯及其衍生物,统称为脂质或脂类,是动物和植物体的重要组成成分。脂类是广泛存在与自然界的一大类物质,它们的化学组成、结构理化性质以及生物功能存在着很大的差异,但它们都有一个共同的特性,即可用非极性有机溶剂从细胞和组织中提取出来。

3
2. 分类 脂肪 真脂或中性脂肪(甘油三酯) 类脂 卵磷脂 脑磷脂 甘油磷脂 鞘氨醇磷脂 磷脂 糖脂 异戊二烯酯 甾醇 萜类
 类脂 卵磷脂 脑磷脂 甘油磷脂 鞘氨醇磷脂 磷脂 糖脂 异戊二烯酯 甾醇 萜类")
4
3. 脂类的功能 1g 脂肪在体内彻底氧化供能约38KJ,而1g 糖彻底氧化仅供销能 16.7KJ。
3. 脂类的功能 ① 贮藏物质/能量物质: 脂肪是机体内代谢燃料的贮 存形式,它在体内氧化可释 放大量能量以供机体利用。 ② 提供给机体必需脂成分: (1)必需脂肪酸 亚油酸 18碳脂肪酸,含两个不饱和键; 亚麻酸 18碳脂肪酸,含三个不饱和键; 花生四烯酸 20碳脂肪酸,含四个不饱和键; (2)生物活性物质 激素、胆固醇、维生素等。 1g 脂肪在体内彻底氧化供能约38KJ,而1g 糖彻底氧化仅供销能 16.7KJ。
必需脂肪酸. 亚油酸 18碳脂肪酸,含两个不饱和键; 亚麻酸 18碳脂肪酸,含三个不饱和键; 花生四烯酸 20碳脂肪酸,含四个不饱和键; (2)生物活性物质. 激素、胆固醇、维生素等。 1g 脂肪在体内彻底氧化供能约38KJ,而1g 糖彻底氧化仅供销能 16.7KJ。")
5
③生物体结构物质 (1)作为细胞膜的主要成分: 几乎细胞所含的磷 脂都集中在生物膜中,是生物膜 结构的基本组成成分。 (2)保护作用: 脂肪组织较为柔软,存在于各重 要的器官组织之间,使器官之间 减少摩擦,对器官起保护作用。 ④用作药物:卵磷脂、脑磷脂可用于肝病、神经衰 弱及动脉粥样硬化的治疗等。

6
二、脂肪的分解代谢

7
1.脂肪的水解 酯酶为限速酶! 脂肪动员:储存在脂肪细胞中的脂肪,被脂肪酶逐步水解为游离的脂肪酸及甘油,并释入血以供其他组织氧化利用的过程。

8
2.甘油的分解 磷酸酯酶

9
3. 脂肪酸的氧化分解(β-氧化) ① 脂肪酸的活化——脂酰CoA的生成 长链脂肪酸氧化前必须进行活化,活化在线粒体外进行。内质网和线粒体外膜上的脂酰CoA合成酶在ATP、CoASH、Mg2+存在条件下,催化脂肪酸活化,生成脂酰CoA。

10
②脂酰CoA的穿膜(脂酰CoA进入线粒体)
酯酰肉碱穿梭系统 肉毒碱脂酰CoA转移酶Ⅰ( 限速酶 ) 肉毒碱脂酰CoA转移酶Ⅱ 移位酶
 肉毒碱脂酰CoA转移酶Ⅱ. 移位酶.")
11
③ 脂肪酸的β氧化 长链脂酰CoA的β氧化是在线粒体脂肪酸氧化酶系作用下进行的,每次氧化断去二碳单位的乙酰CoA,再经TCA循环完全氧化成二氧化碳和水,并释放大量能量。偶数碳原子的脂肪酸β氧化最终全部生成乙酰CoA。 脂酰CoA的β氧化反应过程如下:

12
(1)脱氢 脂酰CoA经脂酰CoA脱氢酶催化,在其α和β碳原子上脱氢,生成△2反烯脂酰CoA,该脱氢反应的辅基为FAD。
(2)加水(水合反应) △2反烯脂酰CoA在△2反烯脂酰CoA水合酶催化下,在双键上加水生成L-β-羟脂酰CoA。
加水(水合反应) △2反烯脂酰CoA在△2反烯脂酰CoA水合酶催化下,在双键上加水生成L-β-羟脂酰CoA。")
13
(3)脱氢 L-β-羟脂酰CoA在L-β-羟脂酰CoA脱氢酶催化下,脱去β碳原子与羟基上的氢原子生成β-酮脂酰CoA,该反应的辅酶为NAD+。
(4)硫解 在β-酮脂酰CoA硫解酶催化下,β-酮脂酰CoA与CoA作用,硫解产生 1分子乙酰CoA和比原来少两个碳原子的脂酰CoA。
硫解 在β-酮脂酰CoA硫解酶催化下,β-酮脂酰CoA与CoA作用,硫解产生 1分子乙酰CoA和比原来少两个碳原子的脂酰CoA。")
14
氧化的生化历程 TCA + 乙酰CoA 乙酰CoA 乙酰CoA 乙酰CoA 乙酰CoA 乙酰CoA 乙酰CoA 乙酰CoA H20 H2O
RCH2CH2CO-SCoA FAD FADH2 脂酰CoA 脱氢酶 H20 呼吸链 RCH=CH-CO-SCoA 氧化的生化历程 H2O β-烯脂酰CoA 水化酶 RCHOHCH2CO~ScoA NAD + NADH β-羟脂酰CoA 脱氢酶 H20 呼吸链 RCOCH2CO-SCoA β-酮酯酰CoA 硫解酶 CoASH 脂酰CoA R-CO~ScoA + CH3CO~SCoA 乙酰CoA 乙酰CoA 乙酰CoA 乙酰CoA TCA 乙酰CoA 乙酰CoA ATP 乙酰CoA 乙酰CoA

15
脂肪酸β-氧化作用小结: (1)脂肪酸β-氧化时仅需活化一次,消耗1个ATP的 两个高能键,生成脂酰CoA 。
经肉碱运到线粒内;中、短链脂肪酸直接进入线 粒体,由线粒体内的脂酰CoA合成酶活化。 (3)β-氧化包括脱氢、水化、脱氢、硫解4个重复步骤。
β-氧化包括脱氢、水化、脱氢、硫解4个重复步骤。")
16
(4) 脂肪酸β氧化最终的产物为乙酰CoA、NADH和FADH2。假如碳原子数为Cn的脂肪酸进行β氧化,则需要作(n/2-1)次循环才能完全分解为n/2个乙酰CoA,产生n/2-1个NADH和n/2-1个FADH2;生成的乙酰CoA通过TCA循环彻底氧化成二氧化碳和水并释放能量,而NADH和FADH2则通过呼吸链传递电子生成ATP。至此可以生成的ATP数量为: 以软脂酸(16C)为例计算其完全氧化所生成的ATP分子数:
为例计算其完全氧化所生成的ATP分子数:")
17
4. 脂肪酸的其它氧化分解方式 1)奇数碳原子脂肪酸的分解 丙酰CoA有两条代谢途径: ① 羧化:丙酰CoA转化成琥珀酰CoA,进入TCA。
4. 脂肪酸的其它氧化分解方式 1)奇数碳原子脂肪酸的分解 丙酰CoA有两条代谢途径: ① 羧化:丙酰CoA转化成琥珀酰CoA,进入TCA。 动物体内存在这条途径,因此,在动物肝脏中奇数碳脂肪酸最终能够异生为糖。 ② 脱羧:丙酰CoA转化成乙酰CoA,进入TCA。 植物、微生物中较普遍。
奇数碳原子脂肪酸的分解. 丙酰CoA有两条代谢途径: ① 羧化:丙酰CoA转化成琥珀酰CoA,进入TCA。 动物体内存在这条途径,因此,在动物肝脏中奇数碳脂肪酸最终能够异生为糖。 ② 脱羧:丙酰CoA转化成乙酰CoA,进入TCA。 植物、微生物中较普遍。")
18
2)脂肪酸的α-氧化(不需活化,直接氧化游离脂
肪酸) RCH2COOH→RCOOH+CO2 对于降解支链脂肪酸、奇数碳脂肪酸、过分长链脂肪酸(如脑中C22、C24)有重要作用。 3)脂肪酸的-ω氧化(ω端的甲基羟基化,氧化成 醛,再氧化成酸) 少数的12C以下的脂酸可通过ω-氧化途径,产生二羧酸。
 RCH2COOH→RCOOH+CO2. 对于降解支链脂肪酸、奇数碳脂肪酸、过分长链脂肪酸(如脑中C22、C24)有重要作用。 3)脂肪酸的-ω氧化(ω端的甲基羟基化,氧化成. 醛,再氧化成酸) 少数的12C以下的脂酸可通过ω-氧化途径,产生二羧酸。")
19
△3顺—△2反烯脂酰CoA异构酶(改变双键位置和顺反构型)
4)不饱和脂肪酸的分解 ① 单不饱和脂肪酸的氧化 油酸的β氧化: △3顺—△2反烯脂酰CoA异构酶(改变双键位置和顺反构型) ② 多不饱和脂肪酸的氧化 亚油酸的β氧化: △3顺—△2反烯脂酰CoA异构酶(改变双键位置和顺反构型) β-羟脂酰CoA差向酶(改变β-羟基构型:D→L型)
不饱和脂肪酸的分解. ① 单不饱和脂肪酸的氧化. 油酸的β氧化: △3顺—△2反烯脂酰CoA异构酶(改变双键位置和顺反构型) ② 多不饱和脂肪酸的氧化. 亚油酸的β氧化: △3顺—△2反烯脂酰CoA异构酶(改变双键位置和顺反构型) β-羟脂酰CoA差向酶(改变β-羟基构型:D→L型)")
20
5. 乙酰CoA的去路 ①进入TCA循环最终氧化生成二氧化碳和水以及大量的ATP。 ②生成酮体参与代谢(动物体内)
脂肪酸β氧化产生的乙酰CoA,在肌肉细胞中可进入TCA循环进行彻底氧化分解;但在肝脏及肾脏细胞中还有另外一条去路,即形成乙酰乙酸、D-β-羟丁酸和丙酮,这三者统称为酮体。

21
6. 酮体的代谢 (1)酮体的生成(肝细胞的线粒体内)
肝脏线粒体中的乙酰CoA走哪一条途径(生成酮体或进入TCA ),主要取决于草酰乙酸的可利用性。 饥饿状态下,草酰乙酸离开TCA,用于异生合成葡萄糖。只有少量乙酰CoA进入TCA,大多数乙酰CoA用于合成酮体。 酮体的生成过程:
,主要取决于草酰乙酸的可利用性。 饥饿状态下,草酰乙酸离开TCA,用于异生合成葡萄糖。只有少量乙酰CoA进入TCA,大多数乙酰CoA用于合成酮体。 酮体的生成过程:")
22
图 酮体的生成-1 ① 2分子的乙酰CoA在肝脏线粒体β-酮酯酰硫解酶的作用下,缩合成乙酰乙酰CoA,并释放1分子的CoASH。

23
图 酮体的生成-2 ②乙酰乙酰CoA与另一分子乙酰CoA缩合成羟甲基戊二酰CoA(HMG CoA),并释放1分子CoASH。
,并释放1分子CoASH。")
24
图 酮体的生成-3 ③ HMG CoA在HMG CoA裂解酶催化下裂解生成乙酰乙酸和乙酰CoA。

25
④ 乙酰乙酸在线粒体内膜β-羟丁酸脱氢酶作用下,被还原成β-羟丁酸。
图 酮体的生成-4 ④ 乙酰乙酸在线粒体内膜β-羟丁酸脱氢酶作用下,被还原成β-羟丁酸。

26
图 酮体的生成-5 ⑤ 部分乙酰乙酸可自动脱羧而成为丙酮。

27
酮体的生成途径 CoA-SH CH3COCH2CO~SCoA 乙酰乙酰CoA CoA-SH HMG-CoA 合酶 CH3CO~SCoA
硫解酶 CH3CO~SCoA 乙酰CoA CH3—C—CH2CO~SCoA OH CH2COOH β-羟β-甲基戊二酸单酰CoA CH3CO~SCoA 乙酰CoA HMG-CoA 裂解酶 关键酶 乙酰乙酸 脱羧酶 NADH+H+ CH3COCH2COOH 乙酰乙酸 NAD+ CH3—C—CH2COOH OH β-羟丁酸 CH3COCH3 丙酮 CO2 β -羟丁酸 脱氢酶

28
(2)酮体的分解(肝外) 肝脏是生成酮体的器官,但不能使酮体进一步氧化分解,而是采用酮体的形式将乙酰CoA经血液运送到肝外组织,作为它们的能源,尤其是肾、心肌、脑等组织中主要以酮体为燃料分子。在这些细胞中,酮体进一步分解成乙酰CoA参加三羧酸循环。
酮体的分解(肝外) 肝脏是生成酮体的器官,但不能使酮体进一步氧化分解,而是采用酮体的形式将乙酰CoA经血液运送到肝外组织,作为它们的能源,尤其是肾、心肌、脑等组织中主要以酮体为燃料分子。在这些细胞中,酮体进一步分解成乙酰CoA参加三羧酸循环。")
29
A. 乙酰乙酸在肌肉线粒体中经β-酮脂酰CoA转移酶催化,能被琥珀酰CoA活化成乙酰乙酰CoA。
B. 乙酰乙酰CoA被β氧化酶系中的硫解酶裂解成乙酰CoA进入三羧酸循环。 C. β-羟丁酸在β-羟丁酸脱氢酶作用下,脱氢生成乙酰乙酸,然后再转变成乙酰CoA而被氧化。 D. 丙酮可在一系列酶作用下转变成丙酮酸或乳酸,进而异生成糖。

30
酮体的氧化过程 心、肾、脑细胞 心、肾、脑、骨骼肌细胞

31
(3)酮体生成的生理意义 酮体是肝输出能量的一种形式,为肝外组织提供可利用的能源。
酮体溶于水,分子小,能通过血脑屏障及肌肉毛细管壁。脑组织细胞不能氧化脂肪酸,但能利用酮体。长期饥饿,糖供应不足时,酮体可以代替葡萄糖,成为脑组织及肌肉的重要能源。

32
饥饿、糖供给不足或糖尿病: 脂肪动员加强,酮体生成增加,超过肝外组织氧化能力,“酮血症”、“酮尿症”。可引起酸中毒。

33
三、脂肪的生物合成 1.脂肪酸的生物合成 生物机体内脂类的合成是十分活跃的,特别是在高等动物的肝脏、脂肪组织和乳腺中占优势。脂肪酸合成的碳源主要来自糖酵解产生的乙酰CoA。脂肪酸合成步骤与氧化降解步骤完全不同。脂肪酸的生物合成是在细胞液中进行,需要CO2和柠檬酸参加;而氧化降解是在线粒体中进行的。

35
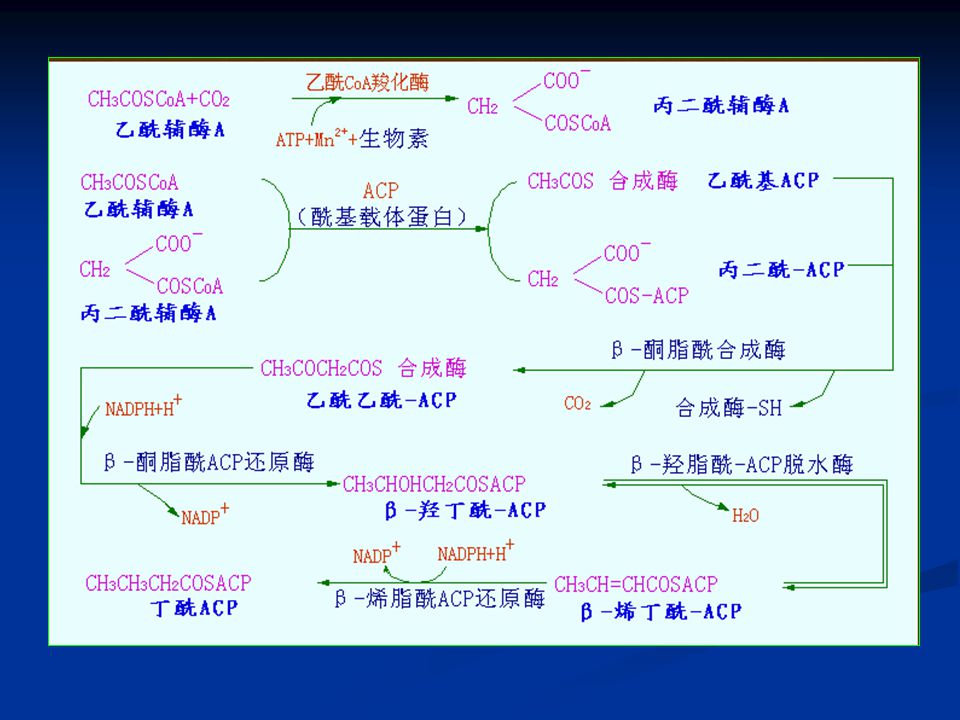
合成过程可以分为三个阶段: (1)原料的准备——乙酰CoA羧化生成丙二酸单酰CoA(在细胞液中进行),由乙酰CoA羧化酶催化,辅基为生物素,是一个不可逆反应。 乙酰CoA羧化酶可分成三个不同的亚基: 生物素羧化酶(BC) 生物素羧基载体蛋白(BCCP) 羧基转移酶(CT)
 生物素羧基载体蛋白(BCCP) 羧基转移酶(CT)")
36
乙酰CoA的穿膜转运: 柠檬酸穿梭系统 肉毒碱转运

37
(2)合成阶段 ——— 以软脂酸(16碳)的合成为例(在细胞液中进行)。催化该合成反应的是一个多酶体系,共有七种蛋白质参与反应,以没有酶活性的脂酰基载体蛋白(ACP)为中心,组成一簇。
原初反应(初始反应) 原初反应 缩合反应 还原反应 脱水反应
 原初反应. 缩合反应. 还原反应. 脱水反应.")
38
至此,生成的丁酰-ACP比开始的乙酰-ACP多了两个碳原子;然后丁酰基再从ACP上转移到β-酮脂酰合成酶的-SH上,再重复以上的缩合、还原、脱水、还原4步反应,每次重复增加两个碳原子,释放一分子CO2,消耗两分子NADPH,经过7次重复后合成软脂酰-ACP,最后经硫脂酶催化脱去ACP生成软脂酸(16碳)。
。")
39
(3)延长阶段(在线粒体和微粒体中进行)生物体内有两种不同的酶系可以催化碳链的延长,一是线粒体中的延长酶系,另一个是粗糙内质网中的延长酶系。
线粒体脂肪酸延长酶系 以乙酰CoA为C2供体,不需要酰基载体,由软脂酰CoA与乙酰CoA直接缩合。 内质网脂肪酸延长酶系 用丙二酸单酰CoA作为C2的供体,NADPH作为H的供体,中间过程和脂肪酸合成酶系的催化过程相同。

40
2. 不饱和脂肪酸的合成 不饱和脂肪酸中的不饱和键由去饱和酶催化形成。人体内含有的不饱和脂肪酸主要有棕榈油酸(16C,一个不饱和键)、油酸(18C,一个不饱和键)、亚油酸(18C,两个不饱和键)、亚麻酸(18C,三个不饱和键)以及花生四烯酸(20C,四个不饱和键)等,前两种单不饱和脂肪酸可由人体自己合成,后三种为多不饱和脂肪酸,必须从食物中摄取,因为哺乳动物体内没有△9以上的去饱和酶。
、油酸(18C,一个不饱和键)、亚油酸(18C,两个不饱和键)、亚麻酸(18C,三个不饱和键)以及花生四烯酸(20C,四个不饱和键)等,前两种单不饱和脂肪酸可由人体自己合成,后三种为多不饱和脂肪酸,必须从食物中摄取,因为哺乳动物体内没有△9以上的去饱和酶。")
41
本章重点: 脂类概述 脂肪的分解 脂肪的合成 脂肪与类脂,功能 脂肪酸的β -氧化,酮体 乙酰CoA羧化生成丙二酸单酰CoA
脂肪酸的从头合成

Podobné prezentace

 2012 1-3.>")










 2012 1-5 捷克語(一) 2012 1-5. 第五課 第一單元 課文【】 第一單元 課文【購物 -1 Nakupování-1 】 第二單元 文法與句型 第二單元 文法與句型 【數字 Číslovky základní 】 【數字 Číslovky základní 】 【主格複數.>")

 jap. Róši 老子>")



>")
