Stáhnout prezentaci
Prezentace se nahrává, počkejte prosím

1
Fotosyntéza Jiří Šantrůček Přírodovědecká fakulta Jihočeské univerzity
Katedra fyziologie rostlin Ústav molekulární biologie AV ČR České Budějovice Jiří Šantrůček

2
Máte na to !? 1/ Jaký je význam slova „foto-syntéza“ ? 2/
Jaký má fotosyntéza význam pro život na Zemi ? 3/ Dokážete pojmenovat vstupy a výstupy ? Odkud pochází kyslík? 4/ Kde k fotosyntéze dochází ? Pochod, kterým jsou autotrofní rostliny, řasy a někteří prokaryonti schopni energii slunce svázat do chemických vazeb se nazývá fotosyntéza (syntéza světlem). Pro naprostou většinu života na Zemi je prvotní energií zářivá energie slunce. Fotosyntéza je primárně zodpovědná za existenci veškeré organické hmoty, fosilních paliv a za podporu heterotrofního života na Zemi. Kromě toho jako vedlejší produkt „může“ fotosyntéza za přítomnost kyslíku v atmosféře nutného pro respirační aktivitu organismů. Fotosyntéza zahrnuje komplex reakcí a dějů provázejících zachycení sluneční energie, štěpení kyslíku, transport elektronů a produkci ATP a NADPH – to vše na membránách uvnitř chloroplastů (thylakoidech). Dále pak enzymové procesy fixace CO2 z atmosféry a jeho primární přeměny v cukry – ve stromatu (polotekuté fázi) choroplastů. Chloroplasty jsou zelené buněčné organely v buňkách listové tkáně nazývané mezofyl.
. Pro naprostou většinu života na Zemi je prvotní energií zářivá energie slunce. Fotosyntéza je primárně zodpovědná za existenci veškeré organické hmoty, fosilních paliv a za podporu heterotrofního života na Zemi. Kromě toho jako vedlejší produkt „může fotosyntéza za přítomnost kyslíku v atmosféře nutného pro respirační aktivitu organismů. Fotosyntéza zahrnuje komplex reakcí a dějů provázejících zachycení sluneční energie, štěpení kyslíku, transport elektronů a produkci ATP a NADPH – to vše na membránách uvnitř chloroplastů (thylakoidech). Dále pak enzymové procesy fixace CO2 z atmosféry a jeho primární přeměny v cukry – ve stromatu (polotekuté fázi) choroplastů. Chloroplasty jsou zelené buněčné organely v buňkách listové tkáně nazývané mezofyl.")
3
cukry O2 foton CO2 ATP, NADPH H2O
Fotosyntéza – zjednodušená časoprostorová škála O2 foton ( nm) CO2 ATP, NADPH Fotosyntéza zahrnuje komplex reakcí a dějů provázejících zachycení sluneční energie (fotonu), štěpení vody na kyslík a protony, transport elektronů a produkci ATP a NADPH – to vše na membránách uvnitř chloroplastů (thylakoidech). Dále pak enzymové procesy fixace CO2 z atmosféry a jeho primární přeměny v cukry – ve stromatu (polotekuté fázi) choroplastů. Chloroplasty jsou zelené buněčné organely v buňkách listové tkáně nazývané mezofyl. H2O Passioura, Plants in Action
 CO2. ATP, NADPH. Fotosyntéza zahrnuje komplex reakcí a dějů provázejících zachycení sluneční energie (fotonu), štěpení vody na kyslík a protony, transport elektronů a produkci ATP a NADPH – to vše na membránách uvnitř chloroplastů (thylakoidech). Dále pak enzymové procesy fixace CO2 z atmosféry a jeho primární přeměny v cukry – ve stromatu (polotekuté fázi) choroplastů. Chloroplasty jsou zelené buněčné organely v buňkách listové tkáně nazývané mezofyl. H2O. Passioura, Plants in Action.")
4
Obsah Přehled fotosyntézy:
Bilance: Má dáti, dal. Místo činu: Chloroplast (anatomie) Co se v chloroplastech děje? A jak se na to přišlo? Thylakoidní membrána, primární procesy Enzymatické pochody ve stromatu (bude příště) Primární pochody a struktury ve fotosyntéze: Absorpce světelného kvanta - pigmenty Separace náboje Transport elektronů Syntéza ATP, NADPH Ochrana fotosyntetického aparátu a regulace pochodů při fotosyntéze
 Co se v chloroplastech děje A jak se na to přišlo Thylakoidní membrána, primární procesy. Enzymatické pochody ve stromatu (bude příště) Primární pochody a struktury ve fotosyntéze: Absorpce světelného kvanta - pigmenty. Separace náboje. Transport elektronů. Syntéza ATP, NADPH. Ochrana fotosyntetického aparátu a regulace pochodů při fotosyntéze.")
5
Má dáti – dal: souhrnná reakce
CO2 + 2H2A (CH2O) + 2A + H2O h (1) Donor elektronů foto.obecně: CO2 + 2H2O (CH2O) + O2 + H2O h (2) foto. oxygenní: vyšší i nižšší rostliny, řasy sinice CO2 + 2H2S (CH2O) + 2S + H2O h (3) foto. anoxygenní: fotosyntetické bakterie
 + 2A + H2O. h (1) Donor elektronů. foto.obecně: CO2 + 2H2O (CH2O) + O2 + H2O. h (2) foto. oxygenní: vyšší i nižšší rostliny, řasy. sinice. CO2 + 2H2S (CH2O) + 2S + H2O. h (3) foto. anoxygenní: fotosyntetické bakterie.")
6
Místo činu

7
Chloroplast = fotosyntetická organela eukaryot
vnější membrána vnitřní membrána Chloroplast = fotosyntetická organela eukaryot Am = škrobová zrna Th = thylakoid (stromatální) G = grana Pg = plastoglobuly
 G = grana. Pg = plastoglobuly.")
8
Chloroplast - fotosyntetická organela eukaryot

9
Eukaryontní buňka: chloroplast
A = škrobové zrno, G = grana Pg = plastoglobuly Co se děje v chloroplastech? A jak se na to přišlo? Robert Hill (30-tá léta, akceptory elektronů, na světle měřil produkci O2 z vody) Daniel Arnon (50-tá léta, izolované chloroplasty váží CO2 a vytvářejí z něj cukry) Melvin Calvin & …… (50-60 l., sled enzym. reakcí) Co se děje v chloroplastech? A jak se na to přišlo? Začátky: Joseph Priestley - produkce kyslíku (1771), Jan Ingenhousz - světlo (1779). Koncem 19. Století se už znala bilanční rovnice fotosyntézy. To, že fixace CO2 a produkce kyslíku nejsou nutně spojeny ukázal už ve 30-tých letech Robert Hill, který dodával různé i anorganické akceptory elektronů a na světle měřil produkci O2 z vody (tzv. Hillova reakce). Tak se dodnes měří aktivita komplexů na thylakoidních membránách, které se účastní zachycení světelné energie a štěpení vody. Mezi léty Daniel Arnon ukázal, že izolované chloroplasty jsou sto vázat CO2 a vytvářet z něj cukry (použil izotopově značený 14CO2). Ukázalo se také, že celý tento proces se skládá ze dvou relativně nezávislých pochodů: 1) na světle se produkuje ATP (energie), NADPH (látka schopná redukce) a kyslík a 2) i bez světla pak může proběhnout fixace CO2. Brzy poté se zjistilo, že fixace CO2 probíhá cyklickou enzymatickou cestou nazvanou po objevitelích Calvinův nebo Calvin-Benson-Basshamův cyklus nebo také cyklus fotosyntetické redukce uhlíku (photosynthetic carbon reduction cycle, PCR). Objeveno Melvin Calvin-em, James Bassham-em a Andrew Bensonem na kalifornské univerzitě v Berkeley 6C → chloroplastový škrob U,K 101
 Daniel Arnon (50-tá léta, izolované chloroplasty váží CO2 a vytvářejí. z něj cukry) Melvin Calvin. & …… (50-60 l., sled enzym. reakcí) Co se děje v chloroplastech A jak se na to přišlo Začátky: Joseph Priestley - produkce kyslíku (1771), Jan Ingenhousz - světlo (1779). Koncem 19. Století se už znala bilanční rovnice fotosyntézy. To, že fixace CO2 a produkce kyslíku nejsou nutně spojeny ukázal už ve 30-tých letech Robert Hill, který dodával různé i anorganické akceptory elektronů a na světle měřil produkci O2 z vody (tzv. Hillova reakce). Tak se dodnes měří aktivita komplexů na thylakoidních membránách, které se účastní zachycení světelné energie a štěpení vody. Mezi léty Daniel Arnon ukázal, že izolované chloroplasty jsou sto vázat CO2 a vytvářet z něj cukry (použil izotopově značený 14CO2). Ukázalo se také, že celý tento proces se skládá ze dvou relativně nezávislých pochodů: 1) na světle se produkuje ATP (energie), NADPH (látka schopná redukce) a kyslík a 2) i bez světla pak může proběhnout fixace CO2. Brzy poté se zjistilo, že fixace CO2 probíhá cyklickou enzymatickou cestou nazvanou po objevitelích Calvinův nebo Calvin-Benson-Basshamův cyklus nebo také cyklus fotosyntetické redukce uhlíku (photosynthetic carbon reduction cycle, PCR). Objeveno Melvin Calvin-em, James Bassham-em a Andrew Bensonem na kalifornské univerzitě v Berkeley. 6C → chloroplastový škrob. U,K 101.")
10
Škrobový sluneční tisk list Geranium, negativ obrazu ...
D. Walker 57

11
Mechanismus primárních fotosyntetických procesů

12
pokus - dedukce - hypotéza - test
Musí existovat dva spřažené mechanismy přeměny světla v chloroplastu. Z čeho to vyplynulo ? pokus - dedukce hypotéza - test Experimentální efekt ukazující, že musí existovat dva spolupracující fotosystémy. Rychlost fotosyntézy při společné funkci obou (pík vpravo) je více než dvojnásobkem každého zvlášť.
 je více než dvojnásobkem každého zvlášť.")
13
Karikatura toku elektronů thylakidní membránou
Karikatura, která znázorňuje jak se vytváří rozdíl elektrochemického potenciálu na thylakoidní membráně chloroplastu. Dopad a absorpce fotonů (dodání energie z vnějšího prostředí) reakčními centry PS II a PS I a jejich anténami jsou znázorněny v podobě „skřítků s kladivem“, kteří vyrazí elektron na vyšší hladinu energie. Ten jde potom samospádem dolů (a přitom někde i roztáčí „kolo“ a žene tak další procesy jako transport protonů přes membránu) což je analogie membránových přenašečů elektronů a končí zachycen v koši – analogie vysoce-energetických redukovaných sloučenin (NADPH) využitelných při fixaci CO2 v Calvin-Bensonově cyklu ve stromatu thylakoidů.
 reakčními centry PS II a PS I a jejich anténami jsou znázorněny v podobě „skřítků s kladivem , kteří vyrazí elektron na vyšší hladinu energie. Ten jde potom samospádem dolů (a přitom někde i roztáčí „kolo a žene tak další procesy jako transport protonů přes membránu) což je analogie membránových přenašečů elektronů a končí zachycen v koši – analogie vysoce-energetických redukovaných sloučenin (NADPH) využitelných při fixaci CO2 v Calvin-Bensonově cyklu ve stromatu thylakoidů.")
14
schéma thylakoidní membrány s vybranými základními bílkovinnými komplexy
K oběma fázím fotosyntézy, dříve zvaným světelná a temnotní, dochází v různých částech chloroplastu. V membránách thylakoidů jsou komplexy bílkovin nazývané fotosystém dva (PS II) a fotosystém jedna (PS I) v jejichž reakčních centrech se mění energie světla (tj. fotonů zachytávaných molekulami chlorofylu na jiných membránových chlorofyl-bílkovinných komplexech, tzv. anténách nebo světlosběrných komplexech - light harvesting complex LHC a předávaných do reakčního centra PS II nebo PS I) na elektrochemickou energii separovaného náboje. PS II a PS I musí spolupracovat - jsou součástí řetězce transportu elektronů, který přesouvá elektrony z lumenu (vnitřku thylakoidů), kde se produkují při štěpení vody na části PS II zvané kyslík vyvíjející komplex, na vodorozpustnou redox- sloučeninu NADP+ vně thylakoidů. V lumenu chloroplastů se na světle hromadí protony (H+) z rozštěpené vody. Gradientu protonů mezi vnitřkem a vnějškem thylakoidu využívá v thylakoidní membráně umístěná ATP-áza k syntéze ATP. Tento bílkovinný v membráně umístěný komplex ATP-ázy se také nazývá „spřažovací faktor“ (angl. coupling factor nahoře v obrázku německy „Kopplungsfaktor“). Za zmínku stojí, že PS II a jeho LHC (LHC2) jsou umístěny v thylakoidních membránách nahloučených v granech, zatímco PS I a LHC I je thylakoidech mimo grana (viz obr.). U,K 100
 a fotosystém jedna (PS I) v jejichž reakčních centrech se mění energie světla (tj. fotonů zachytávaných molekulami chlorofylu na jiných membránových chlorofyl-bílkovinných komplexech, tzv. anténách nebo světlosběrných komplexech - light harvesting complex LHC a předávaných do reakčního centra PS II nebo PS I) na elektrochemickou energii separovaného náboje. PS II a PS I musí spolupracovat - jsou součástí řetězce transportu elektronů, který přesouvá elektrony z lumenu (vnitřku thylakoidů), kde se produkují při štěpení vody na části PS II zvané kyslík vyvíjející komplex, na vodorozpustnou redox- sloučeninu NADP+ vně thylakoidů. V lumenu chloroplastů se na světle hromadí protony (H+) z rozštěpené vody. Gradientu protonů mezi vnitřkem a vnějškem thylakoidu využívá v thylakoidní membráně umístěná ATP-áza k syntéze ATP. Tento bílkovinný v membráně umístěný komplex ATP-ázy se také nazývá „spřažovací faktor (angl. coupling factor nahoře v obrázku německy „Kopplungsfaktor ). Za zmínku stojí, že PS II a jeho LHC (LHC2) jsou umístěny v thylakoidních membránách nahloučených v granech, zatímco PS I a LHC I je thylakoidech mimo grana (viz obr.). U,K 100.")
15
Řetězce přesunu elektronů v thylakoidní membráně - Z schéma
Jiné (vědečtější než předchozí) schéma téhož – tzv. Z-schéma přenosu elektronu v thylakoidní membráně se škálami redox potenciálu a volné energie. P680 a P700 jsou reakční centra PS II a PS I napojená na své antény (světlosběrné komplexy). Označení podél dráhy elektronu (plastochinon, cytochrom b6f, plastocyanin, feredoxin (FD)) jsou názvy přenašečů elektronů v nebo na povrchu thylakoidní membrány.
 schéma téhož – tzv. Z-schéma přenosu elektronu v thylakoidní membráně se škálami redox potenciálu a volné energie. P680 a P700 jsou reakční centra PS II a PS I napojená na své antény (světlosběrné komplexy). Označení podél dráhy elektronu (plastochinon, cytochrom b6f, plastocyanin, feredoxin (FD)) jsou názvy přenašečů elektronů v nebo na povrchu thylakoidní membrány.")
16
Elektronový transport ve fotosyntetické membráně
při oxygenní fotosyntéze (Z-schéma) Buch s.595
 Buch s.595.")
17
Řetězce přesunu elektronů a protonů v thylakoidní membráně
Ještě realističtější schéma prostorového uspořádání chlorofyl-proteinových a proteinových komplexů v thylakoidní membráně chloroplastů včetně kyslík vyvíjejícího komplexu (ZY) v blízkosti P680 centra PS II a protonové ATP-ázy.
 v blízkosti P680 centra PS II a protonové ATP-ázy.")
18
Membránová organizace Z-schématu

19
Vstupy a výstupy látek a energie v primárních procesech
Souhrn Vstupy a výstupy látek a energie v primárních procesech oxygenní fotosyntézy

20
Pigmenty a antény

21
Zelenou barvu rostlin způsobuje chlorofyl
Zelenou barvu rostlin způsobuje chlorofyl. Téměř všechny fotosyntetické organismy jej/je obsahují. Rostliny, řasy a sinice (cyanobacteria) syntetizují světlo absorbující pigment = chlorofyl. Anaerobní fotosyntetické bakterie produkují variantu chlorofylu, které se říká bakteriochlorofyl. Molekula chlorofylu je složena z tetrapyrolového kruhu (porfyrinu), struktury kterou lze nalézt také v hemoglobinu a v cytochromech. Ale chlorofyl váže v centru tetrapyrolového kruhu hořčík (Mg), zatímco v hemu je atom železa. Navíc v chlorofylu je na jeden kruh vázán dlouhý (C20) hydrofóbní řetěz – tzv. fytol, který jej ukotvuje v membráně thylakoidu. Syntéza chlorofylu v rostlinách a hemu u živočichů mají společné prekurzory – glutamovou kyselinu, z které se syntetizuje kyselina d-aminolevulová (ALA). Předposlední krok syntézy chlorofylu – redukce protochlorofylidu na chlorofyl vyžaduje u většiny rostlinných druhů světlo. Výjimkou jsou nahosemené, řasy a fotosyntetické bakterie, které mohou syntetizovat chlorofyl i ve tmě. Poslední krok v syntéze chlorofylu je esterifikace fytolového ocasu. Chlorofyl b se syntetizuje z chlorofylu a oxygenázou, která převádí metylovou skupinu na druhém kruhu na formylovou.
 syntetizují světlo absorbující pigment = chlorofyl. Anaerobní fotosyntetické bakterie produkují variantu chlorofylu, které se říká bakteriochlorofyl. Molekula chlorofylu je složena z tetrapyrolového kruhu (porfyrinu), struktury kterou lze nalézt také v hemoglobinu a v cytochromech. Ale chlorofyl váže v centru tetrapyrolového kruhu hořčík (Mg), zatímco v hemu je atom železa. Navíc v chlorofylu je na jeden kruh vázán dlouhý (C20) hydrofóbní řetěz – tzv. fytol, který jej ukotvuje v membráně thylakoidu. Syntéza chlorofylu v rostlinách a hemu u živočichů mají společné prekurzory – glutamovou kyselinu, z které se syntetizuje kyselina d-aminolevulová (ALA). Předposlední krok syntézy chlorofylu – redukce protochlorofylidu na chlorofyl vyžaduje u většiny rostlinných druhů světlo. Výjimkou jsou nahosemené, řasy a fotosyntetické bakterie, které mohou syntetizovat chlorofyl i ve tmě. Poslední krok v syntéze chlorofylu je esterifikace fytolového ocasu. Chlorofyl b se syntetizuje z chlorofylu a oxygenázou, která převádí metylovou skupinu na druhém kruhu na formylovou.")
22
Rostlina je zelená díky chlorofylu. Ale proč je chlorofyl zelený?
Absorpce světla (fotonu) vede k přesunu molekuly do tzv. „excitovaného stavu“ Malé změny v substituentech na pyrolech způsobují významné změny v absorpci záření (světla) – viz absorpční spektra chorofylu a a b na horním obrázku. Chlorofyl je zelený proto, že absorbuje světlo vlnové délky 430 nm (modré) a 680 nm (červené) více než zelené (kolem 530 nm). Zelené světlo se tedy odráží a my ho tedy můžeme vidět více než červeného nebo modrého.
 vede k přesunu molekuly do tzv. „excitovaného stavu Malé změny v substituentech na pyrolech způsobují významné změny v absorpci záření (světla) – viz absorpční spektra chorofylu a a b na horním obrázku. Chlorofyl je zelený proto, že absorbuje světlo vlnové délky 430 nm (modré) a 680 nm (červené) více než zelené (kolem 530 nm). Zelené světlo se tedy odráží a my ho tedy můžeme vidět více než červeného nebo modrého.")
23
závislost fotosyntézy (produkce kyslíku, rychlosti fixace CO2)
Akční spektrum = závislost fotosyntézy (produkce kyslíku, rychlosti fixace CO2) na kvalitě světla. První akční spektra měřená Engelmannem koncem 19 století. Vláknitou řasu Spirogyra se spirálním chloroplastem a v suspensi s bakteriemi vyžadujícími kyslík a pohybujícími se za ním ozářil spektrem. U modré a červené je kumulace bakterií.
 na kvalitě světla. První akční spektra měřená Engelmannem koncem 19 století. Vláknitou řasu Spirogyra se spirálním chloroplastem a v suspensi s bakteriemi vyžadujícími kyslík a pohybujícími se za ním ozářil spektrem. U modré a červené je kumulace bakterií.")
24
Fluorescence chlorofylu je vždy v červené oblasti.
Modré světlo může excitovat chlorofyl do vyššího energetického stavu než červené světlo (Ee-Eg=hc/). Deexcitace se děje: teplem nebo fluorescencí nebo fotochemií neboli rezonančním transportem Fluorescence chlorofylu je vždy v červené oblasti. Co se děje s molekulou chlorofylu při absorpci světla? Existuje tzv. základní stav molekuly (ground state) a excitovaný stav chlorofylu ve dvou stupních. Ten s nižší energií se nazývá první excitovaný singletní stav (hladina) a ten s vyšší druhý excitovaný singletní stav (singletní znamená, že elektrony ve valenční slupce mají opačnou rotaci – spin). Excitované singletní stavy mají velmi krátkou dobu života (řádově pikosekundy tj s) , po jejichž uplynutí mají tendenci se vrátit do základního stavu. Foton červeného světla má nižší energii, která stačí jen na excitaci do prvního singletního stavu. Do druhého excitačního stavu může molekulu chlorofylu excitovat modré světlo. Návrat do základního stavu musí být provázen disipací (rozptýlením) přebytečné energie. V zásadě existují tři formy disipace: tepelná, zářivá tj. tzv. fluorescence nebo fotochemická – tj. rezonanční přenos energie (excitonu) na jinou sousední molekulu chlorofylu. Fotochemický přenos v rostlinách dominuje ale jsou situace, kdy vzrůstá fluorescence a teplotní disipace. Fotochemickému přenosu napomáhá uspořádání molekul chlorofylu do tzv. světlosběrných komplexů – antén. Energetický rozdíl excitovaného a základního stavu (Ee-Eg) se dá spočítat z Planckovy konstanty h (6,626 x J s), rychlosti světla c (3,0 x 108 m s-1) a vlnové délky světla l ( x 10-9 m).
. Deexcitace se děje: teplem nebo fluorescencí nebo fotochemií neboli rezonančním transportem. Fluorescence chlorofylu je vždy v červené oblasti. Co se děje s molekulou chlorofylu při absorpci světla Existuje tzv. základní stav molekuly (ground state) a excitovaný stav chlorofylu ve dvou stupních. Ten s nižší energií se nazývá první excitovaný singletní stav (hladina) a ten s vyšší druhý excitovaný singletní stav (singletní znamená, že elektrony ve valenční slupce mají opačnou rotaci – spin). Excitované singletní stavy mají velmi krátkou dobu života (řádově pikosekundy tj s) , po jejichž uplynutí mají tendenci se vrátit do základního stavu. Foton červeného světla má nižší energii, která stačí jen na excitaci do prvního singletního stavu. Do druhého excitačního stavu může molekulu chlorofylu excitovat modré světlo. Návrat do základního stavu musí být provázen disipací (rozptýlením) přebytečné energie. V zásadě existují tři formy disipace: tepelná, zářivá tj. tzv. fluorescence nebo fotochemická – tj. rezonanční přenos energie (excitonu) na jinou sousední molekulu chlorofylu. Fotochemický přenos v rostlinách dominuje ale jsou situace, kdy vzrůstá fluorescence a teplotní disipace. Fotochemickému přenosu napomáhá uspořádání molekul chlorofylu do tzv. světlosběrných komplexů – antén. Energetický rozdíl excitovaného a základního stavu (Ee-Eg) se dá spočítat z Planckovy konstanty h (6,626 x J s), rychlosti světla c (3,0 x 108 m s-1) a vlnové délky světla l ( x 10-9 m).")
25
Chl A Chl* A Chl+ A- Schéma fykobilisomu
PE=fykoeritrin, PC=plastocyanin, AP=allofykocyanin Model antény a reakčního centra Proč musí existovat antény? Představte si, že každý z vás zde sedících stavíte dům, každý si najmete si partu dělníků, jeřáb … Máte ale jen jedno auto na dovoz cihel a ještě takové, které uveze jen dvě cihly. Za takové situace bude každý z vás čekat třeba týden na další dvě cihly a přitom budete živit a platit partu stavařů a stroje kolem. V takové nějaké situaci by bylo reakční centrum PS pokud by neexistovaly antény. Chlorofylové antény rostlin (vpravo nahoře): Anténní chlorofyly (s vyšším podílem chl. b) sbírají energii absorbovaných fotonů a předávají ji na speciální pár molekul chlorofylů v reakčním centru fotosystému. Vysokoenergetický elektron pak odchází do elektron-transportního řetězce. Nechlorofylové antény (fykobilisomy) sinic a červených řas (vlevo dole) obsahují fykobilinové pigmenty kovalentně vázané na proteiny a tvoří tyčovité struktury. Světlo kratších vlnových délek se absorbuje na periferii fykobilisomů fykoerytrinem (PE) a energie se přenáší na PS II v thylakoidech vnitřními pigmenty, které také absorbují světlo o delší vlnové délce, fykocyaninem (PC) a alofykocyanninem (AP). Pro resonanční přenos energie v anténách je důležité těsné sousedství molekul chl. Účinnost přenosu se zmenšuje s 6-tou mocninou vzdálenosti molekul. Jde o přenos rezonancí většinou s účinnosti přes 99%. Ke každému reakčnímu centru je v anténě vázáno asi 250 molekul clorofylu. Chl A Chl* A Chl+ A- Alberts s.433
: Anténní chlorofyly (s vyšším podílem chl. b) sbírají energii absorbovaných fotonů a předávají ji na speciální pár molekul chlorofylů v reakčním centru fotosystému. Vysokoenergetický elektron pak odchází do elektron-transportního řetězce. Nechlorofylové antény (fykobilisomy) sinic a červených řas (vlevo dole) obsahují fykobilinové pigmenty kovalentně vázané na proteiny a tvoří tyčovité struktury. Světlo kratších vlnových délek se absorbuje na periferii fykobilisomů fykoerytrinem (PE) a energie se přenáší na PS II v thylakoidech vnitřními pigmenty, které také absorbují světlo o delší vlnové délce, fykocyaninem (PC) a alofykocyanninem (AP). Pro resonanční přenos energie v anténách je důležité těsné sousedství molekul chl. Účinnost přenosu se zmenšuje s 6-tou mocninou vzdálenosti molekul. Jde o přenos rezonancí většinou s účinnosti přes 99%. Ke každému reakčnímu centru je v anténě vázáno asi 250 molekul clorofylu. Chl A Chl* A Chl+ A- Alberts s.433.")
26
Reakční centrum PSII včetně kyslík vyvíjejícího komplexu (OEC)
")
27
Struktura reakčního centra PSII u oxygenních organismů
Struktura reakčního centra PSII u oxygenních organismů (vyšších rostlin, řas, sinic) Struktura reakčního centra PS II – schéma. V reakčním centru jsou dva hlavní proteinové podjednotky D1 a D2. Elektrony se přenášejí z P680 (speciální pár molekul chlorofylu) na pheofytin (Pheo) a pak na molekuly plastochinonů QA a QB. P680+ kation je redukován elektronem z vody, který přešel na Z (tj. tyrosin obsahující část proteinu D1). Spodní část ukazuje manganový klastr, který oxiduje (štěpí) vodu. CP43 a CP47 jsou proteiny, které váží chlorofyl a. D1 podjednotka je náchylná k poškození nadměrnou ozářeností (silným světlem) a proto má rychlý obrat syntézy a degradace.
 Struktura reakčního centra PS II – schéma. V reakčním centru jsou dva hlavní proteinové podjednotky D1 a D2. Elektrony se přenášejí z P680 (speciální pár molekul chlorofylu) na pheofytin (Pheo) a pak na molekuly plastochinonů QA a QB. P680+ kation je redukován elektronem z vody, který přešel na Z (tj. tyrosin obsahující část proteinu D1). Spodní část ukazuje manganový klastr, který oxiduje (štěpí) vodu. CP43 a CP47 jsou proteiny, které váží chlorofyl a. D1 podjednotka je náchylná k poškození nadměrnou ozářeností (silným světlem) a proto má rychlý obrat syntézy a degradace.")
28
Kinetika separace náboje
Primární akceptor a Kinetika separace náboje u bakterie Rhodopseudomonas viridis V roce 1980 zveřejnili Hatmut Michel, Johann Diesenhofer a Robert Huber stukturu komplexu reakčního centra fotosyntetické bakterie Rhodopseudomonas viridis s vysokým rozlišením zjištěnou pomocí rentgenove krystalografie. Ukázalo se, že je osově symetrická a má povahu diméru (všechny složky jsou zdvojeny ve dvou symentrických ramenech).Osa prochází specifickým párem bakteriochlorofylů P865 a jediným atomem železa. Kromě nich je v každé větvi monomer bakteriochlorofylu a, bakteriopheofytinu a a chinon jako primární akceptor elektronů (QA) a sekundární chinonový akceptor (QB). Kromě elektronových přenašečů nakreslených na obr. je v centru jedenáct transmembránových alfa helixů, podjednotka na vnější straně vážící cytochromy. V roce 1988 dostali všichni tři za svůj objev Nobelovu cenu. Přenos elektronů (separace náboje) je velmi rychlý proces – řádově jednotky pikosekund. Pomalejší je pak přechod z bakteriopheofytinu na primární akceptor (200 ps) a až o tři řády pomalejší je pak přechod z primárního na sekundární akceptor. Přenos elektronů v reakčním centru je nezávislý na teplotě a probíhá stejně rychle i při teplotě kapalného helia (4 oK) protože přenos nevyžaduje molekulární kolize.
.Osa prochází specifickým párem bakteriochlorofylů P865 a jediným atomem železa. Kromě nich je v každé větvi monomer bakteriochlorofylu a, bakteriopheofytinu a a chinon jako primární akceptor elektronů (QA) a sekundární chinonový akceptor (QB). Kromě elektronových přenašečů nakreslených na obr. je v centru jedenáct transmembránových alfa helixů, podjednotka na vnější straně vážící cytochromy. V roce 1988 dostali všichni tři za svůj objev Nobelovu cenu. Přenos elektronů (separace náboje) je velmi rychlý proces – řádově jednotky pikosekund. Pomalejší je pak přechod z bakteriopheofytinu na primární akceptor (200 ps) a až o tři řády pomalejší je pak přechod z primárního na sekundární akceptor. Přenos elektronů v reakčním centru je nezávislý na teplotě a probíhá stejně rychle i při teplotě kapalného helia (4 oK) protože přenos nevyžaduje molekulární kolize.")
29
Struktura PSII sinice Thermosynechococcus elongatus s 3,5 A rozlišením.
Žluté- D1,D2 anténní proteiny CP43(zelené) a CP 47(červené) …
 a CP 47(červené) …")
30
Oxidace vody 2H2O O2+4H+ + 4e-
Přenos energie na P680 u vyšších rostlin vede k oxidaci P680 a produkci silného oxidantu, který je sto oxidovat vodu a uvolnit přitom molekulární kyslík (O2). (redox potenciál přechodu P680/P680* musí být větší než +820 mV, protože to minimálně je třeba k rozštěpení vody). Oxidace vody ale není přímý proces. Vyžaduje kumulaci náboje. To bylo potvrzeno experimentálně pomocí krátkých jednofotonových záblesků. První dva záblesky nevyvolaly téměř žádnou produkci kyslíku (viz graf vlevo). Ta byla maximální až při třetím záblesku a pak při každém čtvrtém. Po velkém počtu záblesků se produkce kyslíku ustálí na nějaké stálé hodnotě. Zařízení na kumulaci náboje vypadá asi tak jak je namalované ve schématu vpravo. Stav S1 je charakteristický pro temnotně adaptovaný chloroplast. Teprve stav S4 je dost silný oxidant a je schopen oxidovat vodu (uvolnit 4 elektrony a dva protony) a vrátit se do stavu S0. Po absorpci dalšího kvanta fotonu pak přejde do stavu S1. Při experimentu může dojít k „minutí“ P680, dvojité excitaci nebo k relaxaci zpět na S0. To vše způsobí, že po mnoha excitacích je produkce kyslíku ustálená a nevykazuje čtyřdobou periodicitu. Oxidace vody vyžaduje jako kofaktor v proteinovém klastru mangan (Mn), anion chloru (Cl-) a vápenaté kationty (Ca2+). Detaily struktury klastru se zatím neznají.
. (redox potenciál přechodu P680/P680* musí být větší než +820 mV, protože to minimálně je třeba k rozštěpení vody). Oxidace vody ale není přímý proces. Vyžaduje kumulaci náboje. To bylo potvrzeno experimentálně pomocí krátkých jednofotonových záblesků. První dva záblesky nevyvolaly téměř žádnou produkci kyslíku (viz graf vlevo). Ta byla maximální až při třetím záblesku a pak při každém čtvrtém. Po velkém počtu záblesků se produkce kyslíku ustálí na nějaké stálé hodnotě. Zařízení na kumulaci náboje vypadá asi tak jak je namalované ve schématu vpravo. Stav S1 je charakteristický pro temnotně adaptovaný chloroplast. Teprve stav S4 je dost silný oxidant a je schopen oxidovat vodu (uvolnit 4 elektrony a dva protony) a vrátit se do stavu S0. Po absorpci dalšího kvanta fotonu pak přejde do stavu S1. Při experimentu může dojít k „minutí P680, dvojité excitaci nebo k relaxaci zpět na S0. To vše způsobí, že po mnoha excitacích je produkce kyslíku ustálená a nevykazuje čtyřdobou periodicitu. Oxidace vody vyžaduje jako kofaktor v proteinovém klastru mangan (Mn), anion chloru (Cl-) a vápenaté kationty (Ca2+). Detaily struktury klastru se zatím neznají.")
31
Fotofosforylace, ATP-syntáza

32
Pokus Jagendorfa a spol. (uvedeno v Taiz a Zeiger č
Pokus Jagendorfa a spol. (uvedeno v Taiz a Zeiger č. vydání), který ukazuje, že delta pH přes thylakoidní membránu je hnací silou pro syntézu ATP z ADP a P: 1/ vyrovnat pH v chloroplastech v pufru o pH=4, 2/ přenést do media s pH=8. Tím dojde k tomu, že uvnitř thylakoidů je pH=4 a vně pH=8. 3/ přidat ADP+P, měřit syntézu ATP.
, který ukazuje, že delta pH přes thylakoidní membránu je hnací silou pro syntézu ATP z ADP a P: 1/ vyrovnat pH v chloroplastech v pufru o pH=4, 2/ přenést do media s pH=8. Tím dojde k tomu, že uvnitř thylakoidů je pH=4 a vně pH=8. 3/ přidat ADP+P, měřit syntézu ATP.")
33
Komplex ATP-syntázy Vedle obou fotosystémů, světlosběrných antén a elektrontransportního řetězce je v thylakoidní membráně další významný proteinový komplex, protonová ATP-syntáza. Ta využívá gradientu protonů mezi lumenem thylakoidů a stromatem k fosforylaci ADP na ATP. Má dvě základní části, membránovou CF0 a vyčnívající do stromatu CF1. Membránová část je ze tří podjednotek přičemž podjednotka III se několikrát opakuje. Vnější část se skládá z 5 podjednotek označených písmeny řecké abecedy a zajišťuje syntézu ATP. Roli přitom hraje rotující gama podjednotka. Membránová část CF0 umožňije tok protonů čímž se dodává tzv. proton motorická síla, umožňující rotaci podjednotky gama.

34
Protonová ATP-syntáza

35
Regulace, ochrana, xantofylový cyklus

36
Tři fáze Calvinova cyklu: karboxylace, redukce a regenerace RUBP
Tři fáze Calvinova cyklu: karboxylace, redukce a regenerace RUBP. Energetická potřeba pro fixaci jedne molekuly CO2 jsou 3 ATP a 2 NADPH (na tři molekuly CO2, a tedy jedny nově vytvořenou triózu je to trojnásobek). Z cyklu odchází trióza - fosfogyceraldehyd (GAP)
. Z cyklu odchází trióza - fosfogyceraldehyd (GAP)")
37
Karotenoidy - chemismus
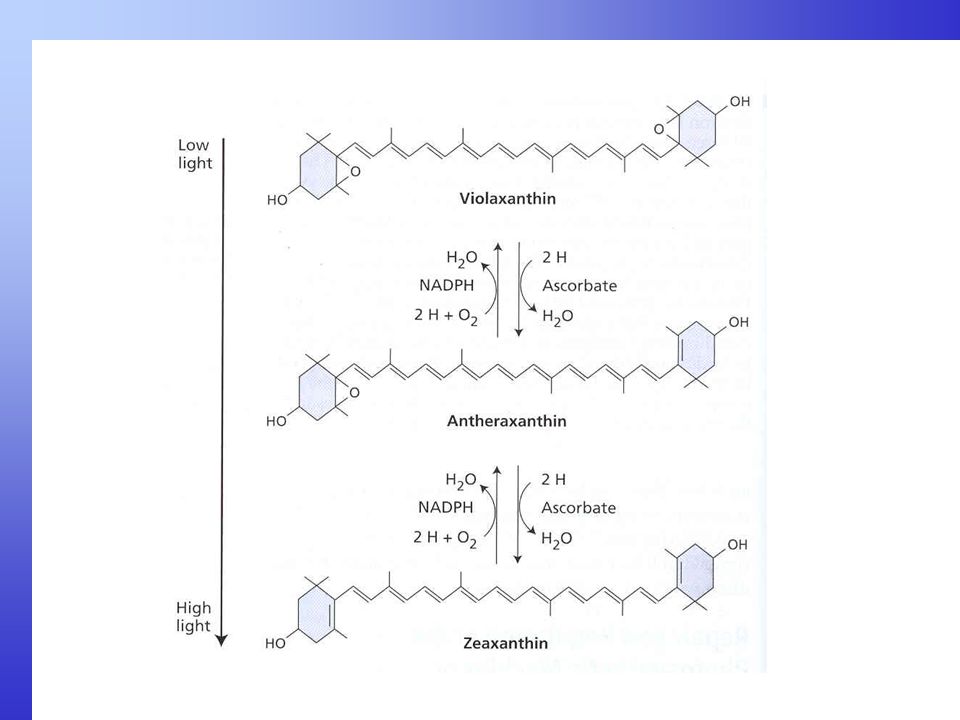
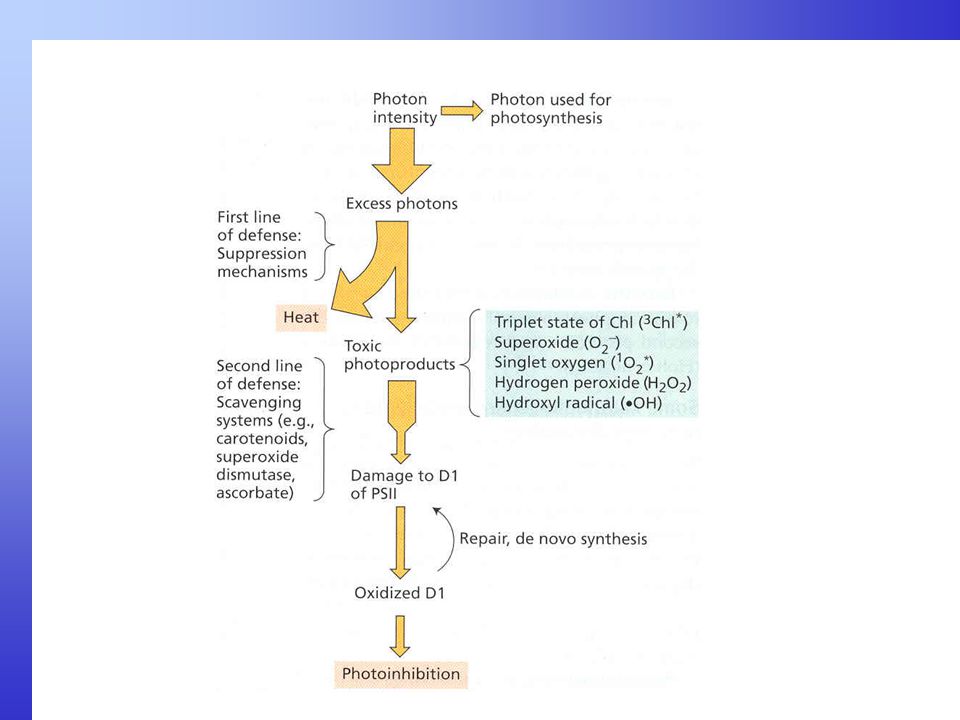
Geranylgeranyl difosfát (20 C) Fytoen (40 C, 9 dvojných vazeb) Fytoflueh (40 C, 10 dvoj. vazeb) -karotén (40 C, 11 dvoj. vazeb) Neurosporen (40 C, 12 dv. v.) Lykopen (40 C, 13 dv. v.) -karotén (40 C, 11 dv. v. 2cykly) Chlorofyly slouží jako fotosyntetické pigmenty ve většině autotrofních fotosyntetických organismu na Zemi. Ale ve všech bez výjimky jsou obsaženy karotenoidy. Mají dvě podskupiny, karotény, které obsahují systém dvojitých vazeb mezi uhlíky v rovinném řetězci a vodík. U beta a alfa karoténu jsou na koncích rovinného řetězce šetiuhlíkaté kruhy. Hydroxylací karoténů vznikají členové druhé podskupiny - xantofyly, které mají kyslík (OH skupiny) v cyklech na koncích rovinného řetězce. Obě mají 40 uhlíků v řetězci a základní stavební jednotkou je isoprén. Příkladem xantofylů je zeaxantin nebo violaxantin. Desaturací jednoduchých vazeb na dvojné v uhlíkovém řetězci se mění absorpční vlastnosti - karotény se stávají barevnými (obr.). Maxima absorpce karotenoidů je mezi nm, v oblasti, kde chlorofyly absorbují jen slabě. Proto karotenoidy slouží jako přídatné pigmenty v anténách. Karotenoidy hrají také roli v ochraně fotosyntetického aparátu před fotopoškozením - fotoinhibicí - při nadměrné ozářenosti (vysokém světle). Je to proto, že karotenoidy jsou sto přebírat excitační energii z tripletního chlorofylu a zabrání tak vzniku kyslíkových radikálů (singletního kyslíku), který jinak poškozuje fotosyntetický aparát (např. poškozuje D1 protein PS II). Při nadbytku světla se mění (deepoxiduje) violaxanthin přes antheraxanthin na zeaxantin, který je hlavně aktivní v disipaci nadměrné energie. V deepoxidační reakci se uvolňuje voda. Při poklesu světla se syntetizuje zpět violaxanthin. Toto se nazývá xantofylový cyklus. zeaxantin (40 C, 11 dv. v. 2cykly, 2 OH skupiny)
 Fytoen (40 C, 9 dvojných vazeb) Fytoflueh (40 C, 10 dvoj. vazeb) -karotén (40 C, 11 dvoj. vazeb) Neurosporen (40 C, 12 dv. v.) Lykopen (40 C, 13 dv. v.) -karotén (40 C, 11 dv. v. 2cykly) Chlorofyly slouží jako fotosyntetické pigmenty ve většině autotrofních fotosyntetických organismu na Zemi. Ale ve všech bez výjimky jsou obsaženy karotenoidy. Mají dvě podskupiny, karotény, které obsahují systém dvojitých vazeb mezi uhlíky v rovinném řetězci a vodík. U beta a alfa karoténu jsou na koncích rovinného řetězce šetiuhlíkaté kruhy. Hydroxylací karoténů vznikají členové druhé podskupiny - xantofyly, které mají kyslík (OH skupiny) v cyklech na koncích rovinného řetězce. Obě mají 40 uhlíků v řetězci a základní stavební jednotkou je isoprén. Příkladem xantofylů je zeaxantin nebo violaxantin. Desaturací jednoduchých vazeb na dvojné v uhlíkovém řetězci se mění absorpční vlastnosti - karotény se stávají barevnými (obr.). Maxima absorpce karotenoidů je mezi nm, v oblasti, kde chlorofyly absorbují jen slabě. Proto karotenoidy slouží jako přídatné pigmenty v anténách. Karotenoidy hrají také roli v ochraně fotosyntetického aparátu před fotopoškozením - fotoinhibicí - při nadměrné ozářenosti (vysokém světle). Je to proto, že karotenoidy jsou sto přebírat excitační energii z tripletního chlorofylu a zabrání tak vzniku kyslíkových radikálů (singletního kyslíku), který jinak poškozuje fotosyntetický aparát (např. poškozuje D1 protein PS II). Při nadbytku světla se mění (deepoxiduje) violaxanthin přes antheraxanthin na. zeaxantin, který je hlavně aktivní v disipaci nadměrné energie. V deepoxidační reakci se uvolňuje voda. Při poklesu světla se syntetizuje zpět violaxanthin. Toto se nazývá xantofylový cyklus. zeaxantin (40 C, 11 dv. v. 2cykly, 2 OH skupiny)")
40
Otázky, shrnutí 1/ Dokázali byste odvodit, kolik fotonů (jednofotonových záblesků) musí chlorofyly RC fotosystémů zachytit, aby se vyvinula jedna molekula kyslíku a proces kontinuálně probíhal? 2/ Na kterých dvou místech je spojen transport elektronů s transpor- tem a tvorbou protonového gradientu? 3/ Jak se rostlina brání nadměrné ozářenosti (na úrovni thylakoidů)?
")
41
Tři fáze Calvinova cyklu: karboxylace, redukce a regenerace RUBP
Tři fáze Calvinova cyklu: karboxylace, redukce a regenerace RUBP. Energetická potřeba pro fixaci jedne molekuly CO2 jsou 3 ATP a 2 NADPH (na tři molekuly CO2, a tedy jedny nově vytvořenou triózu je to trojnásobek). Z cyklu odchází trióza - fosfogyceraldehyd (GAP)
. Z cyklu odchází trióza - fosfogyceraldehyd (GAP)")
42
Biologie buňky Prokaryotní buňka: nefotosyntetická (chemosyntetická) metanotrofní bakterie Methylomonas spec. - thylakoidům podobné struktury Přednášky ZS_ U,K 98

43
Biologie buňky Prokaryontní buňka: fotosyntetická sinice (cyanobacteria) Anabaena spec. Přednášky ZS_ U,K 99

44
Biologie buňky Chloroplast - vývoj Přednášky ZS_ U,K 107

45
Biologie buňky chloroplast - vývoj Přednášky ZS_ U,K 109

Podobné prezentace

>")







>")









